

Abstract
Giant clams (genus Tridacna) are iconic coral reef animals of the Indian and Pacific Oceans, easily recognizable by their massive shells and vibrantly colored mantle tissue. Most Tridacna species are listed by CITES and the IUCN Redlist, as their populations have been extensively harvested and depleted in many regions. Here, we survey Tridacna crocea and Tridacna maxima from the eastern Indian and western Pacific Oceans for mitochondrial (COI and 16S) and nuclear (ITS) sequence variation and consolidate these data with previous published results using phylogenetic analyses. We find deep intraspecific differentiation within both T. crocea and T. maxima. In T. crocea we describe a previously undocumented phylogeographic division to the east of Cenderawasih Bay (northwest New Guinea), whereas for T. maxima the previously described, distinctive lineage of Cenderawasih Bay can be seen to also typify western Pacific populations. Furthermore, we find an undescribed, monophyletic group that is evolutionarily distinct from named Tridacna species at both mitochondrial and nuclear loci. This cryptic taxon is geographically widespread with a range extent that minimally includes much of the central Indo-Pacific region. Our results reinforce the emerging paradigm that cryptic species are common among marine invertebrates, even for conspicuous and culturally significant taxa. Additionally, our results add to identified locations of genetic differentiation across the central Indo-Pacific and highlight how phylogeographic patterns may differ even between closely related and co-distributed species.
Introduction
Giant clams of the genus Tridacna are among the most conspicuous marine invertebrates on coral reefs due to their large size and brilliantly colored mantle that contains photosynthesizing symbionts. Giant clams have traditionally provided raw material for tools, containers, and ornaments [1], and many populations are harvested for meat, shells, and the ornamental aquarium trade [2], [3]. Despite local management efforts, including mariculture [3], wild stocks of giant clams are depleted and some species are locally extinct in many areas of Southeast Asia and the South Pacific [3]–[5]. Consequently, most Tridacna species are listed by CITES (Appendix II)[6] and the IUCN Redlist [7].
There are currently eight [8] described species within the genus Tridacna (T. crocea Lamarck, 1819, T. derasa (Röding 1798), T. gigas (Linnaeus 1758), T. maxima (Röding 1798), T. mbalavuana Ladd, 1934, T. rosewateri Sirenko and Scarlato 1991, T. squamosa Lamarck 1819, and T. squamosina Sturany 1899), differentiated by morphology and habitat preference [9]–[12]. Tridacna squamosina, T. rosewateri, and T. mbalavuana have restricted distributions (Red Sea, Mauritius, and Fiji to Tonga, respectively), whereas T. derasa, T. gigas, T. crocea, T. squamosa and T. maxima are widely distributed in the Indian and Pacific Oceans, with the latter two extending their distribution into the Red Sea [8], [9]. Molecular phylogenetic investigations support monophyly of the described species [13]–[15], albeit with some disagreement among species relationships. An unpublished Master's thesis [16] also reports a morphologically distinct clam from Taiwan and uses mtDNA loci to show that this clam is highly divergent from sympatric T. maxima, potentially indicative of an additional unnamed species.
The juncture between the Indian and Pacific Oceans (Fig. 1), where several species of Tridacna are sympatric [8], is a well-known epicenter of tropical marine biodiversity [17], [18]. Genetic surveys in this region have revealed cryptic species, even among conspicuous and well-studied marine invertebrates [19], [20]. Many species show substantial intraspecific genetic division between the ocean basins (reviewed by [21]), with the Sunda Shelf, Molucca and Flores Seas, Makassar Strait, and Bird's Head region of northwest New Guinea emerging as locations of genetic discontinuities [21], [22]. These locations span the archipelago commonly referred to as Wallacea, which falls between the Sunda (southeast Asia) and Sahul (Australia and New Guinea) continental shelves and was the only point of permanent oceanic connection between the Indian and Pacific Oceans throughout the Pleistocene [23].
Figure 1. Study region.

The light grey outline represents the lowest Pleistocene sea level (120 m depth contour).
Phylogeographic and population genetic surveys have intensely sampled T. maxima and T. crocea throughout Wallacea using mitochondrial (mtDNA) markers [24]–[26], allozymes [27], [28], and microsatellites [29]. Both T. crocea and T. maxima have been shown to contain distinct mtDNA clades associated with Sumatra (Sunda), Wallacea, and northwest New Guinea (Sahul, particularly in Cenderawasih Bay) [24]–[26]. These lineages are sympatric in some populations, for instance T. maxima from northern Java has both Sumatran and Wallacean mitotypes, and similarly T. crocea populations from Halmahera eastward through Cenderawasih Bay contain both Wallacean and northwest New Guinean lineages [26], [29]. Microsatellite genotyping of T. crocea corroborates the distinctiveness of Sumatran and Cenderawasih populations, with evidence for mixing in Wallacea of local genotypes with Cenderawasih-like genotypes [29]. Thus, substantial genetic differentiation typifies at least two Tridacna species in this region.
In the Pacific Ocean, T. derasa, T. gigas, T. maxima and T. crocea have been genetically surveyed, primarily with allozyme markers [27], [28], [30]–[35], but also with mtDNA [36]. These studies show genetic divisions between western and central Pacific populations but with some indication that eastern Australian populations show greater affinities with Philippine populations than they do with other western Pacific populations [30], [33]. Great Barrier Reef populations (eastern Sahul) form a cluster distinct from, but closely related to, Philippine populations for T. maxima and T. derasa but with low sampling in the Philippines (two and one populations, respectively) and no sampling in Wallacea or Sunda regions. Thus, it is unknown whether substantial genetic divergence reflects the geographic distance separating the Philippines and eastern Sahul or is indicative of distinct regional groupings.
