

Abstract
Objectives. This study investigated the association between polymorphisms in the receptor for advanced glycation end products (RAGE) gene and the susceptibility to diabetic retinopathy (DR) in a Chinese population and identified a correlation between serum-soluble RAGE (sRAGE) levels and DR risk. Materials and Methods. We enrolled 1040 patients with type 2 diabetes mellitus: 372 patients with DR and 668 without retinopathy (NDR). All polymorphisms were genotyped by time-of-flight mass spectrometry. Serum levels of sRAGE were assayed by enzyme-linked immunosorbent assays. The interaction of SNPs was analyzed by multifactor dimensionality reduction (MDR). Results. The frequency of the SS genotype for the G82S polymorphism was 12.4% in the DR group and 6.6% in the NDR group; this difference was significant. G82S was associated with sRAGE levels. Specifically, after adjustments for age, sex, duration, and glucose metabolism, serum sRAGE levels were significantly higher in DR subjects with the S/S genotype than in NDR subjects in general. In the DR group, subjects with the G/S genotype had lower sRAGE levels than subjects with the G/G or S/S genotype (P < 0.01). The best multilocus genetic interaction model was assessed using the MDR method; 2184A/G, 1704G/T, G82S, and −429T/C were identified. Conclusions. The findings suggest that the G82S polymorphism in the RAGE gene is associated with DR risk, and G82S was associated with circulating levels of sRAGE. The mechanism by which G82S polymorphism modulates the sRAGE levels remains to be elucidated.
1. Introduction
The incidence of diabetes is rapidly increasing in developing countries, especially in China. According to the latest report from the Chinese Diabetes Society (CDS), the prevalence of diabetes rose to 9.7% in 2010 in China; approximately 92.4 million adults (age ≥ 20 years) are affected.
Diabetic retinopathy (DR) is a potentially devastating microvascular complication in diabetes and one of the leading causes of vision loss and blindness worldwide. DR occurs in approximately 35.7% of patients with diabetes in China [1]. The pathogenesis of DR is complex, but severe hyperglycemia is the major risk factor for developing retinopathy [2]. Prolonged hyperglycemia is required for the development of anatomic retinal vascular lesions in most animal models and in humans with DR [3]. Chronic exposure of the retina to hyperglycemia leads to the formation and accumulation of advanced glycation end products (AGEs), which play an important role in the progression of retinopathy.
AGEs are proteins or lipids that become nonenzymatically glycated after exposure to sugars. Prolonged hyperglycemia, dyslipidemia, and oxidative stress in diabetes increase the production and accumulation of AGEs in the diabetic vasculature [4]. The effects of AGEs are partially mediated by cellular receptors, the most important being the receptor for AGE (RAGE). RAGE is a member of the immunoglobulin superfamily of cell surface molecules and is expressed ubiquitously in various retinal cells. In animal models and humans with diabetes, RAGE expression in the retina increases, concomitant with AGE accumulation [5, 6]. Upregulation of RAGE in the retinas of patients with diabetes activates prooxidant and proinflammatory signaling pathways [7]. The AGE-RAGE interaction activates multiple intracellular signaling pathways and subsequently evokes oxidative stress and inflammatory responses in vascular cells, resulting in the transcription of target genes and oxidative stress, thus contributing to the development and progression of DR [8].
RAGE has a C-terminal-truncated secretory isoform, termed soluble RAGE (sRAGE), which neutralizes AGE-mediated damage by acting as a decoy [9]. In humans, sRAGE is produced by alternative splicing of RAGE messenger RNA or by proteolytic cleavage and shedding of cell-bound RAGE [7]. High levels of sRAGE are present in the circulatory system. Given its AGE-binding properties, sRAGE might play an antagonistic role by competing with cell surface RAGE, thus preventing the RAGE-mediated inflammatory response. As such, the proportion and production of sRAGE may influence RAGE-mediated functions in various tissues and inflammatory conditions [10, 11].
The RAGE gene, composed of a 1.7-kb 5′ flanking region and 11 exons [12], is located on chromosome 6p21.3 in the MHC locus. It is an attractive candidate gene for influencing DR. Sequence variation within the RAGE gene has been studied, and a relatively large number of single nucleotide polymorphisms (SNPs) in the RAGE coding and noncoding regions have been identified [13, 14], including the exon polymorphism G82S, the promoter region polymorphism −429T/C, and several intron polymorphisms (1704G/T, 2184A/G, and rs1035798). Many of the findings, however, are inconsistent.
In this study, we investigated the higher-order interactions among the abovementioned polymorphisms as they relate to DR risk. We performed a cross-sectional study to determine (1) the relationship between genetic polymorphisms in the RAGE gene (2184A/G (rs3134940), 1704G/T (rs184003), rs1035798, G82S (rs2070600), and −429T/C (rs1800625)) and the risk of DR; (2) potential multilocus interactions that affect the risk of DR; (3) the association between RAGE polymorphisms and sRAGE levels; and (4) the association between sRAGE levels and DR.
