

Abstract
Seaweeds are commonly used as functional foods and drugs. A glycoprotein (GP) from the green alga Capsosiphon fulvescens (Cf) has been reported to have anti-tumor activity toward various cancer cells. We previously observed that Cf-GP induced different pathways of apoptosis in AGS human gastric cancer cells. Transforming growth factor (TGF)-β 1 plays an important role in cancer cell migration. Increased TGF-β 1 levels increase the expression of the small GTPases and activate the FAK/PI3K/AKT pathways, resulting in the upregulation of integrin receptor proteins, which mediate the attachment of cells to surrounding tissues, cells or extracellular matrix. Thus, the inhibition of TGF-β 1 signaling would downregulate integrin expression and thereby effectively decrease cell growth and migration. In the present study, we determined the effect of Cf-GP treatment on the proliferation, migration and apoptosis of AGS human gastric cancer cells. To investigate the mechanism by which Cf-GP exerts its anticancer actions, we examined the effect of Cf-GP on the expression levels of TGF-β 1 , FAK, PI3K, AKT, the small GTPases and integrins in AGS cells. Our findings indicate that Cf-GP inhibits AGS cell proliferation and migration by downregulating integrin expression via the TGF-β 1 -activated FAK/PI3K/AKT pathways. These results suggest that Cf-GP may be an important factor in the development of functional foods and therapeutic agents.
Keywords: capsosiphon fulvescens , transforming growth factor-β 1 , FAK/AKT/small GTPase , integrin , cell migration
Introduction
Capsosiphon fulvescens (Cf) is green seaweed that grows mainly in the clean areas of the sea off the coast of Korea. It is commonly used as a traditional health food, and various bioactive effects of Cf have been reported ( 1 ) . A compositional analysis of Cf showed that it contains high levels of proteins, carbohydrates and amino acids ( 2 ) . Specific substances in Cf are known to boost the immune system, bio-activity and have anticancer activities ( 3 – 5 ) . We previously observed the induction of apoptosis in AGS human gastric cancer cells by a glycoprotein of Cf ( 6 , 7 ) . This glycoprotein has also been reported to inhibit cell invasion ( 8 ) .
Transforming growth factor (TGF)-β 1 is a cytokine associated with various human cancers ( 9 , 10 ) involving macrophages, brain cells and keratinocytes. Previous studies have reported that TGF-β 1 modulates cell migration, invasion and proliferation in gastric cancer cells ( 11 ) . TGF-β 1 induces the overexpression of growth factor focal adhesion kinase (FAK) protein in several cancers, activates phosphatidylinositol 3-kinase (PI3K)/AKT and small GTPase proteins ( 12 ) , and upregulates integrin proteins. The small GTPases, which include Rho A, Rho B, Rac-1 and Cdc 42, are involved in the signaling pathways associated with diverse cellular functions, including cell proliferation and migration, in response to different growth factor receptors ( 13 ) . The small GTPases also activate nuclear transcription factor-κB (NF-κB), which upregulates the expression of integrin receptor proteins, thereby contributing to cell migration.
Integrin receptors are located on the cell surface, where they are responsible for the adhesion of cells to extracellular matrix (ECM) proteins such as fibronectin and the transduction of extracellular signals to the cells ( 14 ) . Integrins exist as hetero dimers of two distinct transmembrane glycoprotein chains, called α and β subunits, that are non-covalently linked. The integrin family consists of 24 different αβ heterodimers ( 15 ) . The binding of an integrin receptor to its extracellular ligand causes a signal to be relayed into the cell, resulting in the regulation of specific gene expression ( 16 ) . The binding of integrin receptors to ECM molecules also produces cell adhesion, which is critical for cell migration, proliferation and differentiation.
The present study investigated the possible relationship between the anticancer activity of the C. fulvescens glycoprotein (Cf-GP) and the downregulation of integrin expression via the TGF-β 1 -activated PI3K/AKT/small GTPases pathway in AGS human gastric cancer cells. First, we established the effect of Cf-GP treatment on the proliferation, migration and apoptosis of AGS cells. Then, to investigate the mechanism by which Cf-GP may exert its anticancer activity, we examined the effect of Cf-GP on the expression levels of TGF-β 1 , the small GTPases, and integrins in the AGS cells.
Materials and methods
Preparation of Cf-GP
The C. fulvescens used in this experiment was purchased in 2010 in Republic of Korea. The Cf powder (40 g) was diluted with water (1 liter) and stirred for 3 h at 80°C in a heating mantle. The solution was clarified by centrifugation at 1,500 × g for 15 min at 4°C. Three volumes of 95% ethanol were added to the solution, and precipitates were removed by vacuum filtration. The supernatant was mixed with 80% ammonium sulfate and stirred for 24 h, followed by dialysis (Por Membrane MW 3,500 Da, Spcectrum Laboratories Inc., Rancho Dominguez, CA, USA) for one day at 4°C to remove salts. The concentrated solution was distributed into 1.5-ml tubes and stored at −70°C until use. These samples were named Cf-GP.