Here, we examine DNA sequence diversity of T. crocea and T. maxima whose sampled distributions include the eastern Indian Ocean, Wallacea, and western Pacific Oceans. Data from new samples, predominantly from the western Pacific, are merged with data from previous studies, especially from Wallacea (e.g. [24], [25], [26]), to present a unified summary of phylogeographic patterns and a point of contrast to earlier broadscale studies based on allozymes [30], [32], [33], [35]. We use phylogenetic analyses to assess evolutionary relationships among species and also gauge regional geographical divisions within species.
Materials and Methods
Sampling and permits
Small mantle biopsies were non-lethally collected from animals with morphology characteristic of Tridacna maxima and T. crocea at 0–20 m depth from the Solomon Islands, and in Australia from Ningaloo Reef, Heron Island, Lizard Island, the Torres Strait and Lihou Reef. All sampling and tissue transport was in accordance with local and international regulations. Permit details are as follows: Lihou Reef, Australia: Department of Sustainability, Environment, Water, Population & Communities (Access to Biological Resources in a Commonwealth Area for Non-Commercial Purposes permit number: AU-COM2008042); Lizard Island and Heron Island, Australia: Great Barrier Reef Marine Park Authority and Queensland Parks and Wildlife (Marine Parks Permits: G08/28114.1, G09/31678.1, G10/33597.1, G11/34640.1); Ningaloo Reef, Australia: Western Australia Department of Environment and Conservation (License to take Fauna for Scientific Purposes: SF007126, SF006619, SF008861; Authority to Enter Calm Land/or Waters: CE002227, CE002627, Department of Fisheries, Western Australia Exemption 2046); Queensland: Queensland Government Department of Primary Industries (General Fisheries Permits: 118636, 150981); Torres Strait Islands, Australia: Commonwealth of Australia Torres Strait Fisheries Act 1984 and Australian Fisheries Management Authority (Permit for Scientific Purposes: 8562); Solomon Islands: Solomon Islands Government Ministry of Education and Human Resource Development and Ministry of Fisheries and Marine Resources (research permit: to S Albert, expiry 31/10/2011); Solomon Islands Government Ministry of Environment, Conservation and Meteorology (Convention on International Trade in Endangered Species of Wild Fauna and Flora export permit: EX2010/102); Australian Government Department of the Environment, Water, Heritage and the Arts (Convention on International Trade in Endangered Species of Wild Fauna and Flora import permit: 2010-AU-616020); Australian Quarantine Inspection Service (Permit to Import Quarantine Material: IP10017966).
DNA sequences
DNA was extracted using a modification of the Qiagen DNeasy protocol [37]. Primers that targeted mitochondrial cytochrome oxidase 1 (COI) [24], [26], [38] and ribosomal 16S [39] were used to amplify 390 and 417 basepair segments of the respective gene regions. A subset of samples were amplified for the partial nuclear 18S and ITS1 region (referred to as ITS in text) to provide independent estimates of phylogenetic relationships using primers from [13], [40]. PCR products were purified following a standard Exo-Sap protocol (New England Biolabs) and were sequenced by Macrogen (Korea). Trace files were edited in CodonCode Aligner (ver. 4.0.3). In addition, the NCBI repository of nucleotide sequences was searched for all published Tridacna COI and 16S sequences (August 2012) representing both intraspecific [24]–[26], [41] and interspecific [9], [15], [16] surveys. These sequences were manually aligned [42] against our new sequences and against outgroups (Hippopus hippopus, Hippopus porcellanus, Cerastoderma glaucum, Fragum sueziense, and Corculum cardissa) and trimmed to a common length. For ITS there were several insertions/deletions that could not be reconciled, so these areas of low overlap were masked and not used for phylogenetic analyses.
Phylogenetic analyses
Previous mtDNA surveys have used either 16S [9], [15], [26] or COI [24]–[26], [41] gene regions. To unify these sources of data and address interspecific relationships, we initially took representative sequences across studies and linked them by our samples for which both gene regions had been sequenced in a concatenated search. For samples with only a single gene region (that is, sequences acquired from NCBI), information from the missing gene region was treated as missing data. Up to four individuals per species were retained representing the diversity of their species clade and prioritizing individuals with both 16S and COI sequenced. Using StarBEAST v. 1.6.2 [43] each mtDNA gene region was treated as a separate partition. A general time reversible model with gamma distributed and invariant sites (GTR+G+I) was applied to each gene, with additional partitioning by codon position (1+2, 3) for COI. A relaxed molecular clock with an uncorrelated lognormal mutation rate was used for each gene. The COI and 16S gene trees were linked, as mtDNA is a single linked locus (i.e. concatenated gene regions). Priors were set for nodes defining species as a log normal date (mean = 0, SD = 1) with an offset representing the most recent estimate of the earliest fossil (T. crocea: 1.8, T. maxima: 5.3, and T. squamosa: 1.8 million years). The root of the Tridacninae was set as normal with mean date of 14 and SD of 2.5 million years. All fossil dates were based on [15], [44]. Speciation was modeled both as birth-death and Yule processes in independent runs of 250 million steps, with a burn-in of 25%, and yielded similar results.
