

Abstract
Previously, we demonstrated that prostaglandin E2 (PGE2) induces C-C chemokine receptor type 7 (CCR7) expression on human monocytes, which stimulates their subsequent migration in response to the CCR7 natural ligands CCL19 and CCL21. In this study, we determined whether PGE2 affects CCR7 expression on macrophages. Flow cytometric analysis and chemotaxis assays were performed on Mono Mac-1-derived macrophage (MDMM-1) as well as unpolarized monocyte-derived macrophages (MDMs) to determine the CCR7 expression and functionality in the presence of PGE2. Data revealed that a MDMM-1 exhibited markedly downregulated CCR7 expression and functionality that were partially restored by treatment with PGE2. In MDMs, we observed a drastic downregulation of CCR7 expression and functionality that were unaffected following PGE2 treatment. Our data indicate that monocyte differentiation induces the loss of CCR7 expression and that PGE2 is unable to modulate CCR7 expression and functionality as shown previously in monocytes.
1. Introduction
Monocytes and macrophages orchestrate proper immune responses to pathogens. Monocytes have been demonstrated to be precursors of professional antigen-presenting cells, such as macrophages and dendritic cells (DCs) [1–3]. In response to danger stimuli, circulating blood monocytes migrate into damaged or infected tissues and differentiate into mature macrophages or DCs. After taking up antigens, the activated macrophages and DCs migrate to the draining lymph nodes to present the antigens to T and B cells.
Chemokine receptors confer upon cells the ability to detect and move directionally toward a chemotactic stimulus. C-C chemokine receptor type 7 (CCR7) plays a leading role in the mechanism controlling the entry of lymphocytes and mature DCs into lymph nodes. Within lymph nodes, these cells encounter other immune cells for activation, determining the success of cellular immunity after infection [4]. For immature DCs, the maturation process is initiated upon sensing “danger signals” (tissue damage, inflammatory cytokines, or pathogens) [5]; this process occurs concomitantly with their migration from peripheral tissues to the draining lymph nodes. During maturation, CCR7 expression is upregulated, which guides the migratory DCs to the lymph nodes [6–8]. The chemokines CCL19 and CCL21 are the natural ligands of CCR7, and they are expressed by lymphatic endothelium and/or within lymph nodes by stromal cells, endothelial cells, and DCs [9–13]. Mice deficient in CCL19, CCL21, or CCR7 demonstrate defective DC trafficking and altered immune responses [6, 14, 15].
Recently, CCR7 expression was discovered on human monocytes [16]. As observed with DC migration [17], prostaglandin E2 (PGE2), a pleiotropic immunomodulatory molecule, appears to have a dual role in monocyte migration by regulating the expression and activity of CCR7 through the involvement of EP2 and EP4 receptors [16]. Moreover, in DCs, PGE2 has been reported to synergize with tumor necrosis factor-alpha (TNF-α) to promote CCR7 expression and the chemotactic responsiveness of DCs to CCL19 and CCL21 [17–19]. Recent results indicate that CCL19 binding to CCR7 promotes the activation of p38, extracellular signal-regulated kinase 1/2, and c-Jun N-terminal kinase and leads to monocyte migration. Moreover, the RhoA/ROCK pathway is essential for PGE2-mediated CCR7-dependent monocyte migration [20]. The physiological importance of CCR7 for the immune response has been demonstrated by several studies, and although CCR7 expression and functionality have been extensively characterized in DCs, much less is known about the fate of its expression in macrophages.
As it has been demonstrated that monocytes express the chemokine receptor CCR7 and differentiate upon immune activation, we asked whether monocyte maturation in the presence of PGE2 has a direct effect on CCR7 expression and functionality. In a unpolarized environment, our results demonstrate that monocyte maturation downregulates CCR7 expression in macrophages derived from Mono Mac-1 cells (MDMM-1 cells) and monocyte-derived macrophages (MDMs) from healthy donors. In parallel, chemotaxis assays were performed to establish the influence of differentiation on CCR7 signaling function. Our results clearly indicate that MDMM-1 cells and MDMs lose functional CCR7 expression, which inhibits their migration toward the CCR7 ligands CCL19 and CCL21. Thus, these data reveal that unlike DCs, macrophages lose CCR7 expression and functional activity upon differentiation, even in the presence of PGE2.
2. Materials and Methods
2.1. Reagents
PGE2 was purchased from Sigma Aldrich (Oakville, ON, Canada). The chemokines CCL19 and CCL21 and antibodies anti-CCR7-APC and anti-IgG2a-APC were purchased from R&D Systems (Minneapolis, MN, USA). Anti-CD14-FITC, anti-CD64-FITC (BD Biosciences, Mississauga, ON, Canada), and anti-IgG2a-FITC were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
2.2. Cell Culture
The monocytoid cell line Mono Mac-1 was purchased from the German Collection of Microorganisms and Cell Culture (Braunschweig, Germany). Cells were maintained in RPMI 1640 medium (Wisent, Saint-Bruno, QC, Canada) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Wisent), 1 mM sodium pyruvate (Wisent), 0.1 mM nonessential amino acids, 100 U/mL penicillin G, and 100 μg/mL streptomycin (Wisent). Mono Mac-1 cells were induced to differentiate into macrophage-like (MDMM-1) cells with PMA. Briefly, cells were plated in 100 mm dishes (7 × 106 cells) in fresh medium and incubated with 20 ng/mL PMA for 72 h [21]. Mono Mac-1 and MDMM-1 cells were stimulated for the indicated times using 1 μM PGE2.
