

Abstract
A major challenge in cardiac tissue engineering is the delivery of hemodynamic mechanical cues that play a critical role in the early development and maturation of cardiomyocytes. Generation of functional cardiac tissue capable of replacing or augmenting cardiac function therefore requires physiologically relevant environments that can deliver complex mechanical cues for cardiomyocyte functional maturation. The goal of this work is the development and validation of a cardiac cell culture model (CCCM) microenvironment that accurately mimics pressure-volume changes seen in the left ventricle and to use this system to achieve cardiac cell maturation under conditions where mechanical loads such as pressure and stretch are gradually increased from the unloaded state to conditions seen in vivo. The CCCM platform, consisting of a cell culture chamber integrated within a flow loop was created to accomplish culture of 10 day chick embryonic ventricular cardiomyocytes subject to 4 days of stimulation (10 mm Hg, ~13% stretch at a frequency of 2 Hz). Results clearly show that CCCM conditioned cardiomyocytes accelerate cardiomyocyte structural and functional maturation in comparison to static unloaded controls as evidenced by increased proliferation, alignment of actin cytoskeleton, bundle-like sarcomeric α-actinin expression, higher pacing beat rate at lower threshold voltages and increased shortening. These results confirm the CCCM microenvironment can accelerate immature cardiac cell structural and functional maturation for potential cardiac regenerative applications.
Keywords: CCCM, cardiomyocytes, stimulation, regeneration
Introduction
Cardiovascular disease (CVD) is the leading cause of morbidity and mortality in the world. In the United States alone, CVD claims ~ 1,372,000 lives each year, resulting in an annual healthcare burden of > $300 billion.1 The medical morbidity and mortality associated with heart failure due to acute and chronic heart disease is a major component of this heath care burden and heart transplant remains the only definitive treatment for heart failure. While transplantation can significantly enhance quality of life for patients with heart failure, the limited availability of donor hearts ensures that only a small fraction of the patients suffering from cardiac injury achieve benefit. Because cardiomyocyte cell death is the hallmark of cardiac pathology that precedes heart failure, major efforts are underway to restore cardiac tissue structure, and function through regenerative approaches. 2 Unfortunately, unlike most cells in the body, cardiomyocytes are terminally differentiated and have limited proliferative potential.3 Therefore, repair of cardiac tissue occurs via formation of scar tissue that is incapable of generation of contractile forces resulting in loss of cardiac function.
Various clinical studies support the possibility that cardiac tissue can be regenerated via transplantation of progenitor or differentiated cell populations.4 However, the goal of true regeneration where injured tissue is replaced with functional myocardium has yet to be accomplished. A majority of studies rely on the direct delivery of undifferentiated cardiac stem/progenitor cells to the location of injury. Current results indicate an improvement in cardiac function with transplanted cells augmenting the host myocardium through physical reinforcement and paracrine signaling,5 however only a small percentage of delivered cells remain at the infarct location with no conclusive evidence of myocardial regeneration. In contrast to this approach, several groups have also sought to accomplish differentiation and maturation of cardiomyocytes from stem/progenitor cells in vitro, prior to transplantation the engineered ‘cardiac patch’ in vivo. 6 Current strategies typically accomplish culture of cells cardiac stem/progenitor cells within biodegradable natural or synthetic scaffolds in static culture environments 7,8 with biochemical cues to accomplish differentiation into adult cardiomyocyte phenotypes. The scaffolds establish a 3D environment enabling in vivo -like cell-cell and cell-ECM interactions.8,9 Despite promising results, generation of cardiac tissue constructs that can replace damaged tissue to restore cardiac function (stroke volume > 55 mL, ejection fraction > 55% and heart rate > 60 bpm) in humans has yet to be accomplished.
Several studies using animal models have confirmed that although normal heart development is programmed in the organism’s genetic blueprint, hemodynamic mechanical stresses play significant roles in shaping the formation and ensuring proper function of the heart.10 Therefore it is likely that in vitro mechanical stimulation can impact the rate and fate of cardiomyocyte differentiation into functional cardiac tissues.11 The importance of mechanical stimulation has been understood; however, complex systems that can accomplish culture of cardiac cells in environments that accurately mimic pressure-volume changes seen in heart are currently unavailable. Current approaches to stimulate cells with mechanical stress rely on an array of platforms that include novel fabricated platforms and systems available from commercial suppliers. These systems are able to subject cells and tissues to tension, compression and/or shear stress. Some groups have designed systems to deliver multiple stimuli such as perfusion, stretch and electrical stimulation simultaneously.12 However, these systems fail to mimic the dynamics and complexity of mechanical loads experienced by cardiomyocytes during pressure-volume changes in the heart. For example: cyclic stretch generated using the Flexcell® Tension Systems (http://www.flexcellint.com/documents/FX5000TBrochure.pdf) only replicates the strain waveforms and does not mimic changes in pressure. Whereas during each cardiac cycle, cardiomyocytes in the left ventricle of the heart experience passive stretch during the filling phase, pressure buildup during isovolumetric contraction and finally return to their relaxed state after the ejection.
