

Abstract
Human amniotic membrane is a standard substratum used to culture limbal epithelial stem cells for transplantation to patients with limbal stem cell deficiency. Various methods were developed to decellularize amniotic membrane, because denuded membrane is poorly immunogenic and better supports repopulation by dissociated limbal epithelial cells. Amniotic membrane denuding usually involves treatment with EDTA and/or proteolytic enzymes; in many cases additional mechanical scraping is required. Although ensuring limbal cell proliferation, these methods are not standardized, require relatively long treatment times and can result in membrane damage. We propose to use 0.5 M NaOH to reliably remove amniotic cells from the membrane. This method was used before to lyse cells for DNA isolation and radioactivity counting. Gently rubbing a cotton swab soaked in NaOH over the epithelial side of amniotic membrane leads to nearly complete and easy removal of adherent cells in less than a minute. The denuded membrane is subsequently washed in a neutral buffer. Cell removal was more thorough and uniform than with EDTA, or EDTA plus mechanical scraping with an electric toothbrush, or n-heptanol plus EDTA treatment. NaOH-denuded amniotic membrane did not show any perforations compared with mechanical or thermolysin denuding, and showed excellent preservation of immunoreactivity for major basement membrane components including laminin α2, γ1-γ3 chains, α1/α2 and α6 type IV collagen chains, fibronectin, nidogen-2, and perlecan. Sodium hydroxide treatment was efficient with fresh or cryopreserved (10% dimethyl sulfoxide or 50% glycerol) amniotic membrane. The latter method is a common way of membrane storage for subsequent grafting in the European Union. NaOH-denuded amniotic membrane supported growth of human limbal epithelial cells, immortalized corneal epithelial cells, and induced pluripotent stem cells. This simple, fast and reliable method can be used to standardize decellularized amniotic membrane preparations for expansion of limbal stem cells in vitro before transplantation to patients.
Introduction
Limbal epithelial stem cells (LESC) in the human cornea reside at its periphery known as the corneoscleral limbus and continuously renew the corneal epithelium [1]–[3]. In some conditions these cells degenerate and/or die, leading to limbal epithelial stem cell deficiency (LSCD). It is a fairly common and clinically important cause of corneal blindness. LSCD may develop as a consequence of congenital aniridia, Stevens-Johnson syndrome, chemical and thermal burns (including sulfur mustard gas poisoning in war conditions), ocular cicatricial pemphigoid, chronic inflammation, and microbial infections [4]–[6]. Ocular burns that frequently lead to LSCD comprise up to 18% of all eye injuries [7]. More recently, an increasing number of long-term contact lens wearers have been also diagnosed with LSCD [8]. LSCD results in corneal erosions and vascularization, conjunctival ingrowth (conjunctivalization), and scarring, causing compromised corneal transparency and gradual vision loss [4], [5]. This condition may be hard to treat especially in cases of total stem cell deficiency [4].
Transplantation of LESC cultured on human amniotic membrane (HAM) or fibrin to the affected limbal area has emerged as a promising approach to manage LSCD since the pioneering work of Tseng’s group [1], [2], [9], [10]. Although the procedure is not fully standardized [11] and allograft survival is low, even with immunosuppression, an average 1–3 years success rate of up to 76% was reported [1], [2], [9].
HAM continues to be the most popular substratum for LESC to expand and then use for transplantation purposes in LSCD patients. It is readily available from placenta discarded in delivery rooms, and has been a more successful alternative to previous methods, with hundreds of patients having received culture-expanded LESC transplants [12], [13]. Amniotic cell basement membrane (BM) is largely (but not completely) similar to limbal BM in composition [14], [15]. HAM contains important growth factors [16]–[18], is anti-bacterial, anti-angiogenic, only very weakly immunogenic [19]–[21], and improves wound healing [20], [22], [23]. Although LESC can differentiate on HAM [24], it is still considered the best substratum for their expansion, also allowing secure placement onto the patient’s cornea [11], [13].
HAM has been generally used for clinical purposes as intact (cryopreserved, lyophilized or dry) and denuded, after amniotic epithelial cells removal for better LESC adhesion. Intact HAM mostly supports the growth of limbal explants [24]–[26], whereas denuded HAM can be used as good scaffold for enzymatically dispersed LESC [16], [18], [27]–[30]. Denuded HAM better supports LESC proliferation, shows an increased preservation of clonogenicity, and is less immunogenic [16], [18], [20], [25], [26], [30], [31]. Various methods have been used to denude HAM including treatment with dispase, thermolysin, trypsin, EDTA, sodium dodecyl sulfate (SDS), ammonia or urea [16], [18], [27], [32]–[38]. Most such treatments last for hours, are tedious, may damage HAM or remove some components, fail to uniformly remove the epithelium, require additional scraping, and are not very reproducible [18], . The existing treatments also affect the stroma, which may result in the partial removal of HAM’s growth factors [18]. Realizing the need for a better approach, we took advantage of a long-known ability of alkaline solutions to dissolve cells [41], [42] and applied mild solutions of sodium hydroxide only to the amniotic epithelial cells for less than a minute as a fast and reproducible HAM denuding agent. The procedure was efficient in thoroughly removing amniotic epithelial cells from HAM. This fast and reproducible method using an inorganic agent could significantly streamline and standardize denuded HAM preparation for LESC culture.
Results
The successful removal of amniotic epithelial cells from HAM with NaOH was obtained independently in two laboratories: one at Cedars-Sinai Medical Center (Los Angeles) and another at Charles University (Prague). Gentle rubbing of HAM with 0.25–0.5 M NaOH-soaked cotton tip for less than one minute removed most of the cells, as shown on Fig. 1. To better visualize the debridement, HAM was stained with Trypan blue (TB) that is excluded by live cells. Cryopreservation with glycerol effectively kills amniotic epithelial cells [43], resulting in a membrane that is entirely stained blue. Denuding the right half of the HAM piece with NaOH effectively removed the cells as evidenced by the loss of blue color (Fig. 1). At higher magnification, HAM presented a monolayer of tightly packed epithelial cells, all stained by TB (Fig. 2A,B). Rubbing it with a cotton tip soaked in PBS removed cells only partially, whereas rubbing with NaOH-soaked tip efficiently removed all cells judged by uniform loss of TB-stained material (Fig. 2C–E).