2.3. Blood Monocyte Isolation and MDM Differentiation
Total blood mononuclear cells were isolated from the blood of healthy donors using lymphocyte separation Medium 1077 (Sigma) and washed twice in Hank's balanced salt solution (Wisent). Cells were cultured in RPMI 1640 medium, 20% heat-inactivated FBS, and 10% heat-inactivated human serum for 2 h before use. Monocytes were washed from nonadherent cells with phosphate-buffered saline (PBS). Monocytes were then enriched from peripheral blood mononuclear cells using the MACS Monocyte Isolation Kit II and MACS LS Columns (Miltenyi Biotec, Auburn, CA, USA), yielding an average purity of 98%. The purity was assessed by flow cytometric analyses as recommend by the manufacturer, and isolated monocytes were fluorescently stained with CD14-FITC and anti-Biotin-PE that labeled nonmonocytes. To differentiate monocytes into MDMs, cells (2 × 106/mL) were plated in RPMI 1640 medium with 2 mmol/L l-glutamine (Wisent) containing 20% human AB serum (Wisent) in 24-well plates (Millipore, Nepean, ON, Canada). After 24 h, nonadherent cells were removed, and adherent cells were cultured in RPMI with 20% FBS (Wisent) in 5% CO2 at 37°C for 12 days. Medium was refreshed every 4 days. Macrophages were detached using Cell Dissociation Buffer (Wisent), washed with 1× PBS, and used in experiments. Blood monocytes and MDMs were stimulated for the indicated times using 1 μM PGE2.
2.4. Microscopy
Images of whole-cell morphology were acquired by interferential contrast using a Olympus IX70 microscope (Olympus Canada Inc., Richmond Hill, ON, Canada) and processed by Image Pro Plus software (Media Cybernetics Inc., Bethesda, MD, USA).
2.5. Flow Cytometry
Cells were collected and washed in 1× PBS. Fc receptors were blocked using 1 μg of purified IgG (Santa Cruz Biotechnology) for 15 min at room temperature and washed in PBS containing 3% bovine serum albumin (BSA). Cells were labeled with an anti-CCR7 antibody conjugated with allophycocyanin or the corresponding antibody isotype for 45 min on ice and protected from light. Cells were then washed three times with PBS containing 3% BSA. Samples were analyzed in a BD FACSCalibur flow cytometer (BD Biosciences) using CellQuest software (BD Biosciences).
2.6. Chemotaxis Assay
Chemotaxis was measured by migration through a polycarbonate filter with 5 and 8 μm pores for blood cells and Mono Mac-1 cells, respectively, in 96-well Transwell chambers (Millipore). RPMI with 0.25% BSA (100 μL) containing the indicated concentrations of CCL19 or CCL21 or medium alone as a control for spontaneous migration was added to the lower chamber. To the upper chamber, 1 × 105 cells (150 μL) were added. Monocytes and macrophages were then incubated for 4 h at 37°C. A 150 μL aliquot of the cells that migrated to the bottom chamber was counted by flow cytometry in a FACScan that acquired events for a fixed period of 60 s using CellQuest software. Spontaneous migration was subtracted to calculate the specific migration. The percentage of migrated cells was calculated as follows: the number of migrated cells in response to medium only was subtracted from the number of migrated cells in medium supplemented with CCL19 or CCL21, and this number was reported according to the total input of cells. Each experiment was performed in triplicate, and migration assays were repeated at least three times.
2.7. Reverse Transcription (RT) and Real-Time Polymerase Chain Reaction (PCR)
MDMM-1 cells were incubated with or without 1 μM PGE2 for the indicated times, and total RNA was extracted with the Total RNA Kit E.Z.N.A. (Omega Bio-Tek, Mississauga, ON, Canada) according to the manufacturer's instructions. RNA was reverse-transcribed into cDNA in the presence of 200 U of M-MLV RT (Promega, Madison, WI, USA), 0.5 μg of oligonucleotide d(T)15, and 500 μM deoxyribonucleotide triphosphates at 42°C for 1 h. One microliter of cDNA was used and analyzed using the Platinum SYBR Green qPCR SuperMix System from Invitrogen (Burlington, ON, Canada). The PCR mixture consisted of 0.25 μM forward and reverse primers for CCR7 as described previously [16], 0.4 mM deoxyribonucleotide triphosphate, 2 mM MgCl2, and 1.25 U of Taq DNA polymerase (Roche, Indianapolis, IN, USA). The PCR cycling conditions consisted of initial denaturation at 95°C for 10 min and 40 cycles of 95°C for 30 s, 55°C for 45 s, and 72°C for 45 s. Samples were run in an Applied Biosystems 7500 gel electrophoresis apparatus (Applied Biosystems, Foster City, CA, USA), and the results were analyzed using v1.4 software.
2.8. Statistical Analysis
Each experiment was performed at least three times. Statistically significant differences between experimental groups were evaluated using a paired t-test. Computations were performed using GraphPad PRISM version 5.0b statistical software.
