

Abstract
Humankind's ongoing battle with pest species spans millennia. Pests cause or carry disease, damage or consume food crops and other resources, and drive global environmental change. Conventional approaches to pest management usually involve lethal control, but such approaches are costly, of varying efficiency and often have ethical issues. Thus, pest management via control of reproductive output is increasingly considered an optimal solution. One of the most successful such ‘fertility control’ strategies developed to date is the sterile male technique (SMT), in which large numbers of sterile males are released into a population each generation. However, this approach is time-consuming, labour-intensive and costly. We use mathematical models to test a new twist on the SMT, using maternally inherited mitochondrial (mtDNA) mutations that affect male, but not female reproductive fitness. ‘Trojan females’ carrying such mutations, and their female descendants, produce ‘sterile-male’-equivalents under natural conditions over multiple generations. We find that the Trojan female technique (TFT) has the potential to be a novel humane approach for pest control. Single large releases and relatively few small repeat releases of Trojan females both provided effective and persistent control within relatively few generations. Although greatest efficacy was predicted for high-turnover species, the additive nature of multiple releases made the TFT applicable to the full range of life histories modelled. The extensive conservation of mtDNA among eukaryotes suggests this approach could have broad utility for pest control.
Keywords: biocontrol, fertility control, mtDNA, population viability, sterile male technique, mathematical model
1. Introduction
Pest species (organisms which may cause illness, or damage or consume food crops and other resources important to humans) are critical drivers of global environmental change [1,2]. Pest species are typically abundant, invasive and difficult to control, and can have major impacts on agricultural productivity and native biota worldwide [3–5]. The knock-on effects of such impacts often result in ecosystem-wide consequences for species interaction networks [6], community composition [7], and important ecosystem services such as nutrient cycling and carbon sequestration [8]. Pests are therefore often the target of costly eradication or control programmes [4,9].
Conventional approaches to controlling pests, be they plants, vertebrates or invertebrates, usually involve lethal control techniques, such as poisoning, trapping or shooting vertebrates, herbicide application for weeds, and surfactants for mosquitoes. However, sustained control using these techniques is often limited by cost and effectiveness at low population density, making eradication unlikely except in small geographically defined areas such as islands [10]. These constraints, in addition to adverse non-target effects on other species [9,11], concerns over environmental contamination and animal welfare (in particular for the control of vertebrate pests [12]), and the evolution of resistance with respect to toxin application [13,14], have all spurred a strong demand for more targeted, cost-effective and humane methods of managing pests [15,16].
Management of pest populations via control of reproductive output is generally considered one optimal solution to such issues [17–19]. Theoretically, these approaches are generally more effective at rapidly reducing population size in pest species with relatively high fecundity, low survivorship rates and short generation times [20]. However, the development of such ‘fertility control’ to date has been less than promising, with technologies either unable to reach the levels of efficacy required [21–23], or having undesirable non-target effects [24,25]. Arguably, the most successful of these approaches to date has been the sterile male technique (SMT) commonly applied to invertebrate pests [26,27], in which large numbers of sterile males (SMs) are released into a population each generation. Females mating with a sterile male produce few or no viable progeny. If released in sufficient numbers, then enough females mate with SMs to bring about a reduction in population size in the next or subsequent generations [28], and repeated release will ultimately result in a population being greatly reduced in abundance or even eradicated [29]. For example, use of the SMT has eradicated the parasitic screwworm fly from multiple countries, with an estimated US$796 million per annum saving to the USA alone [27]. However, the SMT has not been a long-term sustainable solution to pest control in all cases. In particular, under current approaches, very large numbers of SMs need to be produced and released year after year, which is a time-consuming, labour-intensive and costly process that may prohibit its use in poorer countries [30]. Here, we explore a new twist on the sterile male control paradigm, using mitochondrial (mtDNA) mutations to introduce ‘Trojan females (TFs)’ into wild populations where they will continuously produce ‘sterile male’ equivalents under natural conditions, across multiple generations.
While it is well established that mtDNA mutations can have dramatic consequences for individual fitness, causing degenerative diseases and cancer through extreme disruptions of the oxidative phosphorylation system (essential for aerobic respiration) [31–33], more subtle effects of mtDNA mutations on male fertility are only now being realized [34–43]. Most tissues can function well when mitochondrial capacity is reduced by as much as 80% [32,33,44,45]. However, the dependence of sperm vigour on mitochondrial capacity, together with the relatively small numbers of mtDNA in a sperm [46,47], means that even a modest reduction in mitochondrial capacity can impact male fertility substantially [35,38,41,43,44,48]. Owing to the asymmetry between egg and sperm dependence of mtDNA function, mtDNA mutations can potentially cause large reductions in male fitness while having little or no effect on female fitness; a view supported by empirical data from mice [38,41] and more recently flies [34,35,37,43]. The maternal inheritance of mtDNA will further exacerbate the fitness costs to males, because deleterious mutations that affect only males will not be subject to natural selection [44,49,50]. Indeed, theory predicts that regardless of the selective pressure in males, mtDNA mutations will be maintained at a mutation–selection equilibrium frequency determined solely by the relative fitness of a female bearing the mutation [49], except under rare situations where there may be high levels of positive-assortative mating or strong inclusive fitness costs to females [51–53]. Surprisingly, the opportunity that this fitness asymmetry presents for biological control of pest organisms has not previously been recognized, even though the predicted effects of such mtDNA mutations on the viability of small populations of endangered species are recognized [44,50]. The purposeful introduction of females carrying such mtDNA mutations into pest populations offers a novel humane approach to their management via fertility control—a concept we term the Trojan female technique (TFT).