2. Materials and Methods
2.1. Study Population
This study enrolled 1040 patients diagnosed with type 2 diabetes mellitus based on clinical features, laboratory data, and the guidelines in the recent Expert Committee Report of the American Diabetes Association [15]. Additionally, all participants underwent a fundus fluorescein angiography by certified ophthalmologists. Three-hundred and seventy-two patients were diagnosed with DR as a complication of diabetes (DR group), and 668 diabetic patients without DR were included in the study for comparison (NDR group). The following were excluded: (1) patients with diabetes undergoing thiazolidinedione therapy; (2) patients with hypertension undergoing angiotensin-converting enzyme inhibitors therapy; (3) patients with diagnosed diabetic nephropathy; (4) patients with acute or chronic inflammatory disease; and (5) patients with type 1 diabetes, maturity-onset diabetes of the young, or mitochondrial diabetes. All participants provided written informed consent. The protocol for this study complies with the Helsinki declaration. It was approved by the Committees on the Ethics of Human Research of Harbin Medical University.
Clinical data were recorded with respect to age, sex, body mass index (BMI), diabetes duration, systolic blood pressure, diastolic blood pressure, waist-hip ratio (WHR), total cholesterol (TC) level, triglyceride (TG) level, high density lipoprotein-cholesterol (HDL-C) level, low density lipoprotein-cholesterol (LDL-C) level, fasting plasma glucose (FPG) level, 2 h postprandial plasma glucose (2hPG) level, and hemoglobin A1c (HbA1c) level.
2.2. Gene Variants and Genotyping
The selection of particular SNPs was based on the following: (1) a population frequency, based on the minor allele frequency (MAF), of >5%; (2) known functional or regulatory impact; and/or (3) previously described association with DR.
Genomic DNA from peripheral blood leukocytes was extracted from 200 μL of whole blood obtained at the time of the interview, using the TIANamp Blood DNA Kit (TIANGEN, China) according to the manufacturer's instructions. The DR and NDR groups were mixed for genotyping, and laboratory personnel were blinded to the patient's diagnosis (DR versus NDR).
Genomic DNA was diluted to a final concentration of 15–20 ng/μL for the genotyping assays. Genotyping for each participant was performed using a MassARRAY compact analyzer based on the chip-based matrix-assisted laser desorption ionization time-of-flight mass spectrometry platform (Sequenom, San Diego, CA, USA). The process was as follows: (1) DNA isolation, (2) primer design, (3) PCR, (4) neutralization of unincorporated dNTPs (SAP Reaction), (5) extension reaction, (6) conditioning of iPLEX Gold reaction products, (7) application to the SpectroCHIP II array, (8) definition of assays and plates, and (9) spectrum acquisition and analysis. Primers for the polymerase chain reaction and single base extension were designed using SEQUENOM Human Genotyping Tools (online) and SEQUENOM MassARRAY Assay Design v4.0 software.
2.3. Biochemical Analysis
Blood samples were obtained after an overnight fast from each participant for baseline measurements, and the samples were stored at −80°C until use. Plasma glucose levels, 2hPG levels, total cholesterol levels, HDL levels, and triglyceride levels were also measured using enzymatic methods. LDL levels were calculated using the Friedewald equation. HbA1c levels were assayed using high-performance liquid chromatography with a cation exchange column.
2.4. ELISA for sRAGE
sRAGE was measured by sandwich colorimetric ELISA using commercially available kits (human sRAGE ELISA kit; R&D Systems, Abingdon, UK) with a standard range of 31.25–2000 pg/mL and intra-assay and inter-assay coefficients of variation (CV) of <10% and <15%, respectively. The results were expressed in pg/mL.
2.5. Statistical Methods
Statistical analysis was performed using the SPSS software package (SPSS 13.0, Beijing, China). Continuous variables were expressed as the mean ± SD. Categorical variables were presented as frequencies. Group differences were analyzed by Student's t-test, the Mann-Whitney U test, and the Chi-square test for normally distributed, abnormally distributed, and noncontiguous variables, respectively. We performed 3 independent tests for sRAGE levels in individuals with G82S genotypes. To avoid type I error induced by multiple tests, Bonferroni's adjustment for multiple comparisons was applied, and a P value of <0.017 was used as the significance threshold.
For genotypic and allelic frequencies, the Hardy-Weinberg equilibrium was applied using SNPStats online tools (http://bioinfo.iconcologia.net/snpstats/start.htm). The linkage disequilibrium (LD) between the RAGE SNPs was assessed with the Haploview software package, and Lewontin's disequilibrium coefficient (D′) was estimated.
Univariate analysis and multiple logistic regression analysis were performed for independent determinants of serum sRAGE levels and DR. Multifactor dimensionality reduction version 1.2.5 was used to detect potential multilocus genetic interactions (http://linux.softpedia.com/progDownload/MDR-Download-35200.html). Statistical power was calculated using the software PASS (http://www.ncss.com/). A P value of <0.05 was considered statistically significant for all analyses.
