

Abstract
Background
A program, supported by the GEMHEP (Groupe d'étude Moléculaire des Hépatites), was established in 2007 in the sanitary district of Tokombéré, to prevent perinatal transmission of hepatitis B virus (HBV). It comprises screening for HBV surface antigen (HBsAg) in all pregnant women and vaccinating the newborn if tests are positive.
Methods/Principal Findings
1276 women were enrolled in the study after providing informed consent. Demographic data and blood samples were available for 1267 of the enrolled patients. HBsAg was determined locally using a rapid test (Vikia HBsAg, Biomerieux). Tests for HBV and HDV virological markers (HBeAg, anti-HDV antibodies (Ab), HBV-DNA, HDV-RNA, HBV and HDV genotypes) were performed on the confirmed HBsAg-positive samples in the virology unit of the Angers University Hospital (France). HBsAg was found in 259 of the 1267 pregnant women (20.4%) between January 2009 and April 2010, of whom 59 were HBeAg-positive (22.7%) with high levels of HBV-DNA. Anti-HDV Ab were found in 19 (7.3%) of the HBsAg-positive women. The prevalence rates of HBsAg and HDV were not age-dependent whereas HBeAg carriers were statistically younger than non carriers. Basal core promoter (BCP) and precore (PC) mutations and genotypes were determined by sequencing. Of 120 amplified sequences, 119 belonged to HBV genotype E (HBV/E) and the 9 HDV strains belonged to HDV clade 1. In the PC region, 83/228 patients (36.4%) harbored a G1896A mutant or mixed phenotype virus. In the BCP region, the double mutation A1762T/G1764A and the G1757A substitution were detected respectively in 26/228 patients (11.4%) and 189/228 patients (82.8%).
Conclusions
Our results confirm the high prevalence and low molecular diversity of HBV in Far Northern Cameroon; more than 20% of the infected women were highly viremic, suggesting a high rate of HBV perinatal transmission and supporting the WHO recommendation to vaccinate at birth against hepatitis B.
Introduction
According to the World Health Organization (WHO), more than 350 million people are chronic carriers of hepatitis B virus (HBV)[1,2]. HBV chronic infection results in 500,000 to 700,000 deaths per year caused by chronic hepatitis, cirrhosis and hepatocellular carcinoma. HBV is the 10th leading cause of death worldwide [3-5]. Africa is reported to have the highest overall prevalence rate, although there are substantial regional variations ranging from less than 7% to more than 20% [6-9]. In general, the prevalence of HBV surface antigen (HBsAg) increases with age. The earlier the infection occurs in life, the higher the probability of becoming a chronic carrier increases [8,10-12]. Currently, eight genomic groups of HBV are recognized, designated as genotypes A to H. Of these, A and E are considered to be major genotypes and are predominant in central, south, and west Africa [13-16]. Hepatitis delta virus (HDV) is also highly endemic in several African countries [17,18]. Extensive analyses of HDV sequences have illustrated a wide genetic diversity grouped into eight clades [19,20]. However, little information is available on the origin, circulation and genetic diversity of both HBV and HDV infections, especially in Central Africa.
More efficacious treatments, mass immunization programs, and safe injection techniques are essential for eliminating HBV infection and reducing global HBV-related morbidity and mortality. Safe and effective vaccines against HBV infection have been available since 1982. The implementation of mass immunization programs, which have been recommended by WHO since 1991, has dramatically decreased the incidence of HBV infection among infants, children, and adolescents in many countries. However, not all countries have adopted these recommendations and many people infected with HBV prior to the implementation of immunization programs are still alive. In infants born to HBeAg-positive mothers, the reported post-vaccination HBsAg carrier rate ranges from 7.4% to 17.3%, depending on the different types of vaccines, injection schedules, and age of assessment [21].
A program was established in 2007 in the sanitary district of Tokombéré, a rural district located in Far Northern Cameroon, to evaluate and prevent mother-to-child transmission of HBV. The program is supported by the GEMHEP (Groupe d'Étude Moléculaire des Hépatites). It involves screening for HBV surface antigen (HBsAg) in all pregnant women and vaccinating newborns when the tests are positive.
The objectives of our study were to establish a more precise measure of the high prevalence of HBV and HDV infection and to analyze HBV genomic diversity in both the polymerase and PC region in this poorly documented area of Cameroon.
Materials and Methods
Patient samples
Since 2005, the GlaxoSmithKline foundation has been supporting a program initially dedicated to the prevention of mother-to-child transmission (PMTCT) of HIV. The reach of the program was broadened in 2007 with a new focus on strengthening healthcare both for mothers and for children less than five years of age. Within this setting and with the support of the GEMHEP, a new PMTCT program for HBV was established. We screened all pregnant women for HIV and HBsAg simultaneously. Our project was approved by the Ethics Committee at the University Hospital of Angers, France (Agreement #2013-49). Our data protection has been registered with the CNIL as statement number 1684412. The patients were informed that the samples could be used for research purposes. Only the pregnant women who provided informed consent were included. From January 2009 to April 2010, we collected 1276 sera from consecutive women who had no clinical or biological signs of severe hepatitis. Then, we proposed immunization for the newborns of HBsAg-positive mothers. Immunization comprised HBV monovalent vaccine at birth plus completion of the primary series at 6, 10 and 14 weeks of the Expanded Programme on Immunization with hepatitis B component (EPI-HB) recommended by WHO.
Serological testing
Blood samples (5 mL) were collected in the 8 sanitary formations (SF) of the Tokombéré district during the first prenatal consultation after HIV counseling. Initial HIV and HBV tests were performed in the Tokombéré hospital laboratory. HBsAg was detected by a rapid immunochromatographic test (VIKIA HBs Ag, Biomerieux, Marcy l'Etoile, France).