Figure 1. Glycerol-cryopreserved HAM after thawing, Trypan blue staining and treatment.

A, mesh Sanatyl 20 support for HAM in a Petri dish. B, HAM on Sanatyl 20 after TB staining (the whole membrane is stained as glycerol kills the cells). C, the right half of HAM treated with NaOH (staining is significantly lighter than on untreated left half). D, the same HAM without Sanatyl 20 support; note lack of TB staining (cell removal) on the most part of the treated right half. Low-magnification pictures are presented.
Figure 2. Cell removal by NaOH from cryopreserved HAM.

A, phase contrast of cryopreserved and thawed out HAM showing a monolayer of amniotic epithelial cells. B, the same HAM stained with TB; all cells are stained. C, cotton swab rubbing of HAM leaves many amniotic epithelial cells behind. D, after 30 sec rub with 0.5 M NaOH-soaked cotton swab no cells are left. E, NaOH effectively removes cells from HAM (gray line shows the boundary of the rubbed zone). Bar in A–D = 50 µm; in E, bar = 100 µm.
It was important to compare NaOH method with several others described before in terms of HAM integrity and expression of major BM components identified by immunofluorescent staining of 1% formalin-fixed cryostat sections. The BM components were chosen from those predominantly expressed in the limbal BM, such as laminin chains γ3 (Fig. 3A), α2, β2 (data not shown here), and collagen type IV α1/α2 chains (Fig. 3B), and those also expressed in central corneal BM, such as laminin γ2 chain (Fig. 4A), nidogen-2 (Fig. 4B), fibronectin (Fig. 5A), perlecan (Fig. 5B), laminin β1 and γ1 chains, and collagen type IV α6 chain (data not shown here). Of these components, laminin β1 and β2 chains could not be detected, whereas all others yielded a positive (although sometimes variable staining (e.g. for laminin α2 chain). As shown in Figs. 3–5, EDTA treatment did not compromise HAM integrity judged by continuous staining for all tested BM components. However, although this treatment removed some cells (single long arrows), many cells were generally left behind and persisted upon gentle scraping. More vigorous scraping with low-speed electric toothbrush caused local ruptures of HAM (double arrows on Figs. 3, 4, 5B). Additional rubbing with n-heptanol-soaked cotton applicator was helpful but still some cells remained. Brief thermolysin treatment efficiently removed most or all cells (few remaining ones are marked with, but HAM became fragile and was easily fragmented during O.C.T. embedding.
Figure 3. Laminin γ3 chain and type IV collagen α1/α2 chain expression in intact HAM and after various treatments.

Laminin γ3 (A) and type IV collagen α1/α2 (B) are found in the limbal epithelial BM but not in the central corneal BM. Here and below, single long arrows show denuded parts with no cells (DAPI-counterstained nuclei). Double arrows show places where HAM is disrupted after scraping the membrane with electric toothbrush on low speed. EDTA together with rubbing or n-heptanol leaves a number of epithelial cells still attached to HAM. In contrast, thermolysin and NaOH leave little (single short arrows) to no epithelium on the treated HAM. Except for EDTA+toothbrush scraping, all treatments preserve normal continuous staining patterns of both BM components. Immunohistochemical staining of O.C.T.-embedded and sectioned HAM. Bar = 20 µm.
Figure 4. Laminin γ2 chain and nidogen-2 expression in intact HAM and after various treatments.

Laminin γ2 (A) and nidogen-2 (B) are also expressed in central and limbal epithelial BM. The effects of various treatments are similar to the ones shown on Fig. 3. EDTA treatment results in incomplete cell removal or even HAM damage (double arrows) after extensive scraping. Thermolysin treatment removes cells well but shows some local irregular staining for proteolysis-sensitive laminin γ2 chain (A). NaOH produces HAM virtually devoid of epithelial cells with continuous staining for both BM components. Immunohistochemical staining of O.C.T.-embedded and sectioned HAM. Bar = 20 µm.
Figure 5. Fibronectin and perlecan expression in intact HAM and after various treatments.

The results on fibronectin (A) and perlecan (B) are similar to other BM components. Only thermolysin and NaOH completely remove amniotic epithelium. Immunohistochemical staining of O.C.T.-embedded and sectioned HAM. Bar = 20 µm.
NaOH treatment was the only one that efficiently removed amniotic epithelium from HAM with only occasional cells remaining (short arrows; Figs. 3B and 4B), while at the same time preserving continuous staining for all BM components (Figs. 3–5). The negative controls showed that this staining was not an artifact due to the known “edge effect” (Fig. 6).
Figure 6. Negative control with omission of primary antibody.
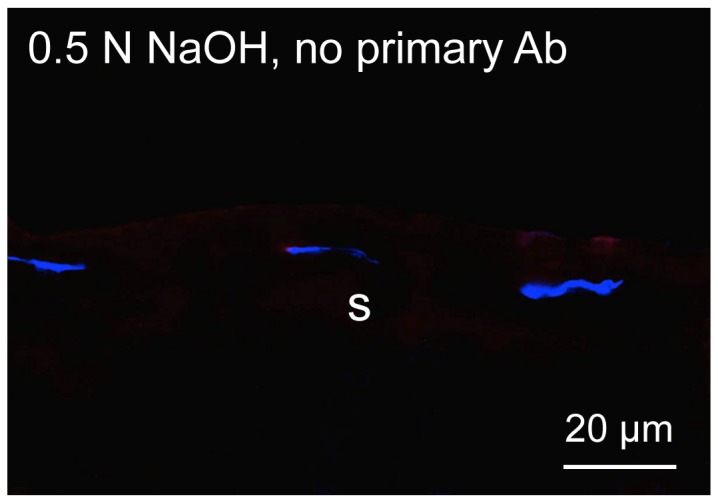
Immunohistochemical staining of OCT-embedded and sectioned HAM treated with NaOH. No staining is seen apart from DAPI nuclear staining of HAM stromal cells. Bar = 20 µm.
