

Abstract
Under physiological conditions, epidermal growth factor receptor (EGFR) tyrosine kinase activity is tightly controlled through the coordinated action of both positive and negative regulators. Aberrant EGFR activation occurs frequently in many cancer types and the endogenous EGFR feedback inhibitor, Mig6/RALT, is more efficiently phosphorylated by oncogenic EGFR variants. We have utilized expressed protein ligation to generate semisynthetic Tyr394-phosphorylated and unphosphorylated forms of the Mig6 protein and show that phosphorylation of Mig6 reduces its ability to inhibit purified, near full-length EGFR (tEGFR). We also demonstrate that the kinetic parameters of tEGFR are similar whether solubilized in detergent or reconstitutued in nanodisc bilayers. These findings suggest a mechanism by which EGFR and its family members evade negative regulation by Mig6 under pathological conditions.
The epidermal growth factor receptor (EGFR) signaling cascade has been one of the most intensively studied signaling pathways in human biology and is the target of several clinically approved anti-cancer drugs 1, 2. EGFR and its family members HER2/Neu, HER3, and HER4 are composed of several domains including an extracellular growth factor-binding domain, a transmembrane domain, an intracellular tyrosine kinase domain, and a C-terminal tail that can be autophosphorylated. Binding of EGF to EGFR stimulates its tyrosine kinase activity via an allosteric mechanism involving formation of an asymmetric dimer of intracellular kinase domains3. This asymmetric dimer juxtaposes the C-lobe of a donor kinase domain with the N-lobe of an acceptor (activated) kinase domain. Activating mutations of EGFR are frequently observed in cancer, including the common alterations L858R and exon 19 (aa746–750) deletion4, 5.
The protein Mig6 (RALT) was identified as an endogenous inhibitor of EGFR signaling6, 7, and loss of Mig6 function can augment tumor development in animal models6. Biochemical analysis of Mig6 has revealed that a 77 amino acid (aa) region (aa 336–412) of Mig6 is responsible for EGFR inhibition. This 77 aa region can be further subdivided into two segments, segment 1 (aa 336–364) and segment 2 (aa 365–412). An X-ray crystal structure of Mig6 segment 1 complexed with the EGFR kinase domain shows an extended surface of interaction of segment 1 with the kinase C-lobe that spans a region critical for forming the asymmetric dimer with the N-lobe of the other subunit 8. Mig6 segment 1 thus physically obstructs asymmetric dimer formation, thereby inhibiting EGF-mediated EGFR activation. Although the structure of Mig6 segment 2 is uncertain, the Mig6 segment 1+2 polypeptide is ~100-fold more potent than Mig6 segment 1 alone in inhibiting EGFR kinase activity8, 9. It is speculated that Mig6 segment 2 interacts with the EGFR kinase domain along a path approaching the active site, thereby augmenting the affinity of the C-lobe targeted segment 1 for EGFR8. The oncogenic L858R and exon 19 (aa746–750) deletion EGFR variants appear to be resistant to Mig6 inhibition, presumably because their asymmetric dimer is more strongly favored9.
Whether and how Mig6 itself is regulated has been unresolved. Mass spectrometry studies showed that Mig6 is tyrosine phosphorylated in cells, and phosphorylation of Tyr394 is enhanced in lung cancer cell lines carrying oncogenic L858R or exon 19 (aa 746–750) deletion mutations as well as in cells that overexpress HER2/Neu 10, 11. Biochemical studies with purified proteins showed that EGFR can directly tyrosine phosphorylate Mig69. As Tyr394 is located within segment 2 of Mig6, it is conceivable that Mig6 phosphorylation could modulate its inhibition of EGFR, which is the subject of this study.
