

Abstract Abstract
Two populations of Astyanax altiparanae (Garutti & Britski, 2000) of the Água dos Patos stream/SP and lake Igapó/PR were analyzed. All individuals showed 2n = 50, however, different karyotypic formulae were observed. The population of the Água dos Patos stream showed 8m +24sm+6st+12a (NF=88) and the population of lake Igapó, 8m+28sm+4st+10a (NF=90). Nucleolus organizing regions (AgNORs) were observed in the terminal position on the short and long arm of different chromosomes of both populations, showing a variation from 3 to 4 chromosomes. Fluorescent in situ hybridization (FISH) using 18S rDNA probes revealed only one pair of chromosomes with fluorescent signals in the terminal site on the short arm in the Igapó lake population, while the population of Água dos Patos stream showed 4 fluorescence terminal signals, characterizing a system of simple and multiple NORs, respectively. 5S rDNA fluorescent signals were detected in the interstitial position of a pair of chromosomes in the two studied populations. Some AgNOR sites revealed to be GC-rich when stained with Chromomycin A3 (CMA3), however, AT positive regions were not observed. The data obtained show that, despite the conservation of the diploid number and location of 5S DNAr, differences in both the distribution of 18S rDNA and karyotypic formula among the populations were found, thus corroborating the existing data on chromosome variability in Astyanax altiparanae that can be significant for cytotaxonomy in this group.
Keywords: Teleostei Characidae, 18S rDNA, 5S rDNA, FISH, karyotypic formula, NORs
Introduction
Astyanax Baird & Girard, 1854, the most common and diversified genus within the family Characidae, has a wide distribution in the Neotropical Region. Due to lack of evidence of monophyly, the genus Astyanax is thought to belong to the Incertae sedis group (Lima et al. 2003). Moreover, the presence of several similarities among the species of this genus allows several species to be considered as a compound from a taxonomic viewpoint (Garutti and Britski 2000).
In Astyanax altiparanae (Garutti & Britski, 2000) from the upper Parana river basin, previously identified as Astyanax bimaculatus (Linnaeus, 1758), all cytogenetic studies accomplished so far reported the occurrence of 2n = 50, with differences in the karyotypic formula among the analyzed populations (Ferreira Neto et al. 2009), which can be explained by the occurrence of chromosome rearrangements, such as pericentric inversions (Domingues et al. 2007).
Besides the differences in karyotypic formula, the nucleolus organizer regions in this species also vary in relation to number and position, as observed by Fernandes and Martins-Santos (2006a), Domingues et al. (2007), Ferreira Neto et al. (2009). However, the same authors found evidence for conservation in relation to the location and number of fluorescent signals of 5S rDNA sites located in the interstitial region of one chromosome pair.
In view of the great chromosome variation observed by other authors in the genus Astyanax, the objective of the present work was to characterize the karyotypes of two populations of Astyanax altiparanae, with emphasis on the location of 18S and 5S DNAr sites, and compare them with data contained in the literature, in an endeavor toward a better understanding of chromosome evolution within this fish group.
Material and methods
Two populations of Astyanax altiparanae were cytogenetically analyzed: twelve specimens (3 males and 9 females) from Água dos Patos stream (22°41'17.7"S; 51° 05'23.9"W ), municipality of Iêpe/SP and sixteen specimens (9 males and 7 females) from Igapó lake (23°19'09.38"S; 51°11'44.72"W ), municipality of Londrina/PR (Fig. 1). Specimens were deposited in the Museum of Zoology of the Universidade Estadual de Londrina (MZUEL). The samples were collected with the permission of Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA), protocol number 11399-1. This study was approved by the ethics committee of our institution and meets all requirements of the Brazilian environmental laws.
Figure 1.

Collection sites of studied specimens. Map of Brazil showing the Paraná and São Paulo states in the selected area (left side). Hydrographic map showing the Paranapanema and Tibagi rivers. In (a) Água dos Patos stream and (b) Igapó lake.
Conventional staining. The specimens were sacrificed after being anesthetized with a solution of benzocaine. Metaphase chromosomes were obtained from kidney cells according to the air drying technique (Bertollo et al. 1978) and stained with 5% Giemsa in phosphate buffer (pH 6.8) The chromosomes were organized as metacentric (m), submetacentric (sm), subtelocentric (st) and acrocentric (a) for the preparation of a karyogram. Metacentric, submetacentric, and subtelocentric chromosomes were considered biarmed and acrocentric uniarmed for determination of the fundamental number (FN) according to Levan et al. (1964).
