

Abstract
The ascomycetes Candida albicans, Saccharomyces cerevisiae and Scheffersomyces stipitis metabolize the pentose sugar xylose very differently. S. cerevisiae fails to grow on xylose, while C. albicans can grow, and S. stipitis can both grow and ferment xylose to ethanol. However, all three species contain highly similar genes that encode potential xylose reductases and xylitol dehydrogenases required to convert xylose to xylulose, and xylulose supports the growth of all three fungi. We have created C. albicans strains deleted for the xylose reductase gene GRE3, the xylitol dehydrogenase gene XYL2, as well as the gre3 xyl2 double mutant. As expected, all the mutant strains cannot grow on xylose, while the single gre3 mutant can grow on xylitol. The gre3 and xyl2 mutants are efficiently complemented by the XYL1 and XYL2 from S. stipitis. Intriguingly, the S. cerevisiae GRE3 gene can complement the Cagre3 mutant, while the ScSOR1 gene can complement the Caxyl2 mutant, showing that S. cerevisiae contains the enzymatic capacity for converting xylose to xylulose. In addition, the gre3 xyl2 double mutant of C. albicans is effectively rescued by the xylose isomerase (XI) gene of either Piromyces or Orpinomyces, suggesting that the XI provides an alternative to the missing oxido-reductase functions in the mutant required for the xylose-xylulose conversion. Overall this work suggests that C. albicans strains engineered to lack essential steps for xylose metabolism can provide a platform for the analysis of xylose metabolism enzymes from a variety of species, and confirms that S. cerevisiae has the genetic potential to convert xylose to xylulose, although non-engineered strains cannot proliferate on xylose as the sole carbon source.
Introduction
Sugars represent an important source of carbon and energy for fungi. However, different species can have widely differing capacities to use sugars for growth. In general, fungal cells prefer to use the 6-carbon sugar glucose, which they can convert to pyruvate and ultimately oxidize to CO2 and water with the concomitant production of up to 36 ATP units through the process of oxidative phosphorylation [1]. Because of this efficiency, many cells suppress the metabolism of other sugars in the presence of glucose [2]. However, in the absence of a direct glucose source other hexoses can be modified into glucose/fructose for entry into the glycolytic pathway, for example galactose can be modified to fructose through the action of the Leloir pathway enzymes [3,4]. As well, disaccharides and more complex sugars can be enzymatically converted to monosaccharides and ultimately to glucose/fructose for entry into the glycolytic pathway [5-7].
Pentose sugars are also a source of carbon and energy for fungi, but the metabolism of these sugars involves a distinct pathway relative to the hexose sugars. Cells make use of the transaldolase and transketolase reactions of the pentose phosphate pathway to generate 3 and 6 carbon intermediates from 5 carbon sugars [8,9]. The resulting hexose-phosphates and glyceraldehyde-3-phosphate enter into glycolysis and oxidative phosphorylation.
Not all fungi make equivalent use of pentose sugars. As one of the most prevalent pentose sugars in nature, xylose makes up to 40% of hemicellulose and is highly abundant in the biosphere. Current efforts to replace fossil fuel with more renewable fuel supplies have led to interest in generating ethanol directly from biomass material; fermentation of xylose to ethanol is thus of commercial interest because it is such a large component of biomass feedstocks [9,10]. However, present production of commercial ethanol depends primarily on the fermentation of hexose sugars by S. cerevisiae. While this yeast ferments glucose to ethanol very efficiently, it is ineffective in metabolizing xylose [8,9]. Xylose fermenting yeasts such as S. stipitis are not as effective in industrial fermentation as S. cerevisiae, so efforts have been made to genetically modify S. cerevisiae by adding metabolic capacity from organisms that can efficiently ferment xylose [8,11].
Although S. cerevisiae is unable to metabolize xylose, it can utilize xylulose as a sole carbon source [12,13], suggesting that S. cerevisiae lacks the capacity of converting xylose to xylulose. Therefore genetic and metabolic engineering of S. cerevisiae in the past decades have involved transferring xylose reductase (XR) and xylitol dehydrogenase (XDH) genes, the two critical enzymatic activities for this conversion, from xylose fermenting organisms such as S. stipitis, but these efforts have generated only limited success [14]. The imbalance of co-factors caused by XR (requiring NADPH) and XDH (requiring NAD) in the process has been proposed to be an important issue, and efforts to overcome the problem have focused on identifying an XR that utilizes NADH instead NADPH [15]. An alternative approach has been to express fungal or bacterial xylose isomerases in S. cerevisiae to bypass the need of co-factors in the conversion of xylose to xylulose [16]. Both of these approaches have reported success in improving the ability of S. cerevisiae to metabolize xylose [11,16,17]. However, commercial application of these modified strains in the industrial fermentation of xylose to ethanol has not been achieved.
Comparative genomic studies show that both XR and XDH exist in S. cerevisiae, and they are well conserved when compared to other xylose metabolizing fungi such as C. albicans and S. stipitis. It is important to know if the S. cerevisiae XR and XDH have the expected enzymatic capacity, as this will help to design strategies to engineering S. cerevisiae for improved xylose metabolism. Here we show that XR and XDH of S. cerevisiae complement the corresponding function of these genes in the xylose metabolizing yeast C. albicans, demonstrating clearly that it is not the enzymatic activities per se, and may not even be the cofactor imbalance, that prevents an effective xylose metabolism in S. cerevisiae. Using C. albicans deletion mutants, we also demonstrate that xylose isomerases can bypass the requirement for XR and XDH to permit C. albicans growth on xylose as a sole carbon source.
Materials and Methods
Media and culture conditions
Yeast strains in this study were grown in YPD or synthetic medium (0.67% Difco yeast nitrogen base without amino acids) supplemented with complete amino acids and filter sterilized D-glucose (2%), D-xylose (2%), xylitol (2%) or xylulose (0.5%)
Plasmid construction
Plasmids and oligonucleotides used in this study are listed in Tables 1 and 2. For complementation studies all genes were cloned into the integration plasmid CIpACT1-CYC, which contains the Candida albicans ACT1 promoter, CYC1 terminator and RPS1 gene, to allow the cloned in gene of interest to be integrated at the RPS1 locus [18] and expressed under the control of the ACT1 promoter. Integration is targeted to RPS1 by linearizing the plasmid with either a StuI or NcoI digest. However a fraction of integrations were also found at ACT1. All plasmids used in this study were linearized with StuI for integration with the exception of pDH271, which was linearized with NcoI. All constructs were sequenced to confirm integrity.
Table 1. Plasmids used in this study.
| Plasmid | Description | Reference |
|---|---|---|
| pGEM-HIS1 | pGEM-T (Promega) with C. albicans HIS1 | [21] |
| pRSARG4Δ SpeI | pRS314 with C. albicans ARG4 | [21] |
| pSN40 | pCR-BluntII- TOPO with C. maltosa LEU2 | [28] |
| pSFS2A | C. albicans SAT1 cassette | [22] |
| CIpACT-CYC | ACT1p-CYC1t-RPS1-cURA3 | [18] |
| pDH270 | CIpACT-CYC with S. stipitis XYL2 | This study |
| pDH271 | CIpACT-CYC with S. stipitis XYL1 | This study |
| pDH275 | CIpACT-CYC with Piromyces sp. XYLA | This study |
| pDH276 | CIpACT-CYC with Orpinomyces sp. UKK1 | This study |
| pDH277 | CIpACT-CYC with T. thermophilus TTHXYLA (codon optimized for Candida albicans) | This study |
| pDH278 | CIpACT-CYC with C. cellulolyticum xylose isomerase (codon optimized for Candida albicans) | This study |
| pDH279 | CIpACT-CYC with S. cerevisiae SOR1 | This study |
| pDH280 | CIpACT-CYC with S. cerevisiae XYL2 | This study |
| Plate 393 | CIpACT-CYC with S. cerevisiae GRE3 | This study |
| Plate 395 | pUC57 with S. cerevisiae GRE3 (CTG/Leu to TTG) | This study |
| Plate 396 | CIpACT-CYC with S. cerevisiae GRE3 (CTG/Leu to TTG) | This study |
| pGen5 | pUC57 with Piromyces sp. XYLA | This study |
| pGen6 | pUC57 with Orpinomyces sp. UKKI | This study |
| pGen7 | pUC57 with T. thermophilus TTHXYLA (codon optimized for Candida albicans) | This study |
| pGen8 | pUC57 with C. cellulolyticum xylose isomerase (codon optimized for Candida albicans) | This study |
Table 2. Oligonucleotides used in this study.