3. Results and Discussion
3.1. Surface Expression of CCR7 on Human Monocytes Is Modulated during Differentiation
Mono Mac-1 is a cell line assigned to the monocytic lineage according to morphological, cytochemical, and immunological criteria [22]. Recently, Mono Mac-1 cells and blood-isolated monocytes were shown to express functional CCR7 chemokine receptors that are upregulated by treatment with PGE2 [16]. In this study, we investigated the impact of the maturation of monocytes into macrophages on CCR7 expression and functionality. First, the differentiation of Mono Mac-1 cells into a more mature, macrophage-like phenotype was induced by treatment with 20 ng/mL PMA for 72 h as previously described [21]. Differentiation was assessed by morphological changes such as cell clustering, cellular adhesion to the bottom surface of the culture dish, and a reduction in the nucleocytoplasmic ratio due to an increase in cytoplasmic volume [23, 24]. As anticipated and shown in Figures 1(a) and 1(b), MDMM-1 cells exhibited increased cytoplasmic volume compared to that in Mono Mac-1 cells. Moreover, we observed that PMA treatment enhanced the clustering of MDMM-1 cells. Another feature of macrophage differentiation is enhanced granularity, as demonstrated by increased side scatter (SSC) on flow cytometry [25]. Compared to Mono Mac-1 cells, MDMM-1 cells have increased SSC as shown in Figure 1(c). As CD14 expression is a phenotypic marker for monocyte differentiation, CD14 expression on Mono Mac-1 cells was determined after differentiation using PMA. The percentage of CD14+ cells increased from 2.1% to 12.6% during maturation, whereas the mean fluorescence intensity (MFI) was 4.3 and 9.9 in Mono Mac-1 and MDMM-1 cells, respectively (Figure 1(d)). In addition, it has been revealed that the expression of the cell surface antigen CD64 is downregulated in macrophages [26]. Thus, we next investigated whether macrophages exhibit a significant decrease in CD64 expression, and the results indicated that CD64 expression was significantly decreased in MDMM-1 cells (34.6% compared to 68.9% in Mono Mac-1) (Figure 1(d)).
Figure 1.
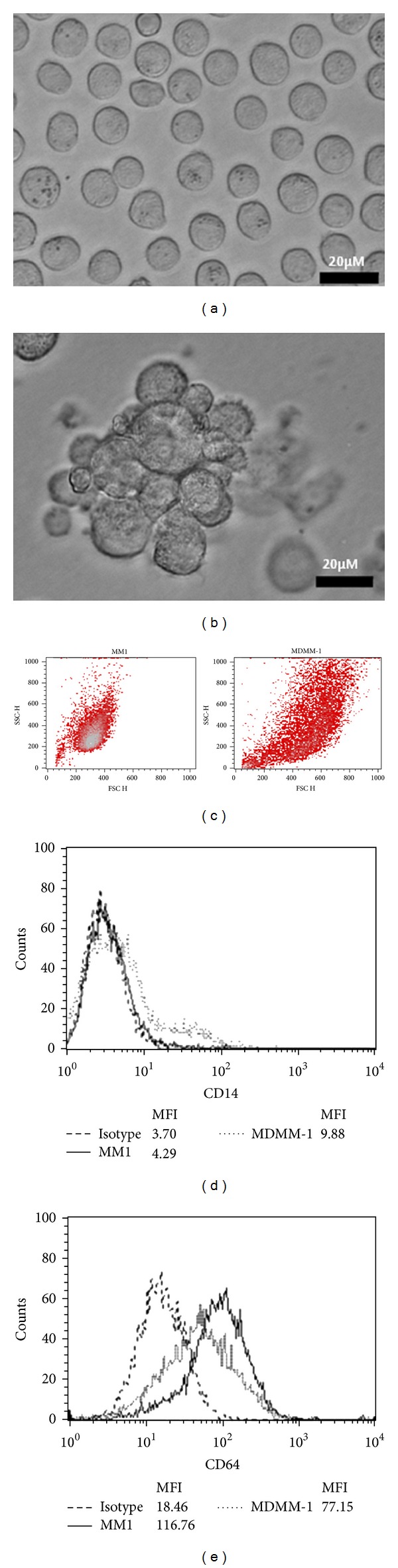
Differentiation of Mono Mac-1 cells into MDMM-1 cells. Representative interferential contrast images of Mono Mac-1 (a) and MDMM-1 (b) cells after 72 h of differentiation with 20 ng/mL PMA, as well as forward light scatter and side light scatter plots (c). CD14 (d) and CD64 (e) expression was analyzed by flow cytometry before and after differentiation. Data shown are representative of three experiments.
We also investigated whether monocytes from the blood of healthy donors that were differentiated into MDMs exhibited the morphological and immunological characteristics of macrophages. As observed with Mono Mac-1 and MDMM-1 cells, morphological differences were noted between monocytes and MDMs (Figures 2(a) and 2(b)), and SSC also increased in macrophages (Figure 2(c)). In human blood cells, monocyte differentiation into macrophages resulted in significantly decreased CD14 expression [27]. We found that macrophage differentiation decreased CD14+ expression (MFI of 25.6 in MDMs; MFI of 138.9 in blood monocytes). Conversely, MDMs exhibited a significant diminution in CD64 expression as expected (MDMs: 4.9% with a MFI of 32.7; blood monocytes: 31.0% with a MFI of 61.1) (Figure 2(d)). Altogether, our results demonstrate that MDMM-1 cells and MDMs exhibit characteristics attributed to macrophages in the literature.
Figure 2.
