

Abstract
How neural circuits associated with sexually dimorphic organs are differentially assembled during development is unclear. Here, we report a sexually dimorphic pattern of mouse mammary gland sensory innervation and the mechanism of its formation. Brain-derived neurotrophic factor (BDNF), emanating from mammary mesenchyme and signaling through its receptor TrkB on sensory axons, is required for establishing mammary gland sensory innervation of both sexes at early developmental stages. Subsequently, in males, androgens promote mammary mesenchymal expression of a truncated form of TrkB, which prevents BDNF-TrkB signaling in sensory axons and leads to a rapid loss of mammary gland innervation independent of neuronal apoptosis. Thus, sex hormone regulation of a neurotrophic factor signal directs sexually dimorphic axonal growth and maintenance, resulting in generation of a sex-specific neural circuit.
Sexually dimorphic body structures require differentially assembled neural circuits to control gender-specific physiological functions and behaviors. One sexually dimorphic organ is the mammary gland, the milk-producing apocrine gland of female mammals. In mice, the initial stages of mammary gland development are virtually identical in males and females, until embryonic day 13 (E13). Then, in response to androgens secreted by the male gonads, the male glands begin to regress, such that sexual dimorphism is first observable at E14 (1, 2). By birth, each of the 10 mammary glands of the female mouse contains a clearly defined nipple and an elaborate ductal tree that is densely innervated by sensory neurons (fig. S1), whereas the male glands are barely detectable. To investigate how development of neuronal innervation coordinates with sexually dimorphic mammary gland organogenesis, we performed immunostaining using an antibody against the neuron-specific class III β-tubulin (Tuj1) to visualize neuronal fibers associated with rudimentary mammary glands from E11 to E14. In both males and females, Tuj1+ fibers innervate the mammary rudiments shortly after their appearance, and the number of Tuj1+ fibers associated with mammary rudiments is comparable for the two sexes until early E13 (Fig. 1, A to D and G, and figs. S2 and S3). These Tuj1+ fibers associated with mammary rudiments at this stage are from sensory neurons, most likely emanating from dorsal root ganglia (DRGs) (fig. S4). The number of Tuj1+ fibers innervating female mammary rudiments dramatically increases from late E12 to late E13, whereas fibers innervating male rudiments increase until early E13, but then they are rapidly lost during the subsequent 8 hours. As a result, a sexually dimorphic pattern of mammary gland sensory innervation is generated by late E13, before male gland regression (Fig. 1, E to G, and fig. S3). As sexually dimorphic development of neuronal populations in both the central nervous system and the peripheral nervous system has been linked to steroid hormone–regulated apoptotic cell death (3–7), we asked whether male-specific axonal loss from mammary rudiments is the result of apoptosis or, alternatively, axon pruning (8). In Bax−/− embryos, in which apoptosis in sensory ganglia is eliminated (9), mammary gland innervation at late E13 exhibits a sexually dimorphic pattern, which is quantitatively indistinguishable from that seen in wild-type embryos (fig. S3). Thus, formation of sexually dimorphic mammary gland innervation is not due to apoptotic loss of male sensory neurons; rather, it is a result of sensory axon pruning from the male gland beginning at early E13.
Fig. 1.

Androgen-dependent, male-specific axon pruning leads to sexually dimorphic mammary gland innervation before male gland regression. (A to F) Tuj1 (yellow) and TO-PRO-3 (blue, nucleic acid stain) staining of mammary gland sections of embryos at different stages. This staining was used in all subsequent mammary gland innervation analyses unless otherwise specified. (G) Quantification of mammary gland innervation. (H to O) Mammary gland innervation of embryos treated with vehicle, flutamide, or testosterone propionate (8 hours). The vehicle group shown in (H) and (I) is the control for flutamide treatments; controls for the testosterone treatments were similar (not shown). Each experiment was done at least three times. (N) and (O) are quantifications of the data shown in (H) to (M). All statistical analyses shown in this figure and all subsequent figures (except Fig. 2, E and F) were done using two-way ANOVA with a Bonferroni post hoc test. Shown are the means ± SEM, n ≥ 5 embryos for each bar, *P < 0.05; **P < 0.01; ***P < 0.001; n.s., not significant. Scale bar: 50 μm.