Additional genealogical searches were performed using MrBayes ver. 3.1.2 [45] and RAxML (Randomized Axelerated Maximum Likelihood, Blackbox interface) [46]. Using the concatenated file of the same mtDNA sequences as above, searches were partitioned such that 16S formed one partition, and COI formed a second partition with third codon positions partitioned separately from first and second (1+2, 3) for COI. In MrBayes, a GTR+G+I (nst = 6, invgamma) model for all three partitions was used, with a search length of 10 million steps, sampling every 10,000 steps, and a burn-in of 25% (2.5 mill steps). Similarly, the GTR+G+I models were applied to these partitions in RAxML in a maximum likelihood search with 100 bootstrap replicates.
Locus-specific genealogies were also inferred for COI, 16S, and ITS using both MrBayes and RAxML. Total data sets for each locus were assembled from all available sequences and then simplified by removing any identical haplotypes. Searches were performed under the same conditions previously described for 16S (no partitions) and for COI (1+2, 3) with four separate searches of 10 million steps and the final 25% percent of trees retained (effectively a burn-in of 7.5 million steps). Search conditions for the partial nuclear ITS sequences were as above with indels treated as missing data and no partitioning.
The software Figtree (Rambaut: http://tree.bio.ed.ac.uk/software/figtree/) was used to assist with tree visualization and graphics preparation.
Phylogeographic patterns
Intraspecific phylogeographic patterns were assessed by examining all available COI and 16S sequences for T. crocea, T. maxima, and the distinct clade (Tridacna sp.) identified in the previous analyses. For each species-locus combination, a heuristic maximum parsimony search was conducted in PAUP* [47]. Because frequencies of published haplotypes are not consistently available, it was not possible to conduct standard population genetic analyses such as measures of diversity and differentiation. For intraspecific parsimony searches, the maximum number of trees was set to 1000 in PAUP*[47].
Results
DNA sequences
New DNA sequence data was generated for individuals from five locations (including 55 COI, 65 16S, and 50 ITS sequences: Genbank Acc. Nos. JX974838-JX975007). Combining these new sequence data with previously published data yielded aggregations of 405 COI, 132 16S, and 50 ITS sequences for Tridacna species, with 335 unique haplotypes for COI and 54 unique haplotypes for 16S. In the new data generated for this study nearly all included individuals were sequenced for both COI and 16S allowing us to link results from these two loci and provide a common context for the aggregated sequences from previous studies. Similarly, ITS sequences were obtained from an overlapping subset of individuals sequenced for COI and 16S. Nexus files have been deposited in Treebase (http://purl.org/phylo/treebase/phylows/study/TB2:S13501).
Phylogenetic analyses
Phylogenetic analyses resulted in well-resolved topologies defining several clades within Tridacna. Tree topologies for the concatenated and single gene datasets were similar (Figs. 2–4), providing evidence for a robust and consistent phylogenetic signal. The concatenated analyses of mitochondrial COI and 16S loci (Fig. 2) strongly support monophyly of T. squamosa, T. crocea, and a previously undescribed clade (but reported in [16]) formed well-supported terminal taxa, with more modest support for the monophyly of T. maxima. This undescribed clade (which we refer to as Tridacna sp.) was also well supported in single gene analyses of COI and 16S (Fig. 3) and ITS (Fig. 4). T. sp. sequences were evolutionarily distinct from other species; the average pairwise COI sequence divergence between T. sp and T. crocea was 14.4% and was 12.6% between T. sp. and T. squamosa, as compared to 9.5% between T. crocea and T. squamosa (uncorrected pairwise distances).
Figure 2. Species relationships within Tridacna based on concatenated mitochondrial DNA (COI and 16S) sequences.

The topology shown is a time calibrated maximum clade credibility tree inferred with StarBEAST under a birth-death model. Bayesian posterior probabilities from StarBEAST and MrBayes are above branches and RAxML bootstrap support percentages are below branches. Individuals with two accession numbers include both COI and 16S sequences. Individuals that are underlined also appear in Fig. 4.
Figure 4. Species relationships within Tridacna based an ITS MrBayes consensus tree.
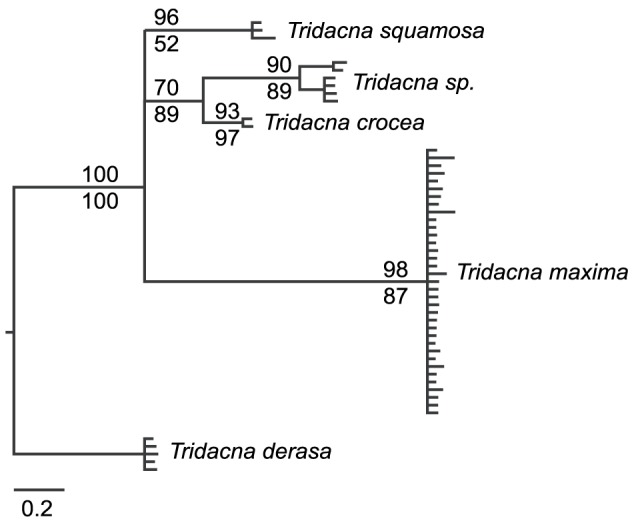
Unalignable regions have been excluded. Bayesian posterior probabilities are above branches and RAxML bootstrap support percentages are below branches.