Sera were then stored at -20C° and sent to the virology laboratory of the Angers for complementary serological and molecular tests. The sera were tested with a commercial immunoassay test (ETI-MAK-4, Diasorin, Antony, France) and HBsAg-positive samples were also confirmed by neutralization (Diasorin, Antony, France). In all HBsAg-positive subjects, HBeAg and hepatitis Delta antibodies (anti-HDV Ab) were searched using two ELISA tests, respectively ETI-EBK-PLUS and ETI-ABDELTAK-2 (Diasorin, Antony, France). Anti-HDV IgM was determined in the anti-HDV Ab positive samples (ETI-DELTA-IGMK-2, Diasorin, Antony, France).
Nucleic Acid extraction
Nucleic acid extractions were performed using the Nuclisens EasyMAG Platform (Biomerieux) according to the manufacturer’s instructions, from 200 µL of serum. HBV DNA and HDV RNA were eluted in 50 µL of elution buffer. Purified nucleic acid was used for HBV DNA and HDV RNA detection and genotyping.
HBV DNA detection
For all patients, the full length HBV genome was amplified with a qualitative PCR using primers HBV T1 (forward, 5'-cca cca ctt tcc acc aaa ct-3') and HBV T2 (reverse, 5'-gta ggc tgc ctt cct gac tg-3') designed in the Angers. 10 µL of extracted DNA was used for PCR in a 50 µL reaction volume containing colorless GoTaq Flexi buffer 1X, 1.5 mM MgCl2, 100 µM dNTP, 5 U GoTaq Flexi DNA polymerase (Promega) and 300 nM of each primer. Thermal profile amplification in the Verity thermocycler (Life technologies) was: pre-heating at 94°C for 5 minutes, 40 cycles including denaturation at 94°C for 1 minute, annealing at 58°C for 1 minute, and extension at 72°C for 4 minutes, followed by 10 minutes at 72°C. PCR products covering the full HBV genome (3162 bp) were used as templates for two nested PCR.
For polymerase region (P) amplification (809 bp), 5 µL of full length HBV amplicons were mixed in a 50 µL reaction volume containing GoTaq Flexi buffer 1X, 1.5mM MgCl2, 200 µM dNTP, 2.5 U GoTaq Flexi DNA polymerase (Promega) and 200 nM of each primer POL3 (forward, 5'-gac tcg tgg tgg act tct ctc a-3') and POL4 (reverse, 5'-ggc att aaa gca gga tat cca cat tg-3') [22] as previously described. Thermal profile amplification was: pre-heating at 94°C for 5 minutes, 30 cycles including denaturation at 94°C for 40 seconds, annealing at 55°C for 1 minute, and extension at 72°C for 1 minute, followed by 10 minutes at 72°C. PCR products of polymerase gene were controlled by electrophoresis. The result of electrophoresis was used to identify positive and negative samples for HBV DNA detection. Based on our experience, only samples with a viral load >3 log IU/mL were detectable with this in-house PCR method. Positive samples were also used to determine HBV genotype by sequencing.
For PC region amplification (363 bp), 10 µL of full length HBV PCR products were mixed in a 90 µL reaction volume containing GoTaq Flexi buffer 1X, 1.5 mM MgCl2, 200 µM dNTP, 2.5 U GoTaq Flexi DNA polymerase (Promega) and 400 nM of each primer PC3 (forward, 5'-TGTCAACGACCGACCTTGAG-3') and PC4 (reverse, 5'- GCAATGCTCAGGAGACTCTAAGGC- 3') [23] as previously described. Thermal profile amplification was: pre-heating at 94°C for 5 minutes, 30 cycles including denaturation at 94°C for 40 seconds, annealing at 55°C for 1 minute, and extension at 68°C for 1 minute, followed by 10 minutes at 72°C. PCR products of PC gene were controlled by electrophoresis and used to determine PC and BCP phenotypes by sequencing.
For 72 consecutive patients, serum HBV DNA levels were determined with the Abbott Real-Time HBV PCR assay (Abbott) on the M2000sp-M2000rt platform. Due to the small volumes, sera were diluted to a final concentration of 1/5 (v/v) in Abbott RealTime negative HBV control. The lower detection limit was also increased from 1 to 1.7 log IU/mL to take the dilution into account.
HDV RNA detection
HDV-RNA was detected by PCR using primers 900s and 1280as, which encompass a region conserved in all HDV genotypes, i.e., the “R0 region” (400 nt) covering the 3’end of the HDV gene as previously described [24]. PCR products of the R0 region were controlled by electrophoresis and used to determine HDV genotype by sequencing.
Sequence analysis of HBV and HDV strains
HBV PCR products (covering the P and BCP/PC regions) and HDV PCR products were purified using Nucleospin Extract II columns (Machery Nagel) and then sequenced bi-directionally using the BigDye Terminator v3.1 kit (Life Technologies). After purification, sequences of amplified nucleic acids were determined using an ABI 3130 xl sequencer (Life Technologies) and the nucleotide sequences analyzed with SeqScape® v2.6 software (Life Technologies). Nucleotide sequences of the P region were compared with reference strains representing each of the genotypes A-H, obtained from GenBank. Genotyping of HBV was then determined by blast analysis, and full genome sequences representing the eight HBV genotypes were used as references.
Phylogenic analyses in both the polymerase gene and the BCP and PC gene were performed to assess the genomic diversity of the strains.
Sequence changes of the PC and BCP regions were used to determine the wild/mixed/mutated PC and BCP phenotypes. In the PC region, two nucleotide changes were analyzed: the presence of a point mutation from G to A at nucleotide 1896 (G1896A), which signals the mutant PC phenotype; and the change C to T at position 1858, which defines the C1858T mutation. In the BCP region, the two-nucleotide substitutions, A-T at nucleotide 1762 and G-A at nucleotide 1764 (A1762T/G1764A), signal the double mutant BCP phenotype. Another mutation was investigated at the 1757 nucleotide position to detect change G1757 to A1757. Mixed phenotype was defined as the simultaneous presence of both wild and mutated phenotypes.
Genotyping of HDV was determined by blast analysis, and full genome sequences representing the eight HDV genotypes were used as references.