In an effort to develop physiologically relevant systems for culture of cardiomyocytes and enable cardiac tissue generation, a microfluidic cardiac cell culture model was previously designed and tested to achieve concomitant stimulation with pressure and stretch.13 However, in this system, pressure-volume changes seen in the left ventricle were not accurately replicated as membrane stretch overlapped with the ejection phase. To overcome this shortcoming and develop a model that accurately mimics left changes experienced by cells during the cardiac cycle, we designed and validated a new cardiac cell culture model (CCCM) where cells are cultured on a thin polymeric membrane in a cell culture chamber integrated within a flow loop. With each cycle, cells cultured within the CCCM undergo isovolumetric contraction, ejection, isovolumetric relaxation, and filling. The CCCM therefore is essentially a left ventricle mimic that can be dynamically tuned to go from static unloaded state (as seen in very early development or during myocardial unloading during ventricular assist device (VAD) support) to pulsatile pressure and cyclic stretch conditions seen in the adult heart.13 We envision that this system will not only provide time varying mechanical cues for differentiation of stem/progenitor cells into adult cardiomyocytes but also provide mechanical conditioning under conditions ranging from embryonic heart to adult heart to generate sufficient contractile forces and integrate within native cardiac tissue to augment cardiac function. In this manuscript we detail the design and construction of the CCCM system and provide proof-of-concept demonstration of the role of mechanical stimulation in accomplishing functional maturation of embryonic chick cardiomyocytes.
MATERIALS AND METHODS
CCCM Design
A schematic detailing the basic setup of the CCCM is illustrated in Fig 1A and a picture of the actual setup is shown in Fig 1B. The CCCM consists of a cell culture chamber sandwiched between two rigid plates (Fig 1C) and assembled within a flow loop. The arrangement allows fluid to flow from the pump flows via a one-way flow control valve into the cell culture chamber and exit the cell culture chamber through a tunable pressure relief valve into a collection tube and recycled within the flow loop. Additionally, a programmable pressure generator is used to supply periodic pulses of pressurized air to the reverse side of the polymeric membrane on which cells are cultured to mimic ventricular contraction. Finally, two tunable compliance elements, a fluidic compliance integrated prior to the one-way valve and air compliance element integrated at the end of the pressure generator flow path are used to modulate pressure waveforms within the cell culture chamber. Pressure measurements are made using a pressure sensor that samples directly from the cell culture chamber and monitored real-time via a LabView program.
Figure 1.

(A) Schematic of the flow loop of the CCCM (a) peristaltic pump, (b) pulmonary compliance, (c) pulmonary resistance, (d) one-way (mitral) valve, (e) cell culture chamber, (f) pressure sensor, (g) pressure generator, (h) air compliance, (i) systemic compliance, (j) systemic resistance and (k) cell culture medium collection reservoir, (B) Picture of actual setup, (C) Sketches of the (a)top plate, (b) cell culture chamber and (c) bottom plate used to assemble the cell culture chamber within the flow loop.
CCCM Fabrication and Components
The CCCM cell culture chamber was 1 cm in diameter and the thin membrane at the bottom was ~ 0.5 mm thick. Both the chamber and the membrane were fabricated using standard soft-lithography techniques described previously.13,14 Thick (1 cm) and thin (0.5 mm) layers of (poly)dimethyl siloxane (PDMS) (Sylgard 184, Dow Corning) were cast by mixing the pre-polymer and cross-linking solution at a ratio of 10:1 and allowed to cure at 80°C for 3 hours. Both the thin and thick layers were cut to 4 cm x 4 cm squares and a 1.36 cm hole was punched through the thick layer using a cork borer. Both layers were autoclaved and irreversibly bonded together after treatment with oxygen plasma (March RIE). The cell culture chamber was assembled between two polycarbonate plates. The top plate has access ports for the inlet and outlet tubing to interface with the setup. The bottom plate contains a 2 cm diameter chamber to allow the membrane to deflect and stretch. Entry and exit ports for supply of compressed air to the chamber were also provided. Flexible, autoclave compatible polymeric tubing was used to establish connections within the flow loop. A 50 mL tube served as a reservoir to supply fluid to the peristaltic pump and to collect fluid that exits the cell culture chamber ensuring continuous supply.
