

Abstract
Patients suffering from heart failure as a result of myocardial infarction are in need of heart transplantation. Unfortunately the number of donor hearts is very low and therefore new therapies are subject of investigation. Cell transplantation therapy upon myocardial infarction is a very promising strategy to replace the dead myocardium with viable cardiomyocytes, smooth muscle cells and endothelial cells, thereby reducing scarring and improving cardiac performance. Despite promising results, resulting in reduced infarct size and improved cardiac function on short term, only a few cells survive the ischemic milieu and are retained in the heart, thereby minimizing long-term effects. Although new capillaries and cardiomyocytes are formed around the infarcted area, only a small percentage of the transplanted cells can be detected months after myocardial infarction. This suggests the stimulation of an endogenous regenerative capacity of the heart upon cell transplantation, resulting from release of growth factor, cytokine and other paracrine molecules by the progenitor cells – the so-called paracrine hypothesis. Here, we focus on a relative new component of paracrine signalling, i.e. exosomes. We are interested in the release and function of exosomes derived from cardiac progenitor cells and studied their effects on the migratory capacity of endothelial cells.
Keywords: ischemic heart disease, cell therapy, paracrine effects, exosomes
We demonstrate that cardiomyocyte progenitor cells (CMPCs) do release exosomes into their environment that can stimulate migration of endothelial cells in an in vitro scratch wound assay. Furthermore, we demonstrate that matrix metalloproteinases (MMPs) and extracellular matrix metalloproteinase inducer (EMMPRIN) are present in exosomes and that an EMMPRIN-mediated activation of CMPC exosomes is involved in the endothelial cell migration.
Cell therapy
Upon myocardial infarction, cardiomyocytes are lost and scar tissue is formed. To compensate this loss, cardiac hypertrophy and morphological remodelling responses try to restore the cardiac output, but can ultimately lead to heart failure. Heart transplantation is currently the only treatment option, but the number of donor hearts does not meet the needs of all heart failure patients. Furthermore, rejection and the complications of immunosuppressive therapy limit the success of heart transplantation. In the last decade, cell transplantation therapy has emerged as a potential therapy to treat patients suffering from heart failure. This approach is a promising strategy to regenerate cardiac tissue after myocardial infarction, thereby replacing dead and/or fibrotic tissue, to reduce or prevent adverse remodelling of the heart and improve cardiac function.
Many different progenitor cells have been used in small animals, and in pre-clinical and clinical settings, thereby suggesting a beneficial effect of the presence of engrafted cells in the injured heart [1–3]. To regenerate the myocardium, the ideal cell for transplantation should be able to differentiate into different cardiac lineages. This includes cardiomyocytes to restore contractile properties, and smooth muscle cells and endothelial cells to restore perfusion and stabilize the heart. Only a few cell types are particularly suitable as a cell source due to their differentiation potential into the cardiac lineages, as was demonstrated both in vitro as well as in vivo. From the perspective of differentiation, embryonic stem cells (ESCs) seem to be an ideal candidate for cell transplantation because of their pluri-potency, i.e. to form cells of all three lineages [4]. However, besides ethical issues, ESCs need careful selection of fully differentiated cells because of the risk of teratoma formation by undifferentiated cells [5]. Recently, other pluripotent cells have been described, termed induced pluripotent stem cells that are capable of full differentiation into all lineages [6]. Although promising and having similar potential as ESCs, these reprogrammed fibroblasts need the introduction of viral- or transient expressed pluri-potency genes and long-term effects of these introductions are not clear yet. Other stem or progenitor cells, isolated from the blood [7], bone marrow [8] or other tissues are also used for cardiac cell transplantation, but their full differentiation potential towards all cardiac lineages is disputed [3].
Since several years, different progenitor cells are isolated from the myocardium itself. They are thought to be a more suitable cell type because of their origin of isolation, potential predisposition for cardiac purposes and their greater potential to differentiate towards cardiomyocytes compared to other adult stem cells [3]. Recently, we have isolated adult and foetal human CMPCs from the heart. These cells can easily be expanded in culture and are very promising as a source of cell therapy [9–11]. These cells are able to differentiate into vascular structures and beating cardiomyocytes in vitro, without co-culturing with neonatal cardiomyocytes. In addition the CMPCs can also form vasculature and cardiomyocytes upon injection into injured myocardium in immunodeficient mice [12].