Cell culture
AGS human gastric cancer cell line (American Type Culture Collection, Manassas, VA, USA) was cultured in RPMI-1640 medium with 10% fetal bovine serum (FBS; Hyclone, Logan, UT, USA), 100 U/ml penicillin, and 100 mg/ml streptomycin, at a temperature of 37°C in a humidified atmosphere of 5% CO 2 . The cells were cultured to 80% confluence in 100-mm dishes. The medium was replaced daily.
Cell proliferation assay
AGS cell proliferation was measured using a CellTiter 96 ® aqueous non-radioactive cell proliferation assay (Promega, Madison, WI, USA), which is based on the cleavage of 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfonyl)-2H-tetrazolium (MTS) into a formazan product soluble in tissue culture medium. The cells were seeded onto 96-well plates at 2×10 4 cells/well and the medium was replaced with serum-free medium (SFM) after culture for 24 h. After another 24 h, the medium was replaced with SFM containing Cf-GP (5, 10 and 20 μ g/ml), followed by incubation for 24 h. For the assay, MTS solution was added to the cells in each well and allowed to react for 30 min at 37°C. The absorbance of the solution in each well was measured at 490 nm using a microplate reader (Benchmark microplate reader; Bio-Rad Laboratories, Hercules, CA, USA).
Cell migration assay
AGS cells were seeded onto 100-mm dishes and grown to 80% confluence. The medium was replaced with SFM, and the cells were cultured for 24 h, after which the cells were wounded by scraping with a pipette tip. The medium was replaced with SFM containing Cf-GP (5, 10 and 20 μ g/ml), and the cells were cultured for 24 h. Wound closure was determined from photographs taken using a microscope at ×200 magnification.
Apoptosis assay
The level of apoptosis induced by Cf-GP treatment was determined using a Muse™ Annexin V and Dead Cell kit (EMD Millipore Co., Hayward, CA, USA). Cells were cultured in 6-well seeds to 60% confluency, and then the medium was replaced with SFM or SFM containing Cf-GP (5, 10 and 20 μ g/ml). After 24 h, the cells were collected in 1% FBS-RPMI-1640 medium, mixed with the Muse Annexin V and Dead Cell Reagent, and analyzed using a Muse Cell Analyzer (EMD Millipore Co.).
mRNA expression assay
The mRNA expression levels of specific genes were evaluated by reverse-transcription polymerase chain reaction (RT-PCR). AGS cells were seeded onto 6-well plates at 2×10 4 cells/well and were cultured for 24 h, after which the medium was replaced with SFM containing Cf-GP (5, 10 and 20 μ g/ml) for 24 h. Total RNA was isolated from the cells using TRIzol reagent (Invitrogen Co., Carlsbad, CA, USA), and total RNA was converted to cDNA using oligo(dT) primers (iNtRON Biotechnology Inc., Seongnam, Korea). For PCR amplification, the cDNA and specific primers ( Table I ) were added to 2X TOPsimple™ DyeMIX-nTaq (Enzynomics, Inc., Daejoen, Korea) and 0.1% diethylpyrocarbonate (DEPC) water. The amplified products were analyzed on 1% agarose gels stained with RedSafe™ nucleic acid staining solution (iNtRON Biotechnology, Inc.).
Table I.
Oligonucleotide sequences of the primer pairs used for RT-PCR.