In this paper, we use a modelling framework to conceptually explore the utility of the TFT for continuous, self-sustaining biological control of pest organisms. Model exploration and sensitivity analyses were conducted to address three fundamental questions: (i) what magnitude of population-scale impact could this technology theoretically have? (ii) Under what circumstances, and over what time scale, could such impacts occur? (iii) What general life-history characteristics might make pest species more or less amenable to population control using such technology? Answers to these questions will guide the future development of this approach as a novel control approach for managing pest populations.
2. Material and methods
(a). General two-sex growth model
We began with a simple and general two-sex model to describe the dynamics of male and female population densities (numbers/area) in a closed community with no spatial heterogeneity. This can be presented via
| 2.1 |
Here, r is the sex ratio at birth, b and d are density-dependent birth and death rates (per capita), respectively, and N is the total population size at each instant of time. Model (2.1) is a deterministic birth–death process with no immigration or emigration effects. P(M) is the female mating rate. In fact, P(M) represents the probability of male–female encounter in the closed population, irrespective of any spatial patterns. A shortage of mating encounters or a sustained disruption in the mating process can affect the reproductive rate of a population. Hence, from a modelling point of view, Allee effects pertinent to mate finding are usually embedded in the fertilization rate. Mate-finding Allee effects can give rise to strong demographic Allee effects, and therefore facilitate pest population reduction or eradication.
We also note from model (2.1) that P only depends on the male population size. This represents a polygynous mating system in which it is assumed that male individuals are not sperm-limited and can achieve arbitrarily high mating rates. Among a variety of functional forms available for the fertilization probability the rectangular hyperbolic form is the most common, mainly owing to its mathematical simplicity and convenience. The rectangular hyperbolic function for P(M) is given by
| 2.2 |
where A scales the strength of the mate-finding Allee effect or any other component Allee effect influencing reproduction.
(b). Generic mtDNA mutant population model
We extended the two-sex model (2.1) by considering a multi-state framework for individuals in the population, distinguishing between their mtDNA genetic make-up (wild-type versus mutant). The population dynamics of all four states are given by the following block of ordinary differential equations:
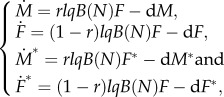 |
2.3 |
where M and F refer to the wild-type individuals, and M* and F*represent mtDNA mutants. A detailed parameter description is given in the next section.
Under the most favourable scenario, where mutant males are assumed to be fully sterile (as opposed to just suffering reduced fertility), we would expect a decline in female fertilization probability as the proportion of mutant males increases. This scenario is predicted by
| 2.4 |
(c). Model assumptions and simulation
Five key assumptions underlie our model simulations:
(i) Breeding is assumed to take place continuously throughout the year.
(ii) Mutant individuals are conceived when a mutant female (F*) mates with a wild-type male (M). Mutant individuals have the same survival probability as their wild-type counterparts. r is the sex ratio at birth and we assume 51% male at birth. l is the average birth rate per female.
(iii) Female fertilization probability is given by
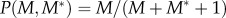 , meaning that in the absence of M*, there is a weak mate-finding Allee effect present in the population.
, meaning that in the absence of M*, there is a weak mate-finding Allee effect present in the population.(iv) The parameter q is an average (per month) maturation rate. We define the quantity 1/q as the generation time (months), being the time that it takes for a juvenile to reach adulthood and commence reproduction.
(v) B(N) is the probability of a successful conception and it depends directly on the proportion of females breeding as well as the mating probability, P(M,M*). The proportion of females breeding denoted by b(N) is calculated via
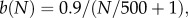 where N is the total population density per 100 ha. The overall B(N) is given by
where N is the total population density per 100 ha. The overall B(N) is given by 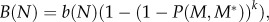 where k is the mating frequency per year (assumed equal to 1, i.e. monogamy, unless noted otherwise).
where k is the mating frequency per year (assumed equal to 1, i.e. monogamy, unless noted otherwise).
Simulations of the generic mtDNA mutant population model were conducted in MATLAB, using a monthly time-step and a generation time of 12 months unless otherwise noted. Management scenarios were explored across a range of maximum birth/death rate combinations, in which b was varied up to 10 per year and d was varied up to five per year. Each simulation began with initial values of 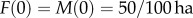 . We then let the computer program run until the time when the wild-type population level reached its stable level with a maximum carrying capacity of K1. Having the wild-type population stabilized around K1, we then released either mutant females (simulating the TFT) or mutant males (simulating the SMT for comparison). The number of mutants released is a fraction of the total population prior to the start of the next simulated year. After the intervention, we let the population stabilize around a new carrying capacity K2, and calculated the relative population decline by
. We then let the computer program run until the time when the wild-type population level reached its stable level with a maximum carrying capacity of K1. Having the wild-type population stabilized around K1, we then released either mutant females (simulating the TFT) or mutant males (simulating the SMT for comparison). The number of mutants released is a fraction of the total population prior to the start of the next simulated year. After the intervention, we let the population stabilize around a new carrying capacity K2, and calculated the relative population decline by 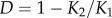 .
.
3. Results
The modelled biological control approach is based on either single or multiple releases into the pest population of TFs—adult females carrying mutant mtDNA (F*, causing male sterility) but which are themselves fully fertile—under the assumption that there is no mortality of released animals. Once established in the population, these females mate with local males and produce a continuous supply of mutant individuals over the subsequent generations so long as there are fertile wild-type males with which to mate.