| Name | Sequence | Gene |
|---|---|---|
| MR2305 | CAATTTTCTTTTTCGCCCTCGCACCACCCCTCCCCCTCCCTCTCCCTCCACACCTTCATATGTGGAATTGTGAGCGGATA | CaGRE3 |
| MR2306 | TAGTTTTTTTTTAATACGAGCCTACCACCACCAACTAATTTTTTTTTAGTCGATTTTGCGCAATTTTCTTTTTCGCCCTC | CaGRE3 |
| MR2307 | TATTTACATATTCAAAGATTTCAAAATATACACATTATATATATATATATATATATATGTGTTTTCCCAGTCACGACGTT | CaGRE3 |
| MR2308 | TATTAGGTAGACAAGCAGTTTTAGATGCCATTTAAAAAGAAAAAAAGAATGTTCACCTTTTATTTACATATTCAAAGATT | CaGRE3 |
| MR2309 | CCTTCCACATTCTTCAGATTGTTATGC | CaGRE3 |
| MR2310 | GGTGGTACCACTTCATTAGTTGATGC | CaGRE3 |
| MR2311 | CGTCTATAAATACACCTTGCAATCCCCCCCCCCATTCATTCGTTTTTTATTTCCCCAACCTGTGGAATTGTGAGCGGATA | CaXYL2 |
| MR2312 | TGATTTTTCGGAGAAATTGTTACTCCCAATTGTTACTCCTTATTTGTTGCTCCTAATTTTCGTCTATAAATACACCTTGC | CaXYL2 |
| MR2313 | GAAAAATTTTTCTTTTTTTTTATTCGACTATACACTTGGTAGCACTATAGATTTCTACATGTTTTCCCAGTCACGACGTT | CaXYL2 |
| MR2314 | GGGAGTGAGTTGGAAAGACATGGTTTGGTTGTTGTTTGGGGTGTGTCTTTTTTTTTTGGAGAAAAATTTTTCTTTTTTTT | CaXYL2 |
| MR2315 | GTAGACAGGAGAACTGGCTTACTCTG | CaXYL2 |
| MR2316 | GTGAGACAGATATTGTTGCCTGATGAG | CaXYL2 |
| MR2511 | CGTCTATAAATACACCTTGCAATCCCCCCCCCCATTCATTCGTTTTTTATTTCCCCAACCGAGCTCCACCGCGGTGGCGGCCGCT | CaXYL2 |
| MR2512 | GAAAAATTTTTCTTTTTTTTTATTCGACTATACACTTGGTAGCACTATAGATTTCTACATGGTACCGGGCCCCCCCTCGAGGAA | CaXYL2 |
| MR2004 | TAAATTTGTGTTGTTCGGTGACTCCATCAC | SAT1 |
| MR2005 | TCTCATATGAAAATTTCGGTGATCCCTGAG | SAT1 |
| MR2357 | CTGAAGAGTTCCCAACTTTGTCTGTTC | LEU2 |
| MR2358 | CCGTTACAGGAGTTAGTCATTCTTCAAC | LEU2 |
| MR1322 | GGAGTTCCATTTAGAGAAACTCATC | ARG4 |
| MR1323 | CGAAAATTCTCATATCGGTAGCAAC | ARG4 |
| MR1324 | GAAGTTGTGTGGGAAAAATCTTCAC | HIS1 |
| MR1325 | ACAGTTCACCTGGTACGGTTTTCTAG | HIS1 |
| ODH322 | TCCcccgggATGACTGCTAACCCTTCCTTGG a | SsXYL2 |
| ODH323 | CTAGtctagaTTACTCAGGGCCGTCAATGAGA | SsXYL2 |
| ODH326 | TCCcccgggATGCCTTCTATTAAGTTGAACTCTGGTTA | SsXYL1 |
| ODH328 | CTAGtctagaTTAGACGAAGATAGGAATCTTGTCCCAGTCCCAAGGGTCGTTGAATCTCAAG | SsXYL1 |
| ODH352 | CTGCAGacgcgtATGTCTCAAAATAGTAACCCT | ScSOR1 |
| ODH353 | GGAATTgtcgacTCATTCAGGACCAAAGATAATAGT | ScSOR1 |
| ODH354 | CTGCAGacgcgtATGACTGACTTAACTACACAAGAA | ScXYL2 |
| ODH355 | GGAATTgtcgacTCATTCCGGGCCCTCAATGATCGT | ScXYL2 |
| ODH346 | TGATACCGACAGTATCATCGAT | ACT1 flanking |
| p517 | TTTCTGGTGAATGGGTCAACGAC | RPS1 |
| p544 | AATAGAGAGAAACTATATTATACAC | RPS1 flanking |
| p565 | TTTTCTAATTTTCACTCCTGG | ACT1 promoter |
| p641 | GCCTTCTATTAAGTTGAACTCTGG | SsXYL1 |
| p642 | ATGACTGCTAACCCTTCCTTGG | SsXYL2 |
| p643 | AATGAGACACTTGACAGCACCC | SsXYL2 |
| p644 | TCGTTGAATCTCAAGTTGATGTCC | SsXYL1 |
| p645 | ACCCAATATCTAATTTAGAAATAGC | CaGRE3 |
| p648 | GAAAATTGGAAAacgcgtAAAGATGTCTTCACTGGTTACTC | ScGRE3 |
| p649 | GATTTTACTGgctagcTCAGGCAAAAGTGGGGAATTTACC | ScGRE3 |
| p650 | AATTTACCATCCAACCAGGTCC | ScGRE3 |
| p657 | AAATTGACAAAAAAGTCTGTGCG | ScGRE3 |
| p658 | TGTATAAATATCAAACGATTTCTCC | CaGRE3 |
Lower case letters are restriction sites.
The open reading frames of the S. stipitis (Ss.) genes XYL1 and XYL2 were amplified by PCR from Ss. (CBS 6054) genomic DNA using primers ODH326/328 and ODH322/323 respectively. The products were cloned into the SmaI/NheI linearized vector CIpACT1-CYC to give rise to pDH271 and pDH270 respectively. ODH328 was designed to mutate the NcoI site in the XYL1 gene so the plasmid could later be linearized with NcoI for integrating at RPS1. The S. cerevisiae (Sc.) SOR1 and XYL2 coding regions were PCR amplified from S. cerevisiae (BY4743) genomic DNA with ODH352/353 and ODH354/355 respectively and the products cloned to the MluI/SalI sites of CIp-ACT1-CYC to produce pDH279 and pDH280. The coding region for the ScGRE3 gene, which contains 5 CTG/Leu codons, was amplified by PCR from S. cerevisiae (BY4743) genomic DNA with primers p648/649 and cloned at the MluI/NheI sites of the vector CIpACT1-CYC to produce plasmid Plate 393. For the “candidized” version, the coding region for the ScGRE3 gene, with the addition of flanking MluI and NheI restriction sites, was synthesized (GenScript, New Jersey) with each of the 5 CTG/Leu codons changed for TTG/Leu codons and the synthetic gene cloned in vector pUC57 to produce plasmid Plate 395. Plasmid Plate 395 was then digested with MluI and NheI; the 1-kb fragment for the “candidized” ScGRE3 coding region was gel-purified and cloned in vector CIpACT1-CYC to produce plasmid Plate 396.