3. Results
3.1. Power Analysis
Power analysis was performed to determine the number of subjects needed for this study. Using the equation of contingency table, we achieved values of 99% for the Chi-square test and determined an appropriate sample size of 311 subjects. For this study, 1040 subjects were included, more than required to detect statistical significance.
3.2. Clinical Characteristics of the Groups
The basic characteristics of the 2 groups are shown in Table 1. Each group was significantly different with respect to duration and the levels of FPG, 2hPG, HbA1c, and LDL-C.
Table 1.
Clinical characteristics of DR and NDR groups.
| DR (n = 372) | NDR (n = 668) | Statistic value | |
|---|---|---|---|
| Gender (male/female) | 146/226 | 242/426 | 0.334 |
| Age (mean ± SD) | 63.39 ± 10.60 | 62.58 ± 11.65 | 0.257 |
| Diabetes duration | 9.27 ± 6.78 | 5.56 ± 5.58 | <0.001* |
| BMI (kg/m2) | 24.43 ± 3.47 | 24.54 ± 3.86 | 0.635 |
| WHR | 0.92 ± 0.14 | 0.91 ± 0.07 | 0.158 |
| Systolic blood pressure (mmHg) | 133.85 ± 16.48 | 132.08 ± 15.32 | 0.091 |
| Diastolic blood pressure (mmHg) | 80.62 ± 10.46 | 79.57 ± 9.13 | 0.116 |
| FPG (mmol/L) | 8.47 ± 3.43 | 7.54 ± 3.19 | <0.001* |
| 2hPG (mmol/L) | 13.01 ± 4.36 | 12.01 ± 4.30 | <0.001* |
| HbA1c (%) | 9.43 ± 2.33 | 8.65 ± 2.36 | 0.049* |
| Total cholesterol (mmol/L) | 5.49 ± 1.02 | 5.43 ± 0.96 | 0.341 |
| Triglycerides (mmol/L) | 2.21 ± 1.27 | 2.13 ± 1.03 | 0.254 |
| HDL-C (mmol/L) | 1.30 ± 0.31 | 1.32 ± 0.32 | 0.297 |
| LDL-C (mmol/L) | 2.81 ± 0.46 | 2.91 ± 0.45 | 0.001* |
| sRAGE (pg/mL) | 526.17 (381.96–798.33) | 522.15 (360.79–739.80) | 0.168 |
*Significant results.
Numbers are given as n (%).
3.3. Polymorphism Distribution in the DR and NDR Groups
The genotype and the allele distributions of the 5 RAGE polymorphisms analyzed in this study are shown in Table 2. The genotype frequencies of the 2 groups are in accordance with those predicted by the Hardy-Weinberg equilibrium for all polymorphisms (except for rs1035798 in DR group). Linkage disequilibrium among the studied polymorphisms was not detected. A significant association was observed between the SS genotype and the risk of DR; the other polymorphisms showed no statistically significant association with DR.
Table 2.
Genotype and allele frequencies of RAGE gene polymorphisms in diabetic patients with and without retinopathy.
| Polymorphism | Genotype | Genotype distribution | Alleles | Allele frequency | |||||
|---|---|---|---|---|---|---|---|---|---|
| Cases | Control | OR (95% CI) | P | Cases | Control | P | |||
| 2184A/G rs3134940 (n = 982) | A/A | 235 | 448 | 1.00 | 0.35 | A | 0.82 | 0.84 | 0.28 |
| A/G | 108 | 167 | 0.81 (0.61–1.08) | G | 0.18 | 0.16 | |||
| G/G | 8 | 16 | 1.05 (0.44–2.49) | ||||||
|
| |||||||||
| 1704G/T rs184003 (n = 1039) | G/G | 266 | 490 | 1.00 | 0.40 | 0.85 | 0.85 | 0.77 | |
| G/T | 98 | 156 | 0.86 (0.64–1.16) | 0.15 | 0.15 | ||||
| T/T | 8 | 21 | 1.42 (0.62–3.26) | ||||||
|
| |||||||||
| rs1035798 (n = 1038) | C/C | 274 | 467 | 1.00 | 0.08 | C | 0.85 | 0.84 | 0.57 |
| C/T | 83 | 183 | 1.29 (0.96–1.74) | T | 0.15 | 0.16 | |||
| T/T | 15 | 16 | 0.63 (0.30–1.29) | ||||||
|
| |||||||||
| G82S rs2070600 (n = 1039) | G/G | 195 | 396 | 1.00 | 0.0035* | G | 0.70 | 0.76 | 0.0015* |
| G/S | 131 | 227 | 1.17 (0.89–1.54) | S | 0.30 | 0.24 | |||
| S/S | 46 | 44 | 2.12 (1.36–3.32) | ||||||
|
| |||||||||
| −429T/C rs1800625 (n = 1037) | T/T | 280 | 502 | 1.00 | 0.42 | T | 0.86 | 0.87 | 0.63 |
| C/T | 80 | 150 | 1.05 (0.77–1.42) | C | 0.14 | 0.13 | |||
| C/C | 12 | 13 | 0.60 (0.27–1.34) | ||||||
The Chi-square test was used to compare the genotype and allele frequencies between DR and NDR groups.