Model simulations were first conducted to explore the relative population decline predicted from the release of TFs equivalent to 1 and 10% of the total standing population abundance (figure 1). Trojan female release at 1% of the standing population (figure 1a) achieved appreciable population declines (approx. 10%) over a large region of the parameter space explored, particularly in the high-birth- and high-death-rate region, with the effect being more sensitive to changing death rate than birth rate. TF releases at 10% achieved high levels of population suppression over a large area of the parameter space explored (figure 1b), and the demographic outcome with respect to varying birth and death rates was maintained (figure 1b).
Figure 1.

(a–c) Contour plots of relative population decline predicted for pests across birth/death rate parameter space (with a set generation time of 12 months), following single or repeated releases of TFs. All rates are yearly. The white region in (a–c) corresponds to parameter space where the death rate exceeds the birth rate. (a) Single release of 1% standing population. (b) Single release of 10% standing population. (c) Multiple releases of 1% per year for 10 consecutive years. (d) The relationship between the total declines achieved versus the number of successive years of releasing TFs at 1% per annum. Blue solid line: birth rate = 2 per year; death rate = 1 per year. Red dashed lines: birth rate = 8 per year; death rate decreases from left to right (5, 3, 1).
For logistical purposes, such as constraints in the breeding of mutant animals or an unwillingness to boost populations in the field by too much (resulting in unwanted consequences such as human annoyance by mosquitoes or vertebrate pests impacting native fauna), frequent low-frequency releases may be a more valid option for TFT application than large one-off releases. We thus explored how yearly low-frequency releases at 1% perform. Over 10 years, this approach greatly increased the effect predicted to be achieved by 1% releases, even markedly increasing the parameter space across which high levels of population suppression occurred relative to a one-off 10% release in year 1 (figure 1c). Furthermore, no matter the size of the yearly release, given a sufficient length of control application, gains always appeared to be additive over time in the models, until target species eradication was achieved (figure 1d).
To compare the TFT with the existing successful SMT, we simulated annual releases for both approaches until residual pest populations reached 1% (at a yearly birth rate per female of 8 and a yearly death rate per capita of 1—pest species life-history parameters amenable to SMT application). A release ratio of 170% (the predicted minimum effective release ratio for successful control using the SMT [27]) reduced residual populations to below 1% in just two releases for the SMT, and in just one release for the TFT. For the SMT, this is not realistic of the situation in the field, probably due to the non-spatial nature of our model. Rather, suppression to below 1% within a decade (a time scale realistic of the SMT [29]) was achieved in our model with release ratios of 65% for the SMT and 22% for the TFT (figure 2). While the simulated population rebounded upon cessation of release under the SMT, population suppression was maintained under the TFT owing to its persistent nature. As a result, continuation of release until eradication would not be required under the TFT; rather, population levels need only to be suppressed sufficiently for environmental or demographic stochasticity to lead to population extinction.
Figure 2.
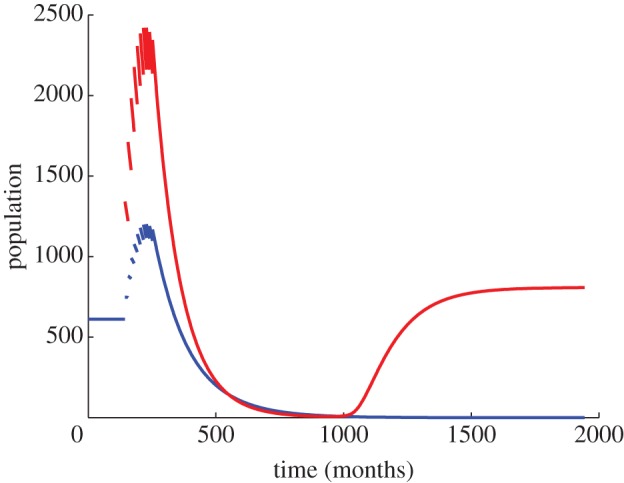
Comparison of the SMT (red) and the TFT (blue) at their minimum yearly release rates sufficient to suppress the modelled population to 1% of its initial level within 10 years (with birth rate = 8 per year and death rate = 1 per year). The comparable yearly release rates of SMs and TFs were 65% and 22% of the standing population, respectively.
To investigate how social systems in which females mate with multiple males might influence the predicted utility of the TFT, we selected four representative life histories from figure 1 and quantified how the level of population suppression achieved by 10 successive releases of TFs at 1% varied with increasing numbers of female mating partners (from 1 up to 10 different males each year). These simulations were carried out under the extreme assumption that a female mating with just a single wild-type male each year produces her full complement of offspring, regardless of how many mutant males she mates with, to illustrate the maximum likely effect of such mating systems. Our results show that while multiple mating reduces the predicted suppression effect of the TFT, substantial effects remain even if females have high rates of multiple mating (figure 3).
Figure 3.
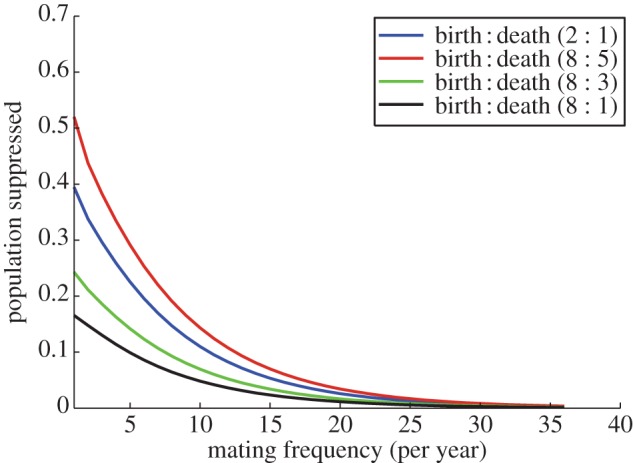
The effect of female multiple mating on the predicted efficiency of the TFT for 10 successive yearly releases of TFs at 1% of the standing population. Each line shows the population suppression achieved, for populations with different yearly per capita birth/death rates, as the number of mating partners a female has during the mating season increases.