CCCM Operation
The working cycle of the CCCM mimics the working cycle of the left ventricle. Pressure is supplied to the reverse side of the membrane using a ventilator (SAR-830/P, CWE Inc., Ardmore, PA), inducing isovolumetric contraction and pressure buildup within the cell culture chamber. Once the pressure exceeds the set value of the pressure relief valve, the accumulated fluid in the CCCM is ejected. Then the pressure applied to the reverse side of the membrane is lowered significantly to cause the CCCM to undergo isovolumetric relaxation. Fluid flow is introduced into the CCCM via a peristaltic pump. Fluid fills the cell culture chamber causing the membrane to stretch passively, mimicking ventricular diastole.
Cell Culture Chamber Preparation for Cell Seeding
To ensure seeded cells adhere to the thin polymeric membrane and maintain attachment during the stretching process, a glutaraldehyde cross-linked type I collagen matrix was established on the surface of the thin polymeric membrane to act as a strong anchor. First, the sterilized PDMS cell culture chamber was treated with oxygen plasma to create a hydrophilic surface via exposure to oxygen plasma (March RIE) (100mmTorr, 100 mmHg, 10% oxygen, 30 sec). Then, a 250μl of the collagen type I solution (0.67mg/ml) was allowed to gel at 37°C for 2 hours. Finally, 400 μl of 0.02% glutaraldehyde solution was added to the collagen surface and maintained for 1 hour at room temperature. The chamber was then washed with deionized (DI) water and 4N sodium hydroxide (NaOH) solution twice (30 mins/each) and then again with DI water and stored at 4°C for later use. Prior to cell seeding, the chambers were incubated with 50μg/ml of FN, 10% attachment fact or in 0.02% gelatin solution for 48 hours at 4°C and 15 mins at 37°C to enhance cell attachment.
Membrane Deflection and Strain Estimation
The increase in volume of the CCCM cell culture chamber following the filling cycle was estimated and used to determine the deflection profile of the membrane at the bottom of the cell culture chamber using finite element analysis (FEA). Using ANSYS 14.5 (Southpointe, PA) FEA software, the PDMS membrane was modeled as a Mooney-Rivlin hyper-elastic material. To minimize computational time, a 2-D model using PLANE 183 or SHELL 181 with an axis-symmetric boundary conditions was used. The membrane was assigned fixed boundary conditions at the edges and the deflection of the membrane for increasing volumes was estimated. The material properties of the PDMS used for the simulation were: Young’s Modulus of 750 kPa and a Poisson’s Ratio of 0.49. The radial strain (εR) from the center to the edge was estimated using (1) where Rf and Ri are the initial and final radius:
| (1) |
The circumferential strain (εR) from the center to the edge was estimated using the Green strain equation (2):
| (2) |
| (3) |
Embryonic Chick Cardiomyocyte Isolation and Culture
Fertile White Leghorn chicken eggs (Utah State University, Logan, UT) were incubated in a forced-draft, constant-humidity incubator until Hamburger-Hamilton (HH) stage 36 (day 10 of a 46-stage, 21-day incubation period). Excised day 10 fetal left ventricles were enzymatically digested using 2 mg/ml of collagenase type II followed by 0.05% trypsin-EDTA solution (Invitrogen, Carlsbad, CA) and cultured on a gyratory shaker (60–70 rotations/min) for 18 hours to re-aggregate viable cardiomyocyte clusters. For each cell culture chamber, approximately of 3.0–3.5 x104 cell clusters were seeded and cultured in a static condition at 37°C, 5% CO2 for 24 hours before being subject to mechanical loading within the CCCM or maintained under static conditions as control.
Mechanical Stimulation within the CCCM
Following 24 hours of culture under static conditions to achieve 100% confluence, the cell culture chamber with cells was transferred to the CCCM for mechanical stimulations for 4 days in a cell culture incubator maintained at 37°C and 5% CO2. The system was setup to enable the fluid flow to generate 15% passive stretch, peak pressure of ~ 10 mm Hg at a frequency of 2 Hz which is similar to conditions seen at stage 36 in vivo. Growth medium was replaced every two days. After removal of the cell culture chamber from the CCCM platform, the constructs were maintained under static conditions for ~ 3 hours prior to analysis.
Electrical Pacing
Electrical pacing of mechanically stimulated constructs and unstimulated controls was accomplished using a platinum wire electrode (d=0.5mm, Alfa Aesar, Ward Hill, MA, 43288) embedded vertically into the tissue-collagen gel matrix and an electrical pacing system (Isolated Programmable Stimulator, Coulbourn Instruments, 02142-1). The maximum pacing tissue beat rate was recorded at increasing while varying the threshold voltage.