*P < 0.05 was considered statistically significant.
3.4. Multilocus Genetic Interactions and the Risk of DR
Multilocus genetic interactions were analyzed by MDR. Each genotype was categorized into 2 subgroups, a high-risk group and a low-risk group, according to the ratio of cases to controls with the genotype. The prediction accuracy of the model was estimated by tenfold cross-validation. The best model chosen had the highest cross-validation consistency (CVC) and the highest prediction accuracy. CVC, training balance accuracy, testing balance accuracy, associated P values, OR, and 95% CI were obtained from the MDR analysis. The best model of multilocus analysis is shown in Table 3. One 4-locus model had a maximum CVC of 10 and a maximum prediction accuracy of 54.92%. The best model consisted of 4 SNPs (2184A/G, 1704G/T, G82S, and −429T/C). Figure 1 shows an interaction dendrogram highlighting the information obtained using MDR. A dashed line indicates moderate interaction, and a dotted line represents independence. The interaction dendrogram indicated that this set, which included the 2184A/G and 1704G/T polymorphisms, exhibited a moderate synergistic effect, whereas G82S and −429T/C had no synergistic effect.
Table 3.
Multilocus genetic interaction model.
| Model | Training balance accuracy | Testing balance accuracy | CVC | OR 95% CI | P value |
|---|---|---|---|---|---|
| G82S | 0.5353 | 0.5238 | 9/10 | 1.3297 (1.03, 1.7163) | 0.0286 |
| 2184A/G, G82S | 0.5565 | 0.5232 | 6/10 | 1.5972 (1.2277, 2.078) | 0.0005 |
| 2184A/G, G82S, −429T/C | 0.5732 | 0.5187 | 4/10 | 2.1518 (1.5975, 2.8983) | <0.0001 |
| 2184A/G, 1704G/T, G82S, −429T/C | 0.5957 | 0.5492 | 10/10 | 2.2254 (1.7115, 2.8935) | <0.0001 |
The best model has the maximum CVC and highest testing balance accuracy.
Figure 1.
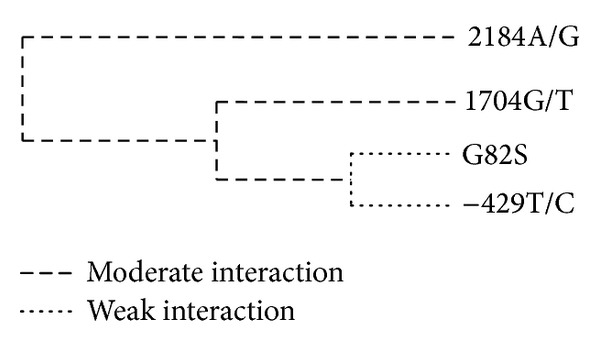
Interaction dendrogram for the best 4-locus SNP model selected by the MDR. The dashed line indicates moderate interaction, and the dotted line indicates independence.
3.5. Relation between RAGE G82S Genotypes and sRAGE Levels
Because sRAGE levels were nonnormally distributed, we conducted the analysis using a nonparametric test. There was no statistically significant difference between the DR and NDR groups in terms of sRAGE concentration in plasma after adjusting for the duration of diabetes, FPG, 2hPG, LDL, and HbA1c (P > 0.05). However, there was a significant association between the G82S genotypes and plasma sRAGE concentrations in all subjects with diabetes (P < 0.01, Figure 2(a)). Serum sRAGE levels were significantly higher in DR subjects with the S/S genotype than in NDR subjects with the S/S genotype. In addition, in the DR group, subjects with the G/S genotype had lower sRAGE levels than those with the G/G or S/S genotype. Besides, sRAGE concentrations in NDR subjects with the G/S genotype were significantly higher than in those with the S/S genotype and significantly lower than in those with the G/G genotype (Figure 2(b)).
Figure 2.