We next analyzed mammary gland innervation in embryos exposed to the androgen receptor antagonist flutamide, testosterone propionate, or the nonaromatizable androgen dihydrotestosterone (DHT), as well as in androgen-insensitive (Tfm) mice (10). It is remarkable that, in flutamide-treated embryos, sensory fibers projecting to the male mammary rudiments fail to be pruned, and quantification of fiber density shows no difference between the two sexes at late E13 (Fig. 1, H to K, and N). Similarly, the amount of mammary gland innervation in Tfm male embryos is comparable to both flutamide-treated male embryos and wild-type female embryos at late E13 (fig. S5). Conversely, in both testosterone- and DHT-treated embryos, sensory fibers projecting to female mammary rudiments are lost within 8 hours (Fig. 1, L, M, and O, and fig S5). Thus, androgen receptor activation is both necessary and sufficient for pruning of sensory axons and, thus, generation of the sexually dimorphic pattern of mammary gland sensory innervation before male gland regression.
The sexually dimorphic pattern of mammary gland innervation results from extension of new sensory fibers into female glands and a coincident pruning of fibers associated with male glands. This suggests that signaling pathways essential for promoting and maintaining axonal projections to female glands are either absent or disrupted in E13 males, presumably because of androgen receptor activation. To identify signals that mediate development of mammary gland sensory innervation in the female, we assessed the expression patterns of the four neurotrophins at E13 because neurotrophins play key roles in primary sensory neuron development (11, 12). Brain-derived neurotrophic factor (BDNF) emerged as a candidate because it is robustly expressed in the developing mammary gland at this stage, whereas the other neurotrophins are not (Fig. 2A and figs. S6 and S7). Staining on tissue sections showed that BDNF is expressed in mammary mesenchymal cells, not in mammary epithelial cells (Fig. 2A, inset). To determine whether the BDNF receptor TrkB is expressed in sensory neurons that innervate the female mammary rudiments, we analyzed TrkBGFP knock-in mice (13) and found that virtually all Tuj1+ fibers projecting to the female mammary rudiments express TrkB at E13 (Fig. 2B). Experiments using a TrkB antibody that recognizes the extracellular domain of TrkB (TrkBECD antibody) confirmed that TrkB is present on most or all axon terminals surrounding female glands at E13 (Fig. 3C). Furthermore, we generated a TrkBCreERT2 knock-in mouse line and used it to permanently label sensory neurons that express TrkB at E13. Postnatally, the labeled fibers innervate the ductal tree structure, but not the nipple, of female mammary glands (fig. S8).
Fig. 2.

BDNF-TrkB signaling is required for sensory innervation of the female mammary gland at E13. (A) Whole-mount X-gal staining of a late E13 female BDNFLacZ/+ embryo (21). (Inset) Cross section of one mammary gland. Arrows: rudimentary mammary glands (five on each side of the body, glands nos. 1 and 5 are located behind the limbs). (B and B’) Costaining of GFP, Tuj1, and TO-PRO-3 of a late E13 female TrkBGFP/+ mammary gland. (C to F) Mammary gland innervation of late E13 female wild-type (WT), BDNFlacZ/LacZ (null), TrkBf/+, and TrkBf/f; AvilCre/+embryos. [The AvilCre/+ mouse is a sensory neuron–specific Cre driver mouse line (22). Statistical analyses in (E) and (F) were done using Student’s t test.] (G to K) Mammary gland innervation of late E13 wild-type and TrkBF616A/F616A female embryos that had been exposed to either vehicle or 1NMPP1 for 5 hours. For each bar, n ≥ 3 embryos in (E), (F), and (K). Scale bar: 50 μm.