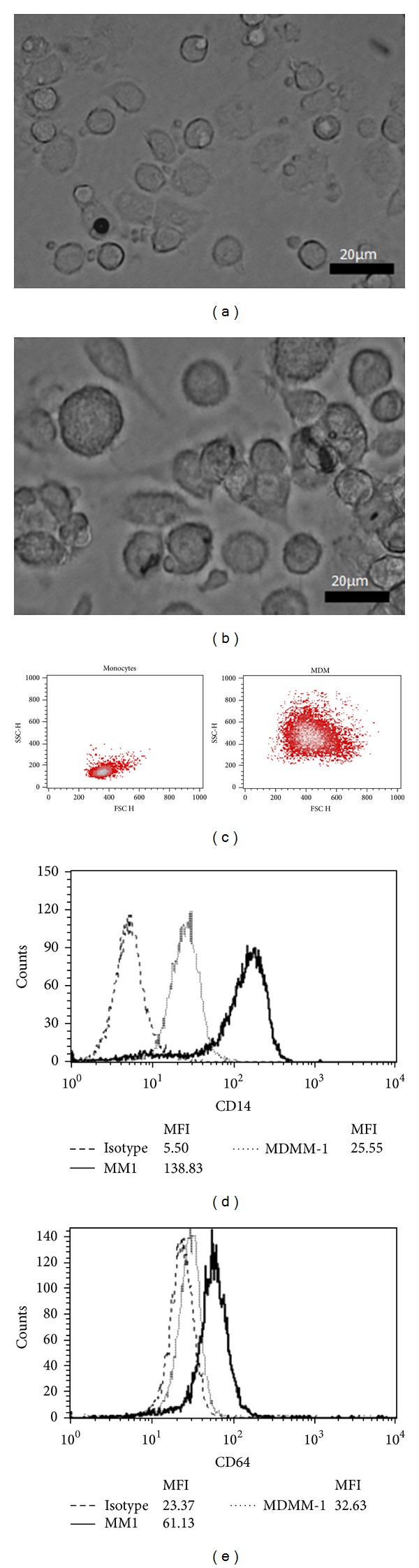
Differentiation of freshly isolated blood monocytes into MDMs. Representative interferential contrast images of blood monocytes (a) and MDMs (b) after 8 days of differentiation in culture, as well as forward light scatter and side light scatter plots (c). CD14 (d) and CD64 (e) expression was analyzed by flow cytometry before and after differentiation. Data shown are representative of three experiments.
To evaluate whether monocyte maturation in the absence of M1/M2 polarizing conditions affects the cell surface expression of CCR7 on Mono Mac-1 and MDMM-1 cells, the cells were incubated in the presence or absence of PGE2 for 24 h, and the expression of CCR7 was analyzed by flow cytometry (Figure 3). Increased cell surface expression of CCR7 has been previously demonstrated on Mono Mac-1 cells by Côté et al [16]. However, an important diminution of CCR7 expression is detected on cell surface of MDMM-1 cells (Mono Mac-1: a MFI of 94.95; MDMM-1: a MFI of 52.23). As it has been demonstrated that PGE2 increases CCR7 expression on Mono Mac-1 cells, we next determined whether PGE2 stimulation also affected CCR7 expression on MDMM-1 cells. After 24 h of treatment, CCR7 expression was significantly increased on MDMM-1 cells (MFI of 52.23). We asked whether CCR7 upregulation was also observed at the mRNA level by using real-time RT-PCR to verify the presence of CCR7 mRNA after Mono Mac-1 differentiation and treatment with PGE2 over a 24 h time course (Figure 4). Our results demonstrated that CCR7 mRNA expression is modulated by PGE2 in MDMM-1 cells. As observed in undifferentiated Mono Mac-1 cells, CCR7 mRNA levels were highest when MDMM-1 cells were stimulated with 1 μM PGE2 for 12 h. Altogether, our results indicate that PMA-induced Mono Mac-1 differentiation decreased the cell surface expression of CCR7, but treatment with the immunomodulatory molecule PGE2 slightly increased the transcription and expression of CCR7 in MDMM-1 cells.
Figure 3.
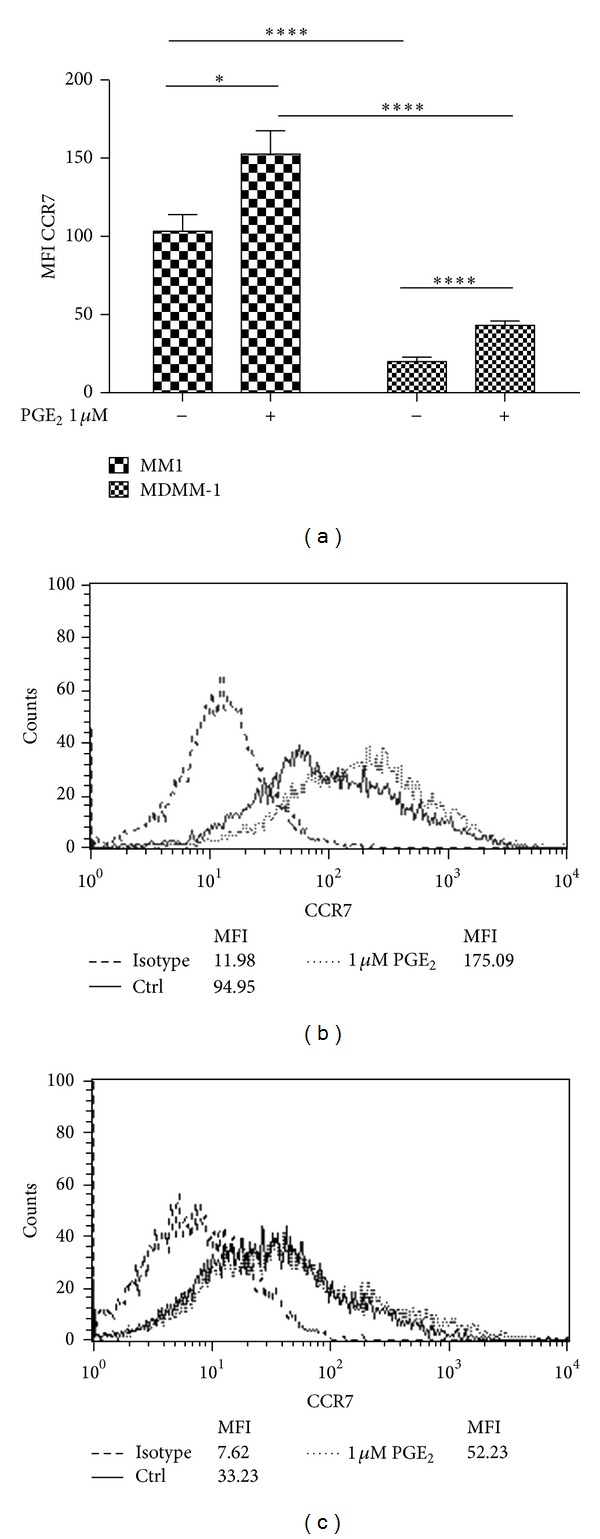
Differentiation of Mono Mac-1 cells into MDMM-1 cells induces the loss of CCR7 expression. Mono Mac-1 cells were differentiated in MDMM-1 cells as described. (a) CCR7 surface expression was evaluated by flow cytometry on Mono Mac-1 and MDMM-1 cells following treatment with 1 μM PGE2 for 24 h. Data are the mean of four representative experiments, and representative histograms obtained for Mono Mac-1 (b) and (c) MDMM-1 cells are presented. *P < 0.5, **P < 0.005, ***P < 0.001, and ****P < 0.0001.