| Name | Sequence of primers (5′→3′) |
|---|---|
| TGF-β 1 | S: GCA-GAA-CCC-AAA-AGC-CAG-AGT-G |
| A: CCA-TAA-CTA-CCG-TGG-AGG-TTG-A | |
| FAK | S: TTC-ATT-ATT-TTG-AAA-GCA-ATA-GT |
| A: CAA-CCC-AAC-TTC-AAA-GCA-ATT-TC | |
| Rho A | S: CTC-ATA-GTC-TTC-AGC-AAG-GAC-CAG-TT |
| A: ATC-ATT-CCG-AAG-ATC-CTT-CTT-ATT | |
| Rho B | S: ATG-GCG-GCC-ATC-CGC-AAG-AAG-C |
| A: TCA-TAG-CAC-CTT-GCA-GCA-GTT-G | |
| Rac-1 | S: GGA-CAC-AGC-TGG-ACA-AGA-AGA |
| A: GGA-CAG-AGA-ACC-GCT-CGG-ATA | |
| Cdc 42 | S: CGA-CCG-CTA-AGT-TAT-CCA-CAG |
| A: GCA-GCT-AGG-ATA-GCC-TCA-TCA | |
| PI3K | S: AGG-AGC-GGT-ACA-GCA-AAG-AA |
| A: GCC-GAA-CAC-CTT-TTT-GAG-TC | |
| Akt | S: CAA-CTT-CTC-TGT-GGC-GCA-GTG |
| A: GAC-AGG-TGG-AAG-AAC-AGC-TCG | |
| IκB | S: TGG-ATG-AAC-TGC-GTG-GTG-CAG |
| A: GCA-GAA-GTG-TCC-CTG-TTC-CAG | |
| NF-κB | S: TCA-GGG-AAT-ATC-CAC-CTA-TCA-CTT-CAG |
| A: CAT-CAG-CAG-CAG-CCA-TGT-ACT-CTT-CAC | |
| Integrin α ν | S: GAA-GCT-TCA-TCT-CCA-GTC-CCT |
| A: TGG-GTA-GGG-CTG-TTT-GTC-ATC-ATA | |
| Integrin β 1 | S: GAC-CTG-CCT-TGG-TGT-CTG-TGC |
| A: AGC-AAC-CAC-ACC-AGC-TAC-AAT | |
| Integrin β 3 | S: CCC-TCG-AAA-ACC-CCT-GCT-AT |
| A: TTA-GCG-TCA-GCA-CGT-GTT-TGT-AG | |
| Integrin β 5 | S: GGC-TGG-GAC-GTC-ATT-CAG-AT |
| A: AGC-TGG-AAG-GTG-GTC-TTG-TCA | |
| β-actin | S: CGT-ACC-ACT-GGC-ATC-GTG |
| A: GTG-TTG-GCG-TAC-AGG-TCT-TTG |
S, sense; A, antisense.
Western blot analysis
AGS cells in 100-mm dishes were cultured to 80% confluence and then incubated in SFM for 4 h. Fresh SFM containing Cf-GP (5, 10 and 20 μ g/ml) was added to the cells, and the incubation continued for 24 h, after which the cells were washed with phosphate-buffered saline (PBS) and mixed with lysis extraction buffer [20 mM Tris (pH 7.5), 150 mM NaCl, 1% Triton X-100, 1 mM EDTA, 1 mM EGTA, 2.5 mM sodium pyrophosphate, 1 mM β-glycerophosphate, 1 mM sodium orthovanadate, 1 μ g/ml aprotinin, 1 μ g/ml leupeptin, 1 μ g/ml pepstatin A, 0.25% Na-deoxycholate, and 1 mM PMSF]. For western blot analysis, the cell lysates were electrophoresed in 10–15% polyacrylamide gels, and the resolved proteins were transferred to Immobilon-P transfer membrane (Millipore Co., Billerica, MA, USA). The membranes were blocked with 1% bovine serum albumin in TBS-T [10 mM Tris-HCl (pH 7.5), 150 mM NaCl, and 0.1% Tween-20] at room temperature, followed by incubation with the following specific primary antibodies (diluted 1:1,000): anti-TGF-β 1 , anti-FAK, anti-PI3K, anti-AKT, anti-Rho A, anti-Rho B, anti-Rac-1, anti-Cdc 42, anti-NF-κB, anti-IκB, anti-integrin α ν , anti-integrin β 1 , anti-integrin β 3 , and anti-integrin β 5 . All primary antibodies were purchased from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA). The secondary antibody was horse-radish peroxidase conjugated goat, mouse or rabbit antibody (1:10,000) (GE Healthcare Bio-Sciences, Piscataway, NJ, USA). Immunoreactive bands were detected with SuperSignal West Pico Chemiluminescent substrate (Thermo Fisher Scientific Inc., Rockford, IL, USA) and visualized on Kodak X-ray film.
Statistical analysis
Data were analyzed using ANOVA. Values of p<0.05 on Duncan’s multiple range test indicated a significant difference between each groups. The results are presented as means ± SD. All analyses were performed with SPSS software (ver. 10.0; SPSS Inc., Chicago, IL, USA).
Results
Cf-GP inhibits AGS cell proliferation
The effects of Cf-GP on AGS cell proliferation were examined using the MTS assay. AGS cells treated with Cf-GP at 5, 10 and 20 μ g/ml showed dose-dependent inhibition of cell proliferation, with a maximum inhibition of 50% at 20 μ g/ml ( Fig. 1A ). We found Cf-GP dose-dependent effect of AGS cell growth inhibition by MTS assay. In addition we determined the toxicity of the normal cells. Treatment with Cf-GP had no toxic effects on the normal human intestinal epithelial IEC-6 cells ( Fig. 1B ).