Fig. 3.
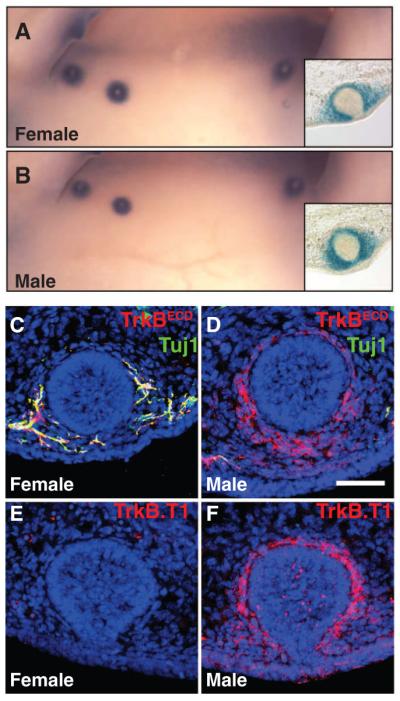
The truncated form of TrkB is expressed in E13 male mammary mesenchymal cells. (A and B) X-gal staining of late E13 female and male BDNFLacZ/+ embryos, respectively. (C to F) Mammary gland sections of late E13 male and female embryos were stained with TO-PRO-3 (blue) and, in addition, (C and D) with TrkBECD and Tuj1 antibodies or (E and F) with a truncated form–specific TrkB antibody (TrkB.T1). Staining observed in the female section in (E) is due to nonspecific antibody binding to blood cells. All experiments were done at least three times. Scale bar: 50 μm.
To ask whether BDNF-TrkB signaling is necessary for establishing sensory innervation of the female mammary gland, we assessed mammary gland innervation in mouse lines harboring targeted mutations of either Bdnf or TrkB. Female embryos with a homozygous null mutation in Bdnf exhibited very few sensory fibers associated with the mammary rudiments at E13 (Fig. 2, C to E). Moreover, both TrkB null and TrkB conditional mutant embryos in which TrkB is ablated exclusively in primary sensory neurons showed a dramatic reduction in female mammary gland innervation at E13 (Fig. 2F and fig. S9). To determine whether BDNF-TrkB signaling is required at E13 for maintaining female mammary gland sensory innervation, we used a chemical-genetic strategy that enables precise temporal inhibition of TrkB signaling in vivo. TrkBF616A knock-in mice harbor a single–amino acid substitution in the TrkB adenosine triphosphate binding pocket that allows specific and potent inhibition of endogenous TrkB kinase signaling by the small-molecule inhibitor 1NMPP1 (14). Five hours of 1NMPP1 exposure at E13 resulted in a dramatic reduction in mammary gland innervation of TrkBF616A/F616A female embryos compared with that of vehicle-treated TrkBF616A/F616A females, whereas 1NMPP1 had no effect on wild-type females (Fig. 2, G to K). Thus, BDNF-TrkB signaling is required in sensory neurons for establishment and maintenance of mammary gland innervation, and disrupting BDNF-TrkB signaling in the female at E13 leads to rapid axonal pruning and a male-type pattern of mammary gland sensory innervation.