Fluorescent in situ hybridization (FISH). The in situ hybridization procedure was performed according to Swarça et al. (2001). The 18S rDNA probe of Prochilodus argenteus Agassiz, 1829 (Hatanaka and Galetti Jr. 2004) and 5S rDNA of Leporinus elongatus Valenciennes, 1850 (Martins and Galetti Jr. 1999) were labeled with biotin-14-dATP by nick translation and used as probes. Slides were treated with 30 µl of hybridization mixture (stringency of 70%) containing 100 ng of labeled probe (4 µl), 50% formamide (15 µl), 50% polyethylene glycol (6 µl), 20´ SSC (3 µl), 100 ng of calf thymus DNA (1 µl) and 10% SDS (1 µl). The probe was denatured at 90°C for 10 min, and hybridization was performed overnight at 37°C in a humidified chamber. Post-hybridization washes were carried out in 2´ SSC, 20% formamide in 0.1´ SSC, 0.1´ SSC and 4´ SSC/0.2% Tween 20, all at 42°C. The probe was detected with a solution of 5% BSA and FITC-conjugated avidin (50:0.5, v:v). The post-detection washes were performed in 4´ SSC/0.2% Tween 20 at room temperature. Slides were mounted with 25 µl of a medium composed of 23 ml of DABCO solution (1,4-diaza- bicyclo (2.2.2)-octane (2,3%), 20 mM Tris HCl, pH 8.0, (2%) and glycerol (90%), in distilled water), 1 ml of 2 mg/ml DAPI and 1 ml of 50 mM MgCl2.
Chromosome banding. Active nucleolus organizer regions (NORs) were detected by silver nitrate staining (Howell and Black 1980). The GC- and AT-rich bands were detected with chromomycin A3 (CMA3) and 4’-6-diamino-2-phenylindole (DAPI), respectively, according to Schweizer (1976). The slides were stained with 0.5 mg/mL CMA3 for 1 h, washed in distilled water and sequentially stained with 2 µg/ml DAPI for 15 min. Slides were mounted with a medium composed of glycerol/McIlvaine buffer (pH 7.0) 1:1, plus 2.5 mM MgCl2.
All the images were acquired with a Leica DM 4500 B microscope equipped with a DFC 300FX camera and Leica IM50 4.0 software, and optimized for best constrast and brightness with iGrafx Image software.
Results
The two populations of Astyanax altiparanae showed 2n = 50, however, different karyotypic formula were evidenced. The population of the Água dos Patos stream showed 8m+24sm+6st+12a (NF=88) (Fig. 2a) and the population of Igapó lake 8m+ 28sm+ 4st+10a (NF=90) (Fig. 2b).
Figure 2.
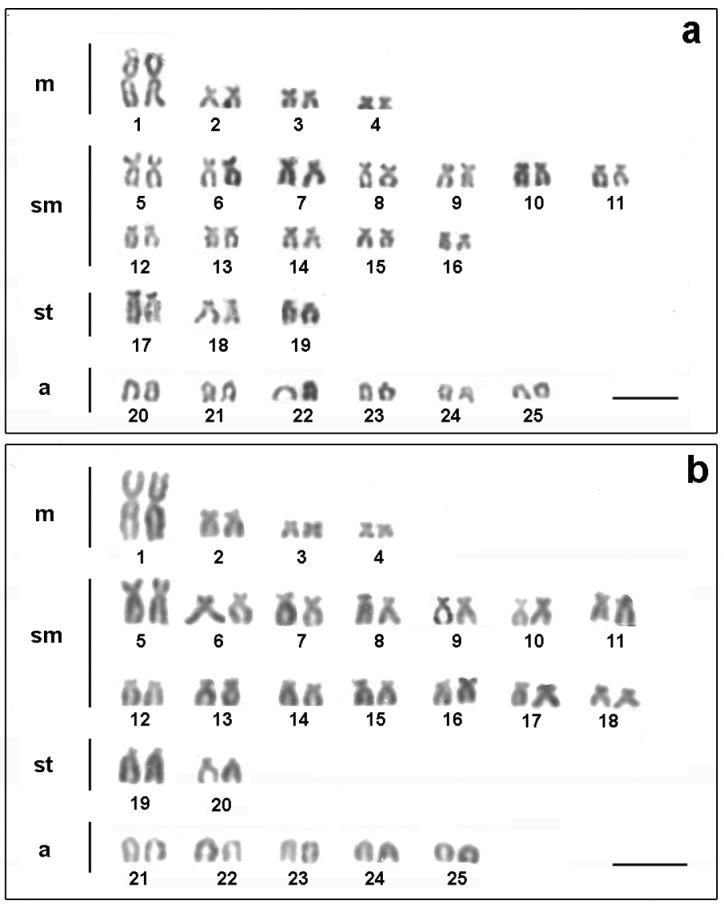
Karyotypes of Astyanax altiparanae after conventional Giemsa staining a Água dos Patos stream b Igapó lake. Bar= 5μm.