Finally, to investigate how variation in target pest species generation time may also influence the utility of the TFT, the simulations presented in figure 1 were repeated for species with generation times of 3 months, 6 months and 2 years (see electronic supplementary material, figure S1). In general, the broad patterns predicted for a target species with a generation time of 12 months were maintained, with the level of population suppression showing a positive relationship with increasing generation time (all else being equal). This relationship is driven by the increase in control effort per generation with increasing generation time.
4. Discussion
Research has identified a situation in sexual organisms where mutations deleterious to male (but not female) fitness can accumulate in maternally inherited mtDNA without selection acting against it, imposing a male-specific genetic load [49]. Authors have previously discussed the consequences of this for endangered populations and their conservation [44,50]. Conversely, here we demonstrate the potential to use this apparent evolutionary loophole for a novel humane approach to pest population control—the TFT.
Our models demonstrate that mtDNA mutations that affect male, but not female reproductive fitness have potential as biological control agents for pests, having predicted population-scale impacts in relatively short time frames (figure 1d). We find that the TFT has the greatest efficacy when applied to species with high turnover rates (figure 1a–c), with only a slight influence of generation time (see electronic supplementary material, figure S1). This is in broad agreement with similar assessments for fertility control in general [20]. We also find that single releases of TFs can provide effective reductions in population sizes when such releases are relatively large (10% of the total population), but the effectiveness of single releases drops when the proportion of mutant females is lower (1%). However, multiple releases of smaller numbers of TFs (1% of the total population) can provide effective and persistent control within a relatively small number of generations. In addition, the additive nature of multiple TFT releases enables high levels of predicted suppression to be achieved even when single releases have limited impact owing to less optimal life-history parameters and/or social structures.
The predicted success of the TFT approach is further improved by the simultaneous release of both the mutant females (TFs) and the SMs that would be produced as a by-product of producing TFs for release (see electronic supplementary material, figure S2). While our main focus here is on the effects driven by the release solely of TFs, this modification would have two key advantages. First, the release of these SMs has an immediate effect on offspring production. Second, it avoids the time-consuming, costly and potentially ethically challenging issue of sorting through and discarding the unwanted sex; a significant shortcoming of the SMT approach. The TFT approach circumvents the need to sort through the sexes, as the release of both females and males will produce population control, albeit in different generations.
Other approaches to pest population management through fertility biocontrol frequently encounter two significant issues. The first is associated with the use of additional species, such as transgenic parasites, to vector the control agents of choice (e.g. genes for zona pellucida proteins, production of which stimulates an auto-immune response [54]). While such vectors could indeed lead to ‘transmissible’ biocontrol (i.e. population control achieved through self-disseminating agents [18]) the potential for non-target impacts may make them ethically unusable [24,25,55]. The second is associated with the non-permanent nature (at the population scale) of the control approach used. Even successful approaches, such as the SMT [26,27,29], require ongoing releases of control agents to maintain population regulatory effects, because the agents (in this case, sterile male individuals) are quite quickly lost from the population (in this case, simply through mortality). The sex ratio distorter in Wolbachia is a more persistent form of male control; however, its efficiency still declines over time owing to the eventual evolution of resistance [56].
While the SMT and other approaches in development, such as transgenic manipulation of sex (e.g. the daughterless carp approach [57]), may circumvent the first of these two issues, with sterile individuals of the target pest species being their own vectors of dissemination, the TFT is the only proposed approach that potentially circumvents both problems. As with the SMT, individuals of the species targeted for control are themselves the agents of biocontrol dissemination. However, while released SMs are rapidly lost from target populations through mortality, and the genetic mutations responsible for daughterless carp are selected against and lost in potentially relatively few generations [58], the TFT is self-perpetuating in nature as long as the mtDNA mutations involved do not impact female fitness. As a result, the TFT offers persistent, humane, intergenerational reproductive control of pest populations, at a frequency that is dependent on the frequency of TFs in the population. Control can be enhanced in subsequent generations by releasing more TFs into the population, or reduced by release of wild-type individuals. This ability to attenuate, or completely reverse, the level of reproductive control makes this approach useful in multiple settings, from eradication (e.g. rats) through to management of conservation species where populations outgrow their available habitat (e.g. elephants). For achieving the goal of eradication, the persistent nature of the TFT and the additive effects of multiple releases also offer additional benefits over conventional control. While eradication by lethal control requires a high proportion of a pest population to be killed to stop population growth (otherwise the gains made are eroded by reproduction of the residual population [59]), a step-by-step management approach can potentially be developed with the TFT, with low erosion of biocontrol gains between control operations. In addition, the additive nature of multiple releases makes the TFT applicable to pest species whose life histories make them less suitable for single-application approaches (figure 2). Furthermore, with the persistent regulatory effect of the TFT being determined by the proportion of mutant females as opposed to the absolute number, the TFT will be equally applicable to pest species whose populations are not regulated by density-dependent processes (as is modelled here).
There are five potential hurdles to the development and application of the TFT for pest population control. The first is the need to identify mutations that have asymmetric effects among the sexes. However, finding these mutations may be increasingly simple. We now have examples of such mutations stemming from work in mice [38], hares [40] and fruitflies [35]. As more examples are described, the core characteristics of such mutations will become increasingly well understood, enabling them to be rapidly identified in screens of wild populations or mutagenized stocks. Such screens will be further simplified by high-throughput molecular capture and next-generation sequencing protocols that allow whole-molecule screens of mtDNA variability within large populations [60].