Using in vitro kinase assays with purified, solubilized tail-truncated EGFR (aa 25–1022, tEGFR), His6-Mig6 segment 1+2 appears to be phosphorylated on multiple Tyr residues, since Y394F/Y395F Mig6 was still efficiently phosphorylated by tEGFR (Supplementary Figure 1). Producing a singly-phosphorylated preparation of pTyr394-Mig6 using tEGFR was thus not possible. We turned instead to protein semisynthesis and the method of expressed protein ligation12–15. In this approach, a recombinant protein fragment containing a C-terminal thioester is generated by the action of a stalled intein and this thioester is chemoselectively ligated to an N-Cys containing peptide to generate a standard amide bond at the ligation junction. After several attempts, we were unable to get adequate soluble His6-Mig6-intein fusion protein production. We next investigated a GST-Mig6 (aa 336–391) intein fusion and this gave acceptable production of GST-Mig6 segment 1+2 after ligation with NCys synthetic peptides aa 392–412 with and without phosphorylation at Tyr394 (Figure 1a, Supplementary Figure 2). We were able to obtain ~80% conversion to ligated products by carrying out the reaction at 4°C. The minor unligated GST-Mig6 may have arisen from non-specific proteolysis or thioester hydrolysis that would prevent protein ligation. Semisynthetic proteins after purification by ion exchange and size-exclusion chromatographies showed a minor impurity of unligated GSTMig6, equally represented in both phosphorylated and unphosphorylated preparations (Figure 1b). Since we showed that unligated GST-Mig6 (IC50 > 3 μM) is only a weak inhibitor of EGFR relative to ligated GST-Mig6 segment 1+2 (Supplementary Figure 3a), we presume the minor contaminant of unligated GSTMig6 to be inconsequential. Semisynthetic GST-Mig6 in phosphorylated and unphosphorylated forms showed the correct molecular weights using MALDI-TOF mass spectrometry (see Experimental Methods). The phosphorylation status of semisynthetic phospho-GST-Mig6 was further confirmed by western blotting with anti-pTyr antibody (4G10) as shown in Figure 1c.
Figure 1.

Preparation of the GST-Mig6 semisynthetic proteins by expressed protein ligation (EPL). (a) Scheme for EPL to generate the semisynthetic GST-Mig6 segment 1+2 containing phosphorylated tyrosine in a full stoichiometry. The residue Tyr394 that can be phosphorylated by tyrosine kinases is highlighted in red. (b) Coomassie stained 10% SDS-PAGE of unligated and semisynthetic GST-Mig6 proteins. (c) The incorporation of phosphorylated Tyr394 into semisynthetic GST-Mig6 was verified with western blot using the antiphosphotyrosine antibody 4G10. The same amount of unligated(1), unphosphorylated (2), and phosphorylated (3) Mig6 proteins obtained via expressed protein ligation were analyzed by 10% SDS-PAGE and probed with 4G10 antibody.
To analyze the effects of semisynthetic GST-Mig6 on EGFR activity, we used solubilized, purified tEGFR, which we have described previously16, as well as tEGFR reconstituted in membrane bilayer nanodiscs17. Nanodiscs were used to test whether detergent solubilization of tEGFR alters its enzymatic properties relative to a native membrane environment (Figure 2a). The steady-state kinetic parameters including apparent kcat and Km values with a synthetic peptide substrate of tEGFR were similar when comparing the nanodisc and detergent-solubilized forms (Figure 2b, Supplementary Figure 4). Likewise, inhibition by unphosphorylated semisynthetic GST-Mig6 segment 1+2 was similar for the nanodisc and solubilized tEGFRs with an IC50 of 0.3–0.4 μM (average of two independent assays) (Figure 3 and Supplementary Figure 3b). Moreover, since we replaced native aa 390–2 (Ser- Ser-Thr) with Pro-Gly-Cys to facilitate the expressed ligation reaction for GST-Mig6 semisynthesis (see Methods), it was important to assess the effects of these substitutions on the potency of Mig6 inhibition of tEGFR. Fortunately, these aa 390– 2 replacements were shown to be well-tolerated as the unphosphorylated, semisynthetic GST-Mig6 showed nearly identical inhibitory potency (IC50 of 0.3–0.4 μM) toward tEGFR as the standard recombinant His6-Mig6 segment 1+2 that was wild type in aa 390–2 (Supplementary Figure5). Remarkably, Tyr394- phosphorylation of GST-Mig6 conferred a sharp reduction in inhibitory potency, with IC50 ~3 μM for pTyr394-GST-Mig6 against both solubilized and nanodiscreconstituted tEGFR (Figure 3 and Supplementary Figure 3b). This weakened potency was comparable to that of unligated GST-Mig6 (aa 336–391) (Supplementary Figure 3a), underscoring that pTyr394 phosphorylation is quite disruptive of the contributions of Mig6 segment 2 to EGFR inhibition. Since the segment 1 region (aa 336–364) in both unphosphorylated and phosphorylated GSTMig6 is identical, and this region alone has previously been shown to bind to the C-lobe of EGFR kinase domain with weak inhibition against EGFR (IC50 ~100 μM)8, 9, it is speculated that the interaction between EGFR kinase domain and Mig6 segment is maintained after phosphorylation of Tyr394 (Figure 4). Interestingly, the His6-Mig6 mutant Y394E (IC50 of 0.74 μM) was only 2-fold less potent than unphosphorylated wt His6-Mig6 (IC50 of 0.35 μM), indicating that the Glu sidechain is a poor mimic of a phosphotyrosine in this case (Supplementary Figure 5). Thus, it would have been difficult to discern the authentic function of Tyr394-phosphorylation of Mig6 using mutagenesis and standard cell-based transfection experiments.