Figure 4.
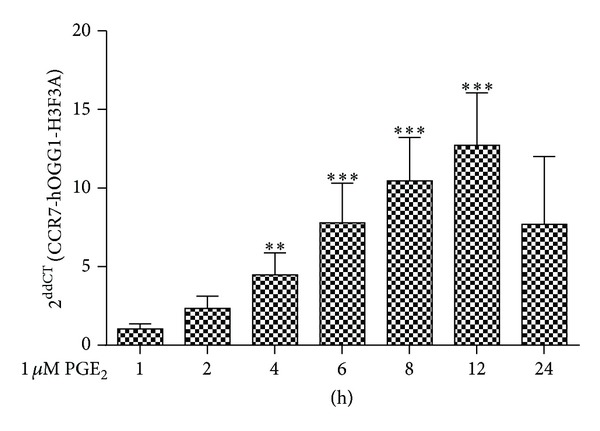
Stimulation with PGE2 induces the upregulation of CCR7 mRNA on MDMM-1 cells. Cells were stimulated with 1 μM PGE2 for the indicated times. Total RNA was extracted and submitted to real-time PCR to detect the presence of CCR7 mRNA. Data represent the means of four different experiments. **P < 0.005, ***P < 0.001.
To further investigate the effects of monocyte maturation on CCR7 expression, we repeated this series of experiments with freshly isolated human blood monocytes. Monocytes were cultured up to 8 days to induce differentiation, and CCR7 mRNA level (Figure 5) as well as CCR7 cell surface expression was analyzed by flow cytometry (Figure 6). As observed in MDMM-1 cells, CCR7 expression was higher on human blood monocytes, and it was downregulated upon the maturation of MDMs (human blood monocytes: 14.0% with a MFI of 31.0; MDMs: 4.9% with a MFI of 16.7). Interestingly, and unlike the effect observed in the differentiated MDMM-1 cell line, no significant change in CCR7 mRNA level and CCR7 expression was detected after PGE2 treatment, although a trend toward lower CCR7 expression was observed. Thus, overall the differentiation of cultured monocytes or freshly isolated monocytes from human blood resulted in a decrease in CCR7 surface expression, and PGE2 slightly restored CCR7 expression in MDMM-1 cells but not MDMs.
Figure 5.
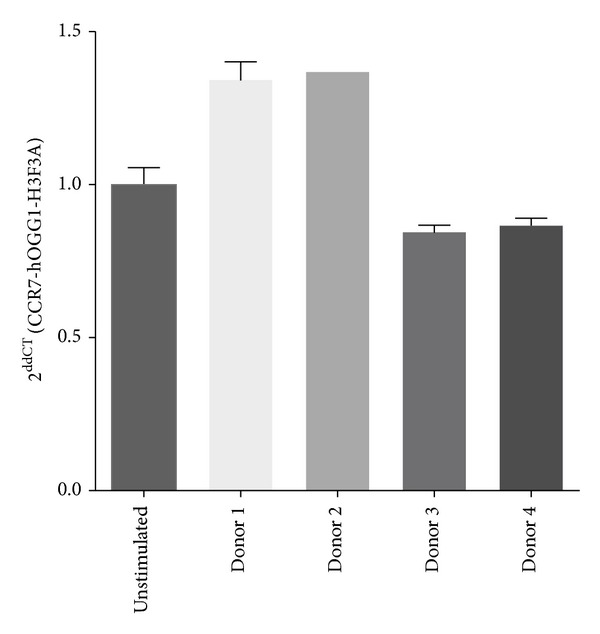
PGE2 does not modulate CCR7 mRNA transcription in MDMs. MDMs were obtained from monocytes from four different healthy donors and were stimulated with 1 μM PGE2 for 6 h. Total RNA was extracted and submitted to real-time PCR to detect the presence of CCR7 mRNA. Results shown are the means (±SD) of four determinations.
Figure 6.