The second hurdle may be the costs of production and release of sufficient TFs to have an appreciable impact at the population scale. However, the TFT could be used in combination with other more traditional forms of pest control, with physical or chemical reduction of the target population immediately prior to the release of TFs, so that fewer individuals are needed to achieve set targets of population suppression. Such an approach is predicted to be beneficial to the efficacy of biocontrol applications in general [20].
The third hurdle may arise if the TF mtDNA type confers major fitness costs to females carrying the mutation versus wild-type females. Under this scenario, the TF mtDNA would be rapidly lost from the population through selection (as for the daughterless carp approach [58]), heavily constraining the prospects for intergenerational control. Male fertility control via TFs would also be compromised by any spontaneous back mutations in the mtDNA, or compensatory nuclear mutations that led to the restoration of mitochondrial function, and thus male fertility. Spontaneous back mutations in the mtDNA that restore fertility are predicted to be infrequent, probably occurring at rates of 1.7–6.2 × 10−8 per site per year [61], and are thus only likely to be an issue at small scales. Compensatory nuclear mutations that restore mitochondrial function are likely to pose more of an issue to the persistence of the TFT as these could rapidly sweep through affected populations as a consequence of their strong selective advantage to males. The existence of mutations in so-called restorer loci have long been known of in plants [62], but we do not yet know whether such compensatory mutations might arise in animals. Nonetheless, both mtDNA back mutation and nuclear compensatory mutations could be readily detected by regular monitoring of mtDNA sequence variation and male fertility parameters within populations under control. The further introduction of original TFs would alleviate the problem of mtDNA back mutation, while the effects of nuclear compensatory mutations might be thwarted using TFs that possess alternative or additional mtDNA genotypes that confer male infertility.
The fourth issue stems from theoretical predictions that under high levels of positive-assortative mating or strong inclusive fitness costs to females [51–53], mtDNA mutations that effect males, but not females will confer indirect selective disadvantages to females, resulting in the elimination of these mutations from the population. Such scenarios are predicted to be rare, but recent work has also demonstrated, at least in theory, that strong spatial population structure could induce selection against such male infertility mutations by increasing the rate of inbreeding and the fitness costs to females of mating with infertile males [63]. Given that many populations exhibit significant population structure, albeit at varying scales, any potential effects of spatial structure on the efficacy of the TFT approach require exploration, probably via spatially explicit modelling in the first instance.
While several factors will act against the success of the TFT, should mtDNA mutations that affect male fertility have direct positive fitness effects on females, or be linked with mtDNA mutations that benefit females, any such fitness advantage would strongly benefit the spread of the mutation conferring male infertility, heightening the prospects for control and potentially eradication. Such sexually antagonistic mutations have been previously identified in fruitflies [64]. Thus, through population screening or experimental evolution it may be possible to generate animals carrying mtDNA variants that not only lead to male sterility, but also confer increased female fitness.
The last hurdle would be if male reproductive fitness were to be only partially affected by the mutations carried by the TF, or if mating was biased towards males that did not carry the TF mutation. Under either scenario the ability of the TFT to affect population control would be reduced significantly. While these issues pose some concerns, the recent report of mtDNA mutations in fruitflies that completely impair male fertility, with no effect on female fertility [35], are reassuring.
Although our results are compelling, we would be the first to point out that our model is relatively simplistic. To reduce the mathematical and computational demands of our simulations, we have used a deterministic framework to provide an averaged and smoothed solution to a problem that clearly has a strong stochastic element, and made several assumptions regarding life-history processes. In addition, we recognize that we have modelled this work assuming a closed population (with a finite and capped population size), while recognizing that most populations in nature are open to migration. Model refinement beyond this first proof of principle is thus desirable to explore further the nuances of how the TFT approach performs under different population and environmental scenarios. Further, the likely effects of both mating system and recruitment compensation on control efficacy at reduced population size, as noted for other fertility control approaches [59,65,66], need to be explored beyond our simple treatment in figure 3. Nonetheless, the conceptual demonstration presented here shows that the TFT has significant theoretical potential—a potential that is readily adaptable to any pest species, because mitochondria are common to all higher organisms [67]. The existence of an analogous situation in plants, cytoplasmic male sterility (CMS), wherein affected plants are unable to produce functional pollen owing to a maternally inherited cytoplasmic factor, adds further weight to the generality of this concept [62]. Indeed, CMS has been widely explored as a tool to limit reproduction of plants under various scenarios, including in biological control [68]. We anticipate that the TFT could have similar and broad utility in other systems.
Acknowledgements
We thank Roger Pech, Phil Cowan, Luke Alphey and an anonymous reviewer for their comments on earlier versions of this manuscript. N.J.G. and R.K.D. conceived this idea, N.J.G., A.J., T.S. and D.M.T. designed the experiments, A.J. and T.S. undertook the modelling, A.J., N.J.G. and D.M.T. wrote a first draft of the manuscript, and all authors contributed substantially to revisions.
Funding statement
N.J.G. acknowledges support from the AgResearch–University of Otago Collaborative Research Fund and the Allan Wilson Centre for Molecular Ecology and Evolution. R.K.D. acknowledges support from an Australian Research Council Future Fellowship (FT100100040). D.M.T. acknowledges Invasive Mammal Impacts MBIE (New Zealand) funding to Landcare Research.