Figure 3. Bayesian phylogenetic trees for mitochondrial CO1 and 16S.

MrBayesian consensus trees constructed for each gene region using all available data. Although different species and regions have differential representation, the two gene trees are concordant, as is expected for linked loci. Thus, overall patterns are consistent among research groups. Branch colors correspond to distinct lineages whose geographic distributions are described in Fig. 5.
Gene trees for COI and 16S show concordant relationships among species (Fig. 3), confirming that independent research groups have sampled similar genotypes. The notable exception to the consistency across studies was the 16S T. derasa sequence from [15] which did not cluster consistently with our 16S T. derasa sequence (specimen ET358) even though our COI sequence from this same individual clustered with other T. derasa sequences including GQ166591 from [48]. For this reason, the T. derasa sequence from [15] was retained in the 16S tree, but excluded from the joint COI and 16S searches. All mtDNA-based genealogies supported T. squamosa and T. crocea as sister species (Figs. 2 and 3) whereas ITS based analyses gave modest support for T. sp. and T. crocea as sister species (Fig. 4). Within the mtDNA-based analyses, T. derasa, T. gigas, and T. mbalavuana appear consistently as basal lineages within Tridacna (Figs. 2 and 3). (No ITS sequences were available for these taxa.)
Phylogeographic patterns
Within T. crocea and T. maxima, there was broadscale phylogeographic concordance of mtDNA gene trees (as shown in Fig. 5). T. crocea and T. maxima haplotypes from the Solomon Islands, the Torres Strait and Lizard Island (and additionally western New Guinea/Cenderwasih Bay, Lihou Reef and Heron Island for T. maxima) formed a distinct monophyletic ‘Pacific’ group (colored blue in Fig. 5). Sequences from the Sunda Shelf formed a second monophyletic group (colored orange in Fig. 5) as described in the original publications [24]–[26], although the location or the genetic break differed slightly for each species. Finally, sequences from Indonesia, Singapore, western New Guinea/Cenderwasih Bay and Taiwan formed a third group (black in Fig. 5). Most sequences published in Genbank are not georeferenced. We were, however, able to deduce the distinct clades typifying major regions from previously published surveys by recreating previously published analyses; T. crocea (yellow haplotypes of [24], grey clade of [26]) are shown in green and orange respectively, and T. maxima (yellow haplotypes of [25]) are shown in blue and orange respectively in Fig. 5.
Figure 5. Unrooted parsimony trees and sampling locations for Tridacna crocea, Tridacna sp., and Tridacna maxima.

Major lineages on networks are colored and the geographic extent of each lineage is indicated on the map. Relative frequencies of each haplotype are not depicted; each haplotype is shown in equal size (see text). Dots on maps indicate sampling locations and locations with two distinct sympatric lineages are shown as bisected circles (not indicative of relative frequencies). Support for monophyly of major clades among COI trees is based on 100 percent consistency of each branch among all equally parsimonious trees (a randomly chosen tree is depicted). Among 16S trees, the single most parsimonious tree for T. sp. and T. maxima are shown, and for T. crocea both green and blue lineages were present in all six equally parsimonious trees. Colors indicate geographic locations of haplotypes and internal branches are in gray.
For T. maxima, the northwest New Guinea clade formed a cluster with the Pacific clade, although no haplotypes were shared between the two locations. For T. crocea, however, haplotypes from northwest New Guinea and the western Pacific were members of two distinct monophyletic groups: the Pacific (blue) and the Wallacea (black) groups (Fig. 5). The T. crocea and T. maxima 16S sequences from [15], described as having been obtained from individuals sourced from aquarium stores, both fell within Pacific haplotype groups, suggesting that these purchased specimens had a Pacific origin.
Despite the reduced sampling for T. sp., a ‘Pacific’ lineage was similarly positioned in the Solomon Islands, and a distinct lineage, comprising samples from western Australia and Taiwan, geographically overlapped with the Wallacea (black) lineage portrayed in T. crocea and T. maxima. Similar phylogeographic patterns were evident for COI and 16S for each species despite only partially overlapping sets of individuals forming the basis for each tree.
Discussion
Despite their distinct shell morphology and longstanding cultural and commercial significance, our data reveal cryptic diversity within giant clams. Here, we find a previously undescribed clade of Tridacna (Tridacna sp.). This clade is supported by both mtDNA and nuclear gene regions (Figs. 2–4), which identify it as a unique, evolutionarily significant unit [49] with reference to previously described species. Our molecular phylogenetic analyses place T. sp. as a sister clade to T. squamosa and/or T. crocea, but in no instance was a close relationship between T. sp. and T. maxima suggested in our gene trees. Thus, molecular data do not support T. sp. being a variety of T. maxima as was suggested by Tang [16]. Clams with T. sp. mitotypes were found both at Ningaloo Reef in western Australia and in the Solomon Islands. Although only T. sp. and T. squamosa were identified among our clam samples from Ningaloo, it is likely that T. maxima also occur at Ningaloo (Penny unpub., [50]), and we found T. sp. sympatric with T. maxima and T. crocea in the Solomons.