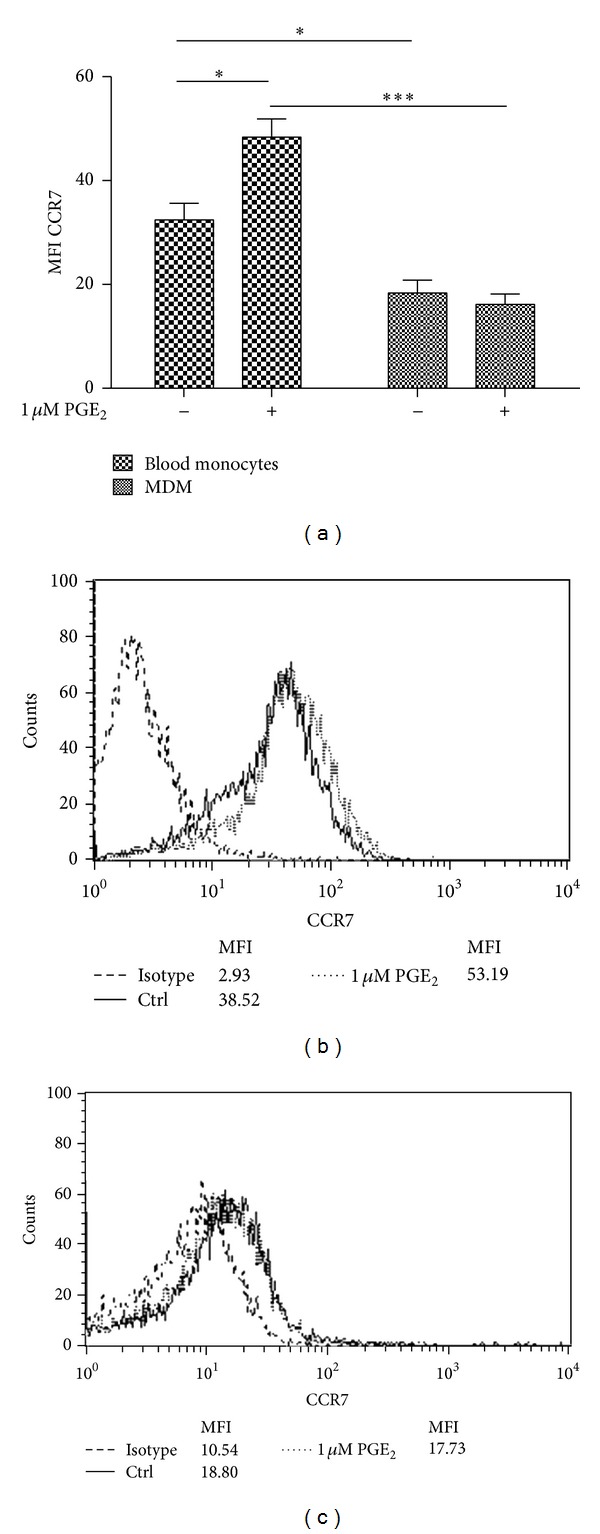
PGE2 has no effect on MDMs CCR7 expression. (a) CCR7 surface expression on blood monocytes and MDMs was evaluated by flow cytometry following treatment with 1 μM PGE2 for 24 h. Data represent the means of five different healthy donors. Histograms from one of five experiments for (b) blood monocytes and (c) MDMs are presented. *P < 0.5, ***P < 0.001.
3.2. MDMs Do Not Migrate in Response to CCR7 Ligands
To confirm the impact of CCR7 functionality on monocyte maturation into macrophages and the effect of PGE2 treatment on this process, we performed chemotaxis assays using these cells and CCR7 ligands (Figure 7). First, Mono Mac-1 and MDMM-1 cells were incubated for 24 h in the presence or absence of PGE2 and evaluated for chemotaxis in response to 300 ng/mL CCL19 or CCL21. The differentiation of Mono Mac-1 cells significantly decreased their cellular responsiveness to both CCR7 ligands (Figure 7(a)); however, the chemotaxis of MDMM-1 cells increased upon treatment with PGE2, which supports the flow cytometry and RT-PCR results. In the case of MDMs, no specific migration was observed compared to that in freshly isolated blood monocytes (Figure 7(b)), confirming the absence of functional CCR7 receptors on the MDMs cell surface. Moreover, PGE2 failed to induce specific migration in MDMs, as was suggested by the flow cytometry results. To confirm the specificity of the observed migration, we incubated cells with a blocking antibody against human CCR7 for 10 min at room temperature prior to the migration assays [16]. This treatment completely abolished cell-specific migration in response to the CCL19 and CCL21 (data not shown).
Figure 7.