The open reading frames of 4 xylose isomerase genes were also synthesized by GenScript: XYLA from Piromyces sp. (CAB76571), Orpinomyces sp. (ACA65427) designated OrpXI, TTHXYLA from Thermus thermophilus (BAA14301) and Clostridium cellulolyticum (YP_002507697) designated as CcelXI. TTHXYLA and CcelXI were codon optimized for Candida albicans. The start codon for TTHXYLA was changed from GTG to ATG and SalI and NheI sites were added 5’ and 3’ respectively. XhoI and SpeI sites were added 5’- and 3’-end respectively to XYLA, OrpXI and CcelXI. The synthesized genes were supplied in the vector pUC57 at the EcoRV site.
To clone these genes into the integrating plasmid, the XYLA, OrpXI and CcelXI genes were each cut out from pUC57 with an XhoI digestion, blunt-ended, and subsequently digested with SpeI. These genes were then cloned into the CIpACT1-CYC plasmid at the SmaI/NheI sites to give rise to pDH275, pDH276 and pDH278 respectively. pDH277 was constructed similarly, except that the TTHXYLA gene was moved out of the pUC57-based construct with a SalI digestion, blunt-ended and followed by an NheI digestion. Cloning in this way removed an out-of-frame ATG between the SmaI and SalI site of the vector.
Strain construction and manipulation
All strains used in this study are listed in Table 3. All C. albicans strain constructions, transformations, and subsequent manipulations were carried out using standard procedures [19,20]. Candida deletion strains were all constructed in strain SN148 using PCR-based gene targeting [21]. To construct the gre3-9 and xyl2-16 deletion strains, two runs of PCR using 80-mers were performed on plasmids pGEM-HIS1 or pRSARG4∆SpeI, and the PCR products were used to replace the entire targeted ORF of either GRE3 or XYL2 at one allele marked with HIS1 and the second allele with ARG4. To construct the Candida gre3/xyl2-3 double deletion mutant, XYL2 was deleted in the gre3-9 strain. PCR products generated from plasmids pSN40 or pSFS2A were used to replace the entire targeted ORF at one allele with LEU2 and the second allele with SAT1. The SAT1-FLP cassette [22] was then looped out to leave behind only the FRT sequence.
Table 3. Strains used in this study.
| Strain | Genotype | Reference |
|---|---|---|
| CBS 6054 | Scheffersomyces stipitis wild-type isolated from insect larvae | ATCC 58785 |
| SC5314 | Candida albicans wild-type clinical isolate | [37] |
| BY4741 | MATa his3∆1 leu2∆0 met15∆0 ura3∆0. | [38] |
| BY4743 | MATa/MATalpha his3∆1/his3∆1 leu2∆0/leu2∆0 lys2∆0/LYS2 MET15/met15∆0 ura3∆0/ura3∆0(4741/4742) | [38] |
| SN148 | arg4∆/arg4∆ leu2∆/leu2∆ his1∆/his1∆ ura3∆::imm434/ura3∆::imm434 iro1∆::imm434/iro1∆::imm434 | [28] |
| gre3-9 | SN148 gre3::HIS1/gre3::ARG4 | This study |
| xyl2-16 | SN148 xyl2::HIS1/xyl2::ARG4 | This study |
| gre3/xyl2-3 | gre3::HIS1/gre3::ARG4 xyl2::LEU2/xyl2::FRT | This study |
| CDH120 | gre3::HIS1/gre3::ARG4 RPS1/RPS1:: [CIpACT-CYC - S. stipitis XYL1 (pDH271)] | This study |
| CDH116 | xyl2::HIS1/xyl2::ARG4 RPS1/RPS1:: [CIpACT-CYC - S. stipitis XYL2 (pDH270)] | This study |
| CDH139 | xyl2::HIS1/xyl2::ARG4 RPS1/RPS1:: [CIpACT-CYC- S. cerevisiae SOR1 (pDH279)] | This study |
| CDH128 | gre3::HIS1/gre3::ARG4 xyl2::LEU2/xyl2::FRT RPS1/RPS1:: [CIpACT-CYC Piromyces sp. XYLA (pDH275)] | This study |
| CDH129 | gre3::HIS1/gre3::ARG4 xyl2::LEU2/xyl2::FRT RPS1/RPS1:: [CIpACT-CYC Orpinomyces sp. UKK1 (pDH276)] | This study |
| CDH130 | gre3::HIS1/gre3::ARG4 xyl2::LEU2/xyl2::FRT RPS1/RPS1:: [CIpACT-CYC T. thermophilus TTHXYLA (pDH277)] | This study |
| CDH131 | gre3::HIS1/gre3::ARG4 xyl2::LEU2/xyl2::FRT RPS1/RPS1:: [CIpACT-CYC C. cellulolyticum xylose isomerase (pDH278)] | This study |
| CDH140 | gre3::HIS1/gre3::ARG4 RPS1/RPS1:: [CIpACT-CYC - Piromyces sp. XYLA (pDH275)] | This study |
| CDH141 | xyl2::HIS1/xyl2::ARG4 RPS1/RPS1:: [CIpACT-CYC - Piromyces sp. XYLA (pDH275)] | This study |
| CA220 | gre3::HIS1/gre3::ARG4 xyl2::LEU2/xyl2::FRT RPS1/RPS1:: [CIpACT-CYC S. stipitis XYL2 (pDH270)] | This study |
| CA227 | gre3::HIS1/gre3::ARG4 xyl2::LEU2/xyl2::FRT RPS1/RPS1:: [CIpACT-CYC S. stipitis XYL2 (pDH270)] ura3-/ura3- | This study |
| CA242 | gre3::HIS1/gre3::ARG4 xyl2::LEU2/xyl2::FRT RPS1/RPS1:: [CIpACT-CYC S. stipitis XYL2 (pDH270)]::[CIpACT-CYC S. stipitis XYL1 (pDH271)] | This study |
| CA243 | gre3::HIS1/gre3::ARG4 xyl2::LEU2/xyl2::FRT RPS1/RPS1:: [CIpACT-CYC S. stipitis XYL2 (pDH270)]::[CIpACT-CYC S. stipitis XYL1 (pDH271)] | This study |
All strains used for complementation analyses were created by chromosomally integrating the desired genes at either the RPS1 or the ACT1 locus. Specifically, the C. albicans gre3-9 strain was transformed with the plasmid pDH271 carrying the SsXYL1 gene to create strain CDH120. Transformation of strain xyl2-16 with pDH270 carrying SsXYL2 gave rise to strain CDH116, and pDH279 carrying ScSOR1 created the strain CDH139. The complementation of the Candida gre3/xyl2-3 double deletion strain was carried out with the S. stipitis XYL1 and XYL2 homologs. The xyl2 complementation was done first with plasmid pDH270 linearized with StuI, and integration at RPS1 was confirmed by PCR (with primers p565/p544). The resulting strain, named CA220, was grown in liquid synthetic medium with xylitol, then selected on a 5-fluoroorotic acid (5-FOA) plate containing xylitol as the sole carbon source to “recycle” the URA auxotrophic marker and retain the xylitol dehydrogenase function to give rise to the ura - strain CA227. Transformation of CA227 with NcoI linearized pDH271 (SsXYL1) followed by selection on 2% xylitol-uridine plates gave rise to strains CA242 and CA243.