Figure 2.
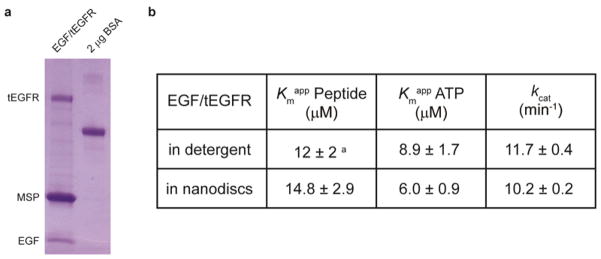
Reconstitution of purified EGF/tEGFR into phospholipid bilayer nanodiscs. (a) WT tEGFR (aa 25–1022) purified from transiently transfected HEK293 GnTicells in the presence of 0.03% dodecyl maltoside was reconstituted into nanodiscs assembled with membrane scaffold protein (MSP) and egg phosphatidylcholine lipids. The reconstituted EGF/tEGFR was subjected to 4–20% gradient SDS-PAGE and Coomassie staining to verify the incorporation of EGF/tEGFR into nanodiscs. BSA is used as a standard to quantify the protein concentrations. (b) The kinetic parameters of reconstituted EGF/tEGFR and EGF/tEGFR in detergent were compared. aPreviously reported data. Enzymatic parameters (±s.d.) were calculated from duplicate experiments.
Figure 3.
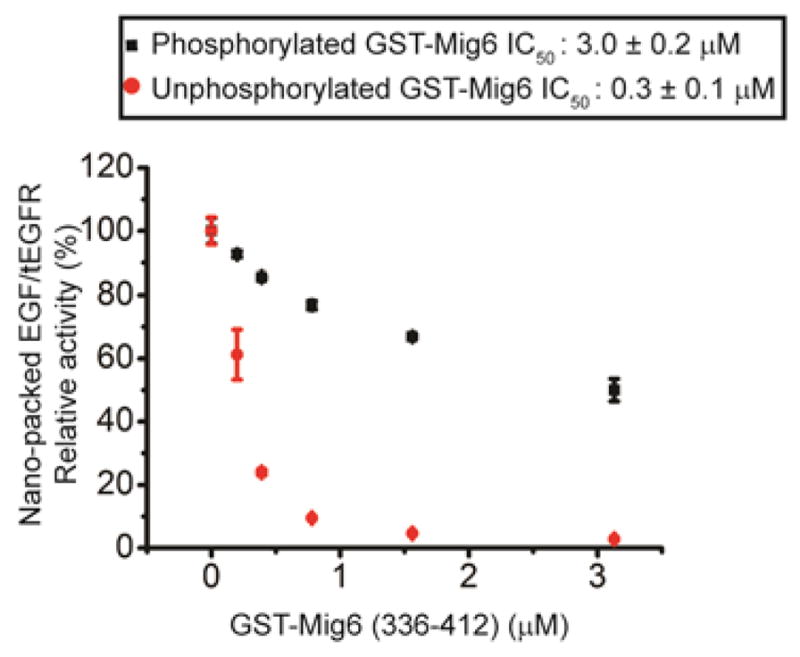
Distinct inhibitory effects of semisynthetic unphosphorylated and phosphorylated GST-Mig6 on EGF/tEGFR reconstituted in nanodiscs. IC50 values of Mig6 were determined by quantifying the relative kinase activity of EGF/tEGFR in the presence of varying concentrations of unmodified and phosphorylated GST-Mig6 (0, 0.20, 0.39, 0.78, 1.56, and 3.13 μM).