The T. sp. clade includes the single haplotype (COI and a 16S) described from Taiwan [16]. Tang et al. (2005) suggested that there are morphological differences between T. sp. and T. maxima, including mantle pattern, shell lip shape, posterior adductor weight and the position of the incurrent aperture. Qualitative examination of an individual from Ningaloo Reef with T. sp. mtDNA shows shell characters typical of T. maxima: asymmetry of the valve with posterior elongation and dense rows of scales on folds (Fig. 6). T. maxima is well known for its morphological variability [51] and thus it is possible that previous morphological examinations of T. sp may have been identified it as T. maxima. (Additional morphological samples are not presently available as most collecting permits only allow non-lethal sampling of giant clams.) Our findings, therefore, lend support to Tang's conclusion that T. sp. is an undescribed species but we show that, rather than being a narrow-range endemic (such as Tridacna rosewateri from Mauritius [10]), T. sp. is widely distributed. Although it is not possible at present to delineate the distribution of T. sp., it seems probable that T. sp. occurs at locations in between Australia, Taiwan and the Solomon Islands. T. sp. individuals from the western Pacific were reciprocally monophyletic from the individuals from Ningaloo (Indian Ocean) and the single sequence from Taiwan (Fig. 5).
Figure 6. An individual with Tridacna sp. mtDNA demonstrating valve morphology consistent with Tridacna maxima.

A) Tridacna maxima from Hibernia Reef, WA, Australia. Accession No# P.52722 (Museum Art Gallery Northern Territory (MAGNT)), original identification based on morphology, B) Tridacna sp. from Five Finger Reef, South of Coral Bay, Ningaloo Marine Park, WA, Australia. Accession No#. P.51911 (Museum Art Gallery Northern Territory (MAGNT)), C) Tridacna maxima, from north western WA, Australia, unregistered (Museum Art Gallery Northern Territory (MAGNT)). Photo credit: Shane Penny.
MtDNA genealogies place T. sp as sister species to T. crocea and T. squamosa, with strong support for monophyly of this group of three species (Figs. 2 and 3). Tridacna maxima and T. squamosina formed a second clade, but with less support across phylogenetic analyses (Fig. 2) probably because only 16S sequences were available for T. squamosina. Monophyly of T. crocea and T. squamosa was reported in previous mtDNA based phylogenetic analyses [9], [15], but not in allozyme analyses [14] where T. squamosa was sister to T. crocea and T. maxima. Monophyly of the Chametrachea subgenus (including T. squamosina, T. crocea, T. maxima, T. sp. and T. squamosa) [15], [44] was supported in individual gene analyses and the concatenated StarBEAST searches (Fig. 2). Monophyly of the Tridacna subgenus (including T. derasa, T. mbalavuana, and T. gigas) was not well supported in any of our mtDNA analyses, with these taxa appearing basal to the Chametrachea, but missing and non-overlapping data may have contributed to the low resolution.
Previous phylogeographic studies of T. crocea [24], [26], [29] and T. maxima [25] from Indonesia show geographic restriction of several clades. The mtDNA gene trees within these papers delineate clusters comprising haplotypes from western Sumatra (Sunda), Wallacea, and northwest New Guinea (Sahul) [24]–[26], [29] with some mixing between clades particularly in the Bird's Head Peninsula of northwest New Guinea [26]. Our samples showed an additional and deeper evolutionary break for T. crocea to the east of Cenderawasih Bay, whereby individuals from the Solomon Islands, Torres Strait, and Great Barrier Reef form a monophyletic group and do not share any mtDNA haplotypes with northwest New Guinea or locations in Wallacea (Fig. 5). Therefore, it appears that the distinct clade of T. crocea haplotypes from northwest New Guinea (with some spillover westward into Wallacea [26]) is regionally endemic and does not extend into the west Pacific. These patterns are not due to differences in DNA sequencing interpretation between research groups, as samples (from [24], [26], [41]) are mutually consistent and a single T. crocea (from [15]) falls within the larger Pacific T. crocea clade. Based on present sampling, we can place this newly discovered genetic discontinuity between Cenderawasih Bay and the Solomon Islands in the north and between the Aru Basin and Torres Strait in the south. For T. maxima, in contrast, the distinct haplotypes from northwest New Guinea fall in the same clade as west Pacific haplotypes. Thus the northwest New Guinea clade of T. maxima can now be viewed as a westward extension of Pacific variants, albeit with no shared haplotypes between locations.
With only two species to compare, we can only speculate as to why the mtDNA patterns differ between species, although greater overall population genetic structure in T. crocea compared to T. maxima is consistent with previously co-sampled regions (for instance, [24] in comparison to [25]). Because of the diffuse sampling for T. crocea, we cannot pinpoint a specific location of geographic differentiation east of Cenderawasih Bay, yet at a macroscale this observation is consistent with mtDNA patterns in a butterflyfish [52], a reef fish [53], and a sea star (Crandall pers. comm.) and may be associated with a long stretch (>700 km) of coastline east of Cenderawasih Bay with sparse reef habitat [54].
In T. maxima, we found that Solomon Islands haplotypes cluster with haplotypes from the Great Barrier Reef; this affinity contrasts with allozyme results that show substantial divergence between Solomon Islands and Great Barrier Reef populations [33]. The nature of these differing patterns cannot be explored further as allozyme results are not directly comparable across research groups.