Statistical analysis
Quantitative variables were compared using Student t test. Qualitative variables were compared by the chi square test or the bilateral Fisher's exact test when the expected value of at least one cell in a contingency table was below 5. A p value of 0.05 or less was considered significant. To assess risk factors, as our study was cross-sectional, we computed risk ratios (i.e., ratio of prevalence in exposed and non-exposed women) and their 95% confidence intervals rather than odds ratios. HBV viral load was expressed as mean ± standard deviation (± SD). All statistical calculations were performed using SPSS software (IBM Corporation, Armonk, NY, USA).
Results
HBV and HDV serological screening
None of the 1276 pregnant women had been vaccinated against HBV or had received antiviral treatment. Demographic data and blood samples were available for 1267 of the enrolled patients.
HBsAg was detected in 259 of 1267 sera (20.4%) by the VIKIA HBsAg test. Negative and positive results for HBsAg were confirmed by EIA and neutralization [25,26]. HBeAg was found in 59 of the 259 HBsAg-positive pregnant women (22.7%). Eight pregnant women were anti-HIVAb-positive. Nineteen of the 259 (7.3 %) HBsAg-positive pregnant women were anti-HDV antibody (Ac) positive with anti-HDV IgM in 10/19 (53%) cases.
No difference in age was observed for HBsAg carriers versus non carriers (24.5 years versus 24.3 years; p=0.73) or HDV Ab carriers versus non carriers (26 years versus 24.3 years; p=0.3). Conversely, HBeAg carriers were statistically younger than non carriers (22.9 years versus 24.95 years; p=0.041, 95% confidence interval for the difference of means: 0.08-3.95 years). Tables 1, 2 and 3 summarize the results of the HBsAg, HBeAg and HDV prevalence according to age range (quartiles).
Table 1. Prevalence of HBsAg by age group in 1276 pregnant women in the district of Tokombéré.
| Variable | HBs Ag |
||
|---|---|---|---|
| n positive/n tested (%) | RR (95% CI)* | P value | |
| Age range (yr) | |||
| < 20 | 74/335 (22.1%) | 1 | |
| 20–23 | 60/330 (18.2%) | 0.95 (0.88-1.03) | 0.21 |
| 24–28 | 48/278 (17.3%) | 0.94 (0.87-1.02) | 0.14 |
| > 28 | 76/324 (23.5%) | 1.02 (0.94-1.11) | 0.68 |
| All | 258/1267 (20.4%) | ||
* risk ratio and 95% confidence interval
Table 2. Prevalence of HBeAg by age group in 1276 pregnant women in the district of Tokombéré.
| Variable | HBe Ag |
||
|---|---|---|---|
| n positive/n tested (%) | RR (95% CI)* | P value | |
| Age range (yr) | |||
| < 20 | 23/74 (31.1%) | 1 | |
| 20–23 | 10/60 (16.7%) | 0.83 (0.68-1) | 0.054 |
| 24–28 | 13/48 (27.1%) | 0.95 (0.74-1.2) | 0.64 |
| > 28 | 11/77 (14.3%) | 0.80 (0.67-0.96) | 0.014 |
| All | 57/259 (22%) | ||
* risk ratio and 95% confidence interval
Table 3. Prevalence of HDV total antibodies by age group in 1276 pregnant women in the district of Tokombéré.
| Variable | HDV Ab |
||
|---|---|---|---|
| n positive/n tested (%) | RR (95% CI)* | P value | |
| Age range (yr) | |||
| < 20 | 3/74 (4.1%) | 1 | |
| 20–23 | 4/60 (6.7%) | 1.03 (0.95-1.12) | 0.7‡ |
| 24–28 | 5/47 (10.6%) | 1.07 (0.97-1.2) | 0.26‡ |
| > 28 | 7/76 (9.2%) | 1.06 (0.97-1.15) | 0.33‡ |
| All | 19/257 (7.4%) | ||
* risk ratio and 95% confidence interval
bilateral Fisher's exact test
HBV DNA detection and quantification
As shown in Figure 1, HBV DNA was detected in 121 of the 259 HBsAg-positive samples (47.1%) by qualitative PCR with a sensitivity ~3 log IU/mL; 57 of these pregnant women (48%) were HBeAg-positive. Thus, in 138 pregnant women (53.3%), HBV DNA was undetectable, 98.5% (136/138) being HBeAg-negative and two HBeAg-positive; interestingly, both of these were co-infected with HDV (HBV DNA levels were <1.7 and 1.97 log IU/mL).
Figure 1. Results of the polymerase gene HBV amplification.

The HBV DNA was amplified using a qualitative PCR with a threshold at 3 log UI/mL and the results were correlated with the HBeAg profile in HBsAg-positive pregnant women.
Serum HBV DNA levels were quantified in 72 consecutive samples (qualitative detection of HBV DNA was negative in 43/72 and positive in 29/72 samples). The mean of the viral HBV DNA was 3.44 log IU/mL using the Abbott Real Time HBV PCR, 21 pregnant women did not have a HBV target detected and 51 had a detectable viral load. All women with an undetectable viral load were HBeAg-negative. Among the 51 pregnant women with detectable viral loads, most had an HBV DNA level inferior to 3 log IU/mL. However the viral load was greater than 7 log IU/mL in 16 women (22.2%) (Figure 2). Figure 3 illustrates the repartition of HBV viral load according to HBeAg phenotype. As expected, HBeAg-positive pregnant women had an HBV viral load significantly higher than HBeAg-negative ones: 7.47 (± 2.09) log IU/mL versus 2.2 (± 1.48) log IU/mL (p <0.001).
Figure 2. HBV DNA quantification.