Complementation of the gre3-9 strain with ScGRE3 was carried out with StuI linearized Plate 393 to produce strain CA247, and with StuI linearized Plate 396 to produce strains CA255 and CA256. Transformation of the gre3-9/xyl2-16 double deletion strain with pDH275, pDH276, pDH277 or pDH278 constructs harboring the heterologous xylose isomerases created strains CDH128 (Piromyces), CDH129 (Orpinomyces), CDH130 (Thermus thermophilus) and CDH131 (Clostridium cellulolyticum) respectively. The single deletion strains gre3-9 and xyl2-16 were also transformed with the Piromyces isomerase plasmid pDH275 to create strains CDH140 and CDH141.
Growth curves
Overnight cell cultures were grown in synthetic complete medium containing dextrose (2%). Cells were washed once with sterile water before diluting to an OD600nm of 0.2 in 5-10 ml of synthetic complete medium containing xylose (2%), xylitol (2%) or xylulose (0.5%). Cells were grown aerobically at 30°C with shaking, and the OD600nm was taken at intervals for up to two weeks.
Microarrays analysis
The strain BY4743 of S. cerevisiae was used for transcription profiling in xylose medium. A 10-ml starter culture in 2% raffinose (to minimize glucose repression effect) was grown for about 2 days and used to inoculate a 400-ml culture in 2% raffinose. This culture was grown to an OD600 of approximately 0.75 - 0.8 (24-34 hr). The cells were then washed twice with sterile water. The cell pellet was resuspended in 8 ml of synthetic medium without sugar (S), and 2 ml was used to inoculate 100 ml cultures, either without sugar (S), with xylose (SX) or with dextrose (SD) for the reference. These cultures were incubated until an increase in OD600 was observed for the culture with dextrose (1.5 - 2 hr); there was no increase in the OD600 for the culture in xylose medium.
The Candida albicans strain SC5314 was used for microarray in xylose medium.
Starter cultures in xylose and in dextrose media were grown overnight and used to inoculate the xylose and dextrose subcultures, respectively, and these were grown to an OD600 of approximately 0.8.
Cells were disrupted with glass beads in the FastPrep-24 instrument (MP Biomedicals). RNA was extracted with the RNeasy kit (QIAGEN cat. 74104), and 20 µg of total RNA was used for the cDNA synthesis with either cyanine-3 or cyanine-5 dyes. Probe synthesis, microarray hybridizations, washings and analyses were as described [23,24]. S. cerevisiae microarrays (Y6.4k7, 6240 ORFs) were obtained from the University Health Network Microarray Centre (http://www.microarrays.ca, Toronto, Canada). C. albicans microarrays (double-spotted 6,394 intragenic 70-mer oligonucleotides) were obtained from the Biotechnology Research Institute microarray facility center (http://www.nrc cnrc.gc.ca/eng/services/bri/microarray.html, Montreal, Canada). The data sets for these array experiments are provided as excel files as supplementary S2, and the full data sets have been registered at NCBI with the GEO accession number GSE50476.
Results
S. cerevisiae, S. stipitis and C. albicans share conserved xylose metabolic enzymes but have different capacities for metabolizing xylose
The baker’s or brewer’s yeast S. cerevisiae, the fungal pathogen C. albicans, and the pentose-sugar fermenting yeast S. stipitis (formerly Pichia stipitis) are related ascomycetes, with C. albicans and S. stipitis in a lineage that uses the codon CTG to encode serine, unlike S. cerevisiae, which, consistent with the universal genetic code, uses CTG to encode leucine. These ascomycetes have distinct ecological niches; S. cerevisiae is almost a domesticated organism primarily associated with human brewing and baking, C. albicans is a human commensal colonizing mucosal surfaces and the gastrointestinal system, while S. stipitis inhabits the guts of some insects. These niches all provide efficient access to the hexose sugar glucose, but only the guts of wood-eating insects provide high levels of pentose sugars such as xylose for possible metabolism.
Although each of these organisms has different access to xylose as a potential carbon source, they all encode the enzymatic capacity to direct xylose into the pentose phosphate pathway. In fungi in general, the metabolism of xylose involves its transformation to xylulose through a two-step process, the first the reduction of xylose to xylitol by the action of a xylose reductase (XR), and the second the production of xylulose from xylitol by the action of a xylitol dehydrogenase (XDH) (Figure 1A). Sequence analysis indicates that all three species have genes that are predicted to encode these functions, and these genes are conserved (Figure S1). The GRE3 genes of C. albicans and S. cerevisiae, and the XYL1 gene of S. stipitis, are highly related sequences encoding predicted xylose reductase activity. The XYL2 genes of S. stipitis and C. albicans, as well as the XYL2, SOR1 and SOR2 genes of S. cerevisiae are also closely related genes encoding putative dehydrogenases. The SOR1 and SOR2 genes, although annotated as sorbitol dehydrogenases, are more similar to the C. albicans and S. stipitis XYL2 genes than is the yeast gene designated XYL2. Intriguingly however, although the species have similar genomic potential for xylose utilization, they actually have very different growth behaviors on this sugar. As shown in Figure 1B, while strains of each organism were able to grow in medium containing xylulose as the sole carbon source, the S. cerevisiae strain was not able to grow when the carbon source was the related pentose sugar xylose. Overall, Candida grows better than the others in both xylose and xylulose, while S. cerevisiae grows slowly in xylulose, and S. stipitis grows well in both xylose and xylulose, but reaches a plateau after 1 day of incubation in both sugars.
Figure 1. Xylose entry to the PPP pathway and xylose metabolizing capacities of different ascomycetes.
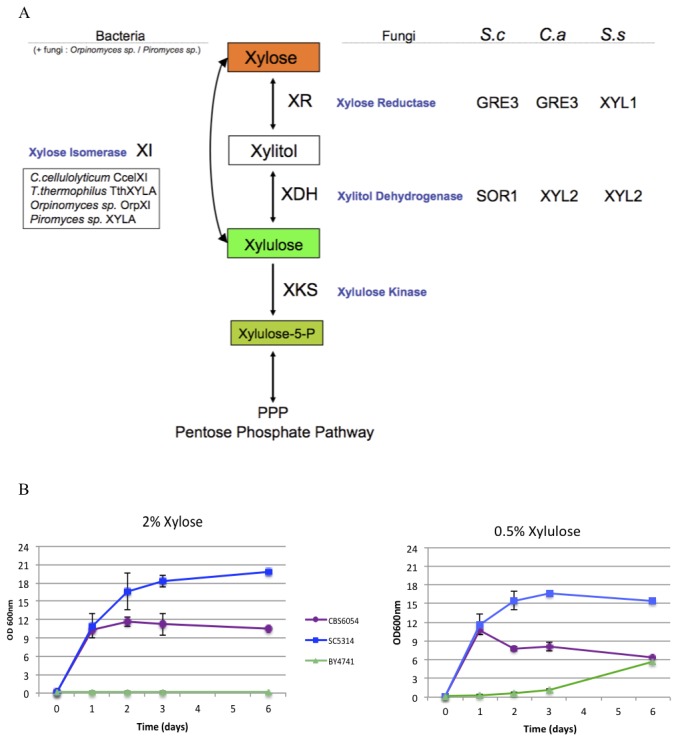
A) The schematic presentation of conserved genes encoding enzymatic activities for the xylose entry into the PPP pathway in ascomycetes. C. a for Candida albicans, S. c for Saccharomyces cerevisiae, and S. s for Schefferomyces stipitis. B) Growth of C. albicans (SC5314), S. stipitis (CBS6054) and S. cerevisiae (BY4741) in SC with 2% xylose or 0.5% xylulose. Strains were grown aerobically at 30°C and the optical density measured over a period of six days, n=3.