Figure 1.
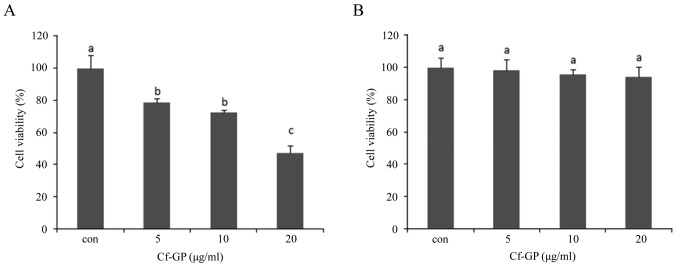
Cf-GP inhibits the proliferation of AGS cells. Cells were treated with Cf-GP (5, 10 and 20 μ g/ml) for 24 h. As a control, (A) AGS cell and (B) IEC-6 cell were similarly treated with Cf-GP. Cell proliferation was determined using a MTS assay. Values represent means ± SD. p<0.05 by ANOVA. Values with different letters are significantly different according to Duncan’s multiple range test.
Cf-GP inhibits AGS cell migration
The effect of Cf-GP on AGS cell migration was examined using a wound-healing assay. As results, in the case of control AGS cells group over time, the mobility of cells in the wounded area was found to increase. But, AGS cells treated with Cf-GP (5, 10 and 20 μ g/ml) exhibited dose-dependent inhibition of migration into the cell-wounded zone on 100-mm dishes, indicating a Cf-GP-induced inhibition of cell mobility ( Fig. 2 ).
Figure 2.

Cf-GP inhibits wound-healing migration of AGS cells. Cells were cultured in 100-mm dishes for 24 h, and then the cell layer was wounded by scraping. The medium was replaced with medium containing 1% FBS and Cf-GP (5, 10 and 20 μ g/ml). The denuded zone of cells was photographed using a microscope at ×200 magnification, and the degree of recovery was measured.
Cf-GP dose-dependently increases cellular apoptosis in AGS cells
AGS cells were treated with Cf-GP (5, 10 and 20 μ g/ml), and apoptotic and necrotic cells were detected by Annexin V and 7-aminoactinomycin D (AAD) staining, respectively ( 17 ) . In this assay, cells in the early stage of apoptosis are Annexin V-positive and 7-AAD-negative, and those in late apoptosis are Annexin V-positive and 7-AAD-positive. In the present study, Cf-GP treatment increased the percentage of apoptotic AGS cells, in a dose-dependent manner. Control cells comprised 5.25% apoptotic cells, 0.85% necrotic cells and 93.90% living cells ( Fig. 3 ). Treatment with 5, 10 and 20 μ g/ml Cf-GP for 24 h resulted in 17.50, 26.65 and 42.68% apoptotic cells, respectively.
Figure 3.

Cf-GP induces apoptosis of the AGS cells. Cells cultured in 6-well plates were treated with Cf-GP (5, 10 and 20 μ g/ml) and then collected in medium containing 1% FBS. The percentages of apoptotic and necrotic cells were determined using a Muse Annexin V and Dead Cell kit as described in the Materials and methods. Cells in the early stage of apoptosis are Annexin V-positive and 7-AAD-negative, and those in late apoptosis are Annexin V-positive and 7-AAD-positive.
Cf-GP dose-dependently alters the expression of TGF-β 1 and small GTPases
The TGF-β 1 receptor activates small GTPases and the FAK/PI3K/AKT pathways. The overexpression of FAK/PI3K/AKT can be induced by several other growth factors as well, and the Rho family of small GTPases plays a critical role in cancer cell growth and migration ( 18 , 19 ) . Thus, the altered expression of proteins associated with growth factors regulates cancer cell growth and migration ( 20 ) . In the present study, the protein and mRNA expression levels of TGF-β 1 , FAK, PI3K, AKT and the small GTPases Rho A, Rho B, Rac-1 and Cdc 42 were determined by western blot analysis and RT-PCR analysis of AGS cells treated with Cf-GP (5, 10 and 20 μ g/ml) for 24 h. Treatment with Cf-GP dose-dependently downregulated the protein ( Fig. 4A ) and mRNA expression levels ( Fig. 4B ) of TGF-β 1 , FAK, PI3K, AKT and the small GTPases Rho A, Rac-1 and Cdc 42. In contrast, Cf-GP treatment upregulated the protein and mRNA expression levels of Rho B, a small GTPase that is involved in the inhibition of cancer cell growth.
Figure 4.