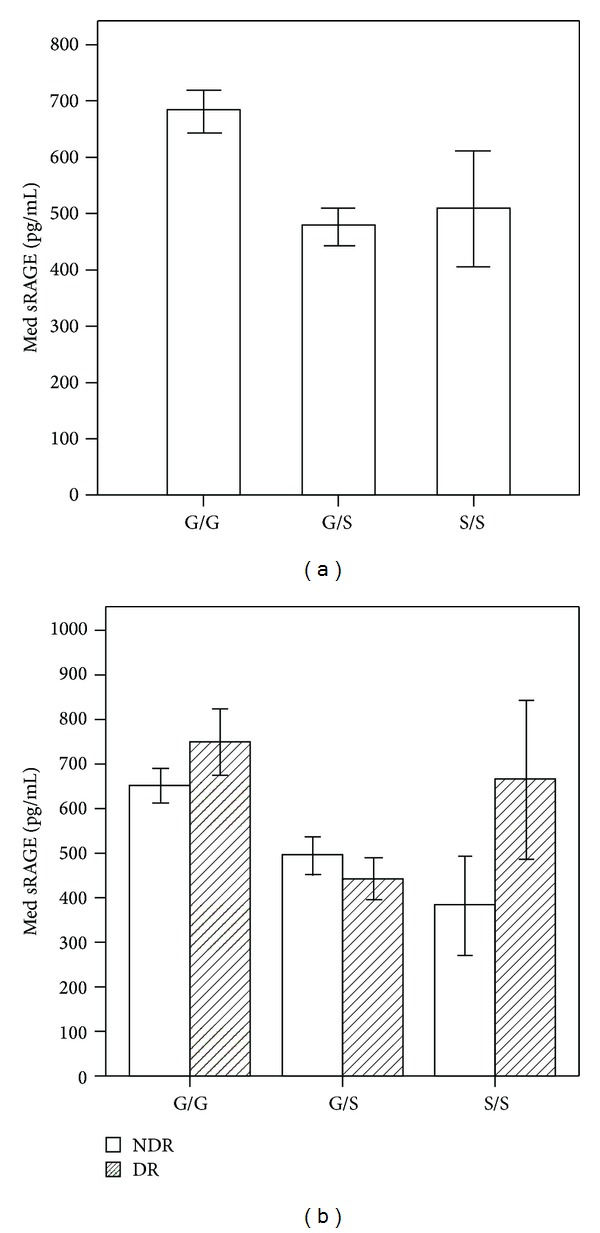
Influence of RAGE G82S genotypes on sRAGE. All diabetic subjects with the G/G genotype had significantly higher sRAGE concentrations than those with G/S genotype and S/S genotype (bonferroni adjustment P < 0.01, (a)). sRAGE concentrations in DR subjects with the S/S genotype were higher than in NDR subjects with the S/S genotype (P < 0.05). DR subjects with the G/S genotype had lower sRAGE levels than DR subjects with the G/G and S/S genotypes after adjusting for the duration of diabetes, FPG, 2hPG, LDL, and HbA1c (Bonferroni adjustment P < 0.01). Note: error bars: 95% CI.
4. Discussion
In this study, the G82S polymorphism in RAGE was associated with DR risk. However, we did not find a significant association between other RAGE SNPs and DR susceptibility. To our knowledge, this is the first study to investigate the association between several RAGE polymorphisms and DR in Chinese populations. Furthermore, we assayed serum sRAGE levels in the DR and NDR groups to better understand the nature of this correlation.
The G82S polymorphism, one of the first polymorphisms identified in the RAGE gene, is potentially interesting because it occurs at a predicted N-linked glycosylation in the AGE binding site and thereby influences AGE-RAGE interactions [16]. Several studies have investigated the association between G82S and DR in various populations. However, the results were inconsistent. In our study, the G82S polymorphism was associated with DR, and the SS genotype was a risk factor for DR. This suggests that the G82S SNP in the RAGE gene might contribute to DR pathogenesis. Our results are similar to those of another study, which found that the 82S allelic variant increased the risk of DR [17]. Other studies did not find a significant association between G82S and DR [18–20]; however, one did not report the sample size of DR subjects, and the sample size in the other studies was smaller than in this study. Different population histories, which could alter haplotype block structure resulting in noncausative allelic association, or differing genetic and environmental contributions of disease risk in various race and geographic groups might also explain the different results [21]. Additionally, the type of diabetes, duration of diabetes, and other characteristics, such as the patients selected and the sample size, might vary between the studies and influence the results.
In this study, there was no significant difference in the serum levels of sRAGE between the DR and NDR groups. This finding differs from previous studies, in which an association between circulating concentrations of sRAGE and DR was demonstrated. Specifically, lower levels of sRAGE were found in patients who developed DR [22, 23]. Another study demonstrated that sRAGE serum levels were significantly higher in patients who had diabetes and proliferative DR than in patients who had diabetes but no retinopathy [24]. Differences in the subject population (age and duration of diabetes) and ethnicity could account for these discrepancies. However, the sample size in the previous studies was less than 50. Therefore, future prospective cohort studies with large sample sizes are necessary to clarify the relationships between serum sRAGE levels and DR.
Although the association between G82S and circulating sRAGE levels has been previously demonstrated in nondiabetic Korean populations as well as diabetic Caucasian populations [25, 26], this is the first study to confirm these findings in DR populations. In this study, the G82S polymorphism correlated with serum sRAGE in all the patients who had diabetes. In addition, S/S genotype was a risk factor for DR, and DR subjects with the S/S genotype had significantly higher levels of sRAGE than the NDR subjects in general, which suggests the significance increase in the level of sRAGE in DR subjects with the S/S genotype might be associated with an increased risk for developing of DR. Although the mechanism that S/S genotype influences the serum levels of sRAGE is still not understood, our hypothesis is supported by the following observations. (1) RAGE 82S allele upregulates the inflammatory response and may contribute to enhanced proinflammatory mechanisms in inflammatory diseases [27]. (2) RAGE expression was enhanced in diabetic vasculature and sRAGE could be generated from the cleavage of cell surface RAGE extracellular domain in endothelial cells by proteolytic means [28]. sRAGE can be formed by cleavage from the cell-surface receptor isoform [29, 30].