In initial experiments, we used 0.5 M solution for less than 30 sec rubbing that was very efficient in removing cells. Later, we also tried lower concentrations that were efficient down to 0.25 M; however, additional gentle rubbing was usually needed.
NaOH-denuded HAM was further used as culture substratum for human cells including telomerase-immortalized corneal epithelial cells (tHCEC), primary LESC, LESC-derived induced pluripotent stem cells (iPSC) or skin fibroblast-derived iPSC. NaOH-treated HAM supported proliferation of epithelial cells and iPSC well (Fig. 7). Cells formed confluent cobblestone monolayers within 1–2 weeks. The expression of some putative LESC markers was further evaluated in primary human LESC cultured on NaOH-denuded HAM. LESC expressed PAX6, keratins 14 and 15, as well as ΔNp63 (Fig. 8), similar to their expression reported in cultured cells and ex vivo corneas [26], [30], [44].
Figure 7. Phase contrast of various cells grown on NaOH-denuded HAM.

Note cobblestone monolayer of telomerase-immortalized corneal epithelial cells (tHCEC), LESC, and LESC-derived iPSC. Fibroblast-derived iPSC also show good spreading of cells and growth. Bar = 40 µm.
Figure 8. Marker analysis of LESC-enriched cultures grown on top of NaOH-denuded HAM.

Note expected positive staining for putative LESC markers PAX6, K14 (A, B; double label), K15, and ΔNp63 (C, D; double label). Immunohistochemical staining of O.C.T.-embedded and sectioned HAM. Bar = 20 µm.
Discussion
Since the initial successful graftings performed in 1960–1970, non-immunogenic HAM has been widely used as a biodegradable patch in corneal surgery and as natural scaffold for culturing LESC as a transplant for treatment of LSCD [10], [32], [45], [46]. The epithelium tightly covering HAM is a poorly adhesive scaffold for most if not all epithelial cells. If amniotic epithelial cells are not removed, LESC monolayers cannot be easily established on HAM. Only limbal explants can develop substantial outgrowths on intact HAM, possibly by displacing amniotic epithelial cells. Cryopreservation of HAM with a mild fixative glycerol that is allowed in the European Union [47] kills amniotic cells [43] allowing for an easier growth of explanted LESC-enriched limbal fragments. However, when compared to denuded HAM, intact membrane supports growth of cultured LESC less well and promotes terminal differentiation of these cells especially when the cultures are air-lifted [31], [48]. Therefore, it is preferable to use decellularized HAM in order to obtain LESC sheets best suitable for transplantation. At the same time, it may be important to keep stromal cells out of contact with denuding agent as much as possible, as they secrete factors needed to promote cell proliferation and wound healing [16], [17].
All existing techniques for denuding HAM with EDTA, dispase, trypsin, thermolysin, NH4OH, SDS or urea [18], [27], [31], [36], [38], [39], [49]–[52] require soaking HAM in cell stripping solutions, from 15 min to 24 hrs, which may impact both its epithelial and stromal compartments. Additionally some drawbacks have been associated with most denuding methods, necessitating further standardization of this important procedure. For instance, dispase routinely used to isolate LESC, and to a certain extent trypsin cause HAM destruction [18], [27], [39]. EDTA even with overnight incubation does not remove amniotic epithelium completely and additional scraping is required [27], [29]. This was also confirmed in our experiments, and EDTA was even reported to cause some damage to HAM [39]. Applying an electric toothbrush at low speed after EDTA resulted in HAM tears in our study (Figs. 3–5). Additional post-EDTA brief rubbing with n-heptanol used to remove corneal epithelial cells for wound healing studies [53] still left some epithelial cells attached. Thermolysin and 5 M urea denude HAM well but some destruction of laminin was noted [18], [39]. In our experiments, thermolysin efficiently removed amniotic epithelial cells but the membranes became fragile and hard to handle. However, after cryopreservation with a mild fixative glycerol, thermolysin was reported to cause no damage to HAM [40]. SDS procedure is very long (up to 48 hrs), involves nuclease treatment and the preservation of HAM components has not been analyzed [16], [30], [36].
Therefore, we set out to design a fast and reliable method for denuding HAM that would not require the incubation of the whole membrane in the decellularizing media and would allow for complete removal of the denuding agent. Mild alkali that are easy to neutralize and can readily solubilize cells [41], [42] could fit these tasks well. Previously, 10–15 min soaking in 0.1%–10% ammonium hydroxide with additional scraping has already been used to denude HAM [33], [49]–[52] for cell culture but this method has been largely abandoned. Consequently, we revisited the old alkaline cell solubilization technique applying faster acting NaOH rather than NH4OH to denude HAM. The advantages of our method are its speed, efficiency, and apparent lack of direct contact of cell stripping solution with the HAM stroma. The denuding agent is inorganic and is easily and immediately neutralized in PBS yielding just salt, which may make it attractive for regulatory agencies. Another important aspect of the NaOH method is that it is very efficient as well on HAM cryopreserved in glycerol (Figs. 1, 2), which is an officially recommended cryopreservative for storage of HAM for grafting in the European Union [47]. In this context, had dispase or thermolysin been effective denuding agents, they could represent regulatory issues, since the residual activity of these proteinases may not be easily neutralized.