The broadscale geographic and multispecies phylogenetic results of this study, consolidated with those of previous investigations, reveal new aspects of regional patterns and highlight key uncertainties in the current knowledge of Tridacna. A common result among population genetic studies of Tridacna species to date is that there is substantial population structure. Such genetic differentiation may be due in part to the relatively short planktonic larval duration of approximately 9 days [12] that is likely to restrict dispersal distances. The discovery of an undescribed species adds to other recent species discoveries in Tridacna [9]–[11], but the broad distribution of T. sp. illustrates that cryptic species can remain undetected even in such conspicuous groups as giant clams.
Both the discovery of a new species and the observation of substantial geographic differentiation are relevant to monitoring of local stocks and human transport of clams. First, the presence of a cryptic sympatric species would result in overestimates of species abundance where clam populations are censused. Second, human-aided movements could cause species to be introduced to regions outside their natural range and, similarly, are likely to introduce foreign genetic material into local populations. Tridacna maxima, T. squamosa, T. derasa, T. mbalavuana and T. gigas were frequently translocated during the 1980's and 1990's (some human assisted movements continuing into this century) by governmental, commercial and conservation organizations to combat local depletion and facilitate the live culture trade [55]. Third, depleted populations are unlikely to receive immigrants from geographically distant locations via planktonic dispersal and, therefore, recovery may be slow or negligible even when local harvesting has ceased. Results from giant clams underscore two important themes emerging from genetic investigations of marine organisms: cryptic species are common [19], [20], [56], [57], and many species are genetically heterogeneous across their geographic range [58].
Supporting Information
Genbank accession numbers for all included sequences.
(XLS)
Acknowledgments
Sampling in the Coral Sea was supported by the Marine Division of the Australian Government Department of Sustainability, Environment, Water, Population & Communities. We are grateful to the staff of the Australian Museum's Lizard Island Research Station and the Heron Island Research Station for their facilities and support. Sampling in the Torres Strait Islands was assisted by the staff and students of Tagai State College, Thursday Island Primary and the Torres Strait Regional Authority. Sampling in the Solomon Islands was made possible via the Pacific Strategy Assistance Program within the Australian Government Department of Climate Change and Energy Efficiency and with the assistance of the Roviana Conservation Foundation. We especially thank JD Aguirre, S Albert, A Denzin, N Gemmell, M Jimuru, F MacGregor, V McGrath (Senior Community Liaison Officer, Land and Sea Management Unit, Torres Strait Regional Authority), A Mirams, R Pearce, Stephen, Lavud and Takenda for their logistical support and field assistance. JS Lucas, LG Cook, A Toon, L Pope and JM Pandolfi provided helpful comments and suggestions, as did several anonymous reviewers.
Funding Statement
Funding for this work was provided by the Australian Research Council (www.arc.gov.au, DP0878306 to CR), the German Research Foundation (www.dfg.de/en, DFG, Hu 1806/1-1, Hu 1806/2-1 to TH), the World Wildlife Fund (worldwildlife.org/initiatives/fuller-science-for-nature-fund, Kathryn Fuller Post-doctoral Research Fellowship to EAT), the Malacological Society of Australasia (www.malsocaus.org, to TH), and the Joyce Vickery Fund (linneansocietynsw.org.au/grants.html, to JK). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Heslinga G (1996) Clams to cash: How to make and sell giant clam shell products. Hawaii, USA: Center for Tropical and Subtropical Aquaculture. Publication Number 125: 47. [Google Scholar]
- 2.Wabnitz C, Taylor M, Green E, Razak T (2003) From ocean to aquarium: The global trade in marine ornamental species. Cambridge, UK: UNEP World Conservation Monitoring Centre. 64 p. [Google Scholar]
- 3.Tisdell C, Menz KM (1992) Giant clam farming and sustainable development: An overview. In: Tisdell C, editor. Giant clams in the sustainable development of the South Pacific: Socioeconomic issues in mariculture and conservation. Canberra, Australia: ACIAR pp. 3–14.
- 4.Tomascik T, Mah AJ, Nontji A, Moosa MK (1997) The ecology of the Indonesian seas: Part one. Hong Kong: Periplus Editions. 656 p. [Google Scholar]
- 5.Wells SM (1997) Giant clams: Status, trade and mariculture, and the role of CITES in management. Gland, Switzerland and Cambridge, UK: IUCN-The World Conservation Union. 77 p. [Google Scholar]
- 6.UNEP-WCMC (2013) Convention on international trade in endangered species of wild fauna and flora. Available: www.cites.org. Accessed 2013 October 20.
- 7.IUCN (2013) IUCN red list of threatened species. Available: www.IUCNredlist.org. Accessed 2013 October 20.
- 8. bin Othman AS, Goh GHS, Todd PA (2010) The distribution and status of giant clams (family Tridacnidae) - a short review. Raffles Bull Zool 58: 103–111. [Google Scholar]
- 9. Richter C, Roa-Quiaoit H, Jantzen C, Al-Zibdah M, Kochzius M (2008) Collapse of a new living species of giant clam in the Red Sea. Curr Biol 18: 1349–1354. [DOI] [PubMed] [Google Scholar]
- 10. Sirenko BI, Scarlato OA (1991) Tridacna rosewateri sp. n., a new species of giant clam from the Indian Ocean. La Conchiglia 22: 4–9. [Google Scholar]
- 11. Lucas JS, Ledua E, Braley RD (1991) Tridacna tevoroa Lucas, Ledua and Braley: A recently-described species of giant clam (Bivalvia; Tridacnidae) from Fiji and Tonga. Nautilus 105: 92–103. [Google Scholar]
- 12.Lucas JS (1988) Giant clams: Description, distribution and life history. In: Copland JW, LucasJS, editor. Giant clams in Asia and the Pacific. pp. 21–32.