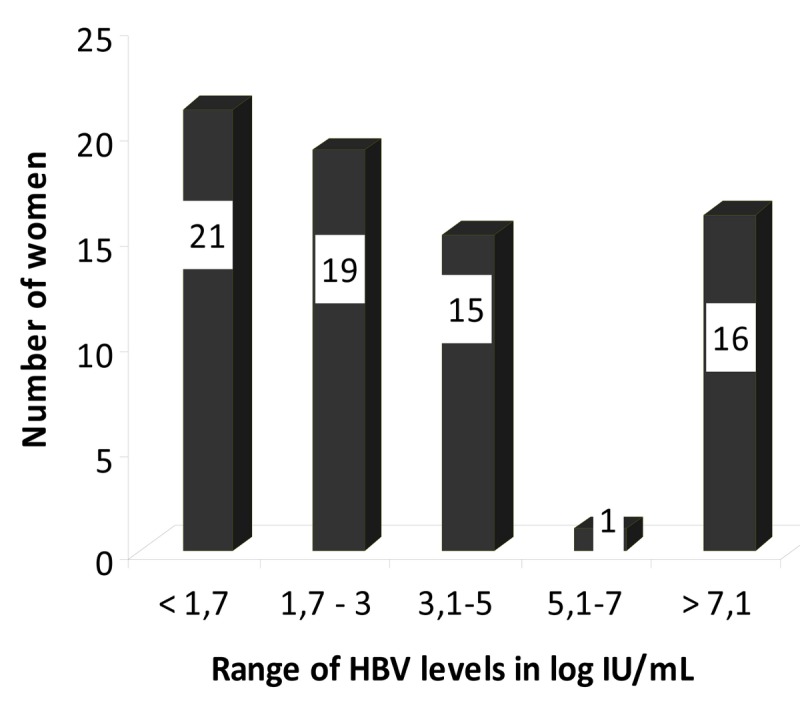
Serum HBV DNA levels were quantified in 72 consecutive samples with the Abbott Real Time HBV PCR (threshold at 1.7 log IU/mL). HBV target was not detected in 21 patients; viral load was detected in 51 patients, 16 of whom had a viral load greater than 7 log IU/mL.
Figure 3. Plots of Log IU/mL HBV-DNA according to the HBe status.

Quantification of HBV DNA using the Abbott Real Time HBV PCR: the median of the log IU/mL HBV viral load (solid black bar in the figure) was significantly higher in HBeAg-positive pregnant women than HBeAg-negative ones (8.33 log IU/mL versus 2.05 log IU/mL respectively).
HBV and HDV genotyping
Of the 120 samples we were able to amplify in the polymerase gene, 99% (119/120) belonged to genotype E; the one exception was genotype D. Phylogenic analyses both in the polymerase gene and the BCP and PC gene showed that the strains were slightly different and closely related to other strains of African origin, notably the strains we have described in Niger and Mauritania [27,28]. As for HDV, HDV RNA was detected by PCR in 9/19 (47%) cases, all being anti-HDV IgM positive. The 9 HDV strains belonged to HDV clade 1.
Characterization of BCP/PC phenotypes
PC/BCP sequences were characterized by direct sequencing in 228 samples: 59 HBeAg positive and 169 HBeAg negative. In the position 1896, wild (1896G) mixed (1896G and A) and mutated (1896A) phenotypes were observed in 145/228 (63.5%), 34/228 (14.9%) and 49/228 (21.4%) women respectively (Table 4). The wild PC phenotype was the dominant detected viral population regardless of the HBeAg profile: 94.9% (56/59) and 52.6% (89/169) in HBeAg-positive and -negative patients respectively (p = 0.01). The frequency of mutant or mixed PC populations was higher in HBeAg-negative (80/169) than in HBeAg-positive (3/59) pregnant women (47% versus 5%, p<0.05). Patients who were infected with a mutant PC virus had lower mean HBV DNA viral load than pregnant women who harbored a wild PC virus (3.11 ± 1.66 logIU/mL versus 5.89 ± 2.72 logIU/mL, p < 0.05, data not shown). The mixed/mutated BCP phenotype was detected in 61/228 patients (26.7%); there was no significant difference in the mutant BCP phenotype distribution between the HBeAg-negative and HBeAg-positive pregnant women: 13/59 (22%) and 48/169 (28.4%) respectively (p=0.8). The C1858T and G1757A substitutions were found in 226/228 (99.1%) and 189/228 patients (82.8%) respectively.
Table 4. Profile of BCP and PC mutations according to HBeAg status.