Figure 4.
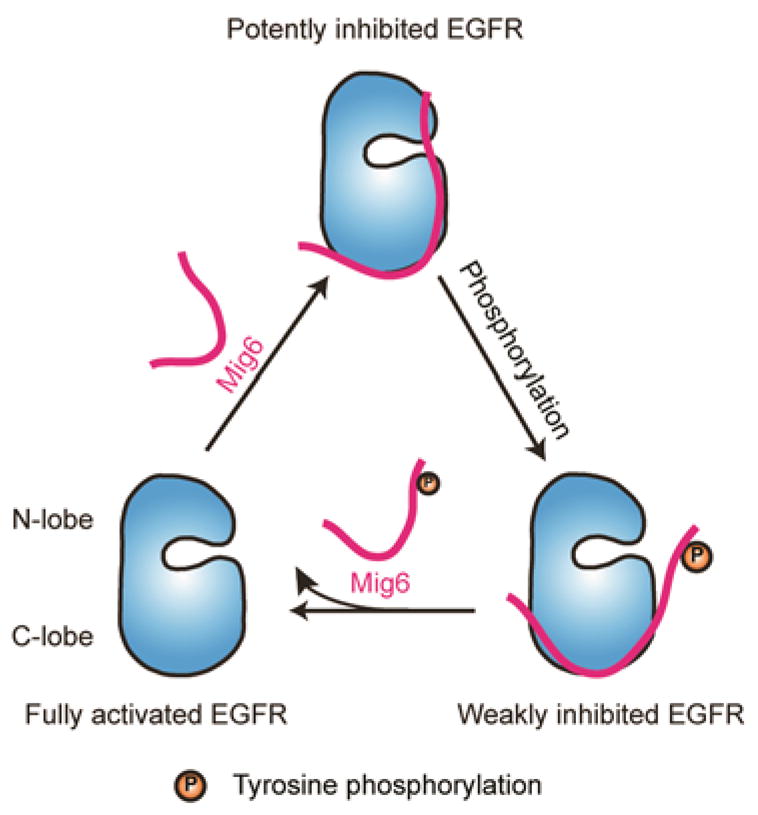
Feedback regulation of EGFR activation by Mig6, an endogenous inhibitor of the EGFR family, is finely tuned by phosphorylation state of Mig6. Mig6 inhibits EGFR by blocking the C-lobe asymmetric dimer interface. After Mig6 is phosphorylated by tyrosine kinases, its inhibitory potency is compromised, suggesting a potential strategy for cancer cells to escape the negative regulation of activated EGFR by Mig6. The EGFR kinase domain is depicted as blue and segment 1+2 of Mig6 is depicted in purple.
These results suggest that tyrosine kinase-catalyzed phosphorylation of Mig6 likely represents a feedback mechanism to antagonize its inhibition of EGFR (Figure 4). As noted, at least two EGFR family members when hyperactivated by mutation (EGFR) or elevated by increased expression (HER2/Neu) appear to target Tyr394 for phosphorylation in cells10, 11. Thus, Mig6 phosphorylation provides a mechanism for these kinases to override the inhibition mediated by Mig6. Such an effect would augment the signaling and cell proliferative properties associated with the EGFR family kinases, all of which are believed to be susceptible to Mig6 inhibition. Thus, the activation of one family member can bolster the signaling from each of the others. Whether other phosphorylation sites on Mig6 also modulate its function will be an interesting direction for future study.
While the structural details of the interactions of Mig6 segment 2 with EGFR are not well understood, it is reasonable to speculate that segment 2 extends near the kinase domain active site where Tyr394 can be targeted for modification. Furthermore, we hypothesize that Tyr394 phosphorylation of Mig6 causes electrostatic repulsion or sterically clashes with the substrate ATP and/or the catalytic base aspartate.. As has been seen with other kinases, phosphorylation of this substrate peptide motif can reduce affinity for the kinase, enhancing product release18, 19. Thus, it will be interesting to investigate whether phosphorylation of endogenous protein inhibitors of kinases represents a general mechanism for overriding their inhibition. Such on/off switches may help account for the highly rapid cellular phospho-protein dynamics often observed in signaling cascades20, 21.