The paracrine hypothesis
Upon injection of cells into the diseased myocardium, cardiac regeneration can potentially be achieved via multiple mechanisms. Although actual differentiation of progenitor cells and active contribution to cardiac function is the main goal, the engraftment of progenitor cells and the number of newly generated cardiomyocytes and vascular cells are in many cases too low to explain the improved cardiac function and morphology [12–14]. In addition to differentiation and active participation, it is suggested that transplanted progenitor cells fuse with endogenous present cells and that release of soluble factors contributes to cardiac repair by inducing cytoprotection, neovascularization and regeneration via endogenous resident cardiac stem cells (CSCs) [13, 15, 16].
Although cardiac progenitor cells are a very promising cell source for therapy because of their differentiation potential, their release of paracrine factors might also have significant effects. In a study by Chimenti et al.[17], paracrine factors of cardiac-derived progenitor cells, so-called cardiosphere-derived cells (CDCs), are investigated. Upon injection of CDCs in the heart, improved cardiac output and reduced scarring was observed. CDCs differentiated into capillaries and cardiomyocytes, thereby explaining 20–50% of the observed increases in angiogenesis and myogenesis. However, CDCs also attracted endogenous cells, thereby inducing blood supply and reducing apoptosis, this is suggestive for paracrine effects to be involved. The collected secretions of CDCs in vitro, labelled as conditioned medium (CM), was studied in detail. The CM was shown to be able to protect neonatal rat ventricular myocytes from apoptosis under hypoxic conditions, and improve angiogenesis of human umbilical vein endothelial cells in a matrigel assay in vitro.
In a model of chronic myocardial infarction in rats, Tang et al.[14] confirmed a role for paracrine factors by intracoronary infusion of rat c-kit+ CSCs in a 4-week-old infarct. In some animals no engrafted cells could be found, but still scar area was reduced and cardiac output improved as compared to the saline infused control animals. Moreover, they observed that more cardiomyocytes, vessels and endogenous cardiac progenitor cells were present in the CSC-treated animals, but without the presence of engrafted transplanted cells. This suggests that secreted factors, released by the transplanted CSC, trigger proliferation and/or migration of endothelial cells and endogenous CSC and thereby stimulate cardiomyocyte survival or CSC myogenic differentiation.
Upon transplantation of human CMPCs into the myocardium of infarcted mouse hearts, we could observe a reduced adverse remodelling of the heart in time. This effect was sustained for at least 3 months and human cells, expressing markers for sarcomeric proteins, as well as smooth muscle and endothelial cells were still present. Although very promising, only few injected cells (∼3–4%) were engrafted and could be observed at 3 months. The positive outcome are partially because of direct differentiation of progenitor cells towards endothelial, smooth muscle cells and cardiomyocytes, but the data suggest that paracrine effects are also involved. Interestingly, increased vascular density was observed upon transplantation of vascular endothelial growth factor (VEGF) producing CMPCs [12].
Thus far, cell-based therapies to regenerate the injured heart are successful in reducing the infarct size and improving cardiac output. However, true regeneration by engrafted cells is limited and suggests paracrine factors to be involved [13, 16]. This effect reflects the communication between different cells via molecules that are released, thereby affecting processes in neighbouring cells. Paracrine factors from progenitor cells are of major interest and have extensively been studied with respect to the release of growth factor, cytokines and chemokines. Many factors are described to be produced by different progenitor cell populations, including insulin like growth factor-1, stem cell derived factor, monocyte chemoattractant protein-1, hepatocyte growth factor and VEGF. Several effects have been described on the survival, proliferation and differentiation of cardiac cells, including cardiomyocytes, smooth muscle cells, endothelial cells and endogenous progenitor cells.