Transcription profiling of cellular response to xylose
Previous work has established that in response to growth in xylose, S. stipitis induces the genes for XR and XDH, as well as for some sugar transporters [25,26]. We have examined the response of both C. albicans and S. cerevisiae to incubation with xylose as the sole carbon source. As shown in Figure 2A, C . albicans generates a robust signal when shifted from growth in glucose to xylose. 175 genes have at least a 2-fold transcription increase in xylose when compared to cells in glucose, of which 44 genes with more than a 5-fold induction in xylose are listed in the table. The induced genes include those that are critical for xylose metabolism such as GRE3 (30 fold) and XYL2 (7 fold), as well as the xylulose kinase XKS1 (5 fold). Some genes encoding sugar transporters, including HGT2, HGT12, HXT5, HGT9 and HGT17, are also strongly induced. This shows that C. albicans is capable of shifting its metabolic constitution to exploit xylose as a carbon source. In contrast, S. cerevisiae has no cell growth under these conditions, has a very modest response to the shift from glucose to xylose. This response is quite similar to that seen when shifting yeast cells from glucose medium to a medium containing no sugar source (Figure 2B). This behavior is consistent with the inability of S. cerevisiae to grow on xylose as the sole carbon source, although S. cerevisiae is capable of transporting xylose [27]. The data sets for the C. albicans and S. cerevisiae transcriptional profiling results are presented as Figure S2.
Figure 2. Transcription profile of C. albicans and S. cerevisiae in xylose.

A) From microarray experiments with C. albicans grown in xylose, 175 genes have at least a 2-fold transcription increase in xylose when compared to cells in dextrose. The 44 genes with more than a 5-fold induction in xylose (SX) are shown with their corresponding value. Data for S. cerevisiae is shown for comparison, and in gray-shade if no reciprocal best-hit ortholog is found. B) Transcription modulation of the S. cerevisiae sugar transporters in xylose. The data from the microarray experiments in xylose (SX) and in no-sugar condition (S) is shown for 18 members of the sugar transporter family, relative to cells in dextrose. HXT5 and HXT2 are likely induced due to the absence of dextrose (glucose). HXT6, HXT7 and HXT4 are induced specifically by the presence of xylose. HXT4, HXT1 and HXT3 show a transcriptional reduction only for the no-sugar condition (S), likely due to the absence of sugar. A summary for the transporters description from SGD is reported in the ‘SGD description’ column.
GRE3 and XYL2 are necessary for xylose metabolism in C. albicans
To examine whether GRE3 and XYL2, encoding potential xylose reductase (XR) and xylitol dehydrogenase (XDH) activities respectively, are solely responsible for the metabolism of xylose in C. albicans, we created disruption mutants of both genes. The two alleles of either GRE3 or XYL2 were replaced by HIS1 and ARG4 in strain SN148 [28]. A double mutant, gre3 xyl2, was created by replacing the two alleles of XYL2 with the LEU2 and SAT1 markers in a strain already deleted for GRE3. All the resulting strains were unable to grow on xylose as a sole carbon source, while as expected the gre3 mutant strain was able to grow on xylitol (Figure S3). This confirms that in C. albicans the oxido-reductase activities encoded by the GRE3 and XYL2 genes are solely necessary for the metabolism of xylose.
Complementation of gre3 and xyl2 mutants
We asked whether the corresponding XR and XDH genes of S. stipitis were capable of complementing the C. albicans gre3 and xyl2 deletions. Because both C. albicans and S. stipitis belong to the CUG clade, there was no need to modify the S. stipitis genes prior to introducing them into the C. albicans mutants. The S. stipitis XYL1 gene integrated at the RPS1 locus of C. albicans was capable of supporting robust growth of the gre3 strain within two to three days in xylose medium (Figure 3A). As well, integration of the S. stipitis XYL2 gene in the C. albicans xyl2 strain complemented the deletion strains growth defect within one day in both xylitol or xylose medium (Figure 3B). We also tested the S. stipitis XR-XDH modular unit in the Candida gre3/xyl2 double deletion mutant in xylose metabolism. Integration of S. stipitis XYL1 and XYL2 into the gre3/xyl2 strain allowed the transformed Candida mutant to grow in medium using xylose as the sole carbon source, indicating that the XR-XDH module from S. stipitis is functional in C. albicans (Figure 3C). However, we noticed that these Candida cells showed a lag time of approximately 48 h when grown in medium with xylose compared to medium with xylitol as a carbon source. This lag was not observed with wild-type C. albicans cells, suggesting that the S. stipitis XR-XDH module may not be perfectly coupled to the other parts of the xylose metabolic pathway of C. albicans.
Figure 3. The xylose reductase and xylitol dehydrogenase genes from S. stipitis complement C. albicans deletion mutants of the equivalent genes.
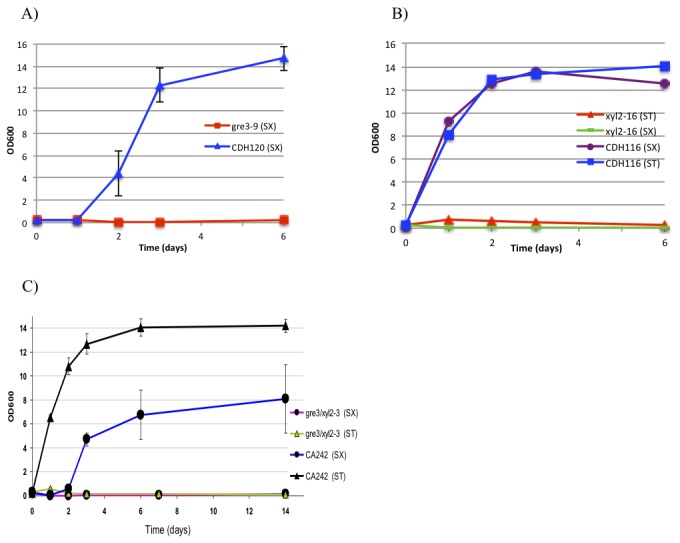
(A) S. stipitis XYL1 (CDH120), and (B) S. stipitis XYL2 (CDH116) complement the C. albicans gre3 and xyl2 deletion mutants respectively. (C) The S. stipitis XYL1-XYL2 module (CA242) complements the C. albicans gre3 xyl2 double deletion mutant. Strains were grown aerobically at 30°C in SC with 2% xylose (SX) or 2% xylitol (ST). The optical density was measured over a period of up to 14 days, n=3.
Similarly, we investigated the ability of the S. cerevisiae homologs to complement the C. albicans mutants. Because ScGRE3 contained 5 CTG codons that would encode leucine in S. cerevisiae but serine in C. albicans we used a synthetic gene with the 5 CTG codons switched to TTG to assess complementation. As shown in Figure 4A, integration of the candidized ScGRE3 gene at RPS1 [18] could complement the gre3 mutation. The CTG codon unmodified ScGRE3 gene was not able to complement when introduced at the same site (data not shown), suggesting the change of one or more leucine residues to serines compromised the enzymatic function. We also assessed the ability of SOR1 and ScXYL2 to complement the xyl2 mutant of C. albicans. The SOR1 gene contained no CTG codons, and when integrated at RPS1 was capable of complementing the xyl2 mutant for growth on xylitol within one day, however complementation of growth in xylose took several days (Figure 4B) The ScXYL2 gene contained 4 CTG codons, and was not capable of complementing the function (data not shown). This could represent problems caused by the leucine/serine discrepancy, or could simply reflect the point that the SOR1 gene is more similar to the C. albicans gene than is ScXYL2. Nonetheless, the observation that ScGRE3 and SOR1 can complement the Candida gre3 and xyl2 mutants shows that S. cerevisiae has XR and XDH enzymatic capacity for the crucial xylose-xylulose conversion steps of xylose metabolism.
Figure 4. The xylose reductase and xylitol dehydrogenase genes from S. cerevisiae complement C. albicans deletion mutants of the equivalent genes.