Multiple NORs sites were detected in the two populations by silver nitrate impregnation, revealing inter- and intra-individual variations. The population of the Água dos Patos stream showes 2 to 4 AgNORs on the short arm, in two equal medium-sized subtelocentric chromosomes, one of which revealed size heteromorphism observed in all metaphases (Fig. 3a).
Figure 3.
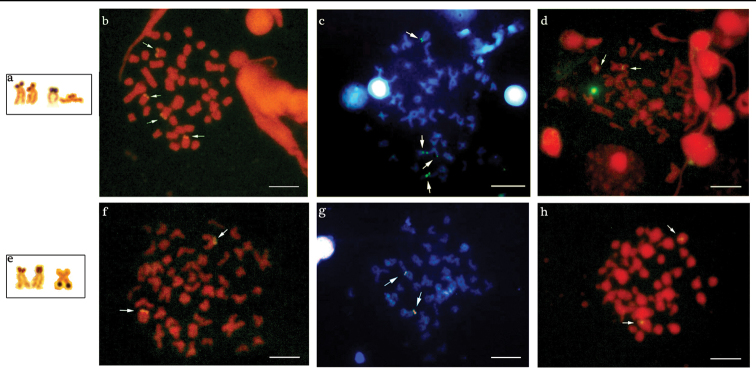
Chromosomes of Astyanax altiparanae bearing AgNORs sites: a Água dos Patos stream e Igapó lake. Metaphase chromosomes with the arrows showing the 18S rDNA, CMA3/DAPI and 5S rDNA sites in Astyanax altiparanae, respectively: b, c, d Água dos Patos stream f, g, h Igapó lake. Bar = 5μm.
One to three AgNORs were detected in the population of Igapó lake: a medium-sized submetacentric chromosome with markings on the long arm and also a medium-sized subtelocentric chromosome with signal on the short arm, presenting a small size heteromorphism. The latter was observed in almost all metaphases (Fig. 3e).
The population of the Água dos Patos stream showed 4 markings on the short arm of two medium-sized subtelocentric chromosome pairs after fluorescent in situ hybridization (FISH) with 18S rDNA probe (Fig. 3b). In the population of Igapó lake, only one pair of medium-sized subtelocentric chromosome presenting fluorescent signals on the short arm was detected (Fig. 3f). Both populations exhibited one pair of chromosomes, bearers of the 5S DNAr sites, in the interstitial position (Fig. 3d and h).
In the population of the Água dos Patos stream, 2 to 4 chromosomes with CMA3+ terminal blocks were detected: a medium-sized subtelocentric pair with signals of size heteromorphism on the short arm, most frequently visualized in the metaphases; a small-sized acrocentric chromosome with a marking on the short arm; and a medium-sized subtelocentric chromosome with a signal on the long arm (Fig. 3c). In the population of Igapó lake, CMA3 markings were detected on the short arm of only one pair of medium-sized subtelocentric chromosomes, which disclosed size heteromorphism (Fig. 3g).
The treatment of the chromosomal preparations of the two populations with DAPI showed a homogeneous staining region, and no regions rich in AT base pairs were detected, as can be seen through the superposition of these fluorochrome, as shown in Fig. 3c-g.
Discussion
Cytogenetic studies in Astyanax altiparanae from the Água dos Patos stream and Igapó lake revealed a conserved diploid number that has been observed within all the analyzed populations of this species (Table 1) so far. However, differences in the karyotypic formula were found in some of these populations, including the one observed in the present study, probably due to occurrence of chromosomal rearrangements, such as pericentric inversions, thus revealing a variability in the karyotypic macrostructure among the species of this group of fish.