Cf-GP dose-dependently alters the expression of growth-regulating factors in AGS cells. (A) Protein and (B) mRNA expression levels of TGF-β 1 , FAK, PI3K, AKT and the small GTPases Rho A, Rho B, Rac-1 and Cdc 42 in Cf-GP-treated (5, 10 and 20 μ g/ml). AGS cells were determined by western blot analysis and RT-PCR analysis, respectively, as described in Materials and methods.
Cf-GP dose-dependently downregulates the expression of NF-κB and IκB
NF-κB activation is necessary for cancer cell growth and migration ( 21 ) . NF-κB inhibits activity through forming an NF-κB/IκB complex by upregulation of IκB, however, almost all cancer cells show downregulation IκB. Thus, degradation of NF-κB/IκB induced nuclear translocation of NF-κB movement. The protein and mRNA expression levels of the transcription factor NF-κB and its regulatory protein IκB in AGS cells treated with Cf-GP (5, 10 and 20 μ g/ml) for 24 h were analyzed by western blot analysis and RT-PCR. Cf-GP treatment downregulated the expression of NF-κB and upregulation of IκB ( Fig. 5 ) in a dose-dependent manner. This indicates that the Cf-GP induced inhibits degradation of IκB. Therefore, inhibition of the translocated NF-κB is by the stable NF-κB/IκB complex.
Figure 5.
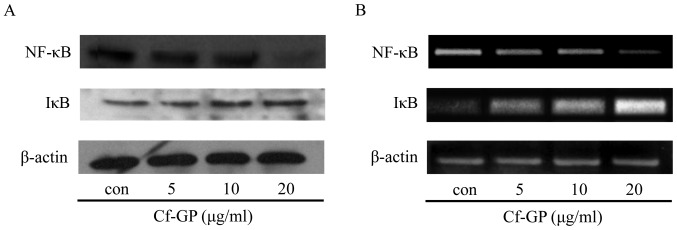
Cf-GP dose-dependently downregulates the expression of NF-κB and downregulates of IκB. (A) Protein and (B) mRNA expression levels of NF-κB and IκB in Cf-GP-treated (5, 10 and 20 μ g/ml) AGS cells were determined by western blot analysis and RT-PCR analysis, respectively, as described in the Materials and methods.
Cf-GP dose-dependently downregulates the expression of integrins
Integrin proteins play important roles in cancer cells growth and migration. Also, involvment in cancer cells angiogenesis, invasion and incessant proliferation was induced. Integrins are a family of heterodimers of α and β subunit, and cancer cells of the specific expression of each subunit have been reported ( 22 ) . Among them, integrin α ν , β 1 , β 3 and β 5 are known to be overexpressed in gastric cancer cells ( 23 ) . We have confirmed inhibition of growth related factor and transcription factor via reduction of TGF-β 1 by Cf-GP. Therefore, we performed expression of integrin proteins and mRNA levels through western blot analysis and RT-PCR. The same conditions as in the treatment by Cf-GP (5, 10 or 20 μ g/ml) for 24 h, were used. The protein ( Fig. 6A ) and mRNA expression levels ( Fig. 6B ) of integrins α ν , β 1 , β 3 and β 5 were reduced by Cf-GP treatment, in a dose-dependent manner.
Figure 6.

Cf-GP dose-dependently downregulates the expression of integrins α ν , β 1 , β 3 and β 5 . Protein and mRNA expression levels of integrins α ν , β 1 , β 3 and β 5 in Cf-GP-treated (5, 10 and 20 μ g/ml) AGS cells were determined by (A) western blot analysis and (B) RT-PCR analysis, respectively, as described in Materials and methods.
Discussion
Gastric cancer is common in many countries, particularly in Asia, and is caused by irregular eating habits, stress and environmental factors. Several studies ( 24 – 27 ) have reported the anticancer effects of various marine algae, including the green alga C. fulvescens , which is commonly consumed in Asia. We previously observed that a glycoprotein of C. fulvescens (Cf-GP) induced apoptosis in AGS human gastric cancer cells through the Fas signaling pathway ( 6 , 7 ) .
In the present study, we showed that Cf-GP also inhibits the migration and proliferation of AGS cells, as well as down-regulates the expression of several growth-related proteins, including TGF-β 1 , FAK, PI3K, AKT and the small GTPases Rho A, Rac-1 and Cdc 42. Cf-GP treatment also resulted in the downregulated expression of NF-κB, IκB, and integrins α ν , β 1 , β 3 and β 5 . Conversely, Cf-GP upregulated the expression of the small GTPase Rho B, which is involved in the inhibition of cell growth and the induction of apoptosis.