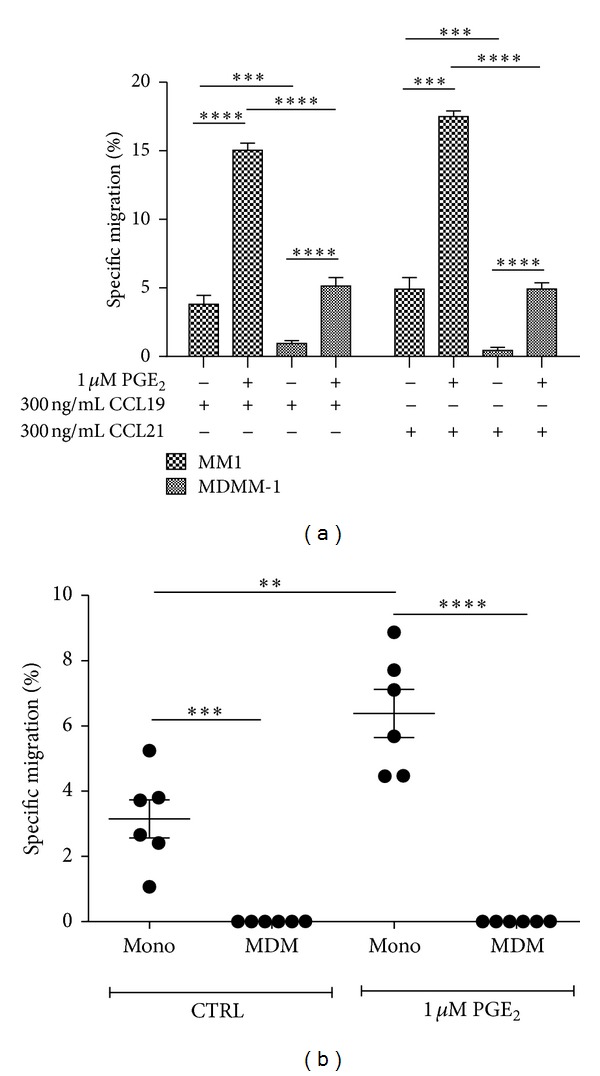
MDMM-1 cells and MDMs do not migrate toward the CCR7-specific ligands CCL19 and CCL21. (a) Chemotaxis assays using 300 ng/mL CCL19 and CCL21 were performed using MM1 and MDMM-1 cells incubated in the presence or absence of 1 μM PGE2 for 24 h. Data represent the means of three different experiments. (b) Blood monocytes from five different healthy donors were differentiated into MDMs and used for chemotaxis assays as described. The mean number of spontaneously migrated cells was subtracted from the number of cells that migrated in response to chemokines. **P < 0.01, ***P < 0.001, and ****P < 0.0001.
4. Conclusion
In response to injury or infection, blood monocytes migrate to tissues to neutralize and eliminate potentially injurious stimuli. Depending on the inflammatory milieu and pathogen-associated pattern recognition receptors, monocytes may differentiate into inflammatory macrophages or DCs [28–30]. Monocytes can first differentiate into immature DCs and then mature upon exposure to antigens while changing their surface expression of chemokine receptors. Indeed, maturing DCs are known to downregulate CCR1, CCR5, and CCR6 expression and upregulate CCR7 expression (see review [31]). As CCR7 expression is not sufficient to ensure the migration of mature DCs in response to CCL19 and CCL21 [19, 32], CCR7 function is dependent on the presence of costimulatory signals such as PGE2 [17, 19]. Alternatively, monocytes can also differentiate into macrophages, long-lived cells that develop specialized functions such as phagocytosis. After phagocytosis, it has been postulated that macrophages will either migrate to the lymphoid organs or die by apoptosis at the inflammatory focus [33, 34]. However, it has recently been demonstrated that inflammatory macrophages, in addition to neutrophils, die via apoptosis during the late stages of resolving a proinflammatory insult [35].
In B cells, CCR9 and CCR10 expression permits the homing of immunoglobulin A plasma cells to mucosal tissues [36, 37], whereas CXCR3 and CXCR4 mediate the migration of IgG+ plasma cells toward inflamed tissues and bone marrow [38–40]. CXCR4 upregulation in T cells is implicated in the migration of leukocytes [41]. However, in bone marrow myeloid cells, CXCR4 downregulation is essential to promote their mobilization from bone marrow to the peripheral blood [42]. In addition, in DCs, PGE2 has been reported to synergize with TNF-α to promote CCR7 expression and the chemotactic responsiveness of DCs to CCL19 and CCL21 in lymph nodes [17–19]. These selected examples illustrate that chemokine receptor expression, together with their natural ligands, is involved and essential in the regulation of cellular migration. Recently, monocytes were shown to express CCR7 in the presence of the proinflammatory molecule PGE2, which is required to ensure migration in response to CCR7 ligands [16]. In the current study, we determined whether monocyte differentiation and the presence of the immunomodulatory molecule PGE2 had an effect on CCR7 expression and CCR7-dependent migration. Our results provide evidence that unpolarized blood macrophages do not express functional CCR7, even in the presence of PGE2 while polarized M1 macrophages have been shown to upregulate CCR7 expression [43]. We used the Mono Mac-1 cell line and freshly isolated blood monocytes that were differentiated into a macrophage-like cell phenotype (Figures 1 and 2). In both cases, we observed downregulation of CCR7 expression (Figures 3 and 6) and CCR7-specific migration (Figure 7). Further, we observed that the addition of PGE2 did not modulate CCR7 mRNA transcription, expression, and functionality in MDMs (Figures 5, 6, and 7). The results obtained using the cell line were different from those obtained using freshly isolated human blood monocytes/macrophages. In MDMM-1 cells, CCR7 expression and functionality were upregulated in response to PGE2. Similar results were found using THP1 cells, a human monocytic cell line. In that study, CCR7 expression in THP1-derived macrophages was increased following activation of the A2A adenosine receptor [44]. The phenotypic differences between cell lines and human blood monocytes/macrophages could be explained by the tumor origins of both Mono Mac-1 and THP1 cells [45].
Our results suggest that human blood macrophages even in the presence of PGE2 do not migrate in response to the CCR7 natural ligands CCL19/CCL21. This reflects in vivo migration patterns because the collection of lymph through cannulation reveals that the major myeloid cell type to enter lymphatic vessels is antigen-presenting DCs rather than classical macrophages [7, 46]. Accordingly, monocyte-derived cells that leave acute inflammatory sites have a DC phenotype [47], and these cells migrate to the T-cell zone of the lymph node [47], which is rich in DCs but not in macrophages [48]. By contrast, little is known regarding macrophage migration. Migration of activated macrophages from the peritoneum to the lymphatics is accelerated by MAC-1, whereas early monocyte accumulation or subsequent redistribution within the peritoneum appears unaffected by this integrin [49].