Ultimately, CM of progenitor cells has been applied in animal models in vivo and ex vivo. In a Langendorf perfused rat heart, mouse ESC-CM enhanced recovery of myocardial function upon global ischemia [18]. Upon injection of concentrated CM from Akt-modified mesenchymal stem cells (MSCs) in infarcted rat hearts, apoptotic cell numbers were reduced and consequently reduced infarct sizes were assessed [19]. Furthermore Timmers et al.[20] demonstrated that injection of CM from ESC-derived MSCs leads to reduced infarct size and improved cardiac function in a pig model of ischemia reperfusion. In later experiments, exosomes within the CM were identified to contain the active component [21].
Exosomes
Exosomes are small membrane vesicles with a lipid bilayer, secreted by many, if not all, cells. These exocytosed internal vesicles of endosomal origin range in size between 50 and 100 nm and can be observed in a cup-shaped morphology by electron microscopy analysis after differential centrifugation [22]. They form by inward budding of the cell membrane to form endosomes, followed by invagination of the limiting membrane of late endosomes to from multivesicular bodies. Fusion of the multivesicular bodies with the plasma membrane results in the release of the internal vesicles, then called exosomes. These vesicles contain cytosolic derivates and express the extracellular domain of membrane-bound receptors at the surface of the membrane [23]. Exosomes contain many different proteins, including growth factors and cytokines, and coding and non-coding RNA molecules [23, 24]. Exosomes are involved in intercellular communication between different cell types, but the mechanisms of their effects are not clear. It is postulated that exosomes can influence target cell behaviour by receptor ligand binding, fusion and endocytosis.
Exosomes are described to be involved in numerous processes, including immune modulating processes and angiogenesis and migration of endothelial cells to facilitate tumour growth and the formation of metastasis. In cardiovascular research, exosomes have been identified as the active component within the CM which was injected in a pig model of ischemia reperfusion injury [21], demonstrating reduced damage and improved outcome. Ultimately these exosomes, derived from MSCs, have been injected in a mouse model of ischemia reperfusion leading to a reduced infarct size [25]. To investigate whether CMPC-derived exosomes could play a role in CMPC transplantation therapy, we studied the effect of these exosomes on endothelial cells migration in vitro and explore the mechanism involved.
Collection CMPC secreted exosomes
CMPCs are isolated as previously described [9–11]. In short, human foetal heart tissue was collected after elective abortion and individual permission using standard informed consent procedures and prior approval of the ethics committee of the University Medical Center Utrecht were obtained. Small pieces of human foetal cardiac tissue were digested with collagenase and put through a cell strainer to obtain a single cell suspension. The cells were then incubated with mouse anti-Sca-1 coated magnetic beads to isolate CMPCs. These CMPCs were cultured and grown to confluency in exosome-depleted medium for 3 to 4 days. From this CMPC-CM, exosomes are isolated by differential centrifugation and visualized using whole-mount transmission electron microscopy (Fig. 1A), demonstrating the characteristic cup-shaped appearance. Exosome vesicles are described to float in a sucrose gradient at a density between 1.10 and 1.20 g/ml [22]. Flotillin-1 was used as a marker protein to identify exosome-containing fractions and we observed that CMPC exosomes float round 1.10–1.12 g/ml (Fig. 1B).
Fig 1.
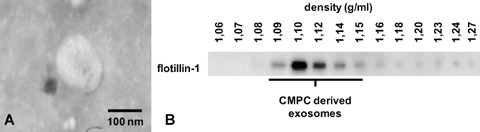
Exosome isolation of CMPCs. (A) Exosomes were isolated from 3–4 days CM from CMPCs by differential centrifugation; by an initial step at 10,000 ×g, floating cells and cell debris were removed from the CM and followed by a centrifugation step at 100,000 ×g. The CMPC-derived exosomes are analysed by whole mount electron microscopy. The size of CMPC-derived exosomes is approximately 100 nm. (B) Exosomes are purified using a sucrose gradient, for which the exosomes are resuspended in 2.5 M sucrose. On top of the isolated exosomes, different molarities of sucrose (from 2.0 M to 0.4 M) are layered. After overnight centrifugation at 200,000 ×g, 14 fractions with sucrose densities ranging from 1.06 to 1.27 g/ml were analysed for exosomal marker flotillin-1 using Western blot. The CMPC-derived exosomes float in a sucrose gradient at 1.10–1.12 g/ml.
Conditioned medium and exosomes in in vitro scratch wound assay
Because exosomes are described to be involved in angiogenesis and endothelial cell migration and based on the observation of increased capillary formation upon CMPC transplantation, we suggested that CMPC-derived exosomes enhance migration of endothelial cells. The in vitro scratch wound assay was performed with human microvascular endothelial cells (HMECs). Upon stimulation of HMECs with CMPC-CM, wound closure of HMECs is increased and, by diminishing the number of exosomes in this medium, this closure is reduced (Fig. 2A). Adding exosomes from 1 × 107 CMPCs in serum-free medium (basal medium), migration of HMECs is significantly enhanced (Fig. 2B).
Fig 2.
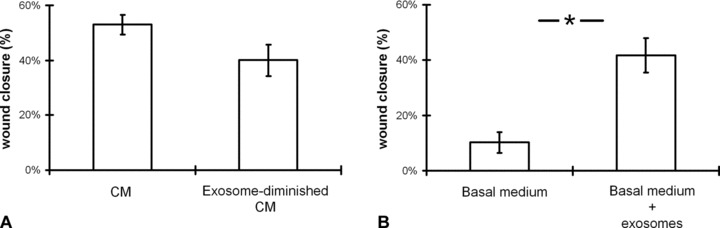
CMPC exosomes enhance migration of HMECs in a scratch wound assay. (A) In a confluent monolayer of HMECs, a scratch was made and cells were incubated for 6 hrs with CM or exosome-depleted CM. Three- to 4-day-old CMPC-CM, depleted from cells and cellular debris, is compared to exosome-depleted CM. Depletion of exosomes from the CM reduces the wound closure (n= 7; error bars = S.E.M.; not significant). (B) Isolated exosomes from 1 × 107 CMPC, resuspended in the basal culture medium of HMEC (MCDB131) without serum additives, is incubated with the HMECs. In the presence of the exosomes the closure of the wound is highly enhanced (n= 7; error bars = S.E.M. *=P < 0.05).
Exosomal signalling via MMP and EMMPRIN
To explore the mechanism how exosomes might stimulate migration of endothelial cells, we studied the role of MMPs. For cell migration, the breakdown and remodelling of matrix is required and this is mediated by MMPs [26]. Interestingly, MMPs can be found in secreted vesicles, including exosomes [27]. To identify the presence of MMP activity within CMPC-derived exosomes, a zymogram was performed as described before [28]. In the exosomes from CMPCs, we observed the presence of gelatinase and collagenase activity, mainly the presence of inactive, intermediate and active MMP-2 (Fig. 3A). Blocking MMP activity with Marimastat, the migratory response of the HMECs upon exosomal stimulation could completely be blocked (data not shown), but because of the direct effect of Marimastat on HMECS we could not exclude the inhibition of MMPs that are released from these endothelial cells.
Fig 3.
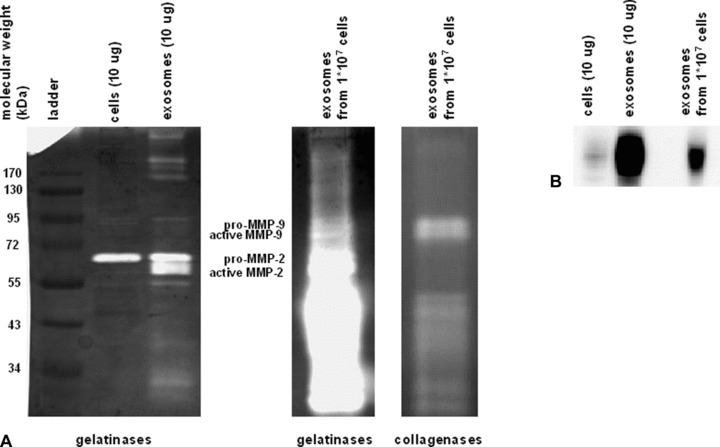
MMPs and EMMPRIN are present in CMPC-derived exosomes. (A) A zymogram was performed to detect gelatolytic MMP activity within the exosomes. The most abundant bands identify MMP-2 in the pro, intermediate and active form being predominantly expressed. (B) Using Western blotting, EMMPRIN (47–55 kD) is detected in the exosomes.
Because a membrane-bound MMP activator, EMMPRIN, has been demonstrated to be released via microvesicle shedding as well [29], we suggested that CMPC-derived exosomes might display EMMPRIN and thereby stimulate HMEC migration. As displayed in Fig. 3B, CMPC-derived exosomes contain an enriched level of EMMPRIN, as compared to total cell lysate. EMMPRIN is known to induce MMP and VEGF release from neighbouring cells, thereby inducing angiogenic responses and cell migration. To elucidate the causal role of exosomal-expressed EMMPRIN on HMEC migration, we performed a scratch assay with exosomes incubated with an EMMPRIN neutralizing antibody [30]. We observed that the enhanced closure of the wound via exosomes is completely blocked when the exosomes are treated with an EMMPRIN neutralizing antibody (Fig. 4).
Fig 4.
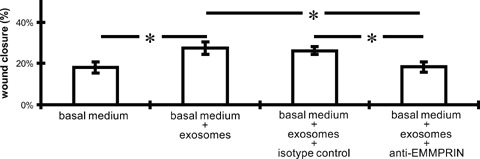
Exosomal signalling enhances migration of HMECs via EMMPRIN. Exosomes, used to stimulate HMEC cells in a scratch wound assay for 6 hrs, are treated with an EMMPRIN neutralizing antibody (Fitzgerald; 10R-CD147AHU; 20 μg/ml) or with an appropriate isotype control. Excess neutralizing antibody is washed away by ultracentrifugation at 100,000 ×g. The beneficial exosome effect (derived from 1 ×107 CMPCs) is blocked when exosomes are treated with the EMMPRIN antibody (n= 4; error bars = S.E.M.; *=P < 0.05).
Discussion
Upon CMPC cell transplantation into the myocardium, the engrafted cells contributed to formation of new capillaries and cardiomyocytes by direct differentiation, which potentially can lead to improved heart function and morphology [10–12]. However, only few donor-derived cells could be detected after 3 months [12], suggesting a loss of transplanted cells but a stimulation of endogenous cardiac recovery. The transplanted CMPCs most likely produce paracrine factors that enhance this endogenous regeneration potential, including cardiomyogenesis and angiogenesis.
Here, we studied in vitro the role of one of these factors, called exosomes. Exosomes from CMPCs are able to enhance migration of endothelial cells via an EMMPRIN-mediated mechanism. This suggests that exosomes, released by CMPCs upon transplantation, might be involved in the activation of endogenous cells and thereby result in increased capillary density. The role of MMPs and their regulators are well established in migration and angiogenesis. Here, we show that CMPC-derived exosomes contain several MMPs and this might suggest that exosomes themselves are able to breakdown the extracellular matrix or activate pro-active MMPs. Additionally, EMMPRIN, located on the exosomes, might activate target cell MMP release and/or activity.
The effect of CMPC-derived exosomes in other processes that are important for cardiac regenerative responses, such as survival, proliferation and differentiation, are unknown. However, CM from progenitor cells, containing exosomes, can affect proliferation, survival and differentiation of cardiomyocytes, cardiac fibroblasts and cardiac progenitor cells [17–20]. Moreover, Lai and colleagues identified exosomes as the active component within MSC-CM that resulted in cardiac protection upon reperfusion injury [20, 21, 25].
The full capacity of CMPCs is of major interest because of its regenerative potential via direct differentiation and through the proposed enhanced activation of endogenous cells. CMPC-derived exosomes can enhance migration of endothelial cells in vitro. The potential of CMPC and other progenitor cell-derived exosomes in different processes, such as proliferation, differentiation and survival on cardiomyocytes, smooth muscle cells and endogenous progenitor cells, are subject to research.
Acknowledgments
Source of funding: This research forms part of the Project P1.04 SMARTCARE of the research program of the BioMedical Materials institute, co-funded by the Dutch Ministry of Economic Affairs (KV). Additionally we would like to thank the Netherlands Heart Foundation and the Prof. R.L.J van Ruyvenstichting for their financial contribution.