In addition, DR subjects with the G/S genotype had significantly lower serum sRAGE levels than those with the G/G or S/S genotype. In our study, we used the R&D immunoassay to assay total circulating sRAGE. The low sRAGE levels assayed in the G/S genotypes (with serine at location 82) could reflect a higher binding affinity of RAGE for ligand [31] and, consequently, a diminished ability of the R&D antibody to capture sRAGE. The mechanism by which G82S affects the serum levels of sRAGE is unknown, but the polymorphism is thought to have functional significance because it is located in the ligand-binding V domain of RAGE. As such, it may alter the N-linked glycosylation state of the protein, resulting in conformational changes in the protein that influence the detection of sRAGE by the R&D antibody. Furthermore, the polymorphism may influence intracellular processes, resulting in increased activation of proinflammatory proteins such as nuclear factor-κB (NF-κB). In turn, this intracellular signaling process may enhance RAGE expression in the retina through a feedback mechanism that would ultimately affect sRAGE concentrations [27].
It is becoming increasingly evident that single-locus effects cannot explain multifactorial chronic diseases. Thus, when the single polymorphism effect is not present alone or is not strong enough, it is important to characterize genes related to susceptibility, keeping in mind the concept of multilocus genetic interactions. Multifactor dimensionality reduction (MDR) is used to analyze gene-environment interactions, and it has been used in many genetics studies of common diseases. To the best of our knowledge, this is the first study to explore the multilocus interactions associated with DR in the Chinese population. Using MDR analysis, we determined that the 4-locus model is the best model to identify DR susceptibility. This conclusion is based on the model's balanced accuracy and CVC. This model included 2184A/G, 1704G/T, G82S, and −429T/C. The interaction dendrogram revealed that the 2184A/G and 1704G/T polymorphisms exhibit a moderate synergistic effect. We did not find an association between 2184A/G and 1704G/T intron polymorphisms and DR. Moreover, Kanková et al. previously established that these 2 polymorphisms might influence oxidative stress [32]. They also found that the 1704G/T and 2184A/G polymorphisms exhibit a similar synergistic tendency in patients with noninsulin-dependent diabetes mellitus and in patients without diabetes. These 2 polymorphisms, located at introns 7 and 8, respectively, may have similar functions and act synergistically.
The advantages of this study are as follows. First, the cross-sectional design provided a sufficient sample size. The power values of each analysis were more than 90%. Second, this is the first comprehensive study to investigate the correlation between several RAGE gene polymorphisms and DR. Third, we examined serum levels of sRAGE in Chinese population. Despite these advantages, the present study has several limitations. The system utilized to detect sRAGE cannot distinguish between sRAGE splice variants. Because the proportion of sRAGE isoforms could differ among individuals, the development of new specific assays will improve the evaluation of sRAGE levels [33]. Additionally, because the number of proliferative retinopathy (PDR) patients was small, we did not further classify DR subjects into nonproliferative retinopathy (NPDR) and proliferative retinopathy (PDR) groups.
Little is known about the mechanisms of sRAGE production and formation. The RAGE gene may participate in the regulation of sRAGE expression in vivo. Further studies are needed to explore the mechanism by which RAGE polymorphisms regulate sRAGE expression. Future prospective cohort studies with large sample sizes are necessary to clarify the cause-effect relationships between serum sRAGE levels and DR.
Conflict of Interests
The authors declare that they have no conflict of interests.
Authors' Contribution
Li Yang, Yuan Li, and Xiaohong Fan contributed equally to this work.
Acknowledgments
The authors would like to thank the participants of the study, as well as the doctors and nurses from Wenhua community clinics in Qiqihar city, China, the first and second hospital of Qiqihar city, and the Harbin hospital for their considerable assistance. This work was supported by the China Medical Board Distinguished Professorships Project (G16916400/F510000).
References
- 1.Chinese Diabetes Society. China guideline for type 2 diabetes. 2011.