Using gentle rubbing of HAM with 0.25–0.5 M NaOH-soaked cotton tip, we were able to nearly completely denude fresh or DMSO-cryopreserved membranes within 1 min without any apparent damage to the BM. In some instances a slightly longer time was needed to remove all cells, which may be due to physico-chemical heterogeneity of HAM depending on the distance from placental disc [54]. Additionally, since HAM is human donor material, some donor-to-donor variability could be expected to marginally affect the denuding time.
Staining patterns of the major HAM components were further examined by immunostaining. Generally, the composition of HAM resembled that of limbal epithelial BM, with some differences. Both structures showed continuous staining for laminin α2 and γ3 chains, and type IV collagen α1/α2 chains that are not revealed in the central corneal epithelial BM [55]–[57], as well as for other components present in both central corneal and limbal epithelial BM, such as type IV collagen α6 chain, laminin γ1 and γ2 chains, nidogen-2, perlecan, and fibronectin (see Figs. 3–5). Contrary to a recent study [17], we were unable to obtain positive HAM staining for laminin β1 and β2 chains, but did find laminin α2 and γ3 chains. These discrepancies could arise from different antibodies and fixation protocols used.
Various treatments tested did not show any significant alterations in the patterns of HAM components. In particular, after NaOH denuding, there was no change in continuity and strength of staining for any component compared to the untreated membranes (Figs. 3–5). One can conclude that brief exposure of the epithelial side of HAM to mild NaOH solutions did not adversely affect its structure. Further proof of NaOH usability was obtained upon seeding of various cells on denuded HAM. All cell types including primary LESC and iPSC grew well on NaOH-denuded HAM and formed monolayers in one-two weeks. Over more than a year, no batch of NaOH-treated HAM showed any adverse effect on cell spreading or growth. Primary limbal cultures on this denuded HAM were positive for several putative LESC markers (Fig. 8) with patterns typical for these cells.
Overall, the new method of denuding HAM by brief exposure of its epithelial side to mild sodium hydroxide solution proved to be fast, easy, and reproducible. NaOH decellularization does not alter normal patterns of major HAM components and supports growth of various cell types including LESC. The NaOH treatment also works well on glycerol-cryopreserved HAM, where other methods failed to provide efficient deepithelialization. Because of these advantages, this method may be easily standardized and could become a treatment of choice for preparing HAM in order to propagate LESC under good manufacturing practice (GMP) protocol for clinical transplantation treatment of patients with LSCD.
Materials and Methods
Ethics Statement
Human placentas were obtained, after written informed consent from the prospective mothers, upon their Cesarean-section deliveries (approved IRB protocol Pro00019230, part of IRB protocol #9313 for Cedars-Sinai Medical Center, and approved by State Institute for Drug Controls for Charles University). Primary LESC cultures were obtained from donor corneoscleral rims discarded after penetrating keratoplasty. Work with these rims including culturing LESC and iPSC generation was fully covered by an approved Cedars-Sinai Medical Center IRB protocol Pro00019393.
Preparation of Denuded Human Amniotic Membrane (HAM)
The placental tissue was placed in low glucose Dulbecco’s Modified Eagle’s medium (DMEM; Invitrogen/Life Technologies, Carlsbad, CA, cat# 11885-084 or Sigma-Aldrich, St. Louis, MO, cat# D5546) with antibiotic/antimycotic solution (Invitrogen, cat# 15240062 or Sigma-Aldrich, cat# A5955). The placenta was washed first with Hanks’ balanced salt solution (HBSS, Life Technologies, cat# 24020117) or phosphate-buffered saline (PBS; Life Technologies, cat# 10010049) to remove excess blood. Then the amniotic membrane was mechanically separated from the chorion with forceps and washed three times with HBSS or PBS to allow the spongy stromal side layer to swell, which was then removed. The HAM was cut into 5×5 cm and cryopreserved in PBS with 10% dimethyl sulfoxide (DMSO; Sigma-Aldrich, cat# 2438). In some experiments, HAM was cryopreserved with 50% glycerol, which is a mild fixative [58] and a common cryoprotectant approved in the European Union for HAM cryopreservation [47]. In these cases HAM (3×3 cm) was placed on Sanatyl 20 support (warp knitted 100% polyester fiber bandage, Tylex, Letovice, Czech Republic) and immersed in 50% DMEM with antibiotic/antimycotic solution, 10% fetal bovine serum (Life Technologies, cat# 26140079), 0.38% sodium bicarbonate (Life Technologies, cat# 25080094), and 50% glycerol (Sigma-Aldrich, cat# G2025). Cryopreserved HAM was stored at −80°C until use. After thawing and sequential washing in PBS for 10–30 min at room temperature to remove DMSO or glycerol, HAM was de-epithelialized using different methods for comparison as follows:
EDTA treatment. HAM was incubated in 0.02% EDTA (Sigma-Aldrich, cat# E-5134) in calcium-magnesium free PBS at 37°C for one hour to loosen amniotic epithelial cells, and then transferred into PBS. Treatment of membranes was followed by immediately washing twice for 15 min in PBS to remove cellular debris.
EDTA-scraping treatment. EDTA-treated HAM was subjected to gentle mechanical scraping with an electric toothbrush on low speed and then washed twice for 15 min in PBS to remove cellular debris.
EDTA-heptanol treatment. EDTA-treated HAM was placed in CellCrown™ inserts (Sigma-Aldrich, cat#Z681792 for 6 well plate insert or cat# Z681830 for 12 well plate insert) with epithelial side facing up and was rubbed for 1–2 min with cotton-tipped applicators (Cardinal Health, Dublin, OH, cat# C15053-006) soaked in n-heptanol (1-heptanol, Sigma-Aldrich, cat# 72954-5ML-F). HAM was then washed twice for 15 min in PBS.
Thermolysin treatment. HAM was incubated in a 125 µg/ml thermolysin (Sigma-Aldrich, cat# T7902) solution in PBS for 9 min at 37°C as described previously [39], [40]. Treated membranes were subjected to gentle mechanical scraping and then rinsed and washed three times for 15 min in PBS to remove cellular debris or immediately rinsed and washed three times for 15 min in PBS with shaking to remove cellular debris.