Table 1.
Cytogenetic data of different populations of Astyanax altiparanae. FN: fundamental number; m: metacentric; sm: submetacentric; st: subtelocentric; a: acrocentric; AgNORs: nucleolar organizer regions; Ref: Reference; PR: Paraná; SP: São Paulo
| Locality | 2n | FN | Chromosome formulae | AgNORs | CMA3 | 18S/28S | 5S | Ref. |
| Mogi Guaçu river/SP | 50 | 88 | 10m+24sm+4st+12a | 1 | ||||
| 50 | 92 | 6m+24sm+12st+8a | 1 a 5 | 2 | ||||
| Paranapanema river/SP | 50 | 88 | 10m+22sm+6st+12a | 3 | ||||
| Tibagi river/Sertanópolis/PR | 50 | 90 | 10m+22sm+8st+10a | 2 a 5 | 5 | 4 | ||
| Tibagi river/Limoeiro/PR | 50 | 86 | 6m+22sm+8st+14a | 2 a 5 | 5 | 4 | ||
| Tibagi river/Limoeiro/PR | 50 | 88 | 10m+22sm+6st+12a | 2 a 5 | 5 | 4 | ||
| Couro de Boi river/PR | 50 | 88 | 8m+20sm+10st+12a | 1 a 4 | 6 | 4 | ||
| Três Bocas stream/PR | 50 | 92 | 10m+28sm+4st+8a | 1 a 6 | 8 | 4 | ||
| Claro river/PR | 50 | 90 | 10m+26sm+4st+10a | 1 a 4 | 6 | 5 | ||
| 88 | 10m+24sm+4st+12a | 1 a 4 | 6 | 5 | ||||
| 86 | 10m+22sm+4st+14a | 1 a 4 | 6 | 5 | ||||
| Paraná river/PR | 50 | 82 | 32m/sm+18st/a | 3 | 4 | 2 | 6 | |
| Índios river/PR | 50 | 90 | 6m+30sm+4st+10a | 10 | 7 | 7 | ||
| Paraná river/PR | 50 | 88 | 6m+26sm+6st+12a | 2 | 5 | 4 | 2 | 7,9 |
| Tibagi river upper/PR | 50 | 92 | 6m+28sm+8st+8a | 4 | 7 | 7 | 2 | 8 |
| Iguaçu river upper/PR | 50 | 94 | 6m+30sm+8st+6a | 2 | 4 | 2 | 2 | 8 |
| Keçaba brook/PR | 50 | 88 | 6m+26sm+6st+12a | 3 | 7 | 2 | 9 | |
| Tatupeba brook/PR | 50 | 88 | 6m+26sm+6st+12a | 3 | 4 | 2 | 9 | |
| Maringá stream/PR | 50 | 88 | 6m+26sm+6st+12a | 1 | 4 | 2 | 9 | |
| 50 | 88 | 10m+22sm+6st+12a | 2 a 3 | 10 | ||||
| Iguaçu river/PR | 50 | 92 | 10m+26sm+6st+8a | 2 a 5 | 10 | |||
| Monjolinho river/SP | 50 | 90 | 8m+20sm+12st+10a | 2 | 2 | 2 | 11 | |
| Água dos Patos stream/SP | 50 | 88 | 8m+24sm+6st+12a | 2 a 4 | 2 a 4 | 4 | 2 | 12 |
| Igapó lake/PR | 50 | 90 | 8m+28sm+4st+10a | 1 a 3 | 2 | 2 | 2 | 12 |
The great variability in the karyotypic macrostructure is also reflected in other chromosome marks of Astyanax altiparanae. Variability in AgNOR sites with respect to the number, location and types of chromosomes bearers of such sites is frequently evidenced in this species (Table 1), as corroborated by the present study. Some authors consider that such variations can be ascribed to chromosomal rearrangements and transfer of ribosomic sites (Fernandes and Martins-Santos 2006a, Peres et al. 2008) However, in several cases, transposition events have been held liable for that variability of NORs in the genoma of these animals (Mantovani et al. 2000).