- 13. Maruyama T, Ishikura M, Yamazaki S, Kanai S (1998) Molecular phylogeny of zooxanthellate bivalves. Biol Bull 195: 70–77. [DOI] [PubMed] [Google Scholar]
- 14. Benzie JAH, Williams ST (1998) Phylogenetic relationships among giant clam species (Mollusca: Tridacnidae) determined by protein electrophoresis. Mar Biol 132: 123–133. [Google Scholar]
- 15. Schneider JA, O'Foighil D (1999) Phylogeny of giant clams (Cardiidae: Tridacninae) based on partial mitochondrial 16S rDNA gene sequences. Mol Phyl Evol 13: 59–66. [DOI] [PubMed] [Google Scholar]
- 16.Tang YC (2005) The systematic status of Tridacna maxima (Bivalvia: Tridacnidae) based on morphological and molecular evidence. Master's thesis. Keelung, Taiwan, Republic of China: National Taiwan Ocean University. 80 p. [Google Scholar]
- 17.Hoeksema B (2007) Delineation of the Indo-Malayan centre of maximum marine biodiversity: The Coral Triangle. In: Renema W, editor. Biogeography, time, and place: Distributions, barriers, and islands. pp. 117–178.
- 18.Ekman S (1953) Zoogeography of the sea. London: Sidgwick and Jackson. 417 p. [Google Scholar]
- 19. Vogler C, Benzie J, Lessios H, Barber P, Wörheide G (2008) A threat to coral reefs multiplied? Four species of crown-of-thorns starfish. Biol Lett 4: 696–699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Duda TF, Kohn AJ, Matheny AM (2009) Cryptic species differentiated in Conus ebraeus, a widespread tropical marine gastropod. Biol. Bull. 217: 292–305. [DOI] [PubMed] [Google Scholar]
- 21. Carpenter KE, Barber PH, Crandall ED, Ablan-Lagman MCA, Ambariyanto, et al (2010) Comparative phylogeography of the Coral Triangle and implications for marine management. J Mar Biol 2011: 1–14. [Google Scholar]
- 22. Barber PH, Erdmann MV, Palumbi SR (2006) Comparative phylogeography of three codistributed stomatopods: Origins and timing of regional lineage diversification in the Coral Triangle. Evolution 60: 1825–1839. [PubMed] [Google Scholar]
- 23. Voris HK (2000) Maps of Pleistocene sea levels in southeast Asia: Shorelines, river systems and time durations. J Biogeogr 27: 1153–1167. [Google Scholar]
- 24. Kochzius M, Nuryanto A (2008) Strong genetic population structure in the boring giant clam, Tridacna crocea, across the Indo-Malay archipelago: Implications related to evolutionary processes and connectivity. Mol Ecol 17: 3775–3787. [DOI] [PubMed] [Google Scholar]
- 25. Nuryanto A, Kochzius M (2009) Highly restricted gene flow and deep evolutionary lineages in the giant clam Tridacna maxima . Coral Reefs 28: 607–619. [Google Scholar]
- 26. DeBoer TS, Subia MD, Ambariyanto, Erdmann MV, Kovitvongsa K, et al. (2008) Phylogeography and limited genetic connectivity in the endangered boring giant clam across the Coral Triangle. Cons Biol 22: 1255–1266. [DOI] [PubMed] [Google Scholar]
- 27. Ravago-Gotanco RG, Magsino RM, Juinio-Meñez MA (2007) Influence of the North Equatorial Current on the population genetic structure of Tridacna crocea (Mollusca: Tridacnidae) along the eastern Philippine seaboard. Mar Ecol Prog Ser 336: 161–168. [Google Scholar]
- 28. Juinio-Meñez MA, Magsino RM, Ravago-Gotanco RG, Yu ET (2003) Genetic structure of Linckia laevigata and Tridacna crocea populations in the Palawan shelf and shoal reefs. Mar Biol 142: 717–726. [Google Scholar]
- 29.DeBoer TS, Naguit MR, Erdmann MV, Ablan-Lagman MCA, Ambariyanto, et al. (2014) Concordant phylogenetic patterns inferred from mitochondrial and microsatellite DNA in the giant clam Tridacna crocea. Bull Mar Sci in press.