NaOH treatment. HAM placed in CellCrown™ inserts (Sigma-Aldrich) with epithelial side facing up was de-epithelialized by rubbing with cotton-tipped applicator soaked in 0.5 M NaOH (Sigma-Aldrich, cat# S5881) for 20–30 seconds and followed by immediate 10–15 min washing in HBSS or PBS. In later experiments, 0.25 M NaOH solution was used with additional gentle rubbing using a cotton-tipped applicator.
HAM pictures were taken with Olympus CKX41 inverted microscope equipped with an Olympus C-3040 camera (Olympus, Tokyo, Japan). In some experiments, glycerol-cryopreserved cells were stained with 0.2% TB (Sigma-Aldrich, cat# T6146) in PBS for 90 sec, followed by rinsing 3–5 times in PBS. Because cells are dead after cryopreservation in glycerol [43], they all stained with TB, which made it easy to monitor their further removal.
Immunostaining
Different types of denuded HAMs from 2–4 independent treatment experiments were embedded for immunocytochemistry in Tissue-Tek® O.C.T. compound (VWR, Visalia, CA, cat# 25608-930). Well-characterized antibodies to BM components (Table 1) and putative LESC markers were used for indirect immunofluorescence as described [44], [56].
Table 1. Antibodies used in the study.
| Antigen | Antibody | Source | Dilution |
| Fibronectin | Mouse mAb 568 | Ref. [56] | 1∶60 |
| α1/α2 type IV collagen | Mouse mAb M3F7 | Developmental Hybridoma Bank | straight |
| α6 type IV collagen | Rat mAb H63 | Ref. [56] | straight |
| Laminin α2 | Mouse mAb 1F9 | Ref. [56] | straight |
| Laminin β1 | Rat mAb LT3 | Ref. [56] | straight |
| Laminin β2 | Mouse mAb C4 | Developmental Hybridoma Bank | straight |
| Laminin γ1 | Rat mAb A5 | Ref. [56] | straight |
| Laminin γ2 | Mouse mAb MAB19562 | Millipore | 1∶50 |
| Laminin γ3 | Rabbit pAb R96 | Ref. [44] | 1∶200 |
| Perlecan | Rat mAb C11L6 | Ref. [64] | straight |
| Nidogen-2 | Rabbit pAb 1080 | Ref. [56] | 1∶200 |
| Keratin 14 | Mouse mAb sc-53253 | Santa Cruz Biotechnology | 1∶10 |
| Keratin 15 | Mouse mAb sc-47697 | Santa Cruz Biotechnology | 1∶10 |
| ΔNp63 | Goat pAb sc-8609 | Santa Cruz Biotechnology | 1∶20 |
| PAX6 | Rabbit pAb PRB-278P | Covance | 1∶100 |
mAb, monoclonal antibody; pAb, polyclonal antibody.
Cell Culture on NaOH-denuded HAM
NaOH-treated HAM was used to culture telomerase-immortalized human corneal epithelial cell line (tHCEC), primary human LESC or iPS cells derived from human LESC or skin fibroblasts. Primary LESC-enriched cultures were prepared from discard corneoscleral rims from healthy donors, obtained from collaborating surgeons under an approved IRB protocol CR00004366. Cells were isolated by the dispase method. Corneoscleral rims with conjunctiva cut out with scissors were treated with 2.4 U/mL Dispase II (Roche Applied Science, Indianapolis, IN, cat# 04942078001) in keratinocyte serum-free medium (KSFM; Life Technologies, cat# 17005042) supplemented with 10% FBS at 37°C for 2 hrs. [59]. The limbal epithelial cells eased off the rims were dissociated with 0.25% trypsin (Life Technologies, cat# 15050065) for 30 min at room temperature. Cells were first seeded on a mixture of human basement membrane proteins including fibronectin (BD Biosciences, San Jose, CA, cat# 354008), laminin (Sigma-Aldrich, cat# L4445), and type IV collagen (Sigma-Aldrich, cat# C6745-1ML), at 0.5–1 µg/cm2, after [60] in KSFM medium. Cells formed confluent monolayers and stained positive for putative LESC markers including PAX6, K14, K15, K17, and ΔNp63. Then they were passaged onto HAM using trypsin and grown in the same medium. LESC-derived induced pluripotent stem cells were obtained using Yamanaka’s non-integrating oriP/EBNA1 (Epstein-Barr nuclear antigen-1)-based episomal plasmid vectors [61], [62] from Addgene (Cambridge, MA). They were first cultured in mTeSRTM1 medium (StemCell Technologies, Vancouver, BC, Canada, cat# 05850), gradually changed to LESC Epilife® medium (Life Technologies, cat# MEPI500CA) with a defined growth supplement (Life Technologies, B-27, cat# 17504044 and N-2, cat# 17502048), antibiotic-antimycotic mixture, and human keratinocyte growth supplement (HKGS, cat# S0015; Invitrogen): bovine pituitary extract, 0.2% v/v; insulin, 5 µg/ml; hydrocortisone, 0.18 µg/ml; transferrin, 5 µg/ml; human epidermal growth factor, 0.2 ng/ml. Non-tumorigenic diploid human corneal epithelial cells immortalized by telomerase gene [63] were cultured in EpiLife® medium (Life Technologies) with human corneal growth supplement (HCGS, Invitrogen, cat# S0095) on type IV collagen-coated plates and passaged using 0.05% trypsin (Life Technologies). These cells are somewhat similar to LESC and express putative LESC markers K14, K17, K19, ABCG2, but not K15 or the differentiated corneal keratin K3 (data not shown here). All cultures were incubated at 37°C in a humidified atmosphere containing 5% CO2.