TGF-β 1 is a secreted cytokine that participates in the regulation of cell migration, growth, apoptosis and differentiation. TGF-β 1 acts through its receptor to induce FAK expression in several cancers, to activate the PI3K/AKT/small GTPase pathway ( 13 ) , and to upregulate integrin receptor expression ( 28 ) . The small GTPases Rho A, Rho B, Rac-1, and Cdc 42 are involved in signaling pathways associated with cell proliferation and migration in response to various growth factor receptors ( 13 ) . They also activate NF-κB, which induces the expression of integrin receptors, thereby contributing to cell migration. Integrin receptors are critical for cell adhesion and cell attachment-dependent growth, migration, invasion and metastasis ( 29 , 30 ) . In particular, integrin has α and β, two subunits of hetero-copolymer and these are associated with the progression of a variety of human cancer cells. Attachment in independent growth is a characteristic of transformed cells, but cancer cell growth and migration depend on the interaction of the cell adhesion receptor integrins and matrix. Therefore, integrins promoted cell proliferation by attachment-dependent effects.
Specific integrin heterodimers are expressed in different cancer cells, and the growth and migration of the cells depend on the interactions between the integrins and their extracellular ligands.
First, we observed the effects of Cf-GP in AGS cell viability by MTS assay. Treatment by Cf-GP (5, 10 or 20 μ g/ml) for 24 h inhibited the AGS cell growth and at the highest concentration of 20 μ g/ml in the apoptotic cells by approximately 50% reduction compared to control group and no toxicity to IEC-6 normal cell ( Fig. 1 ). We used a wound-healing assay to confirm cell migration and observed the denuded zone through a microscope. The gap (denuded zone) between the cells was inhibited dose-dependently by Cf-GP ( Fig. 2 ). In addition, we performed Annexin V staining assay for cell apoptosis rate by Muse Annexin V and Dead Cell kit. The apoptosis rate increased 42.68% in final Cf-GP concentration (20 μ g/ml) compared with the control group ( Fig. 3 ). We found TGF-β 1 decreased FAK/PI3K/AKT/small GTPase expression by using western blot analysis and RT-PCR ( Fig. 4 ). This result shows that the inhibition of FAK inhibited TGF-β 1 . Increased cancer cell proliferation was observed when FAK/PI3K/AKT/small GTPase were activated. Cf-GP-treated cells exhibited significant downregulation of FAK/PI3K/AKT/small GTPase proteins (Rho A, Rac-1, Cdc 42) and mRNA levels compared to the control group, but activation of Rho B induced cancer cells apoptosis. In this study, Rho B increased by Cf-GP ( Fig. 4 ). The results of the transcription factor NF-κB and IκB decreased due to inhibition of these growth factors. Activation of the NF-κB and IκB induces promotion of cancer cell migration and variety of intracellular factors ( 31 ) . We found upregulation of IκB induced downregulation NF-κB through inhibition of the growth factor and TGF-β 1 by the Cf-GP effects ( Fig. 5 ). The integrin α ν pair with multiple integrin subunit β (β 1 , β 3 , β 5 ) and upregulation of integrin proteins induced migration of cancer cells. The integrin expression through the activation of transcription factors to increase was seen from previous experiment, thus, the reduction of the NF-κB and increase of IκB were indentified. Therefore, we performed expression of integrins, and confirmed that Cf-GP induced downregulation of integrin α ν , β 1 , β 3 and β 5 ( Fig. 6 ).
Collectively, our findings suggest that Cf-GP inhibits AGS gastric cancer cell migration and proliferation by downregulating integrin expression via the inhibition of TGF-β 1 -activated FAK/PI3K/AKT pathways. Cf-GP may be an important factor in the development of functional foods and therapeutic agents.
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2012R1A6A1028677).