It is worth noting that an AgNORs pair was the most frequently found in the chromosome preparations of the populations analyzed herein. It can be considered a main pair with NOR always active, together with secondary sites, as observed by Pazza et al. (2006) in Astyanax fasciatus (Cuvier, 1819).
After FISH with 18S rDNA probe, Astyanax altiparanae of Água dos Patos stream showed two chromosome pairs with fluorescent signals on the short arms, coinciding with the sites detected by silver impregnation. The population of Igapó lake, however, showed only one chromosome pair with fluorescent signals on the short arm, coinciding with a pair frequently identified by silver nitrate, thus characterizing a system of simple NORs. The other markings, which had not been identified by FISH, but were observed in this population after the impregnation with silver nitrate, are probably heterochromatic sites with acid proteins that have affinity to silver. Despite the fact that multiple NORs are a common condition among Astyanax altiparanae (Table 1), Domingues et al. (2007) and Ferreira Neto et al. (2009) also found a system of simple NORs confirmed by FISH in different populations of this species.
Multiple 18S rDNA sites were also found among other species of the genus Astyanax, such as Astyanax scabripinnis (Jenyns, 1842) (Souza et al. 2001, Mantovani et al. 2005, Fernandes and Martins-Santos 2006b) and Astyanax fasciatus (Pazza et al. 2006), which are, therefore, a characteristic of this group of fish.
The CMA3+ sites of Astyanax altiparanae of Igapó lake and Água dos Patos stream were consistent with those marked by silver, however, the AgNORs not detected through CMA3 in the individuals of Igapó lake may be very small and not detectable by this fluorochrome, or else not all NORs are rich in GC, as suggested by Artoni et al. (1999).
The Ag-NOR heteromorphism observed in the two populations was not seen by FISH, thence, it should be related to the expression of the genes or be ascribable to a larger amount of heterochromatin that insert in 18S DNAr cistrons, once it was detected by CMA3 and by the impregnation with silver nitrate.
After staining with DAPI, markings were not observed in the chromosomes of the two populations of Astyanax altiparanae, which, therefore, did not possess any region rich in AT bases. Rosa et al. (2009) used CMA3 and DAPI fluorochromes in chromosome preparations of Astyanax laticeps (Cope, 1894) and observed the occurrence of NORs rich in GC and poor in AT bases, respectively.
In all the populations of Astyanax altiparanae, the5S DNAr sites were located interstitially in one chromosome pair, demonstrating a high stability of those sites. This corroborates the data on other populations of Astyanax altiparanae (Table 1) and of other species of the genus Astyanax (Almeida-Toledo et al. 2002, Fernandes and Martins-Santos 2006b, Pazza et al. 2006). The conservation of this pattern can be attributed to the interstitial location of those sites in the chromosomes, whereby the 5S DNAr is protected from the dispersion events that may occur with 45S DNAr, as proposed by Martins and Galetti Jr. (2001).
According to Ferreira Neto et al. (2009), the apparent karyotypic similarity among the populations of Astyanax altiparanae strongly suggests an intimate relationship among them. However, the small karyotypic variations detected indicate some evolutionary divergence, probably due to restrictions on gene flow. The data obtained confirm the occurrence of similarity in relation to diploid number and 5S rDNA location, however, differences in the karyotypic macrostructure and in the distribution of 18S rDNA sites are found among the populations. Thence, from the results obtained in this work we corroborate the existing data and, once again, confirm the great chromosome variability of Astyanax altiparanae that can be significant for cytotaxonomy in this group.
Acknowledgments
The authors are grateful to CAPES, CNPq and Fundação Araucária for financial support and to Dr. Oscar A. Shibatta for the identification of the studied species.