Acknowledgments
The authors are grateful for the support of Dr. Leslie Raffel, Director, The Winnick Family General Clinical Research Center (GCRC), and Associate Director, Common Disease Genetics Program at Cedars-Sinai Medical Center. Dr. Raffel is Principal Investigator on the Cedars-Sinai Medical Center IRB protocol #9313, which made it possible to collect and use human placentas for this research. We also thank GCRC staff for their constant help. Antibodies against laminin β2 chain produced by Dr. Joshua R. Sanes and to type IV collagen α1/α2 chains produced by Dr. Heinz Furthmayr were obtained from the Developmental Studies Hybridoma Bank (DSHB), Department of Biology, University of Iowa (Iowa City, Iowa, United States of America), under contract N01-HD-2-3144 from the NICHD.
Presented in part at the Annual Meeting of the Association for Research In Vision and Ophthalmology, Seattle, WA, USA, May 2013.
Funding Statement
NIH EY13431, EY023429, CTSI grant UL 1RR033176, Cedars-Sinai Regenerative Medicine Institute grants, and PRVOUK-P24/LF1/3 Institutional support program of the Charles University, Prague. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Rama P, Matuska S, Paganoni G, Spinelli A, De Luca M, et al. (2010) Limbal stem-cell therapy and long-term corneal regeneration. N Engl J Med 363: 147–155. [DOI] [PubMed] [Google Scholar]
- 2. Ahmad S, Kolli S, Lako M, Figueiredo F, Daniels JT (2010) Stem cell therapies for ocular surface disease. Drug Discov Today 15: 306–313. [DOI] [PubMed] [Google Scholar]
- 3. Ordonez P, Di Girolamo N (2012) Limbal epithelial stem cells: role of the niche microenvironment. Stem Cells 30: 100–107. [DOI] [PubMed] [Google Scholar]
- 4. Biber JM, Holland EJ, Neff KD (2010) Management of ocular stem cell disease. Int Ophthalmol Clin 50: 25–34. [DOI] [PubMed] [Google Scholar]
- 5. Mason SL, Stewart RM, Kearns VR, Williams RL, Sheridan CM (2011) Ocular epithelial transplantation: current uses and future potential. Regen Med 6: 767–782. [DOI] [PubMed] [Google Scholar]
- 6. Baradaran-Rafii A, Eslani M, Tseng SC (2011) Sulfur mustard-induced ocular surface disorders. Ocul Surf 9: 163–178. [DOI] [PubMed] [Google Scholar]
- 7. Xiang H, Stallones L, Chen G, Smith GA (2005) Work-related eye injuries treated in hospital emergency departments in the US. Am J Ind Med 48: 57–62. [DOI] [PubMed] [Google Scholar]
- 8. Martin R (2007) Corneal conjunctivalisation in long-standing contact lens wearers. Clin Exp Optom 90: 26–30. [DOI] [PubMed] [Google Scholar]
- 9. Baylis O, Figueiredo F, Henein C, Lako M, Ahmad S (2011) 13 years of cultured limbal epithelial cell therapy: a review of the outcomes. J Cell Biochem 112: 993–1002. [DOI] [PubMed] [Google Scholar]
- 10. Kenyon KR, Tseng SC (1989) Limbal autograft transplantation for ocular surface disorders. Ophthalmology 96: 709–722. [DOI] [PubMed] [Google Scholar]
- 11. Tseng SC, Chen SY, Shen YC, Chen WL, Hu FR (2010) Critical appraisal of ex vivo expansion of human limbal epithelial stem cells. Curr Mol Med 10: 841–850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Sangwan VS, Basu S, Vemuganti GK, Sejpal K, Subramaniam SV, et al. (2011) Clinical outcomes of xeno-free autologous cultivated limbal epithelial transplantation: a 10-year study. Br J Ophthalmol 95: 1525–1529. [DOI] [PubMed] [Google Scholar]
- 13. Sejpal K, Ali MH, Maddileti S, Basu S, Ramappa M, et al. (2013) Cultivated limbal epithelial transplantation in children with ocular surface burns. JAMA Ophthalmol 131: 731–736. [DOI] [PubMed] [Google Scholar]
- 14. Fukuda K, Chikama T, Nakamura M, Nishida T (1999) Differential distribution of subchains of the basement membrane components type IV collagen and laminin among the amniotic membrane, cornea, and conjunctiva. Cornea 18: 73–79. [PubMed] [Google Scholar]
- 15. Dietrich-Ntoukas T, Hofmann-Rummelt C, Kruse FE, Schlötzer-Schrehardt U (2012) Comparative analysis of the basement membrane composition of the human limbus epithelium and amniotic membrane epithelium. Cornea 31: 564–569. [DOI] [PubMed] [Google Scholar]
- 16. Riau AK, Beuerman RW, Lim LS, Mehta JS (2010) Preservation, sterilization and de-epithelialization of human amniotic membrane for use in ocular surface reconstruction. Biomaterials 31: 216–225. [DOI] [PubMed] [Google Scholar]
- 17. Schulze U, Hampel U, Sel S, Goecke TW, Thäle V, et al. (2012) Fresh and cryopreserved amniotic membrane secrete the trefoil factor family peptide 3 that is well known to promote wound healing. Histochem Cell Biol 138: 243–250. [DOI] [PubMed] [Google Scholar]
- 18. Zhang T, Yam GH, Riau AK, Poh R, Allen JC, et al. (2013) The effect of amniotic membrane de-epithelialization method on its biological properties and ability to promote limbal epithelial cell culture. Invest Ophthalmol Vis Sci 54: 3072–3081. [DOI] [PubMed] [Google Scholar]
- 19. Kubo M, Sonoda Y, Muramatsu R, Usui M (2001) Immunogenicity of human amniotic membrane in experimental xenotransplantation. Invest Ophthalmol Vis Sci 42: 1539–1546. [PubMed] [Google Scholar]
- 20. Baradaran-Rafii A, Aghayan HR, Arjmand B, Javadi MA (2007) Amniotic membrane transplantation. Iran J Ophthalmic Res 2: 58–75. [Google Scholar]
- 21. Liu J, Sheha H, Fu Y, Liang L, Tseng SC (2010) Update on amniotic membrane transplantation. Expert Rev Ophthalmol 5: 645–661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Tamhane A, Vajpayee RB, Biswas NR, Pandey RM, Sharma N, et al. (2005) Evaluation of amniotic membrane transplantation as an adjunct to medical therapy as compared with medical therapy alone in acute ocular burns. Ophthalmology 112: 1963–1969. [DOI] [PubMed] [Google Scholar]