Evaluation of Cell Proliferation and Maturation
Cell alignment and maturation were evaluated using standard confocal microscopy methods. The evaluate cell proliferation, Bromodeoxyuridine (BrdU, 20μM, Sigma) was added to the tissues 48 hours prior to fixation. After fixation, the tissue was permeablized using 0.1% Triton solution for 30 min at room temperature and washed with 1X phosphate buffered saline (PBS) 3 times. 1N HCl was then added to the tissue for 10 min on ice followed by 2N HCl for another 10 min at room temperature before moving to 37°C incubator for an additional 20 min. After HCl incubation, the tissue was washed with 1X TBE buffer (10X TBE, Mediatech Inc, Manassas, VA, 46-011-CM) followed by a 20 minute incubation in 1X TBE buffer at room temperature. A solution of 1% BSA+0.1% glycine was used as the blocking buffer and incubated for 1 hour at room temperature. The primary antibodies, mouse cardiac α-actinin EA53 IgG1(1:400, Abcam, Ab7817) and anti-BrdU (1:100, Abcam, ab1893) were used for staining with in a staining buffer containing 1X PBS and in 1% bovine serum albumin (BSA) for 24 hours at room temperature. The secondary antibodies, anti-mouse (1:600, 488 green, Invitrogen, A21121) and anti-sheep (1:400, red, Abcam, Ab6743) were then added in 1X PBS with 1% BSA for 1 hour at room temperature. DAPI (1:100, Millipore, 90229) nuclear stain was also added to stain for the nucleus. Finally, the tissue constructs were mounted with anti-fade solution (Invitrogen, San Jose, CA) and imaged using a Nikon A1 confocal microscopy system (Nikon Instruments, Melville, NY).
Cardiomyocyte Contractility
Cardiomyocyte contractility (shortening) was estimated using video analysis and Lagrangian strain calculations for finitely small displacement as previously described by Bonet et al.15 Briefly, if p1 and p2 are two nearby points defined on the first frame and d1 denotes the Euclidian distance between p1 and p2 at the first frame (i.e., reference frame), then dj denotes the Euclidian distance between these two points at time frame j. Then, the engineering strain S, at frame j, is defined as:
| (4) |
A custom algorithm was written to estimate planar Lagrangian strains. First, the peak displacement was determined by tracking two points based on their appearance features through the image frames and calculating the displacement between the two points over the image frames. The maximum displacement was then selected as the reference frame. Then, five pairs of points on the first (reference) frame were determined and their tracked pairs on the selected frame with maximum displacement. Lagrangian strain for each of the points was estimated using (5) using each pair of points. Finally, the mean Lagrangian strain was calculated as expressed as % shortening.
RESULTS
Generation of Pressure and Stretch Waveforms
A synergistic relationship exists between the pump flow rate, membrane thickness and the magnitude of pressure pulses within the CCCM and all three parameters need to be set precisely to ensure proper filling and ejection of fluid from the cell culture chamber, and mechanical stresses. Day 10 embryonic chick cardiomyocytes experience in vivo peak pressures of ~ 10–15 mm Hg and radial and circumferential strains of ~ 10–20%.16 To culture of cardiomyocytes under these conditions, a 500 μm thick membrane was selected and a flow rate of ~ 2.6 mL/min was used to attain a filling volume of 44 μl per cycle. The programmable pressure generator was tuned using the compliance element to compress the fluid within the cell culture chamber and generate a peak pressure of ~ 10 mm of Hg. The pressure waveform and the pressure-volume loop for a single working cycle of the CCCM are shown in Fig 2. For a 500 μm thick PDMS membrane, a filling volume of 44 μl was estimated to produce peak radial strain of 12% and a peak circumferential strain of 13% (Fig 3). The pressure and strain values are consistent with levels experienced by chick embryonic cardiomyocytes at day 10. To ensure that fluid shear stress was not a major issue, we followed design guidelines published in our prior work to ensure that the depth of the cell culture wells was > 0.5 cm resulting in low shear stress recirculation at the bottom of the wells.13
Figure 2.
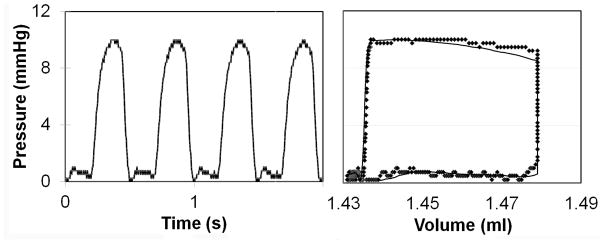
Pressure vs. Time plot representing the changes in pressure experienced by cardiomyocytes within the CCCM and Pressure-Volume loop correlating changes in pressure with changes in volume within the CCCM chamber.