We next asked whether BDNF-TrkB signaling is absent or disrupted in E13 male glands and thus would account for the androgen-dependent axon pruning and the sexually dimorphic pattern of mammary gland sensory innervation. The androgen receptor is expressed in both the male and female mammary mesenchyme but not in DRG sensory neurons at E13 (figs. S10 and S14), which suggests that the site of androgen action is the rudimentary mammary gland itself. We next tested whether BDNF expression in mammary mesenchymal cells differs in males and females. This appears not to be the case, as X-gal staining of BDNFLacZ/+ embryos shows that BDNF expression in the male mammary mesenchyme is comparable to that of the female at late E13 (Fig. 3, A and B, and fig. S11). We next examined TrkB expression and, to our surprise, found that male mammary mesenchymal cells robustly express TrkB (Fig. 3D). This pattern of TrkB expression in the male mammary mesenchyme is not observed in female glands at the same stage and neither is it seen in the glands of either sex 1 day earlier, at E12 (Fig. 3C and fig. S12). These findings raise the possibility that BDNF is sequestered by TrkB on male mammary mesenchymal cells at E13, which prevents BDNF-TrkB signaling in sensory fibers associated with the male mammary rudiments and leads to pruning of fibers in the male and the sexually dimorphic mammary gland innervation pattern.
We tested this model by exploring the nature of TrkB expression in the male mammary mesenchyme, its androgen dependence, and the requirement of mesenchymal TrkB for generating the male pattern of mammary gland sensory innervation. Thus far, two isoforms of TrkB generated by alternative splicing have been described in mice: a full-length form, with the intracellular tyrosine kinase domain required for canonical receptor tyrosine kinase signaling and BDNF-dependent axonal growth (15, 16), and a truncated form, called TrkB. T1, which lacks the tyrosine kinase domain and, instead, has a unique, short, C-terminal intracellular domain (17). Both TrkB forms bind to BDNF and become internalized together with ligand (18, 19), which demonstrates their ability to sequester BDNF. Immunostaining using a TrkB.T1-specific antibody that recognizes the C-terminal intracellular domain of TrkB.T1 revealed that TrkB.T1 is robustly expressed in the male mammary mesenchyme at E13, whereas it is undetectable in the female gland (Fig. 3, E and F). In complementary experiments, in situ hybridizations using two different probes to distinguish between full-length TrkB and TrkB.T1 transcripts show that there is an undetectable amount of full-length TrkB mRNA in mammary mesenchymal cells of either sex (fig. S13). These findings indicate that sexually dimorphic expression of TrkB in mammary mesenchymal cells at E13 is the result of differential transcription of the TrkB gene and expression of the truncated form of TrkB exclusively in the male gland.
We next asked whether TrkB expression in male mammary mesenchymal cells is androgen-dependent. Double immunostaining for green fluorescent protein (GFP) and the androgen receptor on mammary gland sections of TrkBGFP/+ knock-in embryos showed that TrkB-expressing cells surrounding the male mammary rudiment are indeed a major subset of androgen-expressing mammary mesenchymal cells (fig. S14). TrkB expression in mammary mesenchymal cells, assessed by TrkBECD antibody labeling, is abolished in male embryos treated with flutamide and, conversely, it is dramatically induced in female embryos treated with testosterone (Fig. 4, A to F). Experiments that assessed GFP levels in mammary mesenchymal cells of TrkBGFP/+ embryos treated with flutamide are consistent with these TrkBECD antibody labeling experiments and indicate that androgens control TrkB expression through regulation of TrkB gene transcription in mammary mesenchymal cells (fig. S15). Moreover, the increase of TrkB expression in the female mammary mesenchyme after testosterone treatment is temporally coincident with the loss of sensory innervation of female glands (fig. S16). Thus, TrkB transcription in the mammary mesenchyme is androgen-dependent and inversely related to the amount of sensory fibers associated with the rudimentary gland.
Fig. 4.