References
- Abelini E. (2007) Análise citogenética em três espécies do gênero Astyanax (Pisces, Characiformes). Dissertation. Universidade Estadual de Maringá; Paraná. Brazil: 67 pp. [In Portuguese] [Google Scholar]
- Almeida–Toledo LF, Ozouf-Costaz C, Foresti F, Bonillo C, Porto-Foresti F, Daniel-Silva MFZ. (2002)Conservation of the 5S-bearing chromosome pair and co-localization with major rDNA clusters in five species of Astyanax (Pisces, Characidae). Cytogenetic and Genome Research 97(3–4): 229–233 doi: 10.1159/000066609 [DOI] [PubMed] [Google Scholar]
- Artoni RF, Bertollo LAC. (1999) Nature and distribution of constitutive heterochromatin in fishes, genus Hypostomus (Loricariidae). Genetica 106 (3): 209–214 doi: 10.1023/A:1003957719178 [DOI] [PubMed] [Google Scholar]
- Bertollo LAC, Takahashi CS, Moreira-Filho O. (1978) Brazilian Journal of Genetics 1 (2): 103-120 [Google Scholar]
- Daniel-Silva MFZ, Almeida-Toledo LF. (2001) Chromosome R-banding pattern and conservation of a marker chromosome in four species, genus Astyanax (Characidae, Tetragonopterinae). Caryologia 54 (3): 209-215 [Google Scholar]
- Domingues MS, Vicari MR, Abilhoa V, Wamser JP, Cestari MM, Bertollo LAC, Almeida MC, Artoni RF. (2007) Cytogenetic and comparative morphology of two allopatric populations of Astyanax altiparanae Garutti et Britski, 2000 (Teleostei: Characidae) from upper rio Paraná basin. Neotropical Ichthyolology 5(1): 37–44 doi: 10.1590/S1679-62252007000100005 [Google Scholar]
- Fernandes CA, Martins-Santos IC. (2004) Cytogenetic studies in two populations of Astyanax altiparanae (Pisces, Characiformes). Hereditas 141 (3): 328–332 doi: 10.1111/j.1601-5223.2004.01832.x [DOI] [PubMed]
- Fernandes CA, Martins-Santos IC. (2006a) Mapping of the 18S and 5S ribosomal RNA genes in Astyanax altiparanae Garutti et Britski, 2000 (Teleostei, Characidae) from the upper Paraná river basin, Brazil. Genetics and Molecular Biology 29 (1): 1–5 doi: lpage10.1590/S1415-47572006000300011 [Google Scholar]
- Fernandes CA, Martins-Santos IC. (2006b) Chromosomal location of 5S and 18S rRNA genes in three sympatric cytotypes of Astyanax scabripinnis (Characiformes, Characidae) from the Ivaí river basin, state of Paraná, Brazil. Caryologia (Firenze) 59 (3): 253-259 [Google Scholar]
- Ferreira Neto M, Vicari MR, Camargo EF de, Artoni RF, Moreira-Filho O. (2009) Comparative cytogenetics among populations of Astyanax altiparanae (Characiformes, Characidae, Incertae sedis). Genetics and Molecular Biology 32 (4): 792–796 doi: lpage10.1590/S1415-47572009005000078 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garutti V, Britski HA. (2000) Descrição de uma espécie nova de Astyanax (Teleostei: Characidae) da bacia do alto rio Paraná e considerações sobre as demais espécies do gênero na bacia. Comunicações do Museu de Ciência e Tecnologia. Série Zoologia 13: 65-88 [Google Scholar]
- Hatanaka T, Galetti Jr PM. (2004) Mapping of the 18S and 5S ribosomal RNA genes in the fish Prochilodus argenteus Agassiz, 1829 (Characiformes, Prochilodontidae). Genetica (The Hague) 122 (3): 239–244 doi: 10.1007/s10709-004-2039-y [DOI] [PubMed] [Google Scholar]
- Howell WM, Black DA. (1980) Controlled silver-staining of nucleolus organizer regions with the protective coloidal developer: a 1-step method. Experientia 36(8): 1014–1015 doi: 10.1007/BF01953855 [DOI] [PubMed] [Google Scholar]
- Levan A, Fredga K, Sandberg AA. (1964) Nomenclature for centromeric position of chromosomes. Hereditas 52: 201–220 doi: 10.1111/j.1601-5223.1964.tb01953.x [Google Scholar]