| BCP/PC phenotypes | |||||
|---|---|---|---|---|---|
| Wild Number/Total (%) | Mixed Number /Total (%) | Mutated number/Total (%) | |||
| G1896A | HBe (+) Ag | 56/59 (94.9%) | 2/59 (3.3%) | 1/59 (2%) | |
| HBe (-) Ag | 89/169 (52.6%) | 32/169 (19%) | 48/169 (28%) | ||
| Total | 145/228 (63.5%) | 34/228 (14.9%) | 49/228 (21.4%) | ||
| G1757A | HBe (+) Ag | 3/59 (5%) | 0/59 (0%) | 56/59 (95%) | |
| HBe (-) Ag | 12/169 (7%) | 24/169 (14%) | 133/169 (79%) | ||
| Total | 15/228 (6.5%) | 24/228 (10.5%) | 189/228 (82.8%) | ||
| A1762T/G1764A | HBe (+) Ag | 46/59 (77.9%) | 5/59 (8.4%) | 8/59 (13.5%) | |
| HBe (-) Ag | 121/169 (72%) | 30/169 (18%) | 18/169 (11%) | ||
| Total | 167/228 (73.2%) | 35/228 (15.3%) | 26/228 (11.4%) | ||
| C1858T | HBe (+) Ag | 1/59 (1.6%) | 0/59 (0%) | 58/59 (98.3%) | |
| HBe (-) Ag | 0/169 (0%) | 3/169 (2%) | 166/169 (98.2%) | ||
| Total | 1/228 (0.004%) | 3/228 (1.3%) | 226/228 (99.1%) | ||
Discussion
In the present study, we focused on estimating the prevalence of HBV and HDV in pregnant women in the rural district of Tokombéré in Far Northern Cameroon and exploring virus genome diversity. The population of Tokombéré is currently estimated at 130 000 inhabitants with an average density of 260 inhabitants per km2. The prevalence of HBsAg in Cameroon is usually estimated to be between 5 and 10%. The HBsAg prevalence of 20.4% in our study is thus surprisingly high. As concerns other studies conducted in Cameroon, we found only one done recently in pregnant women; it reported, in the Yaoundé area, a prevalence of HBsAg of 7.85% with a very low proportion of HBeAg [29]. Still in Cameroon, the prevalence of HBV infection was assessed at 11.8% in three pygmy groups in 2011 [30] and HBsAg prevalence was reported to be 10.7% in Yaoundé blood donors in 2003 [31]. Our high HBsAg prevalence remains coherent with published data at the scale of central sub-Saharan Africa. Variable endemicity data are reported in other countries: 6.5% in Zambia, 9.5% in Gabon, 10.9 % in Mauritania, 12.6% in Ghana, 13.8% in Senegal, 15.5% in Mali and 25% in Zimbabwe [32-37]. Furthermore, HBV rate also differs intranationally in some countries with areas of high endemicity. In Nigeria, for example, studies reported varying seroprevalence rates of HBsAg seropositivity, i.e., 26.0%, 15.1%, 13.2% and 12.6% according to location [38]. The Tokombéré district is located in a part of Cameroon that is pinched between the politically unstable countries of Chad and Nigeria, both presenting a significant rate of emigration and seemingly high HBsAg prevalence (7-30%) [39]. This may help explain the high rate we found. Another explanation may be the nature of the district, rural and mountainous, making access to health facilities difficult and resulting in home deliveries with a higher risk of perinatal contamination.
Concerning HBeAg prevalence, we found a higher HBeAg prevalence than described by Kfutwah et al. In that study, including 650 pregnant women with a mean age of 26.2 years, none of the HBeAg of the 45 HBsAg-positive samples were reactive [29]. However, Jordis et al reported that the highest HBeAg prevalence (over 50%) was found in 0 to 9 year-old girls. At the reproductive age, HBeAg prevalence was 20-50% [40]. Interestingly, the authors demonstrated that HBeAg prevalence decreased between 1990 and 2005 overall, but that the reduction occurred to a lesser extent in the reproductive age-group of 20-39 year old females. Their systematic literature review showed that, in central sub-Saharan Africa, the highest HBeAg positivity rate (54.86%) was observed in the 0-9 age group. The rate then decreased successively: 40.20% in the 10-19 age group, 29.98% in the 20-29 age group and 22.70% in the 30-39 age-group. Our findings correlated well with those of Jordis et al: the highest HBeAg prevalence we observed was in women <20 years old but it declined thereafter in the others groups, with overall prevalence at 22%.
In our study, 119 of the 120 amplified sequences belonged to HBV genotype E (HBV/E). Inter-sample contaminations were avoided by the phylogenic analysis showing minor nucleotide modification in all strains. In Africa, the spread and evolution of HBV genotypes remain poorly understood, but genotype D seems to be the most prevalent in the north. Genotype A is dominant in the southern and eastern Africa regions while HBV/E predominates throughout a vast region ranging from Senegal to Namibia, then eastward to the Central African Republic. HBV/E is thus the most prevalent genotype found in the countries bordering on or close to Cameroon (e.g., Central African Republic, Democratic Republic of the Congo, Benin, Togo, and Nigeria). In Cameroon, HBV/A and/or HBV/E have both been described, depending on the cohorts studied [41-43]. Co-circulation, recombination and a high number of mixed infections have been detected because of this dual genotypic presence, but no evidence whatsoever of this was detected in the population from the Tokombéré district. A unique genotype, combined with the low genetic diversity of HBV/E, supports the hypothesis of an endemic infection in this area.
Concerning the HBV heterogeneity in the PC region, the most frequently reported mutation is G1896A, which pairs with a T in nucleotide 1858. This base-pairing increases the stability of the stem-loop structure of the encapsidation signal, a critical factor for viral replication; thus non-A genotypes favor the emergence and selection of 1896 PC mutants. Our results confirm those of previous reports indicating a G1896A mutation prevalence in African pregnant women inferior to 50% [33,42]. In Guinea, a similar G1896A mutation prevalence (21%) was described in blood donors infected with HBV/E [44]. In our population, the majority of PC sequences (98%) harbored the C1858T mutation. Many studies have demonstrated the high frequency of this mutation in HBV/E [44,45] as well as in genotypes B-D. In our study, it was surprising to note the low prevalence of the G1896A mutation (21.4%) with the high percentage of the C1858T mutation (99.1%) detected in HBV/E, whereas these two point mutations are usually associated. The low frequency of the PC mutation could be explained by the high prevalence of the G1757A mutation in our population. In the literature, there are few data on the mutation at nucleotide 1757, but it seems to be more prevalent in genotypes D and E and to affect negatively the emergence of BCP mutations [45-47]. A recent paper demonstrated that the G1757A substitution is associated with protection against advanced liver disease (P = 0.001).
Concerning the core promoter mutations, 11.3% of the pregnant women harbored strains with the double mutation T1762/A1764. Similar results for BCP mutants have been reported in Ghana [33] and the Central African Republic [48]. Thus, more than half of the pregnant women were infected with viral strains with one or two mutations in nucleotides at positions 1762, 1764 or 1896. A unique triple mutant T1762/A1764/A1896 was found in 11.4% of the strains.
The impact of mutations in the HBV PC and BCP regions on the course of chronic liver disease is not well established. At the time of inclusion, most of the women were clinically asymptotic, but few biochemical (ALAT) or fibrosis markers were available. However, it could be considered that HBsAg-positive pregnant women with high levels of HBV DNA are in the immune tolerant phase of infection and the others are inactive carriers. The low prevalence of the PC/BCP mutants in conjunction with the high percentage of the G1757A mutation might influence the clinical characteristics of HBV/E. It seems necessary to complete these data with a clinical study aimed at investigating the prevalence of the PC/BCP mutated viruses in HBV/E-infected subjects.