Figure 3.
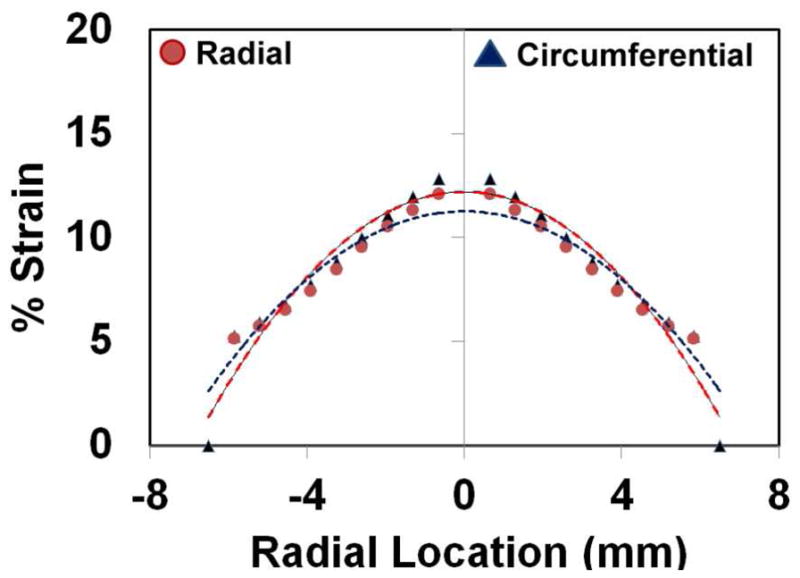
Estimation of radial strain (▲) and circumferential strain (●) on the thin polymeric membrane at the bottom of the cell culture chamber as a function of radial position.
Evaluation of Cell Alignment, Maturation and Proliferation
Alignment of cardiomyocytes, expression of maturation markers, and rate of proliferation were all evaluated using immunofluorescence microscopy. Cardiomyocytes subject to mechanical stimulation within the CCCM align in response to the radial strain and are also influenced by the fluid flow as evidenced by strong directional alignment of actin cytoskeletal filaments in comparison to unstimulated controls (Fig 4A). Cardiomyocyte maturation was evaluated via staining for sarcomeric α-actinin which is only expressed in mature, contracting cardiomyocytes. In comparison to static controls, cardiomyocytes stimulated within the CCCM show prominent and bundle-like expression of sarcomeric α-actinin confirming a more mature and in vivo –like phenotype (Fig 4B). Finally, cell proliferation was evaluated in both static controls and stimulated samples via determination of BrdU incorporation into the DNA of proliferating cells. CCCM stimulated samples proliferated at ~20% higher rate than unstimulated samples (Fig 4C) indicating that mechanical stimulation delivers pro-growth stimuli. Results indicate a statistically significant (p < 0.001) increase in the proliferation rate of CCCM stimulated samples in comparison to controls.
Figure 4.

(A–B) Confocal microscopy images of controls (left) and CCCM stimulated samples (right) stained with (A) phalloidin to visualize polymerized f-actin and (B) antibody specific to cardiac α-actinin. (C) Evaluation of cell proliferation in controls and CCCM stimulated samples via estimation of BrdU incorporation into the nucleus of dividing cells. For cell proliferation studies the sample size was N = 6 with two tailed significance set at p ≤ 0.001.
Cardiomyocyte Contractility
To estimate the contractility of cardiomyocytes, we determined the shortening length in both control and CCCM stimulated samples with and without electrical pacing. Without electrical pacing, shortening of CCCM conditioned cardiomyocytes was ~ 4.49 ± 0.88% whereas controls were at 2.73 ± 1.8% (Fig 5). When both samples were electrically paced to attain their peak beat rates, the CCCM conditioned cardiomyocytes shortening increased to 5.52 ± 0.88% whereas the shortening for controls only increased to 2.96 ± 1.79% (Fig 5). Evaluation using unpaired t-test shows no statistical significance between the shortening length of control and CCCM stimulated samples without pacing. However, when electrically paced to attain the maximum beat rates, s statistically significant (p < 0.05) difference in the shortening lengths between controls and CCCM stimulated samples was observed. Significant variability in shortening length was observed with the control samples as evidenced by the larger standard deviation. These results suggest that electrical pacing increases contractility of CCCM conditioned cardiomyocytes by > 1% whereas the effect on controls was minimal at ~ 0.3%.
Figure 5.

Cardiomyocyte Contractility. Controls (black) and CCCM stimulated (clear) samples evaluated with and without electrical pacing. Measurements were made at 5 locations on each sample for 6 samples. Two-tailed significance was set at p ≤ 0.05.