Our findings are relevant to plaque formation in atherosclerosis because in this chronic inflammatory disease, the migratory process associated with resolution is impaired and macrophages accumulate in plaques, contributing to the build-up of necrotic pools [50]. Indeed, it has been demonstrated that mouse circulating monocyte subsets labeled with fluorescent latex beads did not emigrate out of plaques during disease progression [50, 51]. Restoration of monocyte-derived cell migration out of plaques could be an effective means to stimulate atherosclerosis disease regression. Moreover, some authors postulate that lipid-derived signals such as prostaglandins are strong candidates for impairing migration, and oxidized phospholipids along with lipoprotein A are disease-relevant mediators that may shift the fate of monocyte-derived cells to a more sessile phenotype [50, 52, 53]. Restoration of monocyte-derived cell migration out of plaques could be an effective means to stimulate atherosclerosis disease regression.
In summary, we demonstrated that monocyte differentiation into macrophages directly modifies migratory behavior via CCR7 (Figure 8). Thus, our results suggest that human monocytes express CCR7 as shown with DCs, and T cells can, whereas unpolarized macrophages do not express CCR7 even in the presence of PGE2. Both MDMM-1 and MDM cells lost the functional capacity to migrate in response to the CCR7 ligands CCL19 and CCL21. However, we cannot state conclusively that the migratory behavior of monocytes and macrophages is due solely to CCR7 expression. The implication of other chemokine receptors and their regulation by prostaglandins should be investigated further.
Figure 8.
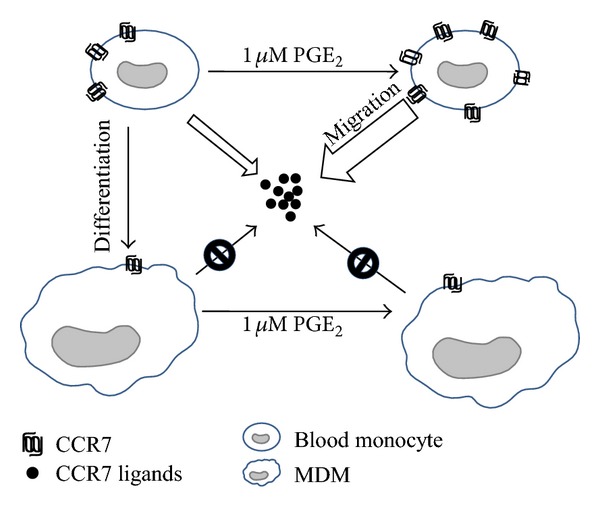
Schematic depicting CCR7 expression and functionality on human monocytes and MDMs.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
Acknowledgments
Marc-André Allaire performed the experiments, prepared the figures, analyzed the data, and drafted the paper. Bérengère Tanné and Sandra C. Côté performed supporting experiments with Mono Mac-1 cells and freshly isolated monocytes. Nancy Dumais conceived the study, participated in its design and coordination, and wrote the paper. All authors read and approved the final paper. This study was supported by the Canadian Institutes of Health Research (CIHR) (Grant no. MOP93677).
References
- 1.Auffray C, Fogg DK, Narni-Mancinelli E, et al. CX3CR1+ CD115+ CD135+ common macrophage/DC precursors and the role of CX3CR1 in their response to inflammation. Journal of Experimental Medicine. 2009;206(3):595–606. doi: 10.1084/jem.20081385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Auffray C, Sieweke MH, Geissmann F. Blood monocytes: development, heterogeneity, and relationship with dendritic cells. Annual Review of Immunology. 2009;27:669–692. doi: 10.1146/annurev.immunol.021908.132557. [DOI] [PubMed] [Google Scholar]
- 3.Liu K, Victora GD, Schwickert TA, et al. In vivo analysis of dendritic cell development and homeostasis. Science. 2009;324(5925):392–397. doi: 10.1126/science.1170540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Förster R, Davalos-Misslitz AC, Rot A. CCR7 and its ligands: balancing immunity and tolerance. Nature Reviews Immunology. 2008;8(5):362–371. doi: 10.1038/nri2297. [DOI] [PubMed] [Google Scholar]
- 5.Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392(6673):245–252. doi: 10.1038/32588. [DOI] [PubMed] [Google Scholar]
- 6.Förster R, Schubel A, Breitfeld D, et al. CCR7 coordinates the primary immune response by establishing functional microenvironments in secondary lymphoid organs. Cell. 1999;99(1):23–33. doi: 10.1016/s0092-8674(00)80059-8. [DOI] [PubMed] [Google Scholar]
- 7.Randolph GJ, Angeli V, Swartz MA. Dendritic-cell trafficking to lymph nodes through lymphatic vessels. Nature Reviews Immunology. 2005;5(8):617–628. doi: 10.1038/nri1670. [DOI] [PubMed] [Google Scholar]
- 8.Gunn MD. Chemokine mediated control of dendritic cell migration and function. Seminars in Immunology. 2003;15(5):271–276. doi: 10.1016/j.smim.2003.08.004. [DOI] [PubMed] [Google Scholar]
- 9.Gunn MD, Tangemann K, Tam C, Cyster JG, Rosen SD, Williams LT. A chemokine expressed in lymphoid high endothelial venules promotes the adhesion and chemotaxis of naive T lymphocytes. Proceedings of the National Academy of Sciences of the United States of America. 1998;95(1):258–263. doi: 10.1073/pnas.95.1.258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Luther SA, Bidgol A, Hargreaves DC, et al. Differing activities of homeostatic chemokines CCL19, CCL21, and CXCL12 in lymphocyte and dendritic cell recruitment and lymphoid neