

Abstract
Study Objectives:
To test the hypothesis that rapid eye movement (REM) sleep contributes to the consolidation of new memories, whereas non-rapid eye movement (NREM) sleep contributes to the prevention of retroactive interference.
Design:
Randomized, crossover study.
Setting:
Two sessions of either a morning nap or wakefulness.
Participants:
Twenty-five healthy young adults.
Interventions:
Declarative learning of word pairs followed by a nap or a wake interval, then learning of interfering word pairs and delayed recall of list A.
Measurements and Results:
After a restricted night (24:00-06:00), participants learned a list of word pairs (list A). They were then required to either take a nap or stay awake during 45 min, after which they learned a second list of word pairs (list B) and then had to recall list A. Fifty percent of word pairs in list B shared the first word with list A, resulting in interference. Ten subjects exhibited REM sleep whereas 13 subjects exhibited NREM stage 3 (N3) sleep. An interference effect was observed in the nap but not in the wake condition. In post-learning naps, N3 sleep was associated with a reduced interference effect, which was not the case for REM sleep. Moreover, participants exhibiting N3 sleep in the post-learning nap condition also showed a reduced interference effect in the wake condition, suggesting a higher protection ability against interference.
Conclusion:
Our results partly support the hypothesis that non-rapid eye movement sleep contributes in protecting novel memories against interference. However, rapid eye movement sleep-related consolidation is not evidenced.
Citation:
Deliens G; Leproult R; Neu D; Peigneux P. Rapid eye movement and non-rapid eye movement sleep contributions in memory consolidation and resistance to retroactive interference for verbal material. SLEEP 2013;36(12):1875-1883.
Keywords: Declarative learning, memory consolidation, nap, retroactive interference, sleep stages
INTRODUCTION
A positive contribution of post-learning sleep in the consolidation of new information in long-term memory is now widely recognized.1–7 Beneficial effects of post-learning sleep have been highlighted for various memory systems, such as verbal declarative memory,8,9 emotional memory,10,11 or spatial learning,12 and non-declarative motor procedural learning13 or priming.14 Like long-term memory, sleep is not a unitary process but comprises different subsystems.15,16 In humans, it is characterized by two prominent stages, rapid eye movement (REM) and non-rapid eye movement (NREM) sleep. NREM is further subdivided17 into stage 1 (N1), stage 2 (N2) and slow wave sleep (SWS or N3). Sleep stages, like memory subsystems, are differentiable at neuroanatomical, neurophysiological, and behavioral levels.3,18 Notwithstanding, how sleep stages specifically contribute to the consolidation of different memory types remains a matter of debate.
According to the dual-process hypothesis,19–21 each sleep stage would preferentially contribute to the post-learning consolidation of a specific memory system. In support of this hypothesis, several studies have highlighted a beneficial effect of SWS for the consolidation of declarative memories and of REM sleep for the consolidation of procedural memories.4,5 However, other studies have shown reverse dissociations according to the task or to the presence of a superimposed emotional modulation,3,16 casting doubts on the operational value of this model. An alternative hypothesis posits that the sequence of sleep stages during a normal sleep cycle (always NREM sleep first, then REM sleep) reflects the succession of cerebral processes supporting the consolidation of new information in memory.22–25 This double-step hypothesis is partially supported by the work of Giuditta et al.24 showing that in rats, regular NREM-REM sleep cycles are necessary to optimize post-learning retention performance. Accordingly in man, Ficca et al.22 found that sleep cycle fragmentation impairs consolidation of declarative memories. It must be emphasized here that these two hypotheses (dual-process and double-step) are not contradictory, but could reflect different facets of post-learning consolidation. Still, the processes implicated at each sleep stage and their relationships with memory consolidation remain open issues. In this perspective, Scrima1 suggested, based on a study in narcoleptic patients, that SWS prevents retroactive interference on recently acquired information, whereas REM sleep consolidates information.
Jenkins and Dallenbach26 proposed in 1924 that sleep would passively protect memories against ongoing interference. However, recent studies have demonstrated that more active sleep mechanisms are at play to support the consolidation of new memories and protect them against interference. For instance, using a classic AB-AC interference paradigm, Ellenbogen et al.27 asked participants to memorize a list of word pairs (AB), before spending 12 h either asleep (at night) or awake (during daytime). Before retrieving list AB, half of the participants had to memorize a series of novel word pairs (AC), in which the first word of the pair was the same as in list AB, hence creating interference, whereas the other half directly proceeded to cued recall of list AB. Results showed better recall of list AB after sleep than wakefulness for material submitted to retroactive interference (i.e., recalling list AB just after learning list AC), and more so than for noninterfering material. This effect was confirmed in a subsequent study28 in which participants were tested first in noninterfering then in interfering conditions within the same retrieval session.
Although an effect of sleep against retroactive interference has been evidenced for declarative learning27,28 in the framework of total post-learning sleep deprivation paradigms, the contribution of different stages of sleep (SWS, REM) in this process remains to be ascertained. In this respect, the aim of the current study was to test the hypothesis1 of a consolidating role of REM sleep on newly formed memory traces, and of a protecting role of SWS from retroactive interference linked to subsequent exposure to a new material interfering with the information already encoded. We tested this hypothesis in a morning nap paradigm aimed at promoting predominant post-training REM or NREM sleep episodes. Indeed, classic sleep architecture is composed of alternating NREM and REM sleep episodes, with NREM at sleep onset and REM latency ranging from 60 to 90 min.17 REM sleep latency shorter than 20 min, called SOREMS (REM at sleep onset) is characteristic of narcolepsy29 but can also be observed in short naps when participants are under appropriate homeostatic and circadian conditions. Indeed, a high circadian drive for REM sleep is observed in late-night and morning sleep30–34 but also when NREM propensity is reduced and REM propensity is increased.35,36 Thus, SOREMS37 can be induced, first by depriving participants from the end of their night of sleep (which is the richer in REM sleep), eventually reducing NREM propensity but also inducing partial REM deprivation; and second, by then allowing a nap in the morning when REM circadian propensity is high.
In our protocol, 25 volunteers slept in the laboratory under polysomnographic (PSG) recording for 6 h after bedtime. In the morning, they learned a list of unrelated word pairs (A), then were allowed to sleep again or were kept awake for 45 min, under electroencephalographic (EEG) control. At noon, a novel list of word pairs (B) was learned just before the delayed recall of list A. List B was composed of 50% word pairs in which the first word of the pair was presented in list A, hence creating interference. The same procedure was repeated 1 mo later in the other condition (morning sleep versus wake), in a within-subject design. Hence our protocol was designed to conduct within-subject comparisons between post-learning sleep and wake conditions, as well as to compare subjects who took a nap predominantly in REM or SWS, allowing us to test the role of sleep stages in consolidation and resistance to interference for newly learned information.
METHOD
Participants
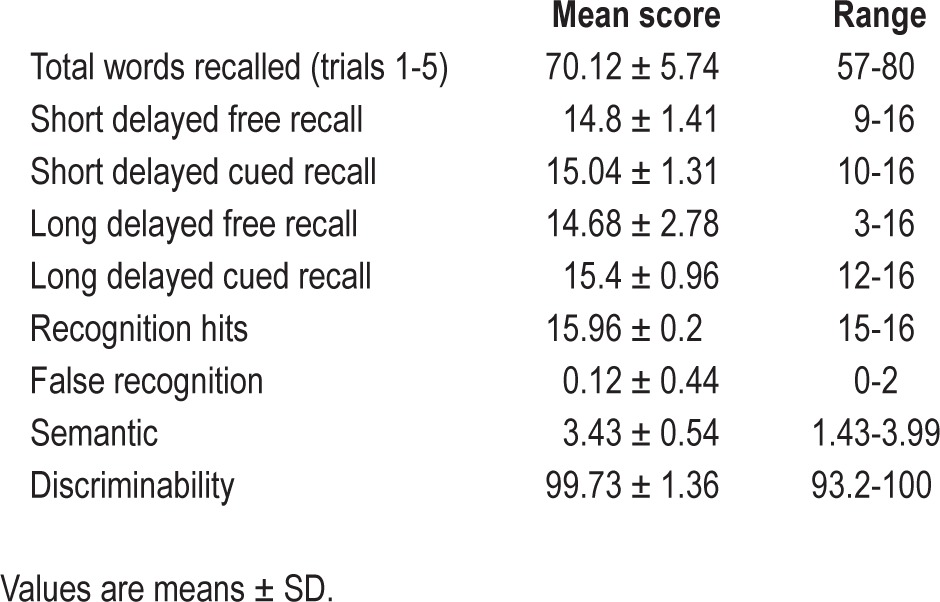
Twenty-six healthy participants gave their written informed consent to participate in this study approved by local ethics committees. They were required to keep regular sleep patterns during the week before and throughout the experiment, and to refrain from alcohol and stimulant drinks. The regularity of their sleep habits was controlled using actimeter recording (Daqtometer, Daqtix GbR, Oetzen, Germany) and the completion of daily sleep logs (St. Mary's Hospital sleep questionnaire38). Due to an irregular sleep pattern, one subject was excluded from the statistical analyses. The 25 remaining participants (26.2 ± 4.7 y, mean ± standard deviation [SD], 11 males) met the following criteria: native French speakers, non- smokers or fewer than 10 cigarettes per day, no medical history of neurological or sleep disorders (Pittsburgh Sleep Quality Index39 total score < 9; Fatigue Severity Scale40 1.88 ± 0.64), no current or past affective disorders (Beck Depression Scale41 < 7; Structured Mini International Neuropsychiatric Interview42 for Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition [DSM-IV]), intermediate or neutral chronotype (Horne and Ostberg Morningness-Eveningness Questionnaire,43 range 31-75), and verbal intelligence within normative values (Mill Hill vocabulary scale,44 36.32 ± 3.4). Finally, to screen for long-term memory deficits, we administered the California Verbal Learning Test (French adaptation45; all participants fell within the normal range for short- and long-term delay recall [Table 1]).
Table 1.
California verbal learning task
Materials
Retroactive Interference Verbal Learning Task
A computerized A-B interference paradigm was adapted from Ellenbogen et al.27 We created four different lists (A1, B1, A2, B2) of 28 semantically unrelated French bisyllabic word pairs, emotionally neutral and matched for lexical frequency,46 imageability, and concreteness.47 Lists B1 (respectively B2) included 50% of word pairs for which the initial word was presented in list A1 (respectively A2) but was associated with a new word (example: A list: cheval - salon [horse - lounge]; B list: cheval - canal [horse - channel]), hence creating interference. The other 50% in list B1 (respectively B2) were new word pairs. Consequently, during delayed recall of list A after learning the corresponding list B, 50% of the word pairs in the list A were subjected to retroactive interference from the list B, whereas the other 50% were not. During learning and recall sessions, word pairs of each list were randomly presented. A2 and B2 lists were paralleled versions of A1 and B1 lists respectively, alternatively used for the nap and wake conditions in our within-subject paradigm. Conditions and list versions were counterbalanced across subjects.
Procedure
After 1 w of actimetry recordings and sleep diaries, participants stayed for 2 consecutive nights in the sleep laboratory. The first night (from 24:00 to 08:00) served as an adaptation night to sleep in the laboratory under PSG recording conditions, but also as a screening night to exclude participants presenting any sleep disorder. On the second night, subjects underwent PSG recording for a time in bed of 6 h (± from 24:00 to 06:00). At 09:00, after an isocaloric breakfast, they learned a list of 28 unrelated word pairs (A). Then at 10:00, they were allowed to sleep again or were kept awake under polygraphic recording for 45 min. In the wake condition, subjects stayed alone lying down in their bed; they were allowed to watch movies or read a book. At 12:00, a novel list of word pairs (B) was learned just before the delayed recall of list A. List B was composed of 50% word pairs in which the first word of the pair was presented in list A, hence creating interference. The same procedure was repeated 1 mo later in the other condition (morning sleep versus wake) but without the adaptation night. An overview of the experimental design is illustrated in Figure 1.
Figure 1.
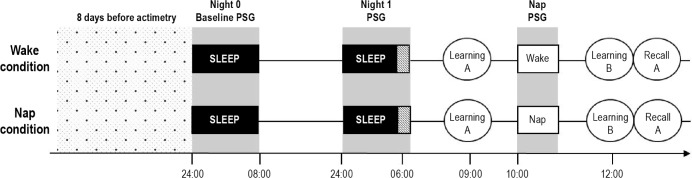
Schematic representation of the protocol. Randomized crossover design with nap or wake conditions. (A) initial word pairs list (version 1 or 2): learning session followed by immediate recall testing; (B) interference word pairs list (version 1 or 2).
For each list, the 28 unrelated word pairs were displayed one by one on the computer screen for 3 sec, followed by a 5-sec white fixation cross, turning red to warn the subject of the next pair's presentation. After the presentation of the 28 word pairs, an immediate cued recall procedure was administered: the first word of each pair appeared on the computer screen, and subjects had to type the associated word. The correct response was then presented on the screen. The cued recall procedure was repeated until subjects were able to recall at least 21 word pairs (75% learning criterion). However, when the response for a pair was correct, it was not presented anymore to avoid over-consolidation. The word pairs were presented in random order over repeated trials to prevent serial learning. The cued delayed recall phase was administered immediately after learning of list B. Cued delay recall was administered using the same procedure as for learning, but pair cues were presented only once and no feedback on performance was provided.
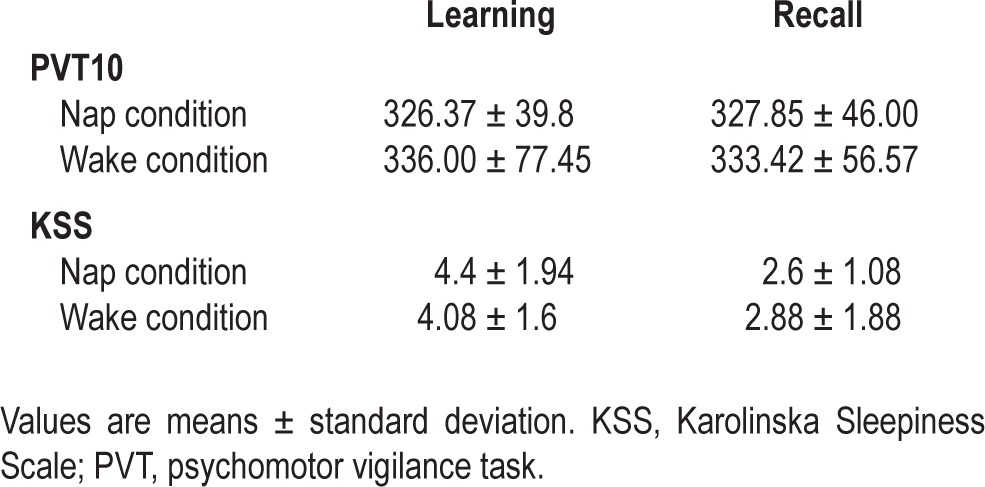
Additionally, the psychomotor vigilance task48 (PVT, 10 min) and the Karolinska Sleepiness Scale49 (KSS) were administered before each learning and test sessions to assess variations in objective and subjective vigilance levels, respectively.
EEG Recordings and Analysis
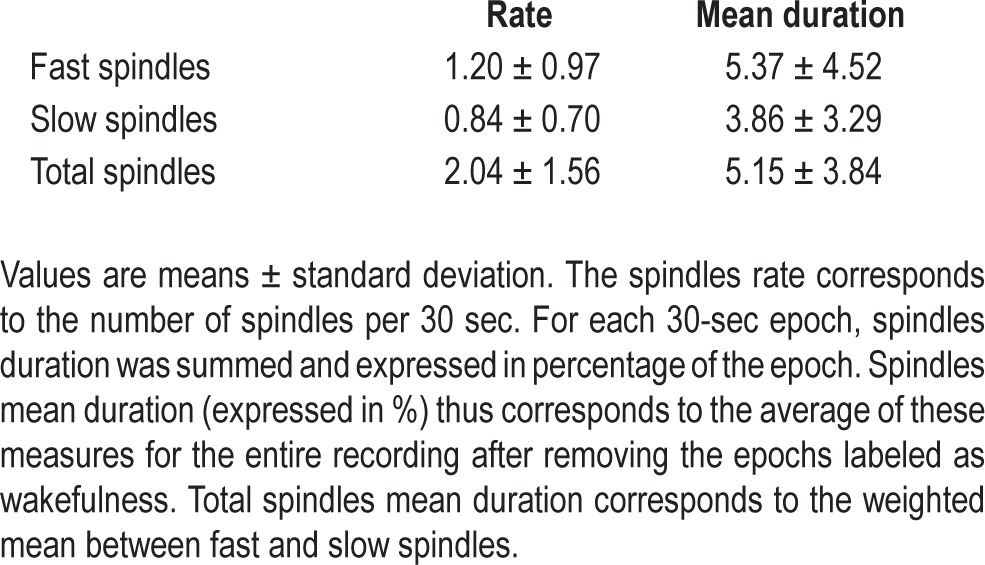
Polygraphic recordings were assessed during nights 0 and 1 (Figure 1), as well as during the scheduled 45-min naps and waking periods. EEG (scalp electrodes at C3-A2, O1-A2, FP1-A2 derivations according to the International 10-20 system), electromyography (EMG), and electrooculograms (EOG) were recorded at a sampling rate of 200 Hz. For the baseline night, recordings included bilateral anterior tibial electromyograms, an oral and nasal airflow recorded by an oronasal cannula (Pro-Flow Plus™ Pro-Tech® Mukilteo, WA, USA), respiratory effort measured by thoracic and abdominal belts (Pro-Tech® CT2™, Mukilteo, WA, USA) and capillary oxygen saturation monitored by photosensitive finger oximetry (Nonin® Flexi-Form® II 7000A Nonin Medical Inc, Minneapolis, MN, USA and LINOP® Adt Masimo Corp., Irvine, CA, USA). For all recordings, a low bandpass filter at 45 Hz and a high pass filter at 0.75 Hz were applied to EEG channels. According to American Academy of Sleep Medicine (AASM) criteria,17 sleep recordings were subsequently visually scored as wake stages N1, N2, SWS (N3) and REM on 30-sec epochs. Sleep onset corresponded to the time of the first 30-sec epoch scored either N1 (provided that there were three consecutive pages of N1), N2, SWS, or REM. Awakening was defined as the time corresponding to the last 30-sec epoch scored N2, SWS, or REM. Sleep latency was the time interval from lights-off to sleep onset, sleep period time (SPT), the time interval from sleep onset to final awakening, total sleep time the sleep period minus the total duration of intrasleep awakenings, and sleep maintenance the total sleep time divided by sleep period time. Sleep scoring was performed by one experimenter (GD); atypical recordings (SOREMS) and sleep staging were systematically reviewed by a specialist physician trained in sleep medicine (DN). Additionally, spindle density was calculated as the mean number of sleep spindles per 30-sec epoch using the PRANA software package (PhiTools, Strasbourg, France). Sleep spindles were detected automatically on C3-A2 channel based on the criteria from Anderer et al.50,51—(1) a minimal amplitude of 12 μV; (2) a spindle duration 0.3-2.0 s; and (3) a frequency range of 11-13 Hz for slow spindles and of 13-16 Hz for fast spindles.
Finally, a spectral analysis was performed on the central EEG signal (C3-A2) using the PRANA software package (PhiTools, Strasbourg, France). Artifacts due to eyes' and muscles' movements were automatically detected and removed prior to the spectral analysis. EEG signal was then visually checked for the accuracy of the computerized artifact detection, and additional epochs of the EEG recording that contained artifacts were marked. Then, a fast Fourier transform was computed on 4-sec intervals (hence a frequency resolution of 0.25 Hz). Power spectra were averaged over 30-sec intervals to match the sleep stage scoring and subsequent power of standard frequency bands was computed (δ: 0.75-4.5 Hz; θ: 4.5-8.5 Hz; α: 8.5-12.5; σ: 12.5-15.5; β: 15.5-22.5; γ: 22.5-45.5). Intervals containing more than 50% of artifacts were considered as missing data. Summary variables included total amount and mean powers during the nap, in addition to the total amount during NREM and total amount during REM sleep.
Statistical Analyses
Statistical analyses were performed using Statistica 7.0 (Statsoft Inc., Tulsa, OK). Analyses were conducted using repeated-measures analyses of variance (ANOVAs), followed by Tukey post hoc tests when appropriate. Significance was set at P < 0.05, two-sided. Relationships between variables were evaluated using Pearson coefficients of correlation.
RESULTS
Sleep and Vigilance
Night: PSG Data
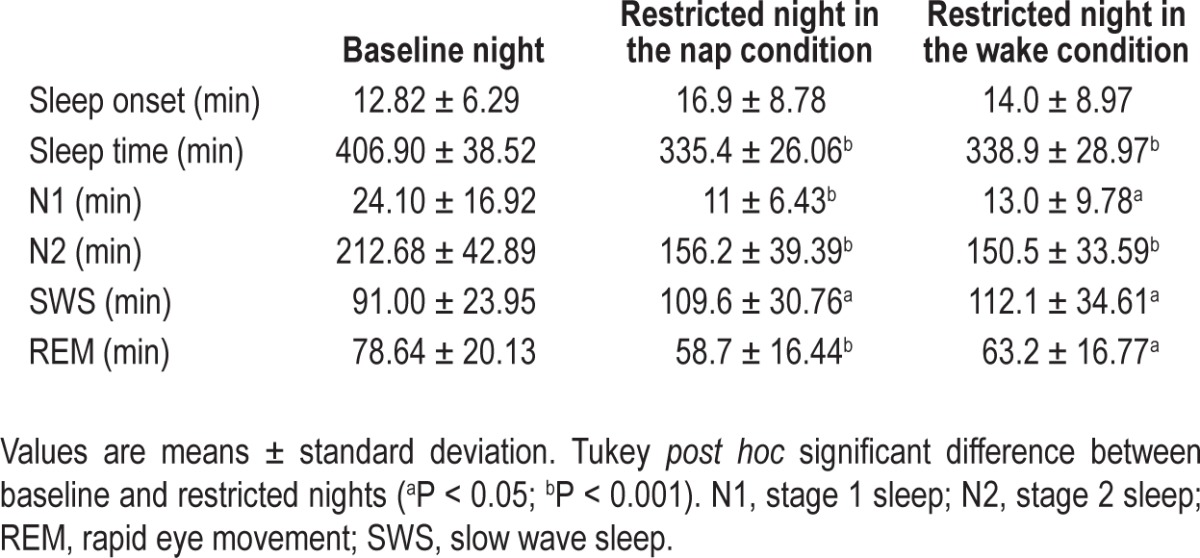
Sleep variables for the baseline night, the restricted night before the nap condition, and the restricted night before the wake condition are summarized Table 2. A repeated-measures ANOVA with the within-subject night factor (baseline versus night before nap versus night before wake) disclosed a main effect for total sleep duration (F2,48 = 49.82; P < 0.001) and for the duration of each sleep stage (N1: F2,48 = 11.51; P < 0.001, N2: F2,48 = 25.09; P < 0.001, SWS: F2,48 = 5.60; P < 0.01, REM: F2,48 = 10.73; P < 0.001) variables, but no significant difference for sleep onset latency (SOL) (F2,48 = 1.98; P > 0.15). Tukey post hoc tests disclosed significant differences between the baseline night and both of the restricted nights with a higher mean duration of total sleep time, stage 1, stage 2, REM, and a lower amount of SWS. Most importantly, no significant difference was found between the restricted night before the nap condition and the restricted night before the wake condition (Table 2).
Table 2.
Sleep
Nap Conditions: PSG Data
Sleep time during the nap condition averaged 32.5 ± 9.6 min with a sleep latency of 5.2 ± 4.6 min. Of the 25 participants, 10 exhibited REM in their sleep (mean REM duration 5.7 ± 5.0 min, NREM S2 16.4 ± 10.9 min) whereas 13 exhibited SWS (mean SWS duration 6.6 ± 6.4 min, S2 26.0 ± 5.8 min). Among the 10 subjects who exhibited REM sleep, eight presented with sleep onset REM sleep (SOREMS; REM latency < 20 min). There were six subjects with REM sleep and no SWS, nine subjects with SWS and no REM sleep, four subjects with both SWS and REM sleep, and six subjects with no SWS nor REM sleep. No sleep features were detected on the EEG recordings during the wake condition, for all participants. Results of the spindles analysis are summarized in Table 3.
Table 3.
Sleep spindles
Vigilance
A repeated-measures ANOVA conducted on PVT mean reaction times (RTs) with within-subjects factors sleep (nap versus awake) and session (learning versus recall) failed to disclose any main effect or any interaction (all P > 0.4), indicating similar vigilance levels all along the experiment. ANOVAs conducted on median reactions times, numbers of lapses with RTs higher than 500 msec, and number of lapses with RTs higher than 2 SDs of the mean also failed to disclosed any main effect or interaction (all P > 0.1). The same analysis conducted on subjective vigilance scores (KSS) disclosed a higher self-reported sleepiness for the recall than for the encoding session (F1,24 = 47.37, P = 0.000), but no effect of sleep and no interaction between sleep and session factors (all P > 0.2). Mean values for PVT and KSS scores are reported in Table 4.
Table 4.
Vigilance parameters
Encoding Data
A repeated-measures ANOVA conducted on the number of trials needed to reach the learning criteria (75% of correct answer) at immediate recall with within-subjects factors sleep (nap versus awake) and session (list A versus list B) failed to disclose any main or interaction effects (nap condition: list A 1.7 ± 0.9, list B 1.9 ± 0.9; wake condition: list A 1.8 ± 0.8, list B 1.9 ± 0.8; all P > 0.2).
We also conducted a repeated-measures ANOVA on the number of interference and new pairs not correctly recalled at the last trial, separately for the wake and sleep conditions. Results disclosed no main effect of sleep (nap versus awake, F1,22 = 0.6; P > 0.4) and no interaction between sleep and word pairs category (new versus interference pairs, F1,22 = 0.9; P > 0.3). However, there was a main effect of pairs category with a higher number of interference than new pairs (mean ± SD 2.3 ± 1.3 versus 1.7 ± 1.1; F1,22 = 6.6; P < 0.05) not recalled. Hence, this result suggests that interference pairs in list B were more difficult to encode, showing a classical proactive interference effect, but that this effect was not modulated by the prior sleep versus nap condition.
Interference Paradigm
Sleep and Interference
At delayed recall, the mean number of correctly recalled word pairs (whether or not subjected to interference) out of a maximum of 14 was 9.64 ± 0.52 in the nap condition and 9.64 ± 0.54 in the wake condition.
A repeated-measures ANOVA on the number of correctly recalled word pairs with within-subject factors sleep (nap versus awake) and interference (word pairs from list A subjected [IP] versus not subjected [NIP] to interference) (Figure 2) disclosed a main interference effect (F1,24 = 9.51, P = 0.005) and a sleep*interference interaction (F1,24 = 5.28, P = 0.03), with no main effect of sleep (F1,24 = 0.00, P = 1). Tukey post hoc test disclosed a highly significant interference effect in the nap condition exclusively, with a lower correct recall score for IP (8.72 ± 0.63) than NIP (10.56 ± 0.50; P = 0.0001) pairs. The effect was not significant in the wake condition (IP 9.24 ± 0.57 versus NIP 10.04 ± 0.63; P = 0.09). Recall of word pairs not subjected to interference (NIP) was similar in the nap and wake conditions (P > 0.3). Recall of word pairs subjected to interference was also similar in both conditions (IP, P > 0.3).
Figure 2.
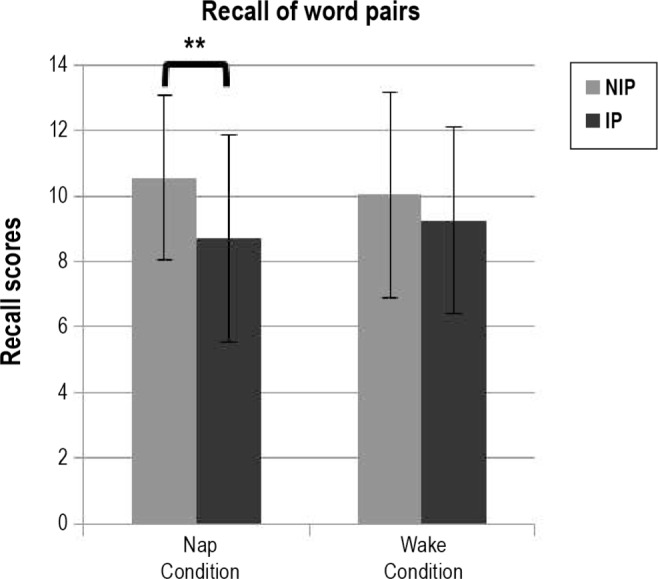
Interference effects. Recall scores (number of correctly recalled words from list A) for pairs subjected (IP) and not subjected (NIP) to interference in nap and wake conditions. Significant differences between conditions and stimulus types are indicated by asterisks, ** < 0.001.
Contributions of Sleep Stages
In the nap condition, independent Student t-tests were conducted to compare recall performance with (≥ 30 sec) or without SWS and with (≥ 30 sec) or without REM sleep. Recall of word pairs not subjected to interference (NIP) was similar in naps (a) with and without REM (9.9 ± 2.1 versus 11.0 ± 2.7; t23 = 1.1; P > 0.3), and (b) with and without SWS (11.2 ± 2.6 versus 9.8 ± 2.3; t23 = -1.4; P > 0.1). Recall of word pairs subjected to interference (IP) was similar in naps with and without REM (7.5 ± 2.6 versus 9.5 ± 3.3; t23 = 1.6; P > 0.1), but was higher in naps with SWS, as compared to those without SWS (10.2 ± 2.2 versus 7.1 ± 3.3; t23 = -2.8; P < 0.01). Interference effects (correct recall of NIP minus correct recall of IP) diminished after a nap with SWS as compared to a nap without SWS (1.0 ± 2.2 versus 2.7 ± 2.0; t23 = 2.1; P < 0.05), whereas no difference was found between naps with versus without REM sleep (2.4 ± 2.0 versus 1.5 ± 2.4; t23 = -1.0; P > 0.3). Additionally, t-tests against the null hypothesis revealed an absence of interference after a nap with SWS (interference score not different from the 0 value; t12 = 1.6; P > 0.1), whereas the interference effect was highly significant after a nap without SWS (t11 = 4.7; P = 0.0006) (Figure 3).
Figure 3.
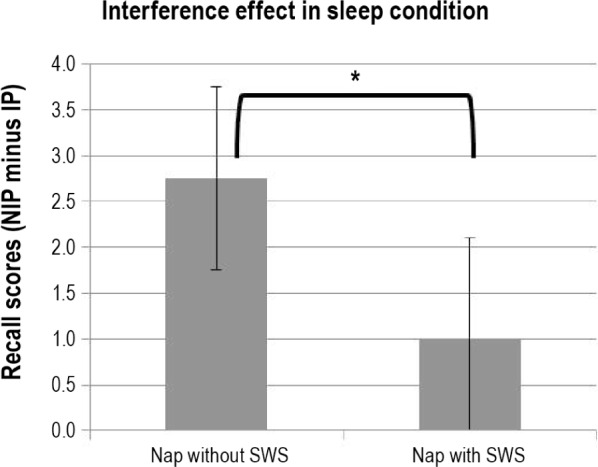
Interference effects in the sleep condition. Interference effect (recall scores for pairs not subjected (NIP) minus pairs subjected (IP) to interference) in nap groups with and without slow wave sleep (SWS). Significant differences between conditions and stimulus types are indicated by asterisks, * < 0.05.
We also investigated whether a higher proportion of SWS during sleep in some individuals (i.e., a trait in sleep architecture) could be associated with a higher ability to be protected against interference after sleep or wakefulness. Therefore, we checked whether protection against interference was also better (i.e., a lower interference effect) after the wake episode in subjects exhibiting more SWS during the post-learning nap. To do so, we conducted a mixed ANOVA on the size of the interference effect with a within-factor session (nap versus awake) and a between-factor SWS (nap with versus without SWS). Results disclosed a significant main effect of SWS presence (F1,23 = 7.0, P < 0.05) with a bigger interference effect in subjects without SWS during their nap (2.4 ± 2.2) than in subjects with SWS (0.3 ± 2.3), and a main effect of session (F1,23 = 5.0, P < 0.05) with a bigger interference effect after a nap (1.9 ± 2.2) than after wakefulness (0.8 ± 2.6). However, the interaction effect was non significant (F1,23 = 0.4, P > 0.5), suggesting a trait-like protection against interference in subjects with a higher proportion of SWS in their post-learning sleep. For the sake of completeness, the same analysis was performed with the within-factor session (nap versus awake) and the between-factor REM sleep (nap with versus without REM sleep). Results disclosed a main effect of session (F1,23 = 4.9, P < 0.05), but no main effect of REM sleep (F1,23 = 1.0, P > 0.3) nor interaction (F1,23 = 0.01, P > 0.9). Finally, we tested whether a “natural” higher proportion of SWS during sleep could be related to a better protection against interference. To do so, the proportion of SWS during the baseline night was compared between subjects with or without SWS in their post-learning nap. Results showed no significant differences in the proportion of SWS during the baseline night (t23 = 1.1, P > 0.2), suggesting that this assumption does not hold true.
Finally, correlational analyses were performed between the size of the interference (NIP minus IP) and the time spent in each sleep stage as well as between the size of the interference effect and delta, theta, alpha, and sigma power frequencies. Correlations between memory scores (separately for noninterfering and interfering word pairs) or interference scores and (a) sleep stages or (b) power frequencies were all non significant (coefficients of correlation ranging from -0.34 to +0.38; all P > 0.06), except for a significant positive correlation between N3 duration and IP scores (r = 0.4; P < 0.05). Nonparametric Spearman correlations confirmed these results. We also computed correlations between slow oscillation power (0.75-1.25 Hz) and memory scores (IP recall, NIP recall) or interference size for all subjects. Positive correlations between IP recall and slow oscillation power (mean, sum, and sum in NREM sleep) were observed (coefficients of correlation ranging from 0.45 to 0.47; all P < 0.03). All other correlations including correlations in subgroups with and without SWS were non-significant (all P > 0.11).
Likewise, no correlation was found between spindle density in N2 (both fast spindle and slow spindle density) and memory scores for noninterfering and interfering word pairs as well as between spindle density in N2 and the size of the interference effect (coefficients of correlation ranging from -0.14 to +0.27; all P > 0.4).
DISCUSSION
The current study investigated retroactive interference effects on the delayed recall of memories in a within-subject paradigm in which participants learned novel word pairs in the morning before taking a 45-min nap or spending the equivalent time awake. Afterward, they learned novel, interfering word pairs and finally had to recall the material learned before the nap or wake episode. Using a partial REM sleep deprivation paradigm by reducing the duration of the preceding night by 2 h, our protocol allowed comparisons between subjects who took a nap predominantly in REM sleep (and/or presenting SOREMS) or in SWS, hence testing the role of these sleep stages in the consolidation and resistance to interference for newly learned information. Scrima's1 hypothesis predicted a role for SWS in protecting memories from retroactive interference, and a role for REM sleep in consolidating memory traces. Our results are only partially consistent with this hypothesis. Indeed, we observed a higher resistance to retroactive interference effects after a nap containing SWS than after a nap without SWS, suggesting that SWS plays a role in protecting memories against retroactive interference.52 Because positive correlations between recall of IP pairs and slow oscillation power were found, we suggest that the beneficial effect of SWS against interference is at least partially linked to slow oscillation power. However, recall of word pairs not subjected to interference was similar in REM sleep and SWS conditions, thus failing to support the hypothesis that REM sleep would specifically consolidate these memories.
A lack of effect of post-learning REM sleep on memory consolidation in the current study might be explained by several factors. First, it is not clarified how much time spent in REM sleep is needed to efficiently initiate the cascade of neurophysiological events eventually leading to the consolidation of verbal material. In this respect, the amount of REM sleep achieved by our participants during naps was relatively low (5.7 ± 4 min). Further studies are needed to replicate our findings with protocols promoting longer REM sleep duration. Also, delayed recall took place 3 h after the initial learning phase in the current study. Therefore, we cannot exclude that REM sleep could play a role for delayed recall at a longer term (after days, weeks, or months).
The analysis of PSG data disclosed REM sleep episodes without atonia in several subjects in whom REM sleep occurred very quickly after sleep onset, i.e. SOREMS.37 It is not known at this point whether SOREMS, and especially SOREMS without atonia, presents the same qualitative features as canonical REM sleep and equally contributes in memory consolidation processes. In the current study, our manipulation during the preceding night with partial sleep deprivation was relatively successful since we observed SOREMS during subsequent morning naps in eight of 25 subjects. Another consideration is that the occurrence of REM sleep within the first 20 min after sleep onset (i.e., SOREMS) actually disrupts the canonical NREM-REM cycle. In this respect, the order of appearance of sleep stages (SWS followed by REM sleep) may be an important determinant for memory consolidation, following the sequential hypothesis24 assuming that, in a first step, SWS selects adapted memory traces and weakens the non-adapted ones by long-term depression and, in a second step REM sleep consolidates selected memories. In this view, cyclic succession of SWS and REM sleep would be required for successful memory consolidation.22–25 Consequently, in our study, the reversed or disrupted order of sleep stages observed in subjects with SOREMS may eventually result in a failure of REM sleep-dependent memory consolidation, potentially explaining why we do not observe a beneficial effect of naps on noninterfering word pairs.
As mentioned previously, our results partially support Scrima's1 hypothesis of a protective role of SWS against interference on recently learned material. Assuming that the absence of beneficial effects of REM sleep might be due to methodological features (SOREM sleep episodes instead of REM sleep episodes, too-short REM sleep duration…), our results are in line with studies proposing a complementary role of sleep stages in memory consolidation processes. Several studies have suggested an experience-dependent neuronal reactivation during post-learning SWS in animals53–55 and humans,19,56–60 and proposed that memories are reactivated and set in a labile form during SWS, before being reconsolidated and strengthened during REM sleep.61,62 During SWS, renewed lability of memories after natural reactivation would allow their integration into previously stored information62 and favors their transfer from hippocampal to neocortical stores61,63,64 throughout subsequent nights.65,66
During sleep, the reduction of external inputs allows memories to be set in a labile form and to be partially sheltered from potential sources of interference, unlike during wakefulness.67 In the current study, we found a beneficial effect of SWS on resistance to interference, as compared to participants who did not exhibit SWS during their nap. We also observed that even if a post-learning nap with SWS reduces the size of the interference effect as compared to a nap without SWS, a wake retention interval remains more beneficial against interference than a nap (with or without SWS). In addition, protection against interference was also better (i.e., a lower interference effect) after the wake episode in subjects exhibiting more SWS during the post-learning nap, suggesting that individuals who are able to dynamically obtain more SWS during a post-learning sleep episode also present a higher protection against interference after a post-training time interval spent awake.
Although these results may seemingly be in contradiction with studies and models proposing a beneficial effect of sleep on memory consolidation and protection against interference,27,28 we argue that our findings can be explained in the framework of the reconsolidation theory68 combined with the notion of pattern completion/separation.69 Indeed, the reconsolidation theory hypothesizes that the recall of previously stored information (e.g., list A) or learning of novel but related material (e.g., list B) reactivates memories in a labile form, making them again sensitive to interference, eventually necessitating their reconsolidation. In this perspective, learning the second list after the nap, and in particular the interfering word pairs, would have reactivated and labilized the corresponding words of the first list that were already consolidated during the nap. Consequently, immediate recall of the first list after its weakening by learning the second list would result in an interference effect. In the wake condition, however, information was less consolidated. Therefore, memories of list A cannot be (or are less) labilized while learning the interfering list and a dual memory trace is created. The formation of a dual memory trace is thought to depend on a pattern separation process in opposition to a pattern completion.70 The pattern separation process refers to the ability to orthogonalize overlapping memories, thus preventing forgetting by creating distinct (non- interfering) traces, whereas the pattern completion would allow to create a single common representation by reactivating old and new memories and integrating them before their consolidation.69–72 In our study, creation of a pattern separation during the second list's encoding would have allowed protection of memories from retroactive interference because the second list does not modify the first list but the two lists coexist in memory, and promotion of the first list's retention because pattern separation does not involve its reactivation during the second list's encoding.70 Although the pattern separation would confer an advantage against retroactive interference in the wake condition, it could also present several drawbacks. Indeed, creating dual memory traces entails costs in terms of energy and space requirements at the neuronal level, progressively leading memory systems to saturation.73 In contrast, sleep-consolidated memories, once reactivated by a second list and labilized, will be reconsolidated via a pattern completion integrating new memories to existing knowledge, thus preventing duplicates in memory68 and saving space and energy. However, without post-learning SWS, this pattern completion would remain sensitive to daytime reactivation. After a sleep episode without SWS, subsequent reactivation by interference learning would set back the single common representation in a labile form again sensitive to interference. By contrast, SWS might protect this single common representation from retroactive interference.
An alternative but not mutually exclusive explanation for the fact that resistance to interference was not superior in the sleep group with SWS than in the wake group could be the relatively small amount of time spent in SWS. Prior studies have suggested that approximately 20 min of SWS episode would be sufficient for memory enhancements74–76 but interfering learning before the recall session was not proposed in these studies. When introducing an interfering task, Alger et al.52 demonstrated a lesser interference effect after a 60-min nap containing 14-min of SWS as compared to a 10-min nap without SWS or to time spent awake. However, these results are in partial contradiction with Diekelmann et al.,77 who found that 40 min of sleep including 17 min of SWS may be sufficient to protect memories against interference, but only if reactivations are externally triggered during sleep (using odours), and that otherwise 90 min of sleep are needed (including 40.4 min of SWS) to observe an enhancement in memory. Like for REM sleep, further studies are needed to investigate how much time spent in SWS is needed to efficiently protect memory from interference.
Finally, as slow oscillations are temporally correlated with sleep spindles78 and spindles are thought to play a role in the hippocampal-neocortical dialogue and brain plasticity processes,56,78–84 we investigated a possible link between spindle activity and memory consolidation. Increased sleep spindle activity was previously observed following successful learning, and correlated with better recall performance,83 with a potential differentiation between slow (11-13 Hz) and fast spindles9,84 (13-16 Hz). Contrary to our expectations, no correlation emerged in our study between recall performance and fast or slow spindle density or duration. Further studies should clarify the reasons why such relationship was not observed in our particular paradigm involving morning naps after partial sleep deprivation.
To conclude, the results of the current study partially support Scrima's hypothesis that SWS would protect recently encoded memories against retroactive interference. In this framework our study confirms that sleep does more than passively shelter declarative memories against interference; SWS helps to actively struggle against interference.
DISCLOSURE STATEMENT
This was not an industry supported study. This study was supported by a FNRS (Belgian National Fund of Scientific Research - Fonds National Belge de la Recherche Scientifique) grant CC 1.5.184.10F. GD is a FNRS Research Fellow. RL is supported by a «Brains-Back-to-Brussels» grant, funded by the « Institut d'encouragement de la Recherche Scientifique et de l'Innovation de Bruxelles » (IRSIB), Brussels Capital-Region (Région de Bruxelles-Capitale). The authors have indicated no financial conflicts of interest.
ABBREVIATIONS
- CVLT
California Verbal Learning Task
- EEG
electroencephalogram
- EMG
electromyography
- EOG
electrooculograms
- IP
word pairs from A list subjected to interference
- KSS
Karolinska Sleepiness Scale
- N1
sleep stage 1
- N2
sleep stage 2
- N3
sleep stage 3
- NIP
word pairs from list A not subjected to interference
- NREM
non-rapid eye movement
- PSG
polysomnography
- PVT
psychomotor vigilance task
- REM
rapid eye movement
- RTs
reaction times
- SD
standard deviation
- SOREMS
REM at sleep onset
- SPT
sleep period time
- SWS
slow wave sleep
- TIB
time in bed
REFERENCES
- 1.Scrima L. Isolated REM sleep facilitates recall of complex associative information. Psychophysiology. 1982;19:252–9. doi: 10.1111/j.1469-8986.1982.tb02556.x. [DOI] [PubMed] [Google Scholar]
- 2.Born J, Rasch B, Gais S. Sleep to remember. Neuroscientist. 2006;12:410–24. doi: 10.1177/1073858406292647. [DOI] [PubMed] [Google Scholar]
- 3.Maquet P, Smith C, Stickgold R, editors. Sleep and brain plasticity. Oxford: Oxford University Press; 2003. [Google Scholar]
- 4.Peigneux P, Laureys S, Delbeuck X, Maquet P. Sleeping brain, learning brain. The role of sleep for memory systems. Neuroreport. 2001;12:A111–24. doi: 10.1097/00001756-200112210-00001. [DOI] [PubMed] [Google Scholar]
- 5.Rauchs G, Desgranges B, Foret J, Eustache F. The relationships between memory systems and sleep stages. J Sleep Res. 2005;14:123–40. doi: 10.1111/j.1365-2869.2005.00450.x. [DOI] [PubMed] [Google Scholar]
- 6.Stickgold R. Sleep-dependent memory consolidation. Nature. 2005;437:1272–8. doi: 10.1038/nature04286. [DOI] [PubMed] [Google Scholar]
- 7.Walker MP, Stickgold R. Sleep-dependent learning and memory consolidation. Neuron. 2004;44:121–33. doi: 10.1016/j.neuron.2004.08.031. [DOI] [PubMed] [Google Scholar]
- 8.Gais S, Lucas B, Born J. Sleep after learning aids memory recall. Learn Mem. 2006;13:259–62. doi: 10.1101/lm.132106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Schmidt C, Peigneux P, Muto V, et al. Encoding difficulty promotes postlearning changes in sleep spindle activity during napping. J Neurosci. 2006;26:8976–82. doi: 10.1523/JNEUROSCI.2464-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hu P, Stylos-Allan M, Walker MP. Sleep facilitates consolidation of emotional declarative memory. Psychol Sci. 2006;17:891–8. doi: 10.1111/j.1467-9280.2006.01799.x. [DOI] [PubMed] [Google Scholar]
- 11.Sterpenich V, Albouy G, Boly M, et al. Sleep-related hippocampo-cortical interplay during emotional memory recollection. PLoS Biol. 2007;5:e282. doi: 10.1371/journal.pbio.0050282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Orban P, Rauchs G, Balteau E, et al. Sleep after spatial learning promotes covert reorganization of brain activity. Proc Natl Acad Sci U S A. 2006;103:7124–9. doi: 10.1073/pnas.0510198103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fogel SM, Smith CT, Cote KA. Dissociable learning-dependent changes in REM and non-REM sleep in declarative and procedural memory systems. Behav Brain Res. 2007;180:48–61. doi: 10.1016/j.bbr.2007.02.037. [DOI] [PubMed] [Google Scholar]
- 14.Peigneux P, Schmitz R, Willems S. Cerebral asymmetries in sleep-dependent processes of memory consolidation. Learn Mem. 2007;14:400–6. doi: 10.1101/lm.551207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Squire LR. Memory systems of the brain: a brief history and current perspective. Neurobiol Learn Mem. 2004;82:171–7. doi: 10.1016/j.nlm.2004.06.005. [DOI] [PubMed] [Google Scholar]
- 16.Tulving E. Organization of memory: Quo vadis? In: Gazzaniga M, editor. The cognitive neurosciences. Cambridge: MA: MIT Press; 1995. pp. 839–47. [Google Scholar]
- 17.Iber C, Ancoli-Israel S, Chesson JA, Quan S. Westchester, IL: American Academy of Sleep Medicine; 2007. The AASM manual for the scoring of sleep and associated events: rules, terminology and technical specifications. [Google Scholar]
- 18.Billiard M, Dauvilliers Y. Les troubles du sommeil. Paris: Elsevier / Masson; 2011. [Google Scholar]
- 19.Plihal W, Born J. Effects of early and late nocturnal sleep on declarative and procedural memory. J Cogn Neurosci. 1997;9:534–47. doi: 10.1162/jocn.1997.9.4.534. [DOI] [PubMed] [Google Scholar]
- 20.Plihal W, Born J. Effects of early and late nocturnal sleep on priming and spatial memory. Psychophysiology. 1999;36:571–82. [PubMed] [Google Scholar]
- 21.Smith C. Sleep states and memory processes. Behav Brain Res. 1995;69:137–45. doi: 10.1016/0166-4328(95)00024-n. [DOI] [PubMed] [Google Scholar]
- 22.Ficca G, Lombardo P, Rossi L, Salzarulo P. Morning recall of verbal material depends on prior sleep organization. Behav Brain Res. 2000;112:159–63. doi: 10.1016/s0166-4328(00)00177-7. [DOI] [PubMed] [Google Scholar]
- 23.Gais S, Plihal W, Wagner U, Born J. Early sleep triggers memory for early visual discrimination skills. Nat Neurosci. 2000;3:1335–9. doi: 10.1038/81881. [DOI] [PubMed] [Google Scholar]
- 24.Giuditta A, Ambrosini MV, Montagnese P, et al. The sequential hypothesis of the function of sleep. Behav Brain Res. 1995;69:157–66. doi: 10.1016/0166-4328(95)00012-i. [DOI] [PubMed] [Google Scholar]
- 25.Stickgold R, Whidbee D, Schirmer B, Patel V, Hobson JA. Visual discrimination task improvement: A multi-step process occurring during sleep. J Cogn Neurosci. 2000;12:246–54. doi: 10.1162/089892900562075. [DOI] [PubMed] [Google Scholar]
- 26.Jenkins JG, Dallenbach KM. Obliviscence during sleep and waking. Am J Psychol. 1924;35:605–12. [Google Scholar]
- 27.Ellenbogen JM, Hulbert JC, Stickgold R, Dinges DF, Thompson-Schill SL. Interfering with theories of sleep and memory: sleep, declarative memory, and associative interference. Curr Biol. 2006;16:1290–4. doi: 10.1016/j.cub.2006.05.024. [DOI] [PubMed] [Google Scholar]
- 28.Ellenbogen JM, Hulbert JC, Jiang Y, Stickgold R. The sleeping brain's influence on verbal memory: boosting resistance to interference. PLoS ONE. 2009;4:e4117. doi: 10.1371/journal.pone.0004117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Bassetti C, Aldrich M. Narcolepsy. Neurol Clin. 1996;14:545–71. doi: 10.1016/s0733-8619(05)70273-5. [DOI] [PubMed] [Google Scholar]
- 30.Bes FW, Jobert M, Müller LC, Schulz H. The diurnal distribution of sleep propensity: experimental data about the interaction of the propensities for slow-wave sleep and REM sleep. J Sleep Res. 1996;5:90–8. doi: 10.1046/j.1365-2869.1996.00020.x. [DOI] [PubMed] [Google Scholar]
- 31.Czeisler CA, Zimmerman JC, Ronda JM, Moore-Ede MC, Weitzman ED. Timing of REM sleep is coupled to the circadian rhythm of body temperature in man. Sleep. 1980;2:329–46. [PubMed] [Google Scholar]
- 32.Dijk DJ, Czeisler CA. Contribution of the circadian pacemaker and the sleep homeostat to sleep propensity, sleep structure, electroencephalographic slow waves, and sleep spindle activity in humans. J Neurosci. 1995;15:3526–38. doi: 10.1523/JNEUROSCI.15-05-03526.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Endo S, Kobayashi T, Yamamoto T, Fukuda H, Sasaki M, Ohta T. Persistence of the circadian rhythm of REM sleep: a variety of experimental manipulations of the sleep-wake cycle. Sleep. 1981;4:319–28. doi: 10.1093/sleep/4.3.319. [DOI] [PubMed] [Google Scholar]
- 34.Webb W, Agnew H, Sternthal J, Sternthal H. Sleep during the early morning. Psychon Sci. 1966;6:277–8. [Google Scholar]
- 35.Dijk DJ, Brunner DP, Borbély AA. EEG power density during recovery sleep in the morning. Electroencephalogr Clin Neurophysiol. 1991;78:203–14. doi: 10.1016/0013-4694(91)90034-2. [DOI] [PubMed] [Google Scholar]
- 36.Werth E, Cote KA, Gallmann E, Borbély AA, Achermann P. Selective REM sleep deprivation during daytime I. Time course of interventions and recovery sleep. Am J Physiol RegulI Integr Comp Physiol. 2002;283:R521–6. doi: 10.1152/ajpregu.00462.2001. [DOI] [PubMed] [Google Scholar]
- 37.Tinguely G, Huber R, Borbély AA, Achermann P. Non-rapid eye movement sleep with low muscle tone as a marker of rapid eye movement sleep regulation. BMC Neurosci. 2006;7:2. doi: 10.1186/1471-2202-7-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ellis BW, Johns MW, Lancaster R, Raptopoulos P, Angelopoulos N, Priest RG. The St. Mary's Hospital sleep questionnaire: a study of reliability. Sleep. 1981;4:93–7. doi: 10.1093/sleep/4.1.93. [DOI] [PubMed] [Google Scholar]
- 39.Buysse DJ, Reynolds CF, 3rd, Monk TH, Berman SR, Kupfer DJ. The Pittsburgh Sleep Quality Index: a new instrument for psychiatric practice and research. Psychiatry Res. 1989;28:193–213. doi: 10.1016/0165-1781(89)90047-4. [DOI] [PubMed] [Google Scholar]
- 40.Krupp LB, La Rocca NG, Muir-Nash J, Steinberg AD. The fatigue severity scale. Application to patients with multiple sclerosis and systemic lupus erythematosus. Arch Neurol. 1989;46:1121–3. doi: 10.1001/archneur.1989.00520460115022. [DOI] [PubMed] [Google Scholar]
- 41.Beck AT, Steer RA, Garbin MG. Psychometric properties of Beck Depression Inventory: twenty-five years of evaluation. Clin Psychol. 1988;8:77–100. [Google Scholar]
- 42.Lecrubier Y, Weiller E, Hergueta T, Amorim P, Bonora L, Lépine J. Paris, France: INSERM; 1998. Mini International Neuropsychiatric Interview French Version 5.0. 0. [Google Scholar]
- 43.Horne JA, Ostberg O. A self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms. Int J Chronobiol. 1976;4:97–110. [PubMed] [Google Scholar]
- 44.Deltour JJ. Braine le Château, Belgique: Application des Techniques Modernes; 1993. Echelle de vocabulaire Mill Hill de J. C. Raven: Adaptation française et normes comparées du Mill Hill et du Standard Progressive Matrices (PM38). Manuel et Annexes. [Google Scholar]
- 45.Delis DC, Kramer JH, Kaplan E, Ober BA, editors. The California Verbal Learning Test. New York: The Psychological Corporation; 1987. [Google Scholar]
- 46.Content A, Mousty P, Radeau M. BRULEX: Une base de données lexicales informatisée pour le Français écrit et parlé. L'Année Psychologique. 1990;90:551–66. [Google Scholar]
- 47.Desrochers A, Bergeron M. Valeurs de fréquence subjective et d'imagerie pour un échantillon de 1916 substantifs de la langue française. Revue canadienne de psychologie expérimentale. 2000;54:274–325. [PubMed] [Google Scholar]
- 48.Dinges DI, Powell JW. Microcomputer analysis of performance on a portable, simple visual RT task sustained operations. Behav Res Meth Instrum Comput. 1985;17:652–5. [Google Scholar]
- 49.Akerstedt T, Gillberg M. Subjective and objective sleepiness in the active individual. Int J Neurosci. 1990;52:29–37. doi: 10.3109/00207459008994241. [DOI] [PubMed] [Google Scholar]
- 50.Anderer P, Gruber G, Parapatics S, et al. An E-health solution for automatic sleep classification according to Rechtschaffen and Kales: validation study of the Somnolyzer 24x7 utilizing the Siesta database. Neuropsychobiology. 2005;51:115–33. doi: 10.1159/000085205. [DOI] [PubMed] [Google Scholar]
- 51.Anderer P, Saletu B, Saletu-Zyhlarz GM, et al. Recent advances in the electrophysiological evaluation of sleep. In: Drinkenburg W, Ruigt G, Jobert M, editors. Essentials and applications of EEG research in preclinical and clinical pharmacology. Berlin: Unipublish Verlag für Studium & Praxis OHG; 2004. pp. 307–39. [Google Scholar]