Androgen-dependent mammary mesenchymal TrkB expression controls formation of the sexually dimorphic pattern of mammary gland innervation before male gland regression. (A to F) Mammary gland sections of late E13 male and female embryos exposed to vehicle, flutamide, or testosterone propionate (8 hours) were stained with TrkBECD (red) and Tuj1 (green) antibodies, and TO-PRO-3 (blue). The same staining is shown in all subsequent images in this figure. (G to K) Mammary mesenchymal TrkB expression and mammary gland innervation of wild-type and TrkB.T1 mutant male and female embryos. (H) A male mammary gland with relatively low mesenchymal TrkB expression. (I) A male mammary gland with relatively high mesenchymal TrkB expression. (L) Quantification of mammary gland innervation of wild-type and TrkB.T1 mutant embryos at late E13 (n ≥ 7 embryos for each bar). (M) The amount of mammary gland innervation of TrkB.T1 mutant male embryos is inversely correlated with residual mesenchymal TrkB expression level. Relative fiber density is fiber density of individual mammary glands divided by the average fiber density of the corresponding female mammary glands at the same position along the anterior-posterior axis at late E13. Each data point represents one mammary gland from a TrkB.T1 mutant male embryo (65 glands from eight mice).
To determine whether the truncated form of TrkB expressed in the male mammary mesenchyme at E13 plays an essential role in the sexually dimorphic patterning of mammary gland innervation, we analyzed mammary gland innervation in TrkB.T1 mutant mice in which the exon encoding the C-terminal residues unique to TrkB.T1 is missing (20). These mice do not express TrkB.T1, but they do express normal levels of full-length TrkB. Because TrkB.T1 is not expressed in DRG neurons at E13 (fig. S17), performing experiments using TrkB.T1 mutant mice allowed us to address the role of TrkB.T1 without directly affecting BDNF-TrkB signaling in DRG neurons themselves. We examined mammary mesenchymal TrkB expression in TrkB.T1 mutant male embryos by immunostaining using the TrkBECD antibody and quantifying immunofluorescence signals in the mammary mesenchyme (fig. S18). The specificity of the TrkBECD antibody was confirmed using late E13 male TrkBGFP/GFP (null) embryos (fig. S19). Our analysis revealed a significant reduction in, but not a complete loss of, TrkB expression in mammary mesenchyme of male TrkB.T1 mutant embryos at late E13, as compared with wild-type males (Fig. 4, G to I, and fig. S18). Although the residual TrkB protein found in TrkB.T1 mutant embryos lacks the TrkB.T1-specific C terminus, it is likely capable of associating with BDNF because it does contain the extracellular domain of TrkB (fig. S20). Quantification of mammary gland innervation revealed a significant increase in fiber density in TrkB.T1 mutant males compared with wild-type males at late E13, and no significant difference between TrkB.T1 mutant females and wild-type females (Fig. 4, G to L). Moreover, because we observed that the amount of residual TrkB varies between glands and embryos (Fig. 4, H and I), we assessed the relation between the amount of residual TrkB expression and relative fiber density for individual glands from several TrkB.T1 mutant males. This analysis revealed a strong inverse correlation between the amount of residual mesenchymal TrkB observed in individual mammary glands of TrkB.T1 mutant males and the number of sensory fibers associated with these glands (correlation coefficient: r = 0.6499, P < 0.0001) (Fig. 4M). We conclude that androgen-dependent expression of the truncated form of TrkB in male mammary mesenchyme is required for inhibition of BDNF-TrkB signaling in sensory fibers, axonal pruning, and generation of the male-type pattern of mammary gland sensory innervation at late E13 (fig. S21). Thus, sex hormone–dependent modulation of neurotrophic factor signaling is a mechanism that controls sexually dimorphic development of the mammalian nervous system.
Supplementary Material
Acknowledgments
We thank members of the Ginty laboratory for assistance and discussions throughout the course of this project, and A. Kolodkin and R. Kuruvilla for comments on the manuscript. This work was supported by NIH grants NS34814 (D.D.G.), EY014998 (K.R.J.), and DE019440 (F.W.). D.D.G. is an investigator of the Howard Hughes Medical Institute. BDNFlox mice are covered under U.S. patent no. 4,959,317, which is licensed to Bristol-Myer Squibb Company, with the right to grant sublicenses. The mice are available for noncommercial biomedical research under a Material Transfer Agreement with the University of Colorado.