- 23. Gomes JA, Romano A, Santos MS, Dua HS (2005) Amniotic membrane use in ophthalmology. Curr Opin Ophthalmol 16: 233–240. [DOI] [PubMed] [Google Scholar]
- 24. Kolli S, Lako M, Figueiredo F, Mudhar H, Ahmad S (2008) Loss of corneal epithelial stem cell properties in outgrowths from human limbal explants cultured on intact amniotic membrane. Regen Med 3: 329–342. [DOI] [PubMed] [Google Scholar]
- 25. Li W, Hayashida Y, He H, Kuo CL, Tseng SC (2007) The fate of limbal epithelial progenitor cells during explant culture on intact amniotic membrane. Invest Ophthalmol Vis Sci 48: 605–613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Sudha B, Sitalakshmi G, Iyer GK, Krishnakumar S (2008) Putative stem cell markers in limbal epithelial cells cultured on intact & denuded human amniotic membrane. Indian J Med Res 128: 149–156. [PubMed] [Google Scholar]
- 27.de Melo GB, Gomes JA, da Glória MA, Martins MC, Haapalainen EF (2007) [Morphological assessment of different amniotic membrane epithelial denuding techniques]. Arq Bras Oftalmol 70: 407–411. Portuguese. [DOI] [PubMed]
- 28. Higa K, Shimmura S, Kato N, Kawakita T, Miyashita H, et al. (2007) Proliferation and differentiation of transplantable rabbit epithelial sheets engineered with or without an amniotic membrane carrier. Invest Ophthalmol Vis Sci 48: 597–604. [DOI] [PubMed] [Google Scholar]
- 29. Shortt AJ, Secker GA, Rajan MS, Meligonis G, Dart JK, et al. (2008) Ex vivo expansion and transplantation of limbal epithelial stem cells. Ophthalmology 115: 1989–1997. [DOI] [PubMed] [Google Scholar]
- 30. Shortt AJ, Secker GA, Lomas RJ, Wilshaw SP, Kearney JN, et al. (2009) The effect of amniotic membrane preparation method on its ability to serve as a substrate for the ex-vivo expansion of limbal epithelial cells. Biomaterials 30: 1056–1065. [DOI] [PubMed] [Google Scholar]
- 31. Baharvand H, Heidari M, Ebrahimi M, Valadbeigi T, Salekdeh GH (2007) Proteomic analysis of epithelium-denuded human amniotic membrane as a limbal stem cell niche. Mol Vis 13: 1711–1721. [PubMed] [Google Scholar]
- 32. Koizumi N, Inatomi T, Quantock AJ, Fullwood NJ, Dota A, et al. (2000) Amniotic membrane as a substrate for cultivating limbal corneal epithelial cells for autologous transplantation in rabbits. Cornea 19: 65–71. [DOI] [PubMed] [Google Scholar]
- 33. Shimazaki J, Aiba M, Goto E, Kato N, Shimmura S, et al. (2002) Transplantation of human limbal epithelium cultivated on amniotic membrane for the treatment of severe ocular surface disorders. Ophthalmology 109: 1285–1290. [DOI] [PubMed] [Google Scholar]
- 34. Grueterich M, Espana EM, Tseng SC (2003) Ex vivo expansion of limbal epithelial stem cells: amniotic membrane serving as a stem cell niche. Surv Ophthalmol 48: 631–646. [DOI] [PubMed] [Google Scholar]
- 35. Grueterich M, Espana EM, Tseng SC (2003) Modulation of keratin and connexin expression in limbal epithelium expanded on denuded amniotic membrane with and without a 3T3 fibroblast feeder layer. Invest Ophthalmol Vis Sci 44: 4230–4236. [DOI] [PubMed] [Google Scholar]
- 36. Wilshaw SP, Kearney JN, Fisher J, Ingham E (2006) Production of acellular amniotic membrane matrix for use in tissue engineering. Tissue Eng 12: 2117–2129. [DOI] [PubMed] [Google Scholar]
- 37. Fatima A, Sangwan VS, Iftekhar G, Reddy P, Matalia H, et al. (2006) Technique of cultivating limbal derived corneal epithelium on human amniotic membrane for clinical transplantation. J Postgrad Med 52: 257–261. [PubMed] [Google Scholar]
- 38. Mariappan I, Maddileti S, Savy S, Tiwari S, Gaddipati S, et al. (2010) In vitro culture and expansion of human limbal epithelial cells. Nat Protoc 5: 1470–1479. [DOI] [PubMed] [Google Scholar]
- 39. Hopkinson A, Shanmuganathan VA, Gray T, Yeung AM, Lowe J, et al. (2008) Optimization of amniotic membrane (AM) denuding for tissue engineering. Tissue Eng Part C Methods 14: 371–381. [DOI] [PubMed] [Google Scholar]
- 40. Zakaria N, Koppen C, Van Tendeloo V, Berneman Z, Hopkinson A, et al. (2010) Standardized limbal epithelial stem cell graft generation and transplantation. Tissue Eng Part C Methods 16: 921–927. [DOI] [PubMed] [Google Scholar]
- 41. Ljubimov AV, Martel N, Yamasaki H (1985) Response of cultured rat liver epithelial cell lines to tumour-promoting phorbol esters. Exp Cell Res 156: 311–326. [DOI] [PubMed] [Google Scholar]
- 42. Vivek Kumar PR, Cheriyan VD, Seshadri M (2009) Could a strong alkali deproteinization replace the standard lysis step in alkaline single cell gel electrophoresis (comet) assay (pH>13)? Mutation Res 678: 65–70. [DOI] [PubMed] [Google Scholar]
- 43. Kruse FE, Joussen AM, Rohrschneider K, You L, Sinn B, et al. (2000) Cryopreserved human amniotic membrane for ocular surface reconstruction. Graefes Arch Clin Exp Ophthalmol 238: 68–75. [DOI] [PubMed] [Google Scholar]
- 44. Saghizadeh M, Soleymani S, Harounian A, Bhakta B, Troyanovsky SM, et al. (2011) Alterations of epithelial stem cell marker patterns in human diabetic corneas and effects of c-met gene therapy. Mol Vis 17: 2177–2190. [PMC free article] [PubMed] [Google Scholar]
- 45. Shukla IM (1962) Amniotic membrane grafts in corneal ulcer. J All India Ophthalmol Soc 10: 55–60. [PubMed] [Google Scholar]
- 46. Kim JC, Tseng SC (1995) The effects on inhibition of corneal neovascularization after human amniotic membrane transplantation in severely damaged rabbit corneas. Korean J Ophthalmol 9: 32–46. [DOI] [PubMed] [Google Scholar]
- 47.Euro GTPs website: Available: http://eurogtps.com/Portals/0/pdf/Euro%20GTP%20Final%20Delivery.pdf Accessed 2013 October 4.