We also found a HDV prevalence of about 7% in this relatively young asymptotic population. This is substantially lower than the prevalence of 6/17 found in the Tokombéré medical staff [25] and quite lower than the 20% prevalence found in pregnant women in Niger or in Mauritania in recent papers [27,28].
Conclusions
The high prevalence of HBV infection in pregnant women in the Far Northern Region of Cameroon, together with the relatively high proportion of women with high viral load, may be responsible for:
1-A high risk of vertical transmission of HBV in this population, justifying routine immunization at birth with HBV monovalent vaccine. The EPI-HB started at 6 weeks of age at best may be insufficient to avoid certain mother-to-child contaminations. This justifies the addition of Hepatitis B immunoglobulin (HBIg) or preemptive treatment with nucleos(t)ides analogs, as previously suggested, to immunization at birth [49].
2-A probably higher risk of severe outcome particularly in HBe Ag-positive highly viremic mothers. This may include, in the absence of treatment, decompensated cirrhosis and later hepatocellular carcinoma.
In view of the implementation of vaccination for all newborns in Cameroon, we are now assessing the effectiveness of the targeted HBV MTCTP without HBIg in an at-risk population (HbeAg+ mothers). Should there be a significant vaccination failure rate (transmission > 10%), causes (delay in immunization, mother's viral load, HIV co-infection, HBV escape mutants) will be analyzed to determine the possible need for additional measures (administration of IgHBs at birth or antivirals during pregnancy).
Acknowledgments
We thank Christian Aurenche, Director of Tokombéré Hospital, for providing us the opportunity to work in the Health Promotion Center of Tokombéré, which has been recognized by the WHO Sasakawa health prize. We also thank all the pregnant women who participated in this study as well as the sanitary formations of the Tokombéré district for their collaboration.
We also express our thanks to all those who helped us carry out our study particularly in Tokombéré: Jean-Pierre ADOUKARA, François BEAUFILS, Amour BOUNE, Adèle CORTET, Rosette DAMZA, Christiane HURAUX, Sastique KARSIKAM, Amos LITINI, Joseph MANGAVE, RATOUA, Jean-Paul TADA, Raymond TAOWE, Thierry YAOUBA, and in France, Emmanuel GORDIEN, Ségolène BRICHLER and K. L. Erwin for proofreading the English manuscript.
We thank Abbott and DiaSorin for their assistance.
Funding Statement
This study was initiated by the Angers University Hospital with grants from the GEMHEP (Groupe d'étude moléculaire sur les virus des hépatites virales). Number : CA 20.02.2007. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Fattovich G, Bortolotti F, Donato F (2008) Natural history of chronic hepatitis B: special emphasis on disease progression and prognostic factors. J Hepatol 48: 335-352. doi: 10.1016/S0168-8278(08)60894-6. PubMed: 18096267. [DOI] [PubMed] [Google Scholar]
- 2. Kwon SY, Lee CH (2011) Epidemiology and prevention of hepatitis B virus infection. Korean J Hepatol 17: 87-95. doi: 10.3350/kjhep.2011.17.2.87. PubMed: 21757978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Liang TJ (2009) Hepatitis B: the virus and disease. Hepatology.49: S13-S21. doi: 10.1002/hep.22881. PubMed: 19399811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Lok AS, McMahon BJ (2009) Chronic hepatitis B: update 2009. Hepatology.50: 661-662. doi: 10.1002/hep.23190. PubMed: 19714720. [DOI] [PubMed] [Google Scholar]
- 5. WHO (2008) orld Health Organization fact sheet n°204 (2004 Revised August 2008) [Google Scholar]
- 6. Modi AA, Feld JJ (2007) Viral hepatitis and HIV in Africa. AIDS Rev 9: 25-39. PubMed: 17474311. [PubMed] [Google Scholar]
- 7. Franco E, Bagnato B, Marino MG, Meleleo C, Serino L et al. (2012) Hepatitis B: Epidemiology and prevention in developing countries. World. J Hepatol 4: 74-80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Ott JJ, Stevens GA, Groeger J, Wiersma ST (2012) Global epidemiology of hepatitis B virus infection: new estimates of age-specific HBsAg seroprevalence and endemicity. Vaccine.30: 2212-2219. doi: 10.1016/j.vaccine.2011.12.116. PubMed: 22273662. [DOI] [PubMed] [Google Scholar]
- 9. Hennig BJ, Hall AJ (2012) Host genetic factors in hepatitis B infection, liver cancer and vaccination response: A review with a focus on Africa. Sci Total Environ 423: 202-209. doi: 10.1016/j.scitotenv.2010.09.036. PubMed: 20970823. [DOI] [PubMed] [Google Scholar]
- 10. Edmunds WJ, Medley GF, Nokes DJ, Hall AJ, Whittle HC (1993) The influence of age on the development of the hepatitis B carrier state. Proc Biol Sci 253: 197-201. doi: 10.1098/rspb.1993.0102. PubMed: 8397416. [DOI] [PubMed] [Google Scholar]
- 11. Hyams KC (1995) Risks of chronicity following acute hepatitis B virus infection: a review. Clin Infect Dis 20: 992-1000. doi: 10.1093/clinids/20.4.992. PubMed: 7795104. [DOI] [PubMed] [Google Scholar]