These studies also provide a cautionary lesson for using Glu residues as mimics of phosphotyrosine22, 23. While the negative charge of the Glu can nominally substitute for the anionic pTyr, its smaller size, reduced charge, and different shape potentially limit its utility in this regard.
The use of nanodisc reconstituted tEGFR shows that the key features of the kinase activity seen in the detergent solubilized tEGFR are recapitulated in the lipid bilayer-embedded protein. Thus, the transmembrane helices, which have recently been suggested to be crucial for transmitting signaling information from the ectodomain to the intracellular region24–27, appear to be functional in the detergent solubilized systems. It will be interesting in future studies to determine if different lipid compositions in nanodiscs affect EGFR kinase activity.
Methods
Plasmid construction
DNA encoding the N-terminal truncated Mig6 segment 1+2 (56 amino acids, residues 336–391) with an N-terminal GST-fusion and two mutations, S390P and S391G, was subcloned into the pTYB2 vector (New England Biolads) to yield pTYB2-GST-Mig6 plasmid. Initial semisynthetic Mig6 experiments showed that the two native C-terminal Ser-Ser residues of Mig6 gave poor yields of thioester product and impeded subsequent ligation. As also noted in the NEB brochure for IMPACT vectors, C-terminal Pro-Gly has been reported to be very efficient for thioester generation and expressed protein ligation12, 28. Plasmid pT7HT-His6-Mig6 encoding Mig6 segment 1+2 (77 aa, residues 336–412) with an Nterminal His6 tag was prepared previously and contained WT 390–2 residues9. Using Quikchange mutagenesis, the mutations Y394E, Y394F, Y395F were introduced into His6-Mig6 segment 1+2, respectively.
Peptide synthesis
The C-terminal fragment of Mig6 (21 aa, residues 392–412) and EGFR peptide substrate (Biotin-RAHEEIYHFFFAKKK-COOH) were synthesized using the standard Fmoc solid phase peptide synthesis strategy described previously29. Two Mig6 peptides were synthesized, one without any post-translational modification, termed the unphosphorylated Mig6 peptide (CHYYLLPERPPYLDKYEKFFR), and the other containing a phosphorylated tyrosine residue to replace the native Tyr394, termed the phosphorylated Mig6 peptide (CHpYYLLPERPPYLDKYEKFFR). After C-18 reversed phase HPLC purification, synthetic peptides were >95% pure and structures were confirmed by mass spectrometry.
Mig6 semisynthesis
Plasmid pTYB2-GST-Mig6 was transformed into E. coli BL21 (DE3) codon plus cells. Transformed cells were cultured at 37 °C until A600 reached 0.6, and then the culture was incubated at 16°C for 20 h in the presence of 500 μM IPTG to induce expression of GST-Mig6 (aa 336–391)-intein-CBD fusion protein. Cell pellets were resuspended in lysis buffer (50 mM HEPES pH 7.4, 400 mM NaCl, 1 mM EDTA, 10% (v/v) glycerol, 1 tablet of protease inhibitor cocktail (Roche)) and lysed by passage through a French Pressure cell three times. After centrifugation at 27,000g for 30 min to remove cell debris, the supernatant was collected and incubated with chitin beads (New England Biolabs) for affinity purification. The chitin beads bound with GST-Mig6-intein-CBD fusion protein was washed thoroughly with 1 M NaCl in washing buffer (50 mM HEPES pH 7.4, 1 mM EDTA, 0.1% (v/v) Triton X-100), followed by one quick wash with 1 M urea in washing buffer. After equilibration with three times the bead volume of cleavage buffer (50 mM HEPES pH 7.4, 150 mM NaCl, 1 mM EDTA), the chitin beads were incubated with 200 mM 2-mercaptoethane sulfonate (MESNA, Sigma-Aldrich) in cleavage buffer at 25 °C for 16 h to release the GST-Mig6 protein with a C-terminal thioester. 