Footnotes
Supplementary Materials www.sciencemag.org/cgi/content/full/338/6112/1357/DC1
Materials and Methods
Figs. S1 to S21
References (23–30)
References and Notes
- 1.Robinson GW. Nat. Rev. Genet. 2007;8:963. doi: 10.1038/nrg2227. [DOI] [PubMed] [Google Scholar]
- 2.Dunbar ME, et al. Development. 1999;126:3485. doi: 10.1242/dev.126.16.3485. [DOI] [PubMed] [Google Scholar]
- 3.Nordeen EJ, Nordeen KW, Sengelaub DR, Arnold AP. Science. 1985;229:671. doi: 10.1126/science.4023706. [DOI] [PubMed] [Google Scholar]
- 4.Wright LL, Smolen AJ. Int. J. Dev. Neurosci. 1987;5:305. doi: 10.1016/0736-5748(87)90005-0. [DOI] [PubMed] [Google Scholar]
- 5.Simerly RB. Annu. Rev. Neurosci. 2002;25:507. doi: 10.1146/annurev.neuro.25.112701.142745. [DOI] [PubMed] [Google Scholar]
- 6.Morris JA, Jordan CL, Breedlove SM. Nat. Neurosci. 2004;7:1034. doi: 10.1038/nn1325. [DOI] [PubMed] [Google Scholar]
- 7.Forger NG. Neuroscience. 2006;138:929. doi: 10.1016/j.neuroscience.2005.07.006. [DOI] [PubMed] [Google Scholar]
- 8.Low LK, Cheng HJ. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006;361:1531. doi: 10.1098/rstb.2006.1883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Patel TD, Jackman A, Rice FL, Kucera J, Snider WD. Neuron. 2000;25:345. doi: 10.1016/s0896-6273(00)80899-5. [DOI] [PubMed] [Google Scholar]
- 10.Lyon MF, Hawkes SG. Nature. 1970;227:1217. doi: 10.1038/2271217a0. [DOI] [PubMed] [Google Scholar]
- 11.Bibel M, Barde YA. Genes Dev. 2000;14:2919. doi: 10.1101/gad.841400. [DOI] [PubMed] [Google Scholar]
- 12.Huang EJ, Reichardt LF. Annu. Rev. Neurosci. 2001;24:677. doi: 10.1146/annurev.neuro.24.1.677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Li L, et al. Cell. 2011;147:1615. doi: 10.1016/j.cell.2011.11.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chen X, et al. Neuron. 2005;46:13. [Google Scholar]
- 15.Klein R, Parada LF, Coulier F, Barbacid M. EMBO J. 1989;8:3701. doi: 10.1002/j.1460-2075.1989.tb08545.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Atwal JK, Massie B, Miller FD, Kaplan DR. Neuron. 2000;27:265. doi: 10.1016/s0896-6273(00)00035-0. [DOI] [PubMed] [Google Scholar]
- 17.Klein R, Conway D, Parada LF, Barbacid M. Cell. 1990;61:647. doi: 10.1016/0092-8674(90)90476-u. [DOI] [PubMed] [Google Scholar]
- 18.Biffo S, Offenhäuser N, Carter BD, Barde YA. Development. 1995;121:2461. doi: 10.1242/dev.121.8.2461. [DOI] [PubMed] [Google Scholar]
- 19.Fryer RH, Kaplan DR, Kromer LF. Exp. Neurol. 1997;148:616. doi: 10.1006/exnr.1997.6699. [DOI] [PubMed] [Google Scholar]
- 20.Dorsey SG, et al. Neuron. 2006;51:21. doi: 10.1016/j.neuron.2006.06.009. [DOI] [PubMed] [Google Scholar]
- 21.Gorski JA, Zeiler SR, Tamowski S, Jones KR. J. Neurosci. 2003;23:6856. doi: 10.1523/JNEUROSCI.23-17-06856.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.da Silva S, et al. Proc. Natl. Acad. Sci. U.S.A. 2011;108:3395. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.