References
- 1.Pimentel D, Lach L, Zuniga R, Morrison D. 2000. Environmental and economic costs of nonindigenous species in the United States. Bioscience 50, 53–65 (doi:10.1641/0006-3568(2000)050[0053:EAECON]2.3.CO;2) [Google Scholar]
- 2.Pimentel D, Zuniga R, Morrison D. 2005. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 52, 273–288 (doi:10.1016/j.ecolecon.2004.10.002) [Google Scholar]
- 3.Salo P, et al. 2007. Alien predators are more dangerous than native predators to prey populations. Proc. R. Soc. B 274, 1237–1243 (doi:10.1098/rspb.2006.0444) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Stenseth NC, et al. 2003. Mice, rats, and people: the bio-economics of agricultural rodent pests. Front. Ecol. Environ. 1, 367–375 (doi:10.1890/1540-9295(2003)001[0367:MRAPTB]2.0.CO;2) [Google Scholar]
- 5.Medina FM, et al. 2011. A global review of the impacts of invasive cats on island endangered vertebrates. Glob. Change Biol. 17, 3503–3510 (doi:10.1111/j.1365-2486.2011.02464.x) [Google Scholar]
- 6.Wardle DA, Verhoef HA, Clarholm M. 2004. Trophic relationships in the soil microfood-web: predicting the responses to a changing global environment. Glob. Change Biol. 4, 713–727 (doi:10.1046/j.1365-2486.1998.00206.x) [Google Scholar]
- 7.Lindenmayer DB. 2009. Forest wildlife management and conservation. Ann. NY Acad. Sci. 1162, 284–310 (doi:10.1111/j.1749-6632.2009.04148.x) [DOI] [PubMed] [Google Scholar]
- 8.Peltzer D, Allen R, Lovett G, Whitehead D, Wardle D. 2010. Effects of biological invasions on forest carbon sequestration. Glob. Change Biol. 16, 732–746 (doi:10.1111/j.1365-2486.2009.02038.x) [Google Scholar]
- 9.Bergstrom DM, Lucieer A, Kiefer K, Wasley J, Belbin L, Pedersen TK, Chown SL. 2009. Indirect effects of invasive species removal devastate World Heritage Island. J. Appl. Ecol. 46, 73–81 (doi:10.1111/j.1365-2664.2008.01601.x) [Google Scholar]
- 10.Clout M, Russell J. 2006. The eradication of mammals from New Zealand islands. In Assessment and control of biological invasion risks (eds Koike F, Clout M, Kawamichi M, De Poorter M, Iwatsuki K.), pp. 127–141 Kyoto, Japan: Shoukadoh Book Sellers [Google Scholar]
- 11.Tompkins DM, Veltman CJ. 2006. Unexpected consequences of vertebrate pest control: predictions from a four-species community model. Ecol. Appl. 16, 1050–1061 (doi:10.1890/1051-0761(2006)016[1050:UCOVPC]2.0.CO;2) [DOI] [PubMed] [Google Scholar]
- 12.Warburton B, Tompkins D, Choquenot D, Cowan P. 2012. Minimising number killed in long-term vertebrate pest management programmes, and associated economic incentives. Anim. Welfare 21(Suppl. 1), 141–149 (doi:10.7120/096272812X13345905674123) [Google Scholar]
- 13.Innes J, Barker G. 1999. Ecological consequences of toxin use for mammalian pest control in New Zealand: an overview. N.Z. J. Ecol. 23, 111–127 [Google Scholar]
- 14.Tabashnik BE, Gassmann AJ, Crowder DW, Carrière Y. 2008. Insect resistance to Bt crops: evidence versus theory. Nat. Biotechnol. 26, 199–202 (doi:10.1038/nbt1382) [DOI] [PubMed] [Google Scholar]
- 15.Grandy J, Rutberg A. 2002. An animal welfare view of wildlife contraception. Reproduction (Suppl.) 60, 1–7 [PubMed] [Google Scholar]
- 16.Singer P. 1997. Neither human nor natural: ethics and feral animals. Reprod. Fertil. Dev. 9, 157 (doi:10.1071/R96060) [DOI] [PubMed] [Google Scholar]
- 17.Courchamp F, Cornell SJ. 2000. Virus-vectored immunocontraception to control feral cats on islands: a mathematical model. J. Appl. Ecol. 37, 903–913 (doi:10.1046/j.1365-2664.2000.00545.x) [Google Scholar]
- 18.Cowan P, Pech R, Curtis P. 2002. Field applications of fertility control for wildlife management. In Reproductive science and integrated conservation (eds Holt WV, Pickard AR, Rodger JC, Wildt DE.), pp. 305–318 Cambridge, UK: Cambridge University Press [Google Scholar]
- 19.McLeod S, Saunders G, Twigg L, Arthur A, Ramsey D, Hinds L. 2008. Prospects for the future: is there a role for virally vectored immunocontraception in vertebrate pest management? Wildl. Res. 34, 555–566 (doi:10.1071/WR07050) [Google Scholar]
- 20.Hone J. 1992. Rate of increase and fertility control. J. Appl. Ecol. 29, 695–698 (doi:10.2307/2404478) [Google Scholar]
- 21.Arthur AD, Krebs CJ, Pech RP, Farroway LN, Singleton GR. 2009. The transmission rate of MCMV in house mice in pens: implications for virally vectored immunocontraception. Wildl. Res. 36, 386–393 (doi:10.1071/WR09006) [Google Scholar]
- 22.Cross M, Zheng T, Duckworth J, Cowan P. 2011. Could recombinant technology facilitate the realisation of a fertility-control vaccine for possums? N.Z. J. Zool. 38, 91–111 (doi:10.1080/03014223.2010.541468) [Google Scholar]
- 23.Tompkins DM. 2007. Minimum specifications for transmissible transgenic biocontrol agents for brushtail possum (Trichosurus vulpecula) population eradication. N.Z. J. Zool. 34, 125–140 (doi:10.1080/03014220709510071) [Google Scholar]