- 12. Lee C, Gong Y, Brok J, Boxall EH, Gluud C (2006) Effect of hepatitis B immunisation in newborn infants of mothers positive for hepatitis B surface antigen: systematic review and meta-analysis. Bmj 332: 328-336. doi: 10.1136/bmj.38719.435833.7C. PubMed: 16443611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Echevarría JM, Avellón A (2006) Hepatitis B virus genetic diversity. J Med Virol 78 Suppl 1: S36-S42. doi: 10.1002/jmv.20605. PubMed: 16622876. [DOI] [PubMed] [Google Scholar]
- 14. Kim BK, Revill PA, Ahn SH (2011) HBV genotypes: relevance to natural history, pathogenesis and treatment of chronic hepatitis B. Antivir Ther 16: 1169-1186. doi: 10.3851/IMP1982. PubMed: 22155900. [DOI] [PubMed] [Google Scholar]
- 15. Kramvis A, Kew M, François G (2005) Hepatitis B virus genotypes. Vaccine.23: 2409-2423. doi: 10.1016/j.vaccine.2004.10.045. PubMed: 15752827. [DOI] [PubMed] [Google Scholar]
- 16. Kurbanov F, Tanaka Y, Mizokami M (2010) Geographical and genetic diversity of the human hepatitis B virus. Hepatol Res 40: 14-30. doi: 10.1111/j.1872-034X.2009.00601.x. PubMed: 20156297. [DOI] [PubMed] [Google Scholar]
- 17. Hsieh TH, Liu CJ, Chen DS, Chen PJ (2006) Natural course and treatment of hepatitis D virus infection. J Formos Med Assoc 105: 869-881. doi: 10.1016/S0929-6646(09)60172-8. PubMed: 17098688. [DOI] [PubMed] [Google Scholar]
- 18. Hughes SA, Wedemeyer H, Harrison PM (2011) Hepatitis delta virus. Lancet.378: 73-85. doi: 10.1016/S0140-6736(10)61931-9. PubMed: 21511329. [DOI] [PubMed] [Google Scholar]
- 19. Le Gal F, Gault E, Ripault MP, Serpaggi J, Trinchet JC et al. (2006) Eighth major clade for hepatitis delta virus. Emerg Infect Dis 12: 1447-1450. doi: 10.3201/eid1209.060112. PubMed: 17073101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Radjef N, Gordien E, Ivaniushina V, Gault E, Anaïs P et al. (2004) Molecular phylogenetic analyses indicate a wide and ancient radiation of African hepatitis delta virus, suggesting a deltavirus genus of at least seven major clades. J Virol 78: 2537-2544. doi: 10.1128/JVI.78.5.2537-2544.2004. PubMed: 14963156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Yeh CT, Lai MW (2012) Eliminating hepatitis B virus through neonatal vaccination: can we make it? J Hepatol. 57: 484-485. [DOI] [PubMed] [Google Scholar]
- 22. Villeneuve JP, Durantel D, Durantel S, Westland C, Xiong S et al. (2003) Selection of a hepatitis B virus strain resistant to adefovir in a liver transplantation patient. J Hepatol 39: 1085-1089. doi: 10.1016/j.jhep.2003.09.022. PubMed: 14642631. [DOI] [PubMed] [Google Scholar]
- 23. Friedt M, Gerner P, Lausch E, Trübel H, Zabel B et al. (1999) Mutations in the basic core promotor and the precore region of hepatitis B virus and their selection in children with fulminant and chronic hepatitis B. Hepatology.29: 1252-1258. doi: 10.1002/hep.510290418. PubMed: 10094972. [DOI] [PubMed] [Google Scholar]
- 24. Ivaniushina V, Radjef N, Alexeeva M, Gault E, Semenov S et al. (2001) Hepatitis delta virus genotypes I and II cocirculate in an endemic area of Yakutia, Russia. J Gen Virol 82: 2709-2718. PubMed: 11602783. [DOI] [PubMed] [Google Scholar]
- 25. Birguel J, Ndong JG, Akhavan S, Moreau G, Sobnangou JJ et al. (2011) [Viral markers of hepatitis B, C and D and HB vaccination status of a health care team in a rural district of Cameroon]. Med Trop (Mars).71: 201-202. [PubMed] [Google Scholar]
- 26. Bottero J, Boyd A, Gozlan J, Lemoine M, Carrat F et al. (2013) Performance of rapid tests for detection of HBsAg and anti-HBsAb in a large cohort, France. J Hepatol 58: 473-478. doi: 10.1016/S0168-8278(13)61164-2. PubMed: 23183527. [DOI] [PubMed] [Google Scholar]
- 27. Abdou Chekaraou M, Brichler S, Mansour W, Le Gal F, Garba A et al. (2010) A novel hepatitis B virus (HBV) subgenotype D (D8) strain, resulting from recombination between genotypes D and E, is circulating in Niger along with HBV/E strains. J Gen Virol 91: 1609-1620. doi: 10.1099/vir.0.018127-0. PubMed: 20147517. [DOI] [PubMed] [Google Scholar]
- 28. Mansour W, Malick FZ, Sidiya A, Ishagh E, Chekaraou MA et al. (2012) Prevalence, risk factors, and molecular epidemiology of hepatitis B and hepatitis delta virus in pregnant women and in patients in Mauritania. J Med Virol 84: 1186-1198. doi: 10.1002/jmv.23336. PubMed: 22711346. [DOI] [PubMed] [Google Scholar]
- 29. Kfutwah AK, Tejiokem MC, Njouom R (2012) A low proportion of HBeAg among HBsAg-positive pregnant women with known HIV status could suggest low perinatal transmission of HBV in Cameroon. Virol J 9: 62. doi: 10.1186/1743-422X-9-62. PubMed: 22401702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Foupouapouognigni Y, Noah DN, Sartre MT, Njouom R (2011) High prevalence and predominance of hepatitis delta virus genotype 1 infection in Cameroon. J Clin Microbiol 49: 1162-1164. doi: 10.1128/JCM.01822-10. PubMed: 21209162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Mbanya DN, Takam D, Ndumbe PM (2003) Serological findings amongst first-time blood donors in Yaoundé, Cameroon: is safe donation a reality or a myth? Transfus Med 13: 267-273. doi: 10.1046/j.1365-3148.2003.00453.x. PubMed: 14617337. [DOI] [PubMed] [Google Scholar]