- 2.Davis MD, Fisher MR, Gangnon RE, et al. Risk factors for high-risk proliferative diabetic retinopathy and severe visual loss: early treatment diabetic retinopathy study report 18. Investigative Ophthalmology and Visual Science. 1998;39(2):233–252. [PubMed] [Google Scholar]
- 3.Engerman RL, Kern TS. Retinopathy in animal models of diabetes. Diabetes/Metabolism Reviews. 1995;11(2):109–120. doi: 10.1002/dmr.5610110203. [DOI] [PubMed] [Google Scholar]
- 4.Yamagishi S-I, Ueda S, Matsui T, Nakamura K, Okuda S. Role of advanced glycation end products (AGEs) and oxidative stress in diabetic retinopathy. Current Pharmaceutical Design. 2008;14(10):962–968. doi: 10.2174/138161208784139729. [DOI] [PubMed] [Google Scholar]
- 5.Ritthaler U, Deng Y, Zhang Y, et al. Expression of receptors for advanced glycation end products in peripheral occlusive vascular disease. American Journal of Pathology. 1995;146(3):688–694. [PMC free article] [PubMed] [Google Scholar]
- 6.Soulis T, Thallas V, Youssef S, et al. Advanced glycation end products and their receptors co-localise in rat organs susceptible to diabetic microvascular injury. Diabetologia. 1997;40(6):619–628. doi: 10.1007/s001250050725. [DOI] [PubMed] [Google Scholar]
- 7.Yonekura H, Yamamoto Y, Sakurai S, et al. Novel splice variants of the receptor for advanced glycation end-products expressed in human vascular endothelial cells and pericytes, and their putative roles in diabetes-induced vascular injury. Biochemical Journal. 2003;370, part 3:1097–1109. doi: 10.1042/BJ20021371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Yamagishi S-I, Matsui T, Nakamura K, et al. Olmesartan blocks advanced glycation end products (AGEs)-induced angiogenesis in vitro by suppressing receptor for AGEs (RAGE) expression. Microvascular Research. 2008;75(1):130–134. doi: 10.1016/j.mvr.2007.05.001. [DOI] [PubMed] [Google Scholar]
- 9.Malherbe P, Richards JG, Gaillard H, et al. cDNA cloning of a novel secreted isoform of the human receptor for advanced glycation end products and characterization of cells co-expressing cell-surface scavenger receptors and Swedish mutant amyloid precursor protein. Molecular Brain Research. 1999;71(2):159–170. doi: 10.1016/s0169-328x(99)00174-6. [DOI] [PubMed] [Google Scholar]
- 10.Pullerits R, Bokarewa M, Dahlberg L, Tarkowski A. Decreased levels of soluble receptor for advanced glycation end products in patients with rheumatoid arthritis indicating deficient inflammatory control. Arthritis Research & Therapy. 2005;7(4):R817–824. doi: 10.1186/ar1749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Basta G, Corciu AI, Vianello A, et al. Circulating soluble receptor for advanced glycation end-product levels are decreased in patients with calcific aortic valve stenosis. Atherosclerosis. 2010;210(2):614–618. doi: 10.1016/j.atherosclerosis.2009.12.029. [DOI] [PubMed] [Google Scholar]
- 12.Goova MT, Li J, Kislinger T, et al. Blockade of receptor for advanced glycation end-products restores effective wound healing in diabetic mice. American Journal of Pathology. 2001;159(2):513–525. doi: 10.1016/S0002-9440(10)61723-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hudson BI, Stickland MH, Futers TS, Grant PJ. Effects of novel polymorphisms in the RAGE gene on transcriptional regulation and their association with diabetic retinopathy. Diabetes. 2001;50(6):1505–1511. doi: 10.2337/diabetes.50.6.1505. [DOI] [PubMed] [Google Scholar]
- 14.Kaňková K, Záhejský J, Márová I, et al. Polymorphisms in the RAGE gene influence susceptibility to diabetes-associated microvascular dermatoses in NIDDM. Journal of Diabetes and its Complications. 2001;15(4):185–192. doi: 10.1016/s1056-8727(00)00135-5. [DOI] [PubMed] [Google Scholar]
- 15.American Diabetes Association. Standards of medical care in diabetes—2009. Diabetes Care. 2009;32(supplement 1):S13–S61. doi: 10.2337/dc09-S013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hudson BI, Stickland MH, Grant PJ. Identification of polymorphisms in the receptor for advanced glycation end products (RAGE) gene: prevalence in type 2 diabetes and ethnic groups. Diabetes. 1998;47(7):1155–1157. doi: 10.2337/diabetes.47.7.1155. [DOI] [PubMed] [Google Scholar]
- 17.Zhang HM, Chen LL, Wang L, et al. Association of 1704G/T and G82S polymorphisms in the receptor for advanced glycation end products gene with diabetic retinopathy in Chinese population. Journal of Endocrinological Investigation. 2009;32(3):258–262. doi: 10.1007/BF03346463. [DOI] [PubMed] [Google Scholar]