- Lima FCT, Malabarba LR, Buckup PA, Silva JFP, Vari RP, Harold H, Benine R, Oyakawa O, Pavanelli CS, Menezes NA, Lucena CAS, Malabarba MCSL, Lucena ZMS, Reis RE, Langeani F, Cassati L, Bertaco VA, Moreira C, Lucinda PHF. (2003) Genera Incertae sedis in Characidae. In: Reis RE, Kullander SO, Ferraris Jr. CJ. (Eds) Check list of the freshwater fishes of South and Central America. EDIPURCS, Porto Alegre, 106–109 [Google Scholar]
- Mantovani M, Abel LDS, Mestriner CA, Moreira-Filho O. (2000) Accentuated polymorphism of heterochromatin and nucleolar organizer regions in Astyanax scabripinnis (Pisces, Characidae): tools for understanding karyotypic evolution. Genetica 109(3): 161–168 doi: 10.1023/A:1017546601065 [DOI] [PubMed] [Google Scholar]
- Mantovani M, Abel LDS, Moreira-Filho O. (2005) Conserved 5S and variable 45S rDNA chromosomal localization revealed by FISH in Astyanax scabripinnis (Pisces, Characidae). Genetica 123 (3): 211–216 doi: 10.1007/s10709-004-2281-3 [DOI] [PubMed]
- Martins C, Galetti Jr. PM. (1999) Chromosomal localization of 5S rDNA genes in Leporinus fish (Anostomidae, Characiformes). Chromosome Research 7 (5): 363–367 doi: 10.1023/A:1009216030316 [DOI] [PubMed] [Google Scholar]
- Martins C, Galetti Jr PM. (2001) Two 5S rDNA arrays in Neotropical fish species: is it a general rule for fishes. Genetica 111 (1–3): 439–446 doi: 10.1023/A:1009216030316 [DOI] [PubMed] [Google Scholar]
- Morelli S, Bertollo LAC, Foresti F, Moreira-Filho O, Toledo-Filho SA. (1983) Cytogenetics considerations on the genus Astyanax (Pisces, Characidae), I Karyotypic variability. Caryologia 36 (3): 235-244 [Google Scholar]
- Pacheco RB. (2001) Estudos citogenéticos em diferentes populações de Astyanax altiparanae (Pisces, Tetragonopterinae). Dissertation. Universidade Estadual de Londrina; Paraná. Brazil: 97pp. [In Portuguese] [Google Scholar]
- Pacheco RB, Giuliano-Caetano L, Dias AL. (2001) Occurrence of cytotypes and multiple NORs in an Astyanax altiparanae population (Pisces, Tetragonopterinae). Chromosome Science 5 (2): 109-114 [Google Scholar]
- Paganelli HH. (1990) Diversidade cromossômica no gênero Astyanax, com especial referência a Astyanax bimaculatus. (Linnaeus, 1758). Considerações citotaxonômicas e evolutivas. Dissertation. Universidade Federal de São Carlos; São Paulo. Brazil: 108pp. [In Portuguese] [Google Scholar]
- Pazza R, Kavalco KF, Bertollo LAC. (2006) Chromosome polymorphism in Astyanax fasciatus (Teleostei, Characidae).1. Karyotype analysis, Ag-NORs and mapping of the 18S and 5S ribosomal genes in sympatric karyotypes and their possible hybrid forms. Cytogenetic Genome Research 112 (3–4): 313–319 doi: 10.1159/000089886 [DOI] [PubMed] [Google Scholar]
- Peres WAM, Bertollo LAC, Moreira-Filho O. (2008) Physical mapping of the 18S and 5S ribosomal genes in nine Characidae species (Teleostei, Characiformes). Genetics and Molecular Biology 31 (1) (suppl): 222–226 doi: 10.1590/S1415-47572008000200009 [Google Scholar]
- Rosa R, Rubert M, Malabarba LR, Martins-Santos IC, Giuliano-Caetano L. (2009) Cytogenetic analysis of Astyanax laticeps (Cope, 1894) (Ostariophysi: Characidae) from the laguna dos Patos system. Neotropical Ichthyology 7(4): 601–605 doi: 10.1590/S1679-62252009000400007 [Google Scholar]
- Schweizer D. (1976) Reverse fluorescent chromosome banding with chromomycin and DAPI. Chromosoma 58(4): 307–324 doi: 10.1007/BF00292840 [DOI] [PubMed]
- Souza IL, Galiann J, Rua PDL, Bertollo LAC, Moreira-Filho O. (2001) Non-radom distribution of the GC-rich heterochromatin and nuclear rDNA sites on Astyanax scabripinnis chromosomes. Cytologia 66 (1): 85-91 [Google Scholar]
- Swarça AC, Cestari MM, Giuliano-Caetano L, Dias AL. (2001) Cytogenetic Characterization of the Large South American Siluriform Fish Species Zungaro zungaro (Pisces, Pimelodidae). Chromosome Science 5 (1): 51-55 [Google Scholar]