0.5 mM phosphorylated Mig6 peptide (CHpYLLPERPPYLDKYEKFFR) was added to the cleaved GST-Mig6 thioester solution and the ligation reaction was allowed to proceed for 48 h at 4°C. Upon ligation reaching 80% completion as estimated by Commassie-stained SDS-PAGE, the semisynthetic phosphorylated GST-Mig6 seg 1+2 was further purified with Mono Q ion exchange chromatography and Superdex 75 size-exclusion chromatography. The unphosphorylated and phosphorylated forms of semisynthetic GST-Mig6 seg 1+2 were obtained using the same conditions. The incorporation of the phosphorylated tyrosine residue was verified with western blot using the antiphosphotyrosine antibody 4G10 (Millipore). The purity and concentration of GST-Mig6 seg 1+2 were determined with SDS-PAGE Coomassie staining and Bradford assay using bovine serum albumin as a standard. The unligated GST-Mig6 was obtained by omitting the peptide ligation step. To confirm semisynthetic GST-Mig6 with mass spectrometry, GST-Mig6 was further purified by reversed-phase HPLC on a C-4 column. The peak corresponding to pure GST-Mig6 was collected and analyzed by MALDI-TOF. The masses of the semisynthetic proteins were internally calibrated versus the unligated GST-Mig6 protein (calcd. m/z: 32,505). The unphosphorylated semisynthetic GST-Mig6 showed m/z 35,287 (calcd. m/z: 35,283) and the phosphorylated semisynthetic GST-Mig6 showed m/z 35,334 (calcd. m/z 35,363). The estimated standard error on these m/z measurements is +/− 50 Da.
Packaging of EGF/tEGFR into nanodiscs and EGFR kinase assays
Our protocol for packaging EGF/tEGFR into nanodiscs is modified from the methods of the Sligar and Springer groups30, 31. L-α-phosphatidylcholine isolated from chicken eggs (Avanti Polar Lipids) that had been suspended in chloroform was dried under nitrogen to produce a film on the interior of a disposable borosilicate glass culture tube. Residual chloroform was removed by keeping this film under vacuum overnight. This lipid film was resuspended in a sodium cholate solution (200 mM sodium cholate, 20 mM Tris-HCl, 0.1 M NaCl, 0.5 mM EDTA, pH 7.4) to a molar ratio of cholate to lipid of 2:1. Detergent solubilized EGF/tEGFR, expressed and purified as described above, and membrane scaffold protein (MSP1E3D1) were added to this solution to a final molar ratio of EGF/tEGFR to MSP1E3D1 to lipid to cholate of 1:10:1590:3180. These molar ratios resulted in packaging of EGF/tEGFR in ~10% of nanodiscs formed. This solution was diluted with buffer (20 mM Tris-HCl, 0.1 M NaCl, 0.5 mM EDTA, pH 7.4) to the desired final volume for the packaging reaction and was incubated for 60 min at 4°C. 250 mg of Bio-Beads (BioRad), polystyrene adsorbent beads that remove detergent from solution, were added per mL of reaction volume and incubated overnight at 4°C. Biobeads were removed by centrifugation, as described previously30, and the final reaction mixture was filtered and stored at 4°C until use.
Supplementary Material
Acknowledgments
We are grateful for support from the NIH.
Abbreviations
- EGFR
epidermal growth factor receptor
- EPL
expressed protein ligation
Footnotes
Supporting Information Available: This material is available free of charge via the Internet at http://pubs.acs.org.