- 24.Vurro M, Gressel J, Butt T, Harman G, St. Leger R, Nuss D, Pilgeram A. 2001. Enhancing biocontrol agents and handling risks. Amsterdam, The Netherlands: IOS Press [Google Scholar]
- 25.Williams C. 2002. Risk assessment for release of genetically modified organisms: a virus to reduce the fertility of introduced wild mice, Mus domesticus. Reproduction (Suppl.) 60, 81. [PubMed] [Google Scholar]
- 26.Dyck VA, Hendrichs J, Robinson A. 2005. Sterile insect technique: principles and practice in area-wide integrated pest management. Dordrecht, The Netherlands: Springer [Google Scholar]
- 27.Alphey L, Benedict M, Bellini R, Clark GG, Dame DA, Service MW, Dobson SL. 2010. Sterile-insect methods for control of mosquito-borne diseases: an analysis. Vector-Borne Zoonot. Dis. 10, 295–311 (doi:10.1089/vbz.2009.0014) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Barclay H. 2005. Mathematical models for the use of sterile insects. In Sterile insect technique: principles and practice in area-wide integrated pest management (eds Dyck VA, Hendrichs J, Robinson AS.), pp. 147–174 Dordrecht, The Netherlands: Springer [Google Scholar]
- 29.Bartlett AC, Staten RT. 1996. The sterile insect release method and other genetic control strategies. In Radcliffe’s IPM world textbook (eds Radcliffe EB, Hutchison WD.). St Paul, MN: University of Minnesota; See http://ipmworld.umn.edu/chapters/bartlett.htm. [Google Scholar]
- 30.Vreysen M, Robinson A, Hendrichs J. 2007. Area-wide control of insect pests from research to field implementation. Dordrecht, The Netherlands: Springer [Google Scholar]
- 31.Nachman MW. 1998. Deleterious mutations in animal mitochondrial DNA. Genetica 102, 61–69 (doi:10.1023/A:1017030708374) [PubMed] [Google Scholar]
- 32.Wallace DC. 1992. Diseases of the mitochondrial DNA. Annu. Rev. Biochem. 61, 1175–1212 (doi:10.1146/annurev.bi.61.070192.005523) [DOI] [PubMed] [Google Scholar]
- 33.Wallace DC. 1999. Mitochondrial diseases in man and mouse. Science 283, 1482–1488 (doi:10.1126/science.283.5407.1482) [DOI] [PubMed] [Google Scholar]
- 34.Camus MF, Clancy DJ, Dowling DK. 2012. Mitochondria, maternal inheritance, and male aging. Curr. Biol. 22, 1–5 (doi:10.1016/j.cub.2012.07.018) [DOI] [PubMed] [Google Scholar]
- 35.Clancy D, Hime G, Shirras A. 2011. Cytoplasmic male sterility in Drosophila melanogaster associated with a mitochondrial CYTB variant. Heredity 107, 374–376 (doi:10.1038/hdy.2011.12) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Holyoake A, McHugh P, Wu M, O'Carroll S, Benny P, Sin I, Sin F. 2001. High incidence of single nucleotide substitutions in the mitochondrial genome is associated with poor semen parameters in men. Int. J. Androl. 24, 175–182 (doi:10.1046/j.1365-2605.2001.00292.x) [DOI] [PubMed] [Google Scholar]
- 37.Innocenti P, Morrow EH, Dowling DK. 2011. Experimental evidence supports a sex-specific selective sieve in mitochondrial genome evolution. Science 332, 845–848 (doi:10.1126/science.1201157) [DOI] [PubMed] [Google Scholar]
- 38.Nakada K, Sato A, Yoshida K, Morita T, Tanaka H, Inoue SI, Yonekawa H, Hayashi JI. 2006. Mitochondria-related male infertility. Proc. Natl Acad. Sci. USA 103, 15 148–15 153 (doi:10.1073/pnas.0604641103) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ruiz-Pesini E, et al. 2000. Human mtDNA haplogroups associated with high or reduced spermatozoa motility. Am. J. Hum. Genet. 67, 682–696 (doi:10.1086/303040) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Smith S, Turbill C, Suchentrunk F. 2010. Introducing mother's curse: low male fertility associated with an imported mtDNA haplotype in a captive colony of brown hares. Mol. Ecol. 19, 36–43 (doi:10.1111/j.1365-294X.2009.04444.x) [DOI] [PubMed] [Google Scholar]
- 41.Trifunovic A, et al. 2004. Premature ageing in mice expressing defective mitochondrial DNA polymerase. Nature 429, 417–423 (doi:10.1038/nature02517) [DOI] [PubMed] [Google Scholar]
- 42.Wolff JN, Gemmell NJ. 2013. Mitochondria, maternal inheritance and asymmetric fitness: why males die younger. BioEssays 35, 93–99 (doi:10.1002/bies.201200141) [DOI] [PubMed] [Google Scholar]
- 43.Xu H, DeLuca SZ, O'Farrell PH. 2008. Manipulating the metazoan mitochondrial genome with targeted restriction enzymes. Science 321, 575–577 (doi:10.1126/science.1160226) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gemmell NJ, Metcalf VJ, Allendorf FW. 2004. Mother's curse: the effect of mtDNA on individual fitness and population viability. Trends. Ecol. Evol. 19, 238–244 (doi:10.1016/j.tree.2004.02.002) [DOI] [PubMed] [Google Scholar]