Cardiomyocyte Beat Rate
The beat rate of the CCCM conditioned cardiomyocytes and static controls were evaluated with and without electrical pacing. The intrinsic beat rate of the controls was 42.7 ± 11.2 bpm whereas CCCM conditioned cardiomyocytes was 62.2 ± 15.2 bpm (Fig 6A). With electrical pacing, the stimulated group attained a maximum beat rate of 245 ± 20.8 bpm in compared to the 194 ± 10.8 bpm of the control group (Fig 6A). The in-vivo heart beat rate for embryonic chick is ~ 260bpm. The threshold voltage necessary to attain the maximum beat rate in the CCCM samples was only 3.05 ± 0.17 V (245 bpm) whereas the control group required 5.03 ± 0.21 V to attain a beat rate of 194bpm (Fig 6B). Evaluation using unpaired t-tests clearly show statistical significance (p < 0.001) for intrinsic beat rate, pacing beat rate and threshold voltage necessary to attain maximum beat rate between unstimulated controls and CCCM stimulated cardiomyocytes. These results suggest that mechanical conditioning enables cellular alignment and electrical coupling between neighboring cardiomyocytes as evidenced by the higher intrinsic and pacing beat rates in cardiomyocytes and the lower threshold voltage necessary to attain the maximum pacing beat rate.
Figure 6.

(Top) Evaluation of intrinsic beat rate (left) and maximum pacing beat rate in controls (black) and CCCM stimulated (clear) samples. (Bottom) Maximum threshold voltage required to attain maximum pacing beat rates in controls and CCCM stimulated samples. For all experiments, the sample size was N = 6 with two-tailed significance set at p ≤ 0.001.
DISCUSSION
In early development, key morphogenetic cascades transform the heart from a tube into a complex 4-chambered structure capable of pumping blood throughout the vertebrate body. During this early developmental stage, blood flow is essential as the heart is extremely sensitive to biomechanical cues and even small variations in normal hemodynamic loads lead to cardiac malformations. Despite significant promise, current strategies for in vitro stem cell based cardiac tissue generation have yet to achieve generation of functional cardiac tissue that can integrate with injured myocardium and restore/augment cardiac function. This can partly be attributed to the fact that in most cases, cardiac tissue generation is accomplished in environments that fail to mimic the complex and time varying hemodynamic loading and unloading seen in vivo. We therefore sought to create a dynamic and tunable CCCM for cardiac tissue generation that can provide suitable biomechanical developmental and maturation stimulus. The CCCM is a left ventricle mimic that replicates the different phases of the cardiac cycle like isovolumetric contraction, ejection, isovolumetric relaxation, and filling. The magnitude and duration of delivery of individual mechanical stimulus experienced during each phase can be varied using external tunable components. This system was designed to provide an environment which can be used to gradually deliver critical biomechanical cues necessary for cardiac tissue differentiation and functional maturation before being surgically implanted in the body to augment and restore cardiac function in injured hearts.
To demonstrate feasibility of the CCCM to direct functional maturation of cardiac tissue, we cultured embryonic chick cardiomyocytes isolated at day 10 which corresponds to Hamburger-Hamilton (HH) stage 36 which represents the period just after cardiac morphogenesis at which the embryonic ventricle has differentiated from a single primitive ventricular chamber into right and left ventricular (LV) chambers. The CCCM delivers hemodynamic stresses similar to that seen in the native ventricle and therefore, the levels of pressure and stretch during the pressure-volume cycle were set to ~ 10 mm Hg peak pressure and ~ 10–20% stretch which are typical of values seen during HH stage 36. Following 4 days of mechanical stimulation, the cells cultured within the CCCM were evaluated for cardiomyocyte maturation, contractility and proliferation in comparison to static unloaded controls.
Our results establish proof-of concept that application of appropriate mechanical cues similar to that seen in vivo during cardiogenesis direct alignment, maturation and proliferation of immature cardiac cells. It can be clearly seen from our results that mechanical stimulation significantly impacts several aspects of cardiac tissue development and maturation. Stimulation within the CCCM impacts proliferation as evidenced by increased incorporation of BrdU into the DNA of dividing cells indicating that mechanical stresses at levels provided within the CCCM promote expansion of cardiomyocytes. Cardiomyocytes within the CCCM respond to cyclic stretch and align perpendicular to the direction of radial stretch which can be seen from staining for cytoplasmic f-actin. The effect of fluid flow direction was not clear as stretch appears to be the primary stimulus that directs cell alignment. Cardiomyocytes cultured within the CCCM showed strong and bundle-like sarcomeric α-actinin expression in comparison to static controls. Both CCCM stimulated cardiomyocytes and static controls were electrically paced with increasing frequency to determine the highest beat frequency in the cardiomyocytes prior to 2:1 heart block. Results show that the CCCM controls not only attained a higher beat rate, close to adult chick beat rate of 300 bpm but this beating rate was accomplished at a lower threshold voltage. These results suggest that mechanical stimulation encourages superior cellular electrophysiological behavior via cell alignment and strong cell-cell electrical coupling. Finally, cardiac cell contractility was evaluated to determine shortening length of individual cardiomyocytes. Results show that shortening length indicative of superior cardiac function is significantly higher for stimulated cells and increases by over 1% in the presence of electrical stimulation whereas the change is minimal in controls.