References
- 1.Lemmon MA, Schlessinger J. Cell signaling by receptor tyrosine kinases. Cell. 2010;141:1117–1134. doi: 10.1016/j.cell.2010.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hynes NE, Lane HA. ERBB receptors and cancer: the complexity of targeted inhibitors. Nat Rev Cancer. 2005;5:341–354. doi: 10.1038/nrc1609. [DOI] [PubMed] [Google Scholar]
- 3.Zhang X, Gureasko J, Shen K, Cole PA, Kuriyan J. An allosteric mechanism for activation of the kinase domain of epidermal growth factor receptor. Cell. 2006;125:1137–1149. doi: 10.1016/j.cell.2006.05.013. [DOI] [PubMed] [Google Scholar]
- 4.Paez JG, Janne PA, Lee JC, Tracy S, Greulich H, Gabriel S, Herman P, Kaye FJ, Lindeman N, Boggon TJ, Naoki K, Sasaki H, Fujii Y, Eck MJ, Sellers WR, Johnson BE, Meyerson M. EGFR mutations in lung cancer: Correlation with clinical response to gefitinib therapy. Science. 2004;304:1497–1500. doi: 10.1126/science.1099314. [DOI] [PubMed] [Google Scholar]
- 5.Pao W, Miller V, Zakowski M, Doherty J, Politi K, Sarkaria I, Singh B, Heelan R, Rusch V, Fulton L, Mardis E, Kupfer D, Wilson R, Kris M, Varmus H. EGF receptor gene mutations are common in lung cancers from “never smokers” and are associated with sensitivity of tumors to gefitinib and erlotinib. Proc Natl Acad Sci U S A. 2004;101:13306–13311. doi: 10.1073/pnas.0405220101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ferby I, Reschke M, Kudlacek O, Knyazev P, Pante G, Amann K, Sommergruber W, Kraut N, Ullrich A, Fassler R, Klein R. Mig6 is a negative regulator of EGF receptor-mediated skin morphogenesis and tumor formation. Nat Med. 2006;12:568–573. doi: 10.1038/nm1401. [DOI] [PubMed] [Google Scholar]
- 7.Anastasi S, Baietti MF, Frosi Y, Alema S, Segatto O. The evolutionarily conserved EBR module of RALT/MIG6 mediates suppression of the EGFR catalytic activity. Oncogene. 2007;26:7833–7846. doi: 10.1038/sj.onc.1210590. [DOI] [PubMed] [Google Scholar]
- 8.Zhang XW, Pickin KA, Bose R, Jura N, Cole PA, Kuriyan J. Inhibition of the EGF receptor by binding of MIG6 to an activating kinase domain interface. Nature. 2007;450:741–745. doi: 10.1038/nature05998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wang ZH, Longo PA, Tarrant MK, Kim K, Head S, Leahy DJ, Cole PA. Mechanistic insights into the activation of oncogenic forms of EGF receptor. Nat Struct Mol Biol. 2011;18:1388–1393. doi: 10.1038/nsmb.2168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Guha U, Chaerkady R, Marimuthu A, Patterson AS, Kashyap MK, Harsha HC, Sato M, Bader JS, Lash AE, Minna JD, Pandey A, Varmus HE. Comparisons of tyrosine phosphorylated proteins in cells expressing lung cancer-specific alleles of EGFR and KRAS. Proc Natl Acad Sci U S A. 2008;105:14112–14117. doi: 10.1073/pnas.0806158105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wolf-Yadlin A, Kumar N, Zhang Y, Hautaniemi S, Zaman M, Kim HD, Grantcharova V, Lauffenburger DA, White FM. Effects of HER2 overexpression on cell signaling networks governing proliferation and migration. Mol Syst Biol. 2006;2:54. doi: 10.1038/msb4100094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Muir TW, Sondhi D, Cole PA. Expressed protein ligation: a general method for protein engineering. Proc Natl Acad Sci U S A. 1998;95:6705– 6710. doi: 10.1073/pnas.95.12.6705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Szewczuk LM, Tarrant MK, Cole PA. Protein Phosphorylation by Semisynthesis: From Paper to Practice. Methods Enzymol. 2009;462:1–24. doi: 10.1016/S0076-6879(09)62001-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Vila-Perello M, Muir TW. Biological applications of protein splicing. Cell. 2010;143:191–200. doi: 10.1016/j.cell.2010.09.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hackenberger CP, Schwarzer D. Chemoselective ligation and modification strategies for peptides and proteins. Angew Chem Int Ed Engl. 2008;47:10030–10074. doi: 10.1002/anie.200801313. [DOI] [PubMed] [Google Scholar]