- 18.Liu L, Xiang K. RAGE Gly82Ser polymorphism in diabetic microangiopathy. Diabetes Care. 1999;22(4):p. 646. doi: 10.2337/diacare.22.4.646. [DOI] [PubMed] [Google Scholar]
- 19.Yoshioka K, Yoshida T, Takakura Y, et al. Relation between polymorphisms G1704T and G82S of RAGE gene and diabetic retinopathy in Japanese type 2 diabetic patients. Internal Medicine. 2005;44(5):417–421. doi: 10.2169/internalmedicine.44.417. [DOI] [PubMed] [Google Scholar]
- 20.Ng ZX, Kuppusamy UR, Poh R, et al. Lack of association between Gly82Ser, 1704G/T and 2184A/G of RAGE gene polymorphisms and retinopathy susceptibility in Malaysian diabetic patients. Genetics and Molecular Research. 2012;11(1):455–461. doi: 10.4238/2012.March.1.2. [DOI] [PubMed] [Google Scholar]
- 21.Balasubbu S, Sundaresan P, Rajendran A, et al. Association analysis of nine candidate gene polymorphisms in Indian patients with type 2 diabetic retinopathy. BMC Medical Genetics. 2010;11(1, article 158) doi: 10.1186/1471-2350-11-158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Grossin N, Wautier M-P, Meas T, Guillausseau P-J, Massin P, Wautier J-L. Severity of diabetic microvascular complications is associated with a low soluble RAGE level. Diabetes and Metabolism. 2008;34(4, part 1):392–395. doi: 10.1016/j.diabet.2008.04.003. [DOI] [PubMed] [Google Scholar]
- 23.Al-Mesallamy HO, Hammad LN, El-Mamoun TA, Khalil BM. Role of advanced glycation end product receptors in the pathogenesis of diabetic retinopathy. Journal of Diabetes and its Complications. 2011;25(3):168–174. doi: 10.1016/j.jdiacomp.2010.06.005. [DOI] [PubMed] [Google Scholar]
- 24.Mona LAM, Abdel Hamid A, El-BeltagI T. Circulating soluble Receptor for advanced glycation end products and total antioxidant status in serum of diabetic patients with and without retinopathy. Australian Journal of Basic and Applied Sciences. 2008;2(3):718–723. [Google Scholar]
- 25.Jang Y, Kim JY, Kang S-M, et al. Association of the Gly82Ser polymorphism in the receptor for advanced glycation end products (RAGE) gene with circulating levels of soluble RAGE and inflammatory markers in nondiabetic and nonobese Koreans. Metabolism. 2007;56(2):199–205. doi: 10.1016/j.metabol.2006.09.013. [DOI] [PubMed] [Google Scholar]
- 26.Gaens KHJ, Ferreira I, van der Kallen CJH, et al. Association of polymorphism in the receptor for advanced glycation end products (RAGE) gene with circulating RAGE levels. Journal of Clinical Endocrinology and Metabolism. 2009;94(12):5174–5180. doi: 10.1210/jc.2009-1067. [DOI] [PubMed] [Google Scholar]
- 27.Hofmann MA, Drury S, Hudson BI, et al. RAGE and arthritis: the G82S polymorphism amplifies the inflammatory response. Genes and Immunity. 2002;3(3):123–135. doi: 10.1038/sj.gene.6363861. [DOI] [PubMed] [Google Scholar]
- 28.Schlueter C, Hauke S, Flohr AM, Rogalla P, Bullerdiek J. Tissue-specific expression patterns of the RAGE receptor and its soluble forms—a result of regulated alternative splicing? Biochimica et Biophysica Acta. 2003;1630(1):1–6. doi: 10.1016/j.bbaexp.2003.08.008. [DOI] [PubMed] [Google Scholar]
- 29.Galichet A, Weibel M, Heizmann CW. Calcium-regulated intramembrane proteolysis of the RAGE receptor. Biochemical and Biophysical Research Communications. 2008;370(1):1–5. doi: 10.1016/j.bbrc.2008.02.163. [DOI] [PubMed] [Google Scholar]
- 30.Raucci A, Cugusi S, Antonelli A, et al. A soluble form of the receptor for advanced glycation endproducts (RAGE) is produced by proteolytic cleavage of the membrane-bound form by the sheddase a disintegrin and metalloprotease 10 (ADAM10) FASEB Journal. 2008;22(10):3716–3727. doi: 10.1096/fj.08-109033. [DOI] [PubMed] [Google Scholar]
- 31.Osawa M, Yamamoto Y, Munesue S, et al. De-N-glycosylation or G82S mutation of RAGE sensitizes its interaction with advanced glycation endproducts. Biochimica et Biophysica Acta. 2007;1770(10):1468–1474. doi: 10.1016/j.bbagen.2007.07.003. [DOI] [PubMed] [Google Scholar]
- 32.Kanková K, Márová I, Záhejský J, et al. Polymorphisms 1704G/T and 2184A/G in the RAGE gene are associated with antioxidant status. Metabolism. 2001;50(10):1152–1160. doi: 10.1053/meta.2001.26757. [DOI] [PubMed] [Google Scholar]
- 33.Basta G, Sironi AM, Lazzerini G, et al. Circulating soluble receptor for advanced glycation end products is inversely associated with glycemic control and S100A12 protein. Journal of Clinical Endocrinology and Metabolism. 2006;91(11):4628–4634. doi: 10.1210/jc.2005-2559. [DOI] [PubMed] [Google Scholar]