Conflict of interest
The authors confirm that there are no conflicts of interest.
References
- 1.Segers VF, Lee RT. Stem-cell therapy for cardiac disease. Nature. 2008;451:937–42. doi: 10.1038/nature06800. [DOI] [PubMed] [Google Scholar]
- 2.Abdel-Latif A, Bolli R, Tleyjeh IM, et al. Adult bone marrow-derived cells for cardiac repair: a systematic review and meta-analysis. Arch Intern Med. 2007;167:989–97. doi: 10.1001/archinte.167.10.989. [DOI] [PubMed] [Google Scholar]
- 3.Chamuleau SA, Vrijsen KR, Rokosh DG, et al. Cell therapy for ischaemic heart disease: focus on the role of resident cardiac stem cells. Neth Heart J. 2009;17:199–207. doi: 10.1007/BF03086247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Thomson JA, Itskovitz-Eldor J, Shapiro SS, et al. Embryonic stem cell lines derived from human blastocysts. Science. 1998;282:1145–7. doi: 10.1126/science.282.5391.1145. [DOI] [PubMed] [Google Scholar]
- 5.Nussbaum J, Minami E, Laflamme MA, et al. Transplantation of undifferentiated murine embryonic stem cells in the heart: teratoma formation and immune response. FASEB J. 2007;21:1345–57. doi: 10.1096/fj.06-6769com. [DOI] [PubMed] [Google Scholar]
- 6.Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–76. doi: 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- 7.Badorff C, Brandes RP, Popp R, et al. Transdifferentiation of blood-derived human adult endothelial progenitor cells into functionally active cardiomyocytes. Circulation. 2003;107:1024–32. doi: 10.1161/01.cir.0000051460.85800.bb. [DOI] [PubMed] [Google Scholar]
- 8.Orlic D, Kajstura J, Chimenti S, et al. Bone marrow cells regenerate infarcted myocardium. Nature. 2001;410:701–5. doi: 10.1038/35070587. [DOI] [PubMed] [Google Scholar]
- 9.Goumans MJ, De Boer TP, Smits AM, et al. TGF-[beta]1 induces efficient differentiation of human cardiomyocyte progenitor cells into functional cardiomyocytes in vitro. Stem Cell Research. 2008;1:138–49. doi: 10.1016/j.scr.2008.02.003. [DOI] [PubMed] [Google Scholar]
- 10.Smits AM, Van Vliet P, Metz CH, et al. Human cardiomyocyte progenitor cells differentiate into functional mature cardiomyocytes: an in vitro model for studying human cardiac physiology and pathophysiology. Nat Protocols. 2009;4:232–43. doi: 10.1038/nprot.2008.229. [DOI] [PubMed] [Google Scholar]
- 11.Van Vliet P, Roccio M, Smits AM, et al. Progenitor cells isolated from the human heart: a potential cell source for regenerative therapy. Neth Heart J. 2008;16:163–9. doi: 10.1007/BF03086138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Smits AM, Van Laake LW, Den Ouden K, et al. Human cardiomyocyte progenitor cell transplantation preserves long-term function of the infarcted mouse myocardium. Cardiovasc Res. 2009;83:527–35. doi: 10.1093/cvr/cvp146. [DOI] [PubMed] [Google Scholar]
- 13.Gnecchi M, Zhang Z, Ni A, et al. Paracrine mechanisms in adult stem cell signaling and therapy. Circ Res. 2008;103:1204–19. doi: 10.1161/CIRCRESAHA.108.176826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Tang XL, Rokosh G, Sanganalmath SK, et al. Intracoronary administration of cardiac progenitor cells alleviates left ventricular dysfunction in rats with a 30-day-old infarction. Circulation. 2010;121:293–305. doi: 10.1161/CIRCULATIONAHA.109.871905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Vrijsen KR, Chamuleau SAJ, Noort WA, et al. Stem cell therapy for end-stage heart failure: indispensable role for the cell. Current Opinion in Organ Transplantation. 2009;14:560–5. doi: 10.1097/MOT.0b013e328330389e. [DOI] [PubMed] [Google Scholar]