References
- 1. Park HY , Lim CW , Kim YK , Yoon HD , Lee KJ . Immunostimulating and anticancer activities of hot water extract from Capsosiphon fulvescens . J Korean Soc Appl Biol Chem . 2006 ; 49 : 343 – 348 . [Google Scholar]
- 2. Jung KJ , Jung CH , Pyeun JH , Choi YJ . Changes of food components in Mesangi ( Capsosiphon fulvescens ), Gashiparae ( Enteromorpha prolifera ), and Cheonggak ( Codium fragile ) depending on harvest times . J Korean Soc Food Sci Nutr . 2005 ; 34 : 687 – 693 . [Google Scholar]
- 3. Na YS , Kim WJ , Kim SM , Park JK , Lee MS , Kim SO , Synytsya A , Park YI . Purification, characterization and immunostimulating activity of water-soluble polysaccharide isolated from Capsosiphon fulvescens . Int Immunopharmacol . 2010 ; 10 : 364 – 370 . doi: 10.1016/j.intimp.2009.12.011. [DOI] [PubMed] [Google Scholar]
- 4. Karnjanapratum S , Tabarsa M , Cho M , You S . Characterization and immunomodulatory activities of sulfated polysaccharides from Capsosiphon fulvescens . Int J Biol Macromol . 2012 ; 51 : 720 – 729 . doi: 10.1016/j.ijbiomac.2012.07.006. [DOI] [PubMed] [Google Scholar]
- 5. Hwang HJ , Kwon MJ , Kim IH , Nam TJ . The effect of polysaccharide extracted from the marine alga Capsosiphon fulvescens on ethanol administration . Food Chem Toxicol . 2008 ; 46 : 2653 – 2657 . doi: 10.1016/j.fct.2008.04.027. [DOI] [PubMed] [Google Scholar]
- 6. Kim YM , Kim IH , Nam TJ . Induction of apoptosis signaling by a glycoprotein of Capsosiphon fulvescens in AGS cell . Kor J Fish Aquat Sci . 2011 ; 44 : 216 – 224 . [Google Scholar]
- 7. Kim YM , Kim IH , Nam TJ . Induction of apoptosis signaling by glycoprotein of Capsosiphon fulvescens in human gastric cancer (AGS) cells . Nutr Cancer . 2012 ; 64 : 761 – 769 . doi: 10.1080/01635581.2012.683228. [DOI] [PubMed] [Google Scholar]
- 8. Kim YM , Kim IH , Nam TJ . Inhibition of AGS human gastric cancer cell invasion and proliferation by Capsosiphon fulvescens glycoprotein . Mol Med Rep . 2013 ; 8 : 11 – 16 . doi: 10.3892/mmr.2013.1492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Levy L , Hill CS . Alterations in components of the TGF-beta superfamily signaling pathways in human cancer . Cytokine Growth Factor Rev . 2006 ; 17 : 41 – 58 . doi: 10.1016/j.cytogfr.2005.09.009. [DOI] [PubMed] [Google Scholar]
- 10. Serra R , Crowley MR . Mouse models of transforming growth factor beta impact in breast development and cancer . Endocr Relat Cancer . 2005 ; 12 : 749 – 760 . doi: 10.1677/erc.1.00936. [DOI] [PubMed] [Google Scholar]
- 11. Hua F , Zhongliang H , Jifang W , Kuansong W , Ying L . TGF-beta promotes invasion and metastasis of gastric cancer cells by increasing fascin1 expression via ERK and JNK signal pathways . Acta Biochim Biophys Sin (Shanghai) 2009 ; 41 : 648 – 656 . doi: 10.1093/abbs/gmp053. [DOI] [PubMed] [Google Scholar]
- 12. Roarty K , Baxley SE , Crowley MR , Frost AR , Serra R . Loss of TGF-β or Wnt5a results in an increase in Wnt/β-catenin activity and redirects mammary tumor phenotype . Breast Cancer Res. 2009;11:R19. doi: 10.1186/bcr2244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Lee YC , Cheng TH , Lee JS , Chen JH , Liao YC , Fong Y , Wu CH , Shih YW . Nobiletin, a citrus flavonoid, suppresses invasion and migration involving FAK/PI3K/Akt and small GTPase signals in human gastric adenocarcinoma AGS cells . Mol Cell Biochem . 2011 ; 347 : 103 – 115 . doi: 10.1007/s11010-010-0618-z. [DOI] [PubMed] [Google Scholar]
- 14. Humphries MJ . Integrin structure . Biochem Soc Trans . 2000 ; 28 : 311 – 339 . [PubMed] [Google Scholar]
- 15. Fong YC , Hsu SF , Wu CL , Li TM , Kao ST , Tsai FJ , Chen WC , Liu SC , Wu CM , Tang CH . Transforming growth factor-β1 increases cell migration and β1 integrin up-regulation in human lung cancer cells . Lung Cancer . 2009 ; 64 : 13 – 21 . doi: 10.1016/j.lungcan.2008.07.010. [DOI] [PubMed] [Google Scholar]