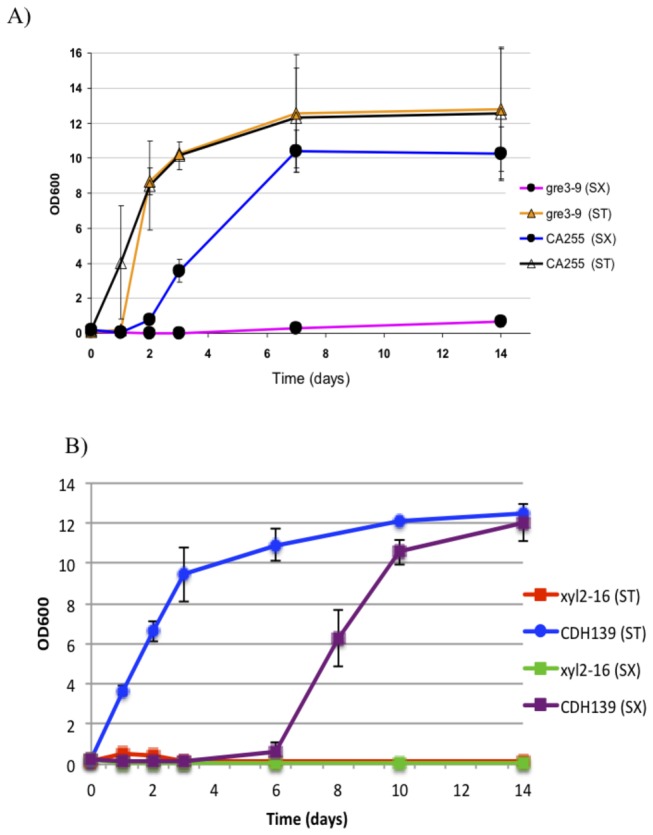
(A) S. cerevisiae GRE3 (CA255), and (B) S. cerevisiae SOR1 (CDH139) complement the C. albicans gre3 and xyl2 deletion mutants respectively. Strains were grown aerobically at 30°C in 2% xylose (SX) or 2% xylitol (ST). The optical density was measured over a period of 14 days, n=3.
When we tested XR-XDH as a modular unit from S. cerevisiae for its capacity to complement the Candida gre3/xyl2 double deletion mutant in xylose. We found the S. cerevisiae XR-XDH module (GRE3-SOR1) showed a clear, but relatively weak ability to support the growth of the C. albicans double mutant on xylose medium. Significant cell growth takes about two weeks (data not shown), which is much slower growth compared to the more rapid growth conferred by either ScGRE3 or ScSOR1 when complementing a gre3 or xyl2 single mutation of C. albicans. These results, along with those from S. stipitis, may suggest that the proper coupling of all the steps in the metabolic pathway is as important as the enzyme activities per se in each step.
Fungal xylose isomerases enable a Candida gre3/xyl2 double mutant to metabolize xylose
We further assessed whether alternate enzymatic activities could function in the xylose utilization pathway of C. albicans. Because the role of XR and XDH is to transform xylose to xylulose, we asked whether expression of a heterologous xylose isomerase could complement the gre3 xyl2 double mutant. Xylose isomerases exist in bacteria, in plants, as well as in some fungi, however, ascomycete fungi like yeast and C. albicans do not have endogenous xylose isomerases. Sequence analysis with phylogenic clustering of 17 xylose isomerases indicate that there are two classes of this enzyme, one represented by the enzyme from Piromyces, and the other from that of Thermus thermophilus as shown in the alignment (Figure S4). Genes corresponding to the XI from fungi (Piromyces sp. and Orpinomyces sp.), and bacteria (Thermus thermophilus and Clostridium cellulolyticum) were synthesized and introduced into the gre3 xyl2 double mutant strain. Piromyces and Orpinomyces have the same codon usage as C. albicans, and thus required no CUG codon modification. Thermus and Clostridium have standard codon usage, so the xylose isomerase genes from these organisms were synthesized with CUG codon modification to reflect C. albicans codon usage. As shown in Figure 5, the isomerase from Piromyces allowed growth of the double mutant in xylose medium within one to two days indicating that the activity of the XI provides effective xylose-xylulose conversion. The same effect was observed with the Orpinomyces XI (data not shown). On the other hand the bacterial isomerases were not able to support the growth of the double the mutant in xylose medium; this could represent a problem with protein production, although the genes were codon optimized for expression in C. albicans, or a problem in enzyme function in situ. The Piromyces isomerase was also able to allow growth of the C. albicans gre3 single deletion mutant in 3-4 days (Figure 5); however, it could not complement the growth of the xyl2 single deletion mutant under the same conditions (data not shown), suggesting an inhibiting effect of accumulating xylitol on XI in xyl2 mutant.
Figure 5. The xylose isomerase gene XYLA from Piromyces sp. complements a double gre3/xyl2 deletion or single gre3 deletion in C. albicans.

Growth analysis in synthetic medium (SC) with 2% xylose as the sole carbon source for C. albicans strains: gre3-9 (gre3 deletion), gre3/xyl2-3 (gre3 xyl2 double deletion), CDH128 (gre3/xyl2-3 with the integrated Pir.XYLA) and CDH140 (gre3-9 with the integrated Pir. XYLA). The optical density was measured over a period of 6 days, n=3.
Discussion
Recent studies [26,29,30] have emphasized the genomic similarities of xylose utilizing fungi such as C. albicans and S. stipitis, and the non-utilizing yeast S. cerevisiae. Here we have extended these studies to show that when put in the common context of the C. albicans cell, genes for xylose metabolism from both the xylose metabolizing yeast S. stipitis and the non-utilizing yeast S. cerevisiae are able to complement the deletion of these functions and permit the growth of mutant C. albicans strains on xylose medium. In the correct context, the GRE3 and SOR1 genes of S. cerevisiae are capable of providing xylose reductase and xylitol dehydrogenase functions and of permitting cell growth on xylose as a sole carbon source. This emphasizes that it is not fundamentally the lack of xylose reductase and xylitol dehydrogenase capacity that prevents yeast cells from growing on xylose, but rather the regulatory and metabolic context of these enzymes.
A significant difference between S. cerevisiae and the xylose-utilizing yeasts C. albicans and S. stipitis is readily apparent when the organisms are examined for their transcriptional response to the sugar. Both C. albicans (Figure 2A; [26]) and S. stipitis [25,31] show robust changes in gene expression in response to the addition of xylose as a carbon source, and many of the induced genes encode components implicated in the transport or metabolism of the sugar. By contrast, as seen in Figure 2A and 2B the response of S. cerevisiae to the presence of xylose is very similar to the response to the absence of any sugar. However, the sugar transporter encoding genes HXT4, HXT6 and HXT7 have a modest induction that was unique to the xylose conditions, suggesting the cells may have been able to distinguish the presence of xylose from the absence of any sugar, even though the cells were not able to use the xylose for growth and proliferation.
It has been proposed that the NADPH/NADH co-factor imbalance created by xylose reductase (XR) and xylitol dehydrogenase (XDH) plays a major role in preventing S. cerevisiae from utilizing xylose. Efforts have been made to change the XR preference for co-factors to alleviate the imbalance and improve xylose consumption [8,32-34]. However, the co-factor imbalance created by the enzymatic activities of XR and XDH with different co-factor preferences appears to be well resolved in C. albicans, suggesting that xylose-metabolizing fungi have the means to deal with the imbalance under aerobic conditions. We do not presently know what activities and processes are responsible for the phenotype. Intriguingly, many genes that are found to be unique to xylose utilizing yeasts are involved in NAD biosynthesis [26], and thus it would be interesting to see if any of these genes could improve xylose metabolism in S. cerevisiae.