- 16.Ebelt H, Jungblut M, Zhang Y, et al. Cellular cardiomyoplasty: improvement of left ventricular function correlates with the release of cardioactive cytokines. Stem Cells. 2007;25:236–44. doi: 10.1634/stemcells.2006-0374. [DOI] [PubMed] [Google Scholar]
- 17.Chimenti I, Smith RR, Li TS, et al. Relative roles of direct regeneration versus paracrine effects of human cardiosphere-derived cells transplanted into infarcted mice. Circ Res. 2010;106:971–80. doi: 10.1161/CIRCRESAHA.109.210682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Crisostomo PR, Abarbanell AM, Wang M, et al. Embryonic stem cells attenuate myocardial dysfunction and inflammation after surgical global ischemia via paracrine actions. Am J Physiol Heart Circ Physiol. 2008;295:H1726–35. doi: 10.1152/ajpheart.00236.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gnecchi M, He H, Noiseux N, et al. Evidence supporting paracrine hypothesis for Akt-modified mesenchymal stem cell-mediated cardiac protection and functional improvement. FASEB J. 2006;20:661–9. doi: 10.1096/fj.05-5211com. [DOI] [PubMed] [Google Scholar]
- 20.Timmers L, Lim SK, Arslan F, et al. Reduction of myocardial infarct size by human mesenchymal stem cell conditioned medium. Stem Cell Res. 2007;1:129–37. doi: 10.1016/j.scr.2008.02.002. [DOI] [PubMed] [Google Scholar]
- 21.Lai RC, Arslan F, Tan SS, et al. Derivation and characterization of human fetal MSCs: An alternative cell source for large-scale production of cardioprotective microparticles. J Mol Cell Cardiol. 2010;48:1215–24. doi: 10.1016/j.yjmcc.2009.12.021. [DOI] [PubMed] [Google Scholar]
- 22.Thery C, Amigorena S, Raposo G, et al. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protoc Cell Biol. 2006:01–29. doi: 10.1002/0471143030.cb0322s30. Chapter 3: Unit 3.22. [DOI] [PubMed] [Google Scholar]
- 23.Thery C, Zitvogel L, Amigorena S. Exosomes: composition, biogenesis and function. Nat Rev Immunol. 2002;2:569–79. doi: 10.1038/nri855. [DOI] [PubMed] [Google Scholar]
- 24.Valadi H, Ekstrom K, Bossios A, et al. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9:654–9. doi: 10.1038/ncb1596. [DOI] [PubMed] [Google Scholar]
- 25.Lai RC, Arslan F, Lee MM, et al. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010 doi: 10.1016/j.scr.2009.12.003. doi: 10.1016/j.scr.2009.12.003. [DOI] [PubMed] [Google Scholar]
- 26.Stetler-Stevenson WG. Matrix metalloproteinases in angiogenesis: a moving target for therapeutic intervention. J Clin Invest. 1999;103:1237–41. doi: 10.1172/JCI6870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hakulinen J, Sankkila L, Sugiyama N, et al. Secretion of active membrane type 1 matrix metalloproteinase (MMP-14) into extracellular space in microvesicular exosomes. J Cell Biochem. 2008;105:1211–8. doi: 10.1002/jcb.21923. [DOI] [PubMed] [Google Scholar]
- 28.Sluijter JPG, Verloop RE, Pulskens WPC, et al. Involvement of furin-like proprotein convertases in the arterial response to injury. Cardiovasc Res. 2005;68:136–43. doi: 10.1016/j.cardiores.2005.05.016. [DOI] [PubMed] [Google Scholar]
- 29.Sidhu SS, Mengistab AT, Tauscher AN, et al. The microvesicle as a vehicle for EMMPRIN in tumor-stromal interactions. Oncogene. 23:956–63. doi: 10.1038/sj.onc.1207070. [DOI] [PubMed] [Google Scholar]
- 30.Tang Y, Nakada MT, Kesavan P, et al. Extracellular matrix metalloproteinase inducer stimulates tumor angiogenesis by elevating vascular endothelial cell growth factor and matrix metalloproteinases. Cancer Res. 2005;65:3193–9. doi: 10.1158/0008-5472.CAN-04-3605. [DOI] [PubMed] [Google Scholar]