- 45.Rossignol R, Faustin B, Rocher C, Malgat M, Mazat JP, Letellier T. 2003. Mitochondrial threshold effects. Biochem. J. 370, 751 (doi:10.1042/BJ20021594) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wolff JN, Gemmell NJ. 2008. Estimating mitochondrial DNA content of chinook salmon spermatozoa using quantitative real-time polymerase chain reaction. Biol. Reprod. 79, 247–252 (doi:10.1095/biolreprod.107.067009) [DOI] [PubMed] [Google Scholar]
- 47.Wolff JN, Gemmell NJ. 2008. Lost in the zygote: the dilution of paternal mtDNA upon fertilization. Heredity 101, 429–434 (doi:10.1038/hdy.2008.74) [DOI] [PubMed] [Google Scholar]
- 48.Cardullo RA, Baltz JM. 2005. Metabolic regulation in mammalian sperm: mitochondrial volume determines sperm length and flagellar beat frequency. Cell Motil. Cytoskeleton 19, 180–188 (doi:10.1002/cm.970190306) [DOI] [PubMed] [Google Scholar]
- 49.Frank S, Hurst L. 1996. Mitochondria and male disease. Nature 383, 224 (doi:10.1038/383224a0) [DOI] [PubMed] [Google Scholar]
- 50.Gemmell NJ, Allendorf FW. 2001. Mitochondrial mutations may decrease population viability. Trends. Ecol. Evol. 16, 115–117 (doi:10.1016/S0169-5347(00)02087-5) [DOI] [PubMed] [Google Scholar]
- 51.Unckless RL, Herren JK. 2009. Population genetics of sexually antagonistic mitochondrial mutants under inbreeding. J. Theor. Biol. 260, 132–136 (doi:10.1016/j.jtbi.2009.06.004) [DOI] [PubMed] [Google Scholar]
- 52.Wade MJ, Brandvain Y. 2009. Reversing mother's curse: selection on male mitochondrial fitness effects. Evolution 63, 1084–1089 (doi:10.1111/j.1558-5646.2009.00614.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Hedrick PW. 2012. Reversing mother's curse revisited. Evolution 66, 612–616 (doi:10.1111/j.1558-5646.2011.01465.x) [DOI] [PubMed] [Google Scholar]
- 54.Duckworth J, Byrom A, Fisher P, Horn C. 2006. Pest control: does the answer lie in new biotechnologies? In Biological invasions in new Zealand (eds Allen RB, Lee WG.), pp. 421–434 Berlin, Germany: Springer [Google Scholar]
- 55.Cooper DW, Larsen E. 2006. Immunocontraception of mammalian wildlife: ecological and immunogenetic issues. Reproduction 132, 821–828 (doi:10.1530/REP-06-0037) [DOI] [PubMed] [Google Scholar]
- 56.Iturbe-Ormaetxe I, Walker T, O'Neill SL. 2011. Wolbachia and the biological control of mosquito-borne disease. EMBO Rep. 12, 508–518 (doi:10.1038/embor.2011.84) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Saunders G, Cooke B, McColl K, Shine R, Peacock T. 2010. Modern approaches for the biological control of vertebrate pests: an Australian perspective. Biol. Control 52, 288–295 (doi:10.1016/j.biocontrol.2009.06.014) [Google Scholar]
- 58.Brown P, Walker TI. 2004. CARPSIM: stochastic simulation modelling of wild carp (Cyprinus carpio L.) population dynamics, with applications to pest control. Ecol. Model. 176, 83–97 (doi:10.1016/j.ecolmodel.2003.11.009) [Google Scholar]
- 59.Hone J. 1999. On rate of increase (r): patterns of variation in Australian mammals and the implications for wildlife management. J. Appl. Ecol. 36, 709–718 (doi:10.1046/j.1365-2664.1999.00439.x) [Google Scholar]
- 60.Vasta V, Ng SB, Turner EH, Shendure J, Hahn SH. 2009. Next generation sequence analysis for mitochondrial disorders. Genome Med. 1, 100 (doi:10.1186/gm100) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Montooth KL, Rand DM. 2008. The spectrum of mitochondrial mutation differs across species. PLoS Biol. 6, e213 (doi:10.1371/journal.pbio.0060213) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Schnable PS, Wise RP. 1998. The molecular basis of cytoplasmic male sterility and fertility restoration. Trends Plant Sci. 3, 175–180 (doi:10.1016/S1360-1385(98)01235-7) [Google Scholar]
- 63.Zhang H, Guillaume F, Engelstädter J. 2012. The dynamics of mitochondrial mutations causing male infertility in spatially structured populations. Evolution 66, 3179–3188 (doi:10.1111/j.1558-5646.2012.01675.x) [DOI] [PubMed] [Google Scholar]
- 64.Rand DM, Clark AG, Kann LM. 2001. Sexually antagonistic cytonuclear fitness interactions in Drosophila melanogaster. Genetics 159, 173–187 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Barlow N, Kean J, Briggs C. 1997. Modelling the relative efficacy of culling and sterilisation for controlling populations. Wildl. Res. 24, 129–141 (doi:10.1071/WR95027) [Google Scholar]
- 66.Caughley G, Pech R, Grice D. 1992. Effect of fertility control on a population's productivity. Wildl. Res. 19, 623–627 (doi:10.1071/WR9920623) [Google Scholar]
- 67.White DJ, Wolff JN, Pierson M, Gemmell NJ. 2008. Revealing the hidden complexities of mtDNA inheritance. Mol. Ecol. 17, 4925–4942 (doi:10.1111/j.1365-294X.2008.03982.x) [DOI] [PubMed] [Google Scholar]
- 68.Hodgins KA, Rieseberg L, Otto SP. 2009. Genetic control of invasive plants species using selfish genetic elements. Evol. Appl. 2, 555–569 (doi:10.1111/j.1752-4571.2009.00102.x) [DOI] [PMC free article] [PubMed] [Google Scholar]