We have demonstrated that the sole xylose metabolic pathway in C. albicans is governed by the enzymatic activities encoded by GRE3 and XYL2. Either single deletions or double deletion mutants are unable to metabolize xylose and therefore provide reliable tools and means to analyze the functionality of heterologous components of xylose metabolic pathways. We have analyzed the ability of S. stipitis XYL1 and XYL2 to complement the C. albicans gre3 xyl2 mutant. While the SsXYL1 and XYL2 complement the gre3 xyl2 in general, growth curve analyses indicate that the transformants grow better in xylitol than in xylose, and there is a 48-hour growth lag time in xylose that is absent in xylitol. However, we did not observe this in the single mutant complementation analyses. This suggests that the conversion of xylose to xylitol catalyzed by the Xyl1 reductase activity may be a limiting step, and the lag time likely reflects the time required for the initial accumulation of a critical amount of xylitol to be utilized as substrate for XDH, pointing to a direction of genetic and metabolic engineering to improve the step of xylose conversion. We have also analyzed four xylose isomerases for their ability to complement the gre3 xyl2 double mutant, and found that the two fungal isomerases (from Piromyces and Orpinomyces) could complement the mutant Candida, whereas the two bacterial xylose isomerases (from Thermus and Clostridium) even when codon optimized for Candida, could not. The Thermus thermophilus XI activity, albeit at a low level, has been reported in S. cerevisiae [35], while the Clostidium phytofermentans (not cellulolyticum) enzyme was expressed in S. cerevisiae [36]. Presently, the molecular basis for this discrepancy in terms of functionality in Candida between the fungal and bacterial xylose isomerases remains unclear. Nevertheless, Candida mutants created in this study, combined with mutagenesis, could be powerful tools to evolve and identify desired enzymatic activities for xylose to xylulose conversion. This may help to engineer the yeast S. cerevisiae with a xylose isomerase function suitable for more robust performance in xylose metabolism.
Supporting Information
Alignments of A) xylose reductases and B) xylose dehydrogenases of C. albicans, S. stipitis and S. cerevisiae.
(DOCX)
Microarray data. The transcription profiles for S. cerevisiae and C. albicans, when shifted to xylose from a glucose starter culture, were determined with two-color cDNA probe hybridizations. The tables show the average signal ratio (xylose/glucose, or no-sugar/glucose) of the replicates with the corresponding P-value for each ORF present on the microarrays. The complete data with the experimental details is available at NCBI under GEO accession number GSE50476.
(XLS)
Growth analysis of C. albicans wild type (SC5314) and deletion mutants for xylose reductase (gre3-9) and xylitol dehydrogenase (xyl2-16) in synthetic medium (SC) with 2% xylose (SX) or 2% xylitol (ST). Strains were grown aerobically at 30°C, and the optical densities were measured over a period of 4 days.
(DOCX)
Alignment of fungal and bacterial xylose isomerases.
(DOCX)
Acknowledgments
This is NRC publication 49551.
Funding Statement
This work was supported by grants from the Canadian Institutes of Health Research (CIHR) to MW (MOP42516) and MW and MR (CTP79843), and from the Agricultural Bioproducts Innovation Program (ABIP). This is NRC publication 49551. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Kay J, Weitzman PDJ (1987) Krebs' citric acid cycle : half a century and still turning Kay J, Weitzman PDJ. London: London Biochemical Society. [Google Scholar]
- 2. Trumbly RJ (1992) Glucose repression in the yeast Saccharomyces cerevisiae. Mol Microbiol 6: 15-21. doi: 10.1111/j.1365-2958.1992.tb00832.x. PubMed: 1310793. [DOI] [PubMed] [Google Scholar]
- 3. Carlson M (1999) Glucose repression in yeast. Curr Opin Microbiol 2: 202-207. doi: 10.1016/S1369-5274(99)80035-6. PubMed: 10322167. [DOI] [PubMed] [Google Scholar]
- 4. Leloir LF (1951) The enzymatic transformation of uridine diphosphate glucose into a galactose derivative. Arch Biochem Biophys 33: 186-190. doi: 10.1016/0003-9861(51)90096-3. PubMed: 14885999. [DOI] [PubMed] [Google Scholar]
- 5. Ha SJ, Galazka JM, Kim SR, Choi JH, Yang X et al. (2011) Engineered Saccharomyces cerevisiae capable of simultaneous cellobiose and xylose fermentation. Proc Natl Acad Sci U S A 108: 504-509. doi: 10.1073/pnas.1010456108. PubMed: 21187422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Saitoh S, Tanaka T, Kondo A (2011) Co-fermentation of cellulose/xylan using engineered industrial yeast strain OC-2 displaying both beta-glucosidase and beta-xylosidase. Appl Microbiol Biotechnol 91: 1553-1559. doi: 10.1007/s00253-011-3357-5. PubMed: 21643701. [DOI] [PubMed] [Google Scholar]
- 7. van Maris AJ, Abbott DA, Bellissimi E, van den Brink J, Kuyper M et al. (2006) Alcoholic fermentation of carbon sources in biomass hydrolysates by Saccharomyces cerevisiae: current status. Antonie Van Leeuwenhoek 90: 391-418. doi: 10.1007/s10482-006-9085-7. PubMed: 17033882. [DOI] [PubMed] [Google Scholar]
- 8. Hahn-Hägerdal B, Karhumaa K, Jeppsson M, Gorwa-Grauslund MF (2007) Metabolic engineering for pentose utilization in Saccharomyces cerevisiae. Adv Biochem Eng/Biotechnol 108: 147-177. doi: 10.1007/10_2007_062. PubMed: 17846723. [DOI] [PubMed] [Google Scholar]
- 9. Jeffries TW (2006) Engineering yeasts for xylose metabolism. Curr Opin Biotechnol 17: 320-326. doi: 10.1016/j.copbio.2006.05.008. PubMed: 16713243. [DOI] [PubMed] [Google Scholar]
- 10. Saha BC (2003) Hemicellulose bioconversion. J Ind Microbiol Biotechnol 30: 279-291. doi: 10.1007/s10295-003-0049-x. PubMed: 12698321. [DOI] [PubMed] [Google Scholar]
- 11. Van Vleet JH, Jeffries TW (2009) Yeast metabolic engineering for hemicellulosic ethanol production. Curr Opin Biotechnol 20: 300-306. doi: 10.1016/j.copbio.2009.06.001. PubMed: 19545992. [DOI] [PubMed] [Google Scholar]
- 12. Bera AK, Ho NW, Khan A, Sedlak M (2011) A genetic overhaul of Saccharomyces cerevisiae 424A(LNH-ST) to improve xylose fermentation. J Ind Microbiol Biotechnol 38: 617-626. doi: 10.1007/s10295-010-0806-6. PubMed: 20714780. [DOI] [PubMed] [Google Scholar]
- 13. Deng XX, Ho NW (1990) Xylulokinase activity in various yeasts including Saccharomyces cerevisiae containing the cloned xylulokinase gene. Scientific note. Appl Biochem Biotechnol 24-25: 193-199. doi: 10.1007/BF02920245. PubMed: 2162148. [DOI] [PubMed] [Google Scholar]
- 14. Laluce C, Schenberg AC, Gallardo JC, Coradello LF, Pombeiro-Sponchiado SR (2012) Advances and Developments in Strategies to Improve Strains of Saccharomyces cerevisiae and Processes to Obtain the Lignocellulosic Ethanol-A Review. Appl Biochem Biotechnol 166: 1908-1926. doi: 10.1007/s12010-012-9619-6. PubMed: 22391693. [DOI] [PubMed] [Google Scholar]
- 15. Verho R, Londesborough J, Penttilä M, Richard P (2003) Engineering redox cofactor regeneration for improved pentose fermentation in Saccharomyces cerevisiae. Appl Environ Microbiol 69: 5892-5897. doi: 10.1128/AEM.69.10.5892-5897.2003. PubMed: 14532041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. van Maris AJ, Winkler AA, Kuyper M, de Laat WT, van Dijken JP et al. (2007) Development of efficient xylose fermentation in Saccharomyces cerevisiae: xylose isomerase as a key component. Adv Biochem Eng/Biotechnol 108: 179-204. doi: 10.1007/10_2007_057. PubMed: 17846724. [DOI] [PubMed] [Google Scholar]