- 30. Macaranas JM, Ablan CA, Pante MJR, Benzie JAH, Williams ST (1992) Genetic structure of giant clam (Tridacna derasa) populations from reefs in the Indo-Pacific. Mar Biol 113: 231–238. [Google Scholar]
- 31. Laurant V, Planes S, Salvat B (2002) High variability of genetic pattern in giant clam (Tridacna maxima) populations within French Polynesia. Biol J Linn Soc 77: 221–231. [Google Scholar]
- 32. Benzie JAH, Williams ST (1995) Gene flow among giant clam (Tridacna gigas) populations in the Pacific does not parallel ocean circulation. Mar Biol 123: 781–787. [Google Scholar]
- 33. Benzie JAH, Williams ST (1997) Genetic structure of giant clam (Tridacna maxima) populations in the west Pacific is not consistent with dispersal by present-day ocean currents. Evolution 51: 768–783. [DOI] [PubMed] [Google Scholar]
- 34. Benzie JAH, Williams ST (1992) No genetic differentiation of giant clam (Tridacna gigas) populations in the Great Barrier Reef, Australia. Mar Biol 113: 373–377. [Google Scholar]
- 35. Benzie JAH, Williams ST (1992) Genetic structure of giant clam (Tridacna maxima) populations from reefs in the western Coral Sea. Coral Reefs 11: 135–141. [Google Scholar]
- 36. Gardner JPA, Boesche C, Meyer JM, Wood AR (2012) Analyses of DNA obtained from shells and brine-preserved meat of the giant clam Tridacna maxima from the central Pacific Ocean. Mar Ecol Prog Ser 453: 297–301. [Google Scholar]
- 37. Huelsken T, Schreiber S, Hollmann M (2011) COI amplification success from mucus-rich marine gastropods (Gastropoda: Naticidae) depends on DNA extraction method and preserving agent. Mitteilungen der Deutschen Malakozoologischen Gesellschaft 85: 17–26. [Google Scholar]
- 38. Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotech 3: 294–299. [PubMed] [Google Scholar]
- 39.Palumbi S (1996) Nucleic acids ii: The polymerase chain reaction. In: Hillis DM Moritz C, Mable BK, editors. Molecular systematics. Sunderland, Massachusetts: Sinauer & Associates Inc. pp. 205–247.
- 40. Yu ET, Juinio-Menez MA, Monje VD (2000) Sequence variation in the ribosomal DNA internal transcribed spacer of Tridacna crocea . Mar Biotechnol 2: 511–516. [DOI] [PubMed] [Google Scholar]
- 41. Neo ML, Todd PA (2012) Population density and genetic structure of the giant clams Tridacna crocea and T. squamosa on Singapore's reefs. Aquatic Biology 14: 265–275. [Google Scholar]
- 42.Rambaut AE (1996) Se-Al: Sequence alignment editor v2.0a11. Available: http://tree.bio.ed.ac.uk/software/. Accessed 2013 October 20.
- 43. Heled J, Drummond AJ (2010) Bayesian inference of species trees from multilocus data. Mol Biol Evol 27: 570–580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Harzhauser M, Mandic O, Piller WE, Reuter M, Kroh A (2008) Tracing back the origin of the Indo-Pacific mollusc fauna: Basal Tridacninae from the Oligocene and Miocene of the Sultanate of Oman. Palaeontology 51: 199–213. [Google Scholar]
- 45. Ronquist F, Huelsenbeck JP (2003) MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19: 1572–1574. [DOI] [PubMed] [Google Scholar]
- 46. Stamatakis A, Hoover P, Rougemont J (2008) A rapid bootstrap algorithm for the RAxML web-servers. Syst Biol 57: 758–771. [DOI] [PubMed] [Google Scholar]
- 47.Swofford DL (1998) PAUP*, phylogenetic analysis using parsimony (*and other methods). Version 4. Sunderland, Massachusetts: Sinauer Associates.
- 48. Plazzi F, Ceregato A, Taviani M, Passamonti M (2011) A molecular phylogeny of bivalve mollusks: Ancient radiations and divergences as revealed by mitochondrial genes. PLoSOne 6: e27147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Moritz C, Dowling TE, Brown WM (1987) Evolution of animal mitochondrial DNA: Relevance for population biology and systematics. Ann Rev Ecol Syst 18: 269–292. [Google Scholar]
- 50. Black R, Johnson MS, Prince J, Brearley A, Bond T (2011) Evidence of large, local variations in recruitment and mortality in the small giant clam, Tridacna maxima, at Ningaloo Marine Park, Western Australia. Mar FW Res 62: 1318–1326. [Google Scholar]
- 51. Rosewater J (1965) The family Tridacnidae in the Indo-Pacific. Mollusc Res 1: 347–396. [Google Scholar]
- 52. McMillan WO, Weigt LA, Palumbi SR (1999) Color pattern evolution, assortative mating, and genetic differentiation in brightly colored butterflyfishes (Chaetodontidae). Evolution 53: 247–260. [DOI] [PubMed] [Google Scholar]
- 53.Sims CA, Riginos C, Blomberg SP, Huelsken T, Drew J, et al. (2014) Cleaning up the biogeography of Labroides dimidiatus using phylogenetics and morphometrics. Coral Reefs: in press.
- 54. Spalding MD, Fox HE, Allen GE, Davidson N, Ferdaña ZA, et al. (2007) Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. Bioscience 57: 573–583. [Google Scholar]
- 55.Kinch J, Teitelbaum A (2009) Proceedings of the regional workshop on the management of sustainable fisheries for giant clams (Tridacnidae) and CITES capacity building. Nadi, Fiji: Secretariat of the Pacific Community. 54 p. [Google Scholar]
- 56. Bickford D, Lohman DJ, Sodhi NS, Ng PK, Meier R, et al. (2006) Cryptic species as a window on diversity and conservation. Tr Ecol Evol 22: 148–155. [DOI] [PubMed] [Google Scholar]
- 57. Knowlton N (1993) Sibling species in the sea. Ann Rev Ecol Syst 24: 189–216. [Google Scholar]
- 58. Hellberg ME (2009) Gene flow and isolation among populations of marine animals. Ann Rev Ecol Syst 40: 291–310. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Genbank accession numbers for all included sequences.
(XLS)