- 16. Woodhouse EC , Chuaqui RF , Liotta LA . General mechanisms of metastasis . Cancer . 1997 ; 80 : 1529 – 1537 . doi: 10.1002/(sici)1097-0142(19971015)80:8+<1529::aid-cncr2>3.3.co;2-#. [DOI] [PubMed] [Google Scholar]
- 17. Krysho DV , Vanden BT , D’Herde K , Vandenabeele P . Apoptosis and necrosis: detection, discrimination and phagocytosis . Methods . 2008 ; 44 : 205 – 221 . doi: 10.1016/j.ymeth.2007.12.001. [DOI] [PubMed] [Google Scholar]
- 18. Sahai E , Marshall CJ . Rho-GTPases and cancer . Nat Rev Cancer . 2002 ; 2 : 133 – 142 . doi: 10.1038/nrc725. [DOI] [PubMed] [Google Scholar]
- 19. Fritz G , Just I , Kaina B . Rho GTPases are over-expressed in human tumors . Int J Cancer . 1999 ; 81 : 682 – 687 . doi: 10.1002/(sici)1097-0215(19990531)81:5<682::aid-ijc2>3.0.co;2-b. [DOI] [PubMed] [Google Scholar]
- 20. Leivonen SK , Kähäri VM . Transforming growth factor-beta signaling in cancer invasion and metastasis . Int J Cancer . 2007 ; 121 : 2119 – 2124 . doi: 10.1002/ijc.23113. [DOI] [PubMed] [Google Scholar]
- 21. Boukerche H , Su ZZ , Emdad L , Sarkar D , Fisher PB . mda-9/Syntenin regulates the metastatic phenotype in human melanoma cells by activating nuclear factor-kappaB . Cancer Res . 2007 ; 67 : 1812 – 1822 . doi: 10.1158/0008-5472.CAN-06-3875. [DOI] [PubMed] [Google Scholar]
- 22. Peng C , Liu X , Liu E , Xu K , Niu W , Chen R , Wang J , Zhang Z , Lin P , Wang J , Agrez M , Niu J . Norcantharidin induces HT-29 colon cancer cell apoptosis through the α ν β 6 -extracellular signal-related kinase signaling pathway . Cancer Sci . 2009 ; 100 : 2302 – 2308 . doi: 10.1111/j.1349-7006.2009.01320.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Zhang ZY , Xu KS , Wang JS , Yang GY , Wang W , Wang JY , Niu WB , Liu EY , Mi YT , Niu J . Integrin α ν β 6 acts as a prognostic indicator in gastric carcinoma . Clin Oncol (R Coll Radiol) 2008 ; 20 : 61 – 66 . doi: 10.1016/j.clon.2007.09.008. [DOI] [PubMed] [Google Scholar]
- 24. Go H , Hwang HJ , Nam TJ . A glycoprotein from Laminaria japonica induces apoptosis in HT-29 colon cancer cells . Toxicology In Vitro . 2010 ; 24 : 1546 – 1553 . doi: 10.1016/j.tiv.2010.06.018. [DOI] [PubMed] [Google Scholar]
- 25. Hwang HJ , Kim IH , Nam TJ . Effect of a glycoprotein from Hizikia fusiformis on acetaminophen-induced liver injury . Food Chem Toxicol . 2008 ; 46 : 3475 – 3481 . doi: 10.1016/j.fct.2008.08.032. [DOI] [PubMed] [Google Scholar]
- 26. Cho EK , Yoo SK , Choi YJ . Inhibitory effects of maesaengi ( Capsosiphon fulvescens ) extracts on angiotensin converting enzyme and α-glucosidase . Korea J Life Sci . 2011 ; 21 : 811 – 818 . [Google Scholar]
- 27. Kown MJ , Nam TJ . A polysaccharide of the marine alga Capsosiphon fulvescens induces apoptosis in AGS gastric cancer cells via an IGF-IR-mediated PI3K/Akt pathway . Cell Biol Int . 2007 ; 31 : 768 – 775 . doi: 10.1016/j.cellbi.2007.01.010. [DOI] [PubMed] [Google Scholar]
- 28. Wei YY , Chen YJ , Hsiao YC , Huang YC , Lai TH , Tang CH . Osteoblasts-derived TGF-β1 enhance motility and integrin upregulation through Akt, ERK, and NF-κB-dependent pathway in human breast cancer cells . Mol Carcinog . 2008 ; 47 : 526 – 537 . doi: 10.1002/mc.20411. [DOI] [PubMed] [Google Scholar]
- 29. Cooper CR , Chay CH , Pienta KJ . The role of α ν β 3 in prostate cancer progression . Neoplasia . 2002 ; 4 : 191 – 194 . doi: 10.1038/sj.neo.7900224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Kumar CC . Integrin α ν β 3 as a therapeutic target for blocking tumor-induced angiogenesis . Curr Drug Targets . 2003 ; 4 : 123 – 131 . doi: 10.2174/1389450033346830. [DOI] [PubMed] [Google Scholar]
- 31. Helbig G , Christopherson KW , II , Bhat-Nakshatri P , Kumar S , Kishimoto H , Miller KD , Broxmeyer HE , Naksharti H . NF-κB promotes breast cancer cell migration and metastasis by inducing the expression of the chemokine receptor CXCR4 . J Biol Chem . 2003 ; 278 : 21631 – 21638 . doi: 10.1074/jbc.M300609200. [DOI] [PubMed] [Google Scholar]