- 48. Chen B, Mi S, Wright B, Connon CJ (2010) Differentiation status of limbal epithelial cells cultured on intact and denuded amniotic membrane before and after air-lifting. Tissue Eng Part A 16: 2721–2729. [DOI] [PubMed] [Google Scholar]
- 49. Hendrix MJ, Gehlsen KR, Wagner HN Jr, Rodney SR, Misiorowski RL, et al. (1985) In vitro quantification of melanoma tumor cell invasion. Clin Exptl Metastasis 3: 221–233. [DOI] [PubMed] [Google Scholar]
- 50. Ingber DE, Madri JA, Jamieson JD (1986) Basement membrane as a spatial organizer of polarized epithelia. Exogenous basement membrane reorients pancreatic epithelial tumor cells in vitro. Am J Pathol 122: 129–139. [PMC free article] [PubMed] [Google Scholar]
- 51. Davis GE, Blaker SN, Engvall E, Varon S, Manthorpe M, et al. (1987) Human amnion membrane serves as a substratum for growing axons in vitro and in vivo. Science 236: 1106–1109. [DOI] [PubMed] [Google Scholar]
- 52. Allen TD, Aplin JD, Campbell S (1988) Surface visualisation of tissue interfaces by scanning electron microscopy. Methods for exposure of the basal lamina and associated structures in human amnion. Scanning Microsc 2: 2067–2076. [PubMed] [Google Scholar]
- 53. Saghizadeh M, Kramerov AA, Yu FS, Castro MG, Ljubimov AV (2010) Normalization of wound healing and diabetic markers in organ cultured human diabetic corneas by adenoviral delivery of c-Met gene. Invest Ophthalmol Vis Sci 51: 1970–1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Connon CJ, Doutch J, Chen B, Hopkinson A, Mehta JS, et al. (2010) The variation in transparency of amniotic membrane used in ocular surface regeneration. Br J Ophthalmol 94: 1057–1061. [DOI] [PubMed] [Google Scholar]
- 55. Ljubimov AV, Burgeson RE, Butkowski RJ, Michael AF, Sun TT, et al. (1995) Human corneal basement membrane heterogeneity: topographical differences in the expression of type IV collagen and laminin isoforms. Lab Invest 72: 461–473. [PubMed] [Google Scholar]
- 56. Kabosova A, Azar DT, Bannikov GA, Campbell KP, Durbeej M, et al. (2007) Compositional differences between infant and adult human corneal basement membranes. Invest Ophthalmol Vis Sci 48: 4989–4999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Schlötzer-Schrehardt U, Dietrich T, Saito K, Sorokin L, Sasaki T, et al. (2007) Characterization of extracellular matrix components in the limbal epithelial stem cell compartment. Exp Eye Res 85: 845–860. [DOI] [PubMed] [Google Scholar]
- 58. Gedrange T, Mai R, Mack F, Zietek M, Borsos G, et al. (2008) Evaluation of shape and size changes of bone and remodelled bone substitute after different fixation methods. J Physiol Pharmacol 59 Suppl 587–94. [PubMed] [Google Scholar]
- 59. Nakatsu MN, Ding Z, Ng MY, Truong TT, Yu F, et al. (2011) Wnt/β-catenin signaling regulates proliferation of human cornea epithelial stem/progenitor cells. Invest Ophthalmol Vis Sci 52: 4734–4741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Blazejewska EA, Schlötzer-Schrehardt U, Zenkel M, Bachmann B, Chankiewitz E, et al. (2009) Corneal limbal microenvironment can induce transdifferentiation of hair follicle stem cells into corneal epithelial-like cells. Stem Cells 27: 642–652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Yu J, Hu K, Smuga-Otto K, Tian S, Stewart R, et al. (2009) Human induced pluripotent stem cells free of vector and transgene sequences. Science 324: 797–801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Okita K, Matsumura Y, Sato Y, Okada A, Morizane A, et al. (2011) A more efficient method to generate integration-free human iPS cells. Nat Methods 8: 409–412. [DOI] [PubMed] [Google Scholar]
- 63.Dimitrijevich SD, Shankardas J (2008) Characterization of telomerized human corneal epithelial cell line. Invest Ophthalmol Vis Sci 49: E-Abstract 4306.
- 64. Couchman JR, Ljubimov AV (1989) Mammalian tissue distribution of a large heparan sulfate proteoglycan detected by monoclonal antibodies. Matrix 9: 311–321. [DOI] [PubMed] [Google Scholar]