Overall, our results clearly suggest that subjecting immature cardiomyocytes to left ventricle –like pressure-volume changes can significantly influence cardiac tissue maturation and function. This approach therefore has potential to deliver biomechanical cues for in vitro cardiac cell differentiation and maturation prior to implanting functionally mature cardiac tissue patches to support injured myocardium. Despite lack of electrical stimulation, mechanical stimulation alone via replication of the pressure-volume cycle directs functional maturation of embryonic cardiomyocytes. Integration of electrical pacing has potential to further enhance differentiation and functional maturation and will fully replicate the in vivo cardiac tissue microenvironment. The CCCM also has potential for cardiac stem cell differentiation where cardiac stem/progenitor cells are seeded within the CCCM and the mechanical loads are gradually increased to mimic loading seen during embryonic development and gradually increased to loading seen in the adult heart.
CONCLUSION
In summary, we designed and validated the CCCM to accurately mimic hemodynamic stresses seen during pressure-volume changes seen in the left ventricle. We envision this platform to be widely applicable for the generation of cardiac tissue via delivery of critical biomechanical cues to enable differentiation and functional maturation of cardiac stem/progenitor cells. To demonstrate proof-of-concept we have shown that embryonic chick cardiomyocytes cultured within this system attain superior structural and functional maturation in comparison to static unloaded controls.
Acknowledgments
MDN was supported via a Ph.D. fellowship from the Southern Regional Education Board (SREB). This work was supported via NSF grant # 0814194 (PS), NIH grant # R01 HL085777-01-A2 (BK) and Kosair Charities Pediatric Heart Research Program (BK).
Non-Standard Abbreviations
- CCCM
Microfluidic Cardiac Cell Culture Model
- PDMS
(poly) dimethyl siloxane
Standard Abbreviations
- BrdU
5-bromo-2′-deoxyuridine
- CVD
Cardiovascular Disease
- PV
Pressure-Volume
- FEA
Finite Element Analysis
- bpm
beats per minute
- Ri
Initial Radius
- Rf
Final Radius
- εR
Radial Strain
- εC
Circumferential Strain
References
- 1.Lanza RP, Langer RS, Vacanti J. Principles of tissue engineering. 3. Elsevier Academic Press; Amsterdam; Boston: 2007. p. xxvii.p. 1307. [Google Scholar]
- 2.Chiong M, Wang ZV, Pedrozo Z, Cao DJ, Troncoso R, Ibacache M, Criollo A, Nemchenko A, Hill JA, Lavandero S. Cell Death Dis. 2011;2:e244. doi: 10.1038/cddis.2011.130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bergmann O, Bhardwaj RD, Bernard S, Zdunek S, Barnabé-Heider F, Walsh S, Zupicich J, Alkass K, Buchholz BA, Druid H, Jovinge S, Frisén J. Science. 2009;324:98–102. doi: 10.1126/science.1164680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Murrow JR, Dhawan SS, Quyyumi AA. 2010:131–151. [Google Scholar]; Reinlib L, Field L. Circulation. 2000;101:e182–187. doi: 10.1161/01.cir.101.18.e182. [DOI] [PubMed] [Google Scholar]; Wollert KC, Drexler H. Circ Res. 2005;96:151–163. doi: 10.1161/01.RES.0000155333.69009.63. [DOI] [PubMed] [Google Scholar]