- 32. Mansour W, Bollahi MA, Hamed CT, Brichler S, Le Gal F et al. (2012) Virological and epidemiological features of hepatitis delta infection among blood donors in Nouakchott, Mauritania. J Clin Virol 55: 12-16. doi: 10.1016/j.jcv.2012.05.011. PubMed: 22704272. [DOI] [PubMed] [Google Scholar]
- 33. Candotti D, Danso K, Allain JP (2007) Maternofetal transmission of hepatitis B virus genotype E in Ghana, west Africa. J Gen Virol 88: 2686-2695. doi: 10.1099/vir.0.83102-0. PubMed: 17872520. [DOI] [PubMed] [Google Scholar]
- 34. Makuwa M, Caron M, Souquière S, Malonga-Mouelet G, Mahé A et al. (2008) Prevalence and genetic diversity of hepatitis B and delta viruses in pregnant women in Gabon: molecular evidence that hepatitis delta virus clade 8 originates from and is endemic in central Africa. J Clin Microbiol 46: 754-756. doi: 10.1128/JCM.02142-07. PubMed: 18077651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Ampofo W, Nii-Trebi N, Ansah J, Abe K, Naito H et al. (2002) Prevalence of blood-borne infectious diseases in blood donors in Ghana. J Clin Microbiol 40: 3523-3525. doi: 10.1128/JCM.40.9.3523-3525.2002. PubMed: 12202610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Touré-Fall AO, Dièye TN, Sall A, Diop M, Seck M et al. (2009) [Residual risk of transmission of HIV and HBV, in Senegalese national blood bank from 2003 to 2005]. Transfus Clin Biol 16: 439-443. doi: 10.1016/j.tracli.2009.09.005. PubMed: 19926508. [DOI] [PubMed] [Google Scholar]
- 37. Buseri FI, Muhibi MA, Jeremiah A (2009) Sero-epidemiology of transfusion-transmissible infectious diseases among blood donors in Osogbo, south-west Nigeria. Blood Transfus 7: 293-299. PubMed: 20011640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Acquaye JK, Mingle JA (1994) Hepatitis B viral markers in Ghanaian pregnant women. West Afr J Med 13: 134-137. PubMed: 7841099. [PubMed] [Google Scholar]
- 39. Emechebe GO, Ikefuna AN, Ilechukwu GC, Igwe WC, Ejiofor OS et al. (2009) Hepatitis B virus infection in Nigeria - A review Nigerian Med J.50: 18-22. [Google Scholar]
- 40. Ott JJ, Stevens GA, Wiersma ST. (2012) The risk of perinatal hepatitis B virus transmission: hepatitis B e antigen (HBeAg) prevalence estimates for all world regions. BMC Infect Dis 12: 131. doi: 10.1186/1471-2334-12-131. PubMed: 22682147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Kurbanov F, Tanaka Y, Fujiwara K, Sugauchi F, Mbanya D et al. (2005) A new subtype (subgenotype) Ac (A3) of hepatitis B virus and recombination between genotypes A and E in Cameroon. J Gen Virol.86: 2047-2056. [DOI] [PubMed] [Google Scholar]
- 42. Olinger CM, Venard V, Njayou M, Oyefolu AO, Maïga I et al. (2006) Phylogenetic analysis of the precore/core gene of hepatitis B virus genotypes E and A in West Africa: new subtypes, mixed infections and recombinations. J Gen Virol 87: 1163-1173. doi: 10.1099/vir.0.81614-0. PubMed: 16603517. [DOI] [PubMed] [Google Scholar]
- 43. Kramvis A, Kew MC (2007) Epidemiology of hepatitis B virus in Africa, its genotypes and clinical associations of genotypes. Hepatol Res 37: S9-S19. doi: 10.1111/j.1872-034X.2007.00098.x. PubMed: 17627641. [DOI] [PubMed] [Google Scholar]
- 44. Garmiri P, Loua A, Haba N, Candotti D, Allain JP (2009) Deletions and recombinations in the core region of hepatitis B virus genotype E strains from asymptomatic blood donors in Guinea, west Africa. J Gen Virol 90: 2442-2451. doi: 10.1099/vir.0.012013-0. PubMed: 19535503. [DOI] [PubMed] [Google Scholar]
- 45. Fujiwara K, Tanaka Y, Orito E, Ohno T, Kato T et al. (2005) Distribution of HBV genotypes among HBV carriers in Benin:phylogenetic analysis and virological characteristics of HBV genotype E. World J Gastroenterol 11: 6410-6415. PubMed: 16425408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Liu C, Gao H, Zhai S, Liu B. (2011) Molecular characterization, chromosomal localization, expression profile and association analysis with carcass traits of the porcine dickkopf homolog1 gene. Mol Biol Rep 38: 1929-1934. doi: 10.1007/s11033-010-0313-x. PubMed: 20848216. [DOI] [PubMed] [Google Scholar]
- 47. Poustchi H, Mohamadkhani A, Bowden S, Montazeri G, Ayres A et al. (2008) Clinical significance of precore and core promoter mutations in genotype D hepatitis B-related chronic liver disease. J Viral Hepat 15: 753-760. doi: 10.1111/j.1365-2893.2008.00998.x. PubMed: 18507754. [DOI] [PubMed] [Google Scholar]
- 48. Bekondi C, Olinger CM, Boua N, Talarmin A, Muller CP et al. (2007) Central African Republic is part of the West-African hepatitis B virus genotype E crescent. J Clin Virol 40: 31-37. doi: 10.1016/j.jcv.2007.05.009. PubMed: 17689139. [DOI] [PubMed] [Google Scholar]
- 49. Ragheb M, Elkady A, Tanaka Y, Murakami S, Attia FM et al. (2012) Multiple intra-familial transmission patterns of hepatitis B virus genotype D in north-eastern Egypt. J Med Virol 84: 587-595. doi: 10.1002/jmv.23234. PubMed: 22337297. [DOI] [PubMed] [Google Scholar]