- 17. Bettiga M, Bengtsson O, Hahn-Hägerdal B, Gorwa-Grauslund MF (2009) Arabinose and xylose fermentation by recombinant Saccharomyces cerevisiae expressing a fungal pentose utilization pathway. Microb Cell Factories 8: 40. doi: 10.1186/1475-2859-8-40. PubMed: 19630951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Tripathi G, Wiltshire C, Macaskill S, Tournu H, Budge S et al. (2002) Gcn4 co-ordinates morphogenetic and metabolic responses to amino acid starvation in Candida albicans. EMBO J 21: 5448-5456. doi: 10.1093/emboj/cdf507. PubMed: 12374745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Chen DC, Yang BC, Kuo TT (1992) One-step transformation of yeast in stationary phase. Curr Genet 21: 83-84. doi: 10.1007/BF00318659. PubMed: 1735128. [DOI] [PubMed] [Google Scholar]
- 20. Dignard D, Whiteway M (2006) SST2, a regulator of G-protein signaling for the Candida albicans mating response pathway. Eukaryot Cell 5: 192-202. doi: 10.1128/EC.5.1.192-202.2006. PubMed: 16400182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Wilson RB, Davis D, Mitchell AP (1999) Rapid hypothesis testing with Candida albicans through gene disruption with short homology regions. J Bacteriol 181: 1868-1874. PubMed: 10074081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Reuss O, Vik A, Kolter R, Morschhäuser J (2004) The SAT1 flipper, an optimized tool for gene disruption in Candida albicans. Gene 341: 119-127. doi: 10.1016/j.gene.2004.06.021. PubMed: 15474295. [DOI] [PubMed] [Google Scholar]
- 23. Askew C, Sellam A, Epp E, Hogues H, Mullick A et al. (2009) Transcriptional regulation of carbohydrate metabolism in the human pathogen Candida albicans. PLOS Pathog 5: e1000612 PubMed: 19816560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Nantel A, Rigby T, Hogues H, Whiteway M (2006) Microarrays for studying pathology in Candida albicans. In: Kavanaugh K. Hoboken NJ: Wiley Press. [Google Scholar]
- 25. Jeffries TW, Van Vleet JR (2009) Pichia stipitis genomics, transcriptomics, and gene clusters. FEMS Yeast Res 9: 793-807. doi: 10.1111/j.1567-1364.2009.00525.x. PubMed: 19659741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Wohlbach DJ, Kuo A, Sato TK, Potts KM, Salamov AA et al. (2011) Comparative genomics of xylose-fermenting fungi for enhanced biofuel production. Proc Natl Acad Sci U S A 108: 13212-13217. doi: 10.1073/pnas.1103039108. PubMed: 21788494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Leandro MJ, Fonseca C, Gonçalves P (2009) Hexose and pentose transport in ascomycetous yeasts: an overview. FEMS Yeast Res 9: 511-525. doi: 10.1111/j.1567-1364.2009.00509.x. PubMed: 19459982. [DOI] [PubMed] [Google Scholar]
- 28. Noble SM, Johnson AD (2005) Strains and strategies for large-scale gene deletion studies of the diploid human fungal pathogen Candida albicans. Eukaryot Cell 4: 298-309. doi: 10.1128/EC.4.2.298-309.2005. PubMed: 15701792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Schwartz K, Wenger JW, Dunn B, Sherlock G (2012) APJ1 and GRE3 homologs work in concert to allow growth in xylose in a natural Saccharomyces sensu stricto hybrid yeast. Genetics 191: 621-632. doi: 10.1534/genetics.112.140053. PubMed: 22426884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Wenger JW, Schwartz K, Sherlock G (2010) Bulk segregant analysis by high-throughput sequencing reveals a novel xylose utilization gene from Saccharomyces cerevisiae. PLOS Genet 6: e1000942 PubMed: 20485559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Jeffries TW, Grigoriev IV, Grimwood J, Laplaza JM, Aerts A et al. (2007) Genome sequence of the lignocellulose-bioconverting and xylose-fermenting yeast Pichia stipitis. Nat Biotechnol 25: 319-326. doi: 10.1038/nbt1290. PubMed: 17334359. [DOI] [PubMed] [Google Scholar]
- 32. Liang L, Zhang J, Lin Z (2007) Altering coenzyme specificity of Pichia stipitis xylose reductase by the semi-rational approach CASTing. Microb Cell Factories 6: 36. doi: 10.1186/1475-2859-6-36. PubMed: 18028553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Watanabe S, Abu Saleh A, Pack SP, Annaluru N, Kodaki T et al. (2007) Ethanol production from xylose by recombinant Saccharomyces cerevisiae expressing protein-engineered NADH-preferring xylose reductase from Pichia stipitis. Microbiology 153: 3044-3054. doi: 10.1099/mic.0.2007/007856-0. PubMed: 17768247. [DOI] [PubMed] [Google Scholar]
- 34. Watanabe S, Pack SP, Saleh AA, Annaluru N, Kodaki T et al. (2007) The positive effect of the decreased NADPH-preferring activity of xylose reductase from Pichia stipitis on ethanol production using xylose-fermenting recombinant Saccharomyces cerevisiae. Biosci Biotechnol Biochem 71: 1365-1369. doi: 10.1271/bbb.70104. PubMed: 17485825. [DOI] [PubMed] [Google Scholar]
- 35. Walfridsson M, Bao X, Anderlund M, Lilius G, Bülow L et al. (1996) Ethanolic fermentation of xylose with Saccharomyces cerevisiae harboring the Thermus thermophilus xylA gene, which expresses an active xylose (glucose) isomerase. Appl Environ Microbiol 62: 4648-4651. PubMed: 8953736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Brat D, Boles E, Wiedemann B (2009) Functional expression of a bacterial xylose isomerase in Saccharomyces cerevisiae. Appl Environ Microbiol 75: 2304-2311. doi: 10.1128/AEM.02522-08. PubMed: 19218403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Gillum AM, Tsay EY, Kirsch DR (1984) Isolation of the Candida albicans gene for orotidine-5'-phosphate decarboxylase by complementation of S. cerevisiae ura3 and E. coli pyrF mutations. Molecular General Genet MGG 198: 179-182. doi: 10.1007/BF00328721. [DOI] [PubMed] [Google Scholar]
- 38. Brachmann CB, Davies A, Cost GJ, Caputo E, Li J et al. (1998) Designer deletion strains derived from Saccharomyces cerevisiae S288C: a useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast 14: 115-132. doi: 10.1002/(SICI)1097-0061(19980130)14:2. PubMed: 9483801. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Alignments of A) xylose reductases and B) xylose dehydrogenases of C. albicans, S. stipitis and S. cerevisiae.
(DOCX)
Microarray data. The transcription profiles for S. cerevisiae and C. albicans, when shifted to xylose from a glucose starter culture, were determined with two-color cDNA probe hybridizations. The tables show the average signal ratio (xylose/glucose, or no-sugar/glucose) of the replicates with the corresponding P-value for each ORF present on the microarrays. The complete data with the experimental details is available at NCBI under GEO accession number GSE50476.
(XLS)
Growth analysis of C. albicans wild type (SC5314) and deletion mutants for xylose reductase (gre3-9) and xylitol dehydrogenase (xyl2-16) in synthetic medium (SC) with 2% xylose (SX) or 2% xylitol (ST). Strains were grown aerobically at 30°C, and the optical densities were measured over a period of 4 days.
(DOCX)
Alignment of fungal and bacterial xylose isomerases.
(DOCX)