- 16.Qiu C, Tarrant MK, Boronina T, Longo PA, Kavran JM, Cole RN, Cole PA, Leahy DJ. In vitro enzymatic characterization of near full length EGFR in activated and inhibited states. Biochemistry. 2009;48:6624–6632. doi: 10.1021/bi900755n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bayburt TH, Sligar SG. Membrane protein assembly into Nanodiscs. FEBS Lett. 2010;584:1721–1727. doi: 10.1016/j.febslet.2009.10.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kim K, Cole PA. Kinetic Analysis of a Protein Tyrosine Kinase Reaction Transition State in the Forward and Reverse Directions. J Am Chem Soc. 1998;120:6851–6858. [Google Scholar]
- 19.Cook PF, Neville ME, Jr, Vrana KE, Hartl FT, Roskoski R., Jr Adenosine cyclic 3′,5′-monophosphate dependent protein kinase: kinetic mechanism for the bovine skeletal muscle catalytic subunit. Biochemistry. 1982;21:5794–5799. doi: 10.1021/bi00266a011. [DOI] [PubMed] [Google Scholar]
- 20.Qiao Y, Molina H, Pandey A, Zhang J, Cole PA. Chemical rescue of a mutant enzyme in living cells. Science. 2006;311:1293–1297. doi: 10.1126/science.1122224. [DOI] [PubMed] [Google Scholar]
- 21.Kleiman LB, Maiwald T, Conzelmann H, Lauffenburger DA, Sorger PK. Rapid phospho-turnover by receptor tyrosine kinases impacts downstream signaling and drug binding. Mol Cell. 2011;43:723–737. doi: 10.1016/j.molcel.2011.07.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Xia F, Li J, Hickey GW, Tsurumi A, Larson K, Guo D, Yan SJ, Silver-Morse L, Li WX. Raf activation is regulated by tyrosine 510 phosphorylation in Drosophila. PLoS Biol. 2008;6:1115–1129. doi: 10.1371/journal.pbio.0060128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tamada M, Farrell DL, Zallen JA. Abl regulates planar polarized junctional dynamics through beta-catenin tyrosine phosphorylation. Dev Cell. 2012;22:309–319. doi: 10.1016/j.devcel.2011.12.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Red Brewer M, Choi SH, Alvarado D, Moravcevic K, Pozzi A, Lemmon MA, Carpenter G. The juxtamembrane region of the EGF receptor functions as an activation domain. Mol Cell. 2009;34:641–651. doi: 10.1016/j.molcel.2009.04.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Scheck RA, Lowder MA, Appelbaum JS, Schepartz A. Bipartite tetracysteine display reveals allosteric control of ligand-specific EGFR activation. ACS Chem Biol. 2012;7:1367–1376. doi: 10.1021/cb300216f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Endres NF, Das R, Smith AW, Arkhipov A, Kovacs E, Huang YJ, Pelton JG, Shan YB, Shaw DE, Wemmer DE, Groves JT, Kuriyan J. Conformational Coupling across the Plasma Membrane in Activation of the EGF Receptor. Cell. 2013;152:543–556. doi: 10.1016/j.cell.2012.12.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Adak S, Yang KS, Macdonald-Obermann J, Pike LJ. The membrane-proximal intracellular domain of the epidermal growth factor receptor underlies negative cooperativity in ligand binding. J Biol Chem. 2011;286:45146–45155. doi: 10.1074/jbc.M111.274175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hackeng TM, Griffin JH, Dawson PE. Protein synthesis by native chemical ligation: expanded scope by using straightforward methodology. Proc Natl Acad Sci U S A. 1999;96:10068–10073. doi: 10.1073/pnas.96.18.10068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Tarrant MK, Rho HS, Xie Z, Jiang YL, Gross C, Culhane JC, Yan G, Qian J, Ichikawa Y, Matsuoka T, Zachara N, Etzkorn FA, Hart GW, Jeong JS, Blackshaw S, Zhu H, Cole PA. Regulation of CK2 by phosphorylation and O-GlcNAcylation revealed by semisynthesis. Nat Chem Biol. 2012;8:262–269. doi: 10.1038/nchembio.771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ritchie TK, Grinkova YV, Bayburt TH, Denisov IG, Zolnerciks JK, Atkins WM, Sligar SG. Chapter 11 - Reconstitution of membrane proteins in phospholipid bilayer nanodiscs. Methods Enzymol. 2009;464:211–231. doi: 10.1016/S0076-6879(09)64011-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mi LZ, Grey MJ, Nishida N, Walz T, Lu C, Springer TA. Functional and structural stability of the epidermal growth factor receptor in detergent micelles and phospholipid nanodiscs. Biochemistry. 2008;47:10314–10323. doi: 10.1021/bi801006s. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.