- 5.Orlic D, Kajstura J, Chimenti S, Limana F, Jakoniuk I, Quaini F, Nadal-Ginard B, Bodine DM, Leri A, Anversa P. Proceedings of the National Academy of Sciences of the United States of America. 2001;98:10344–10349. doi: 10.1073/pnas.181177898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zimmermann WH, Schneiderbanger K, Schubert P, Didié M, Münzel F, Heubach JF, Kostin S, Neuhuber WL, Eschenhagen T. Circulation Research. 2002;90:223–230. doi: 10.1161/hh0202.103644. [DOI] [PubMed] [Google Scholar]; Shimizu T, Yamato M, Isoi Y, Akutsu T, Setomaru T, Abe K, Kikuchi A, Umezu M, Okano T. Circulation Research. 2002;90:e40–e48. doi: 10.1161/hh0302.105722. [DOI] [PubMed] [Google Scholar]; Zimmermann WH, Eschenhagen T. Heart Fail Rev. 2003;8:259–269. doi: 10.1023/a:1024725818835. [DOI] [PubMed] [Google Scholar]; Tobita K, Liu LJ, Janczewski AM, Tinney JP, Nonemaker JM, Augustine S, Stolz DB, Shroff SG, Keller BB. Am J Physiol Heart Circ Physiol. 2006;291:14. doi: 10.1152/ajpheart.00205.2006. [DOI] [PubMed] [Google Scholar]
- 7.Li RK, Yau TM, Weisel RD, Mickle DA, Sakai T, Choi A, Jia ZQ. J Thorac Cardiovasc Surg. 2000;119:368–75. doi: 10.1016/S0022-5223(00)70193-0. [DOI] [PubMed] [Google Scholar]; Masters KS, Shah DN, Leinwand LA, Anseth KS. Biomaterials. 2005;26:2517–25. doi: 10.1016/j.biomaterials.2004.07.018. [DOI] [PubMed] [Google Scholar]; Geckil H, Xu F, Zhang XH, Moon S, Demirci U. Nanomedicine. 2010;5:469–484. doi: 10.2217/nnm.10.12. [DOI] [PMC free article] [PubMed] [Google Scholar]; Lee KY, Money DJ. Chem Rev. 2001;101:1869. doi: 10.1021/cr000108x. [DOI] [PubMed] [Google Scholar]
- 8.Khademhosseini A, Langer R, Borenstein J, Vacanti JP. Proceedings of the National Academy of Sciences of the United States of America. 2006;103:2480–2487. doi: 10.1073/pnas.0507681102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Griffith LG, Swartz MA. Nat Rev Mol Cell Biol. 2006;7:211–24. doi: 10.1038/nrm1858. [DOI] [PubMed] [Google Scholar]
- 10.Al Naieb S, Happel CM, Yelbuz TM. Annals of Anatomy - Anatomischer Anzeiger. doi: 10.1016/j.aanat.2012.10.011. [DOI] [PubMed] [Google Scholar]; Keller BB. New insights into the developmental biomechanics of the atrioventricular valves. Circ Res. 2007 May 25;100(10):1399–401. doi: 10.1161/01.RES.0000269327.83426.d9. [DOI] [PubMed] [Google Scholar]; Keller BB, Liu LJ, Tinney JP, Tobita K. Ann N Y Acad Sci. 2007:15. doi: 10.1196/annals.1389.012. [DOI] [PubMed] [Google Scholar]; Lucitti JL, Visconti R, Novak J, Keller BB. Am J Physiol Heart Circ Physiol. 2006;291:28. doi: 10.1152/ajpheart.01061.2005. [DOI] [PubMed] [Google Scholar]
- 11.Goenezen S, Rennie MY, Rugonyi S. Biomechanics and modeling in mechanobiology. 2012;11:1187–204. doi: 10.1007/s10237-012-0414-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lu L, Mende M, Yang X, Korber HF, Schnittler HJ, Weinert S, Heubach J, Werner C, Ravens U. Tissue Eng Part A. 2013;19:403–14. doi: 10.1089/ten.TEA.2012.0135. [DOI] [PubMed] [Google Scholar]
- 13.Giridharan GA, Nguyen MD, Estrada R, Parichehreh V, Hamid T, Ismahil MA, Prabhu SD, Sethu P. Anal Chem. 2010;82:7581–7. doi: 10.1021/ac1012893. [DOI] [PubMed] [Google Scholar]
- 14.Estrada R, Giridharan G, Prabhu SD, Sethu P. Conf Proc IEEE Eng Med Biol Soc. 2011:6089925. doi: 10.1109/IEMBS.2011.6089925. [DOI] [PubMed]; Estrada R, Giridharan GA, Nguyen MD, Roussel TJ, Shakeri M, Parichehreh V, Prabhu SD, Sethu P. Anal Chem. 2011;83:3170–7. doi: 10.1021/ac2002998. [DOI] [PubMed] [Google Scholar]
- 15.Bonet JRD, Wood RD. Nonlinear continuum mechanics for finite element analysis. Cambridge University Press; 1997. [Google Scholar]
- 16.Tobita K, Keller BB. Am J Physiol Heart Circ Physiol. 2000;279:H959–69. doi: 10.1152/ajpheart.2000.279.3.H959. [DOI] [PubMed] [Google Scholar]