- 52.Alger SE, Lau H, Fishbein W. Slow wave sleep during a daytime nap is necessary for protection from subsequent interference and long-term retention. Neurobiol Learn Mem. 2012;98:188–96. doi: 10.1016/j.nlm.2012.06.003. [DOI] [PubMed] [Google Scholar]
- 53.Euston DR, Tatsuno M, McNaughton BL. Fast-forward playback of recent memory sequences in prefrontal cortex during sleep. Science. 2007;318:1147–50. doi: 10.1126/science.1148979. [DOI] [PubMed] [Google Scholar]
- 54.Ji D, Wilson MA. Coordinated memory replay in the visual cortex and hippocampus during sleep. Nature Neurosci. 2007;10:100–7. doi: 10.1038/nn1825. [DOI] [PubMed] [Google Scholar]
- 55.Wilson MA, McNaughton BL. Reactivation of hippocampal ensemble memories during sleep. Science. 1994;265:676–9. doi: 10.1126/science.8036517. [DOI] [PubMed] [Google Scholar]
- 56.Marshall L, Helgadottir H, Molle M, Born J. Boosting slow oscillations during sleep potentiates memory. Nature. 2006;444:610–3. doi: 10.1038/nature05278. [DOI] [PubMed] [Google Scholar]
- 57.Molle M, Marshall L, Gais S, Born J. Learning increases human electroencephalographic coherence during subsequent slow sleep oscillations. Proc Natl Acad Sci U S A. 2004;101:13963–8. doi: 10.1073/pnas.0402820101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Peigneux P, Laureys S, Fuchs S, et al. Are spatial memories strengthened in the human hippocampus during slow wave sleep? Neuron. 2004;44:535–45. doi: 10.1016/j.neuron.2004.10.007. [DOI] [PubMed] [Google Scholar]
- 59.Rasch B, Buchel C, Gais S, Born J. Odor cues during slow-wave sleep prompt declarative memory consolidation. Science. 2007;315:1426–9. doi: 10.1126/science.1138581. [DOI] [PubMed] [Google Scholar]
- 60.Rudoy JD, Voss JL, Westerberg CE, Paller KA. Strengthening individual memories by reactivating them during sleep. Science. 2009;326:1079. doi: 10.1126/science.1179013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Diekelmann S, Born J. The memory function of sleep. Nature. 2010;11:114–26. doi: 10.1038/nrn2762. [DOI] [PubMed] [Google Scholar]
- 62.Rasch B, Born J. Maintaining memories by reactivation. Curr Opin Neurobiol. 2007;17:698–703. doi: 10.1016/j.conb.2007.11.007. [DOI] [PubMed] [Google Scholar]
- 63.Buzsaki G. Memory consolidation during sleep: a neurophysiological perspective. J Sleep Res. 1998;7:17–23. doi: 10.1046/j.1365-2869.7.s1.3.x. [DOI] [PubMed] [Google Scholar]
- 64.Gais S, Albouy G, Boly M, et al. Sleep transforms the cerebral trace of declarative memories. Proc Natl Acad Sci U S A. 2007;104:18778–83. doi: 10.1073/pnas.0705454104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.McGaugh JL. Memory-a century of consolidation. Science. 2000;287:248–51. doi: 10.1126/science.287.5451.248. [DOI] [PubMed] [Google Scholar]
- 66.Tse D, Langston RF, Kakeyama M, et al. Schemas and memory consolidation. Science. 2007;316:76–82. doi: 10.1126/science.1135935. [DOI] [PubMed] [Google Scholar]
- 67.Diekelmann S, Buchel C, Born J, Rasch B. Labile or stable: opposing consequences for memory when reactivated during waking and sleep. Nature Neurosci. 2011;14:381–6. doi: 10.1038/nn.2744. [DOI] [PubMed] [Google Scholar]
- 68.Dudai Y, Eisenberg M. Rites of passage of the engram: reconsolidation and the lingering consolidation hypothesis. Neuron. 2004;44:93–100. doi: 10.1016/j.neuron.2004.09.003. [DOI] [PubMed] [Google Scholar]
- 69.O'Reilly RC, Rudy JW. Conjunctive representations in learning and memory: principles of cortical and hippocampal function. Psychol Rev. 2001;108:311–45. doi: 10.1037/0033-295x.108.2.311. [DOI] [PubMed] [Google Scholar]
- 70.O'Reilly RC, McClelland JL. Hippocampal conjunctive encoding, storage, and recall: avoiding a trade-off. Hippocampus. 1994;4:661–82. doi: 10.1002/hipo.450040605. [DOI] [PubMed] [Google Scholar]
- 71.Bakker A, Kirwan CB, Miller M, Stark CEL. Pattern separation in the human hippocampal CA3 and dentate gyrus. Science. 2008;319:1640–2. doi: 10.1126/science.1152882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.McClelland JL, Goddard NH. Considerations arising from a complementary learning systems perspective on hippocampus and neocortex. Hippocampus. 1996;6:654–65. doi: 10.1002/(SICI)1098-1063(1996)6:6<654::AID-HIPO8>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- 73.Tononi G, Cirelli C. Sleep function and synaptic homeostasis. Sleep Med Rev. 2006;10:49–62. doi: 10.1016/j.smrv.2005.05.002. [DOI] [PubMed] [Google Scholar]
- 74.Genzel L, Dresler M, Wehrle R, Grozinger M, Steiger A. Slow wave sleep and REM sleep awakenings do not affect sleep dependent memory consolidation. Sleep. 2009;32:302–10. doi: 10.1093/sleep/32.3.302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Tucker MA, Fishbein W. Enhancement of declarative memory performance following a daytime nap is contingent on strength of initial task acquisition. Sleep. 2008;31:197–203. doi: 10.1093/sleep/31.2.197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Tucker MA, Hirota Y, Wamsley EJ, Lau H, Chaklader A, Fishbein W. A daytime nap containing solely non-REM sleep enhances declarative but not procedural memory. Neurobiol Learn Mem. 2006;86:241–7. doi: 10.1016/j.nlm.2006.03.005. [DOI] [PubMed] [Google Scholar]
- 77.Diekelmann S, Biggel S, Rasch B, Born J. Offline consolidation of memory varies with time in slow wave sleep and can be accelerated by cuing memory reactivations. Neurobiol Learn Mem. 2012;98:103–11. doi: 10.1016/j.nlm.2012.07.002. [DOI] [PubMed] [Google Scholar]
- 78.Steriade M, Timofeev I. Neuronal plasticity in thalamocortical networks during sleep and waking oscillations. Neuron. 2003;37:563–76. doi: 10.1016/s0896-6273(03)00065-5. [DOI] [PubMed] [Google Scholar]
- 79.Ayoub A, Molle M, Preissl H, Born J. Grouping of MEG gamma oscillations by EEG sleep spindles. NeuroImage. 2012;59:1491–500. doi: 10.1016/j.neuroimage.2011.08.023. [DOI] [PubMed] [Google Scholar]
- 80.Huber R, Ghilardi MF, Massimini M, Tononi G. Local sleep and learning. Nature. 2004;430:78–81. doi: 10.1038/nature02663. [DOI] [PubMed] [Google Scholar]
- 81.Mölle M, Yeshenko O, Marshall L, Sara SJ, Born J. Hippocampal sharp wave-ripples linked to slow oscillations in rat slow-wave sleep. J Neurophysiol. 2006;96:62–70. doi: 10.1152/jn.00014.2006. [DOI] [PubMed] [Google Scholar]
- 82.Mölle M, Eschenko O, Gais S, Sara SJ, Born J. The influence of learning on sleep slow oscillations and associated spindles and ripples in humans and rats. Eur J Neurosci. 2009;29:1071–81. doi: 10.1111/j.1460-9568.2009.06654.x. [DOI] [PubMed] [Google Scholar]
- 83.Schabus M, Gruber G, Parapatics S, et al. Sleep spindles and their significance for declarative memory consolidation. Sleep. 2004;27:1479–85. doi: 10.1093/sleep/27.7.1479. [DOI] [PubMed] [Google Scholar]
- 84.Schabus M, Hodlmoser K, Gruber G, et al. Sleep spindle-related activity in the human EEG and its relation to general cognitive and learning abilities. Eur J Neurosci. 2006;23:1738–46. doi: 10.1111/j.1460-9568.2006.04694.x. [DOI] [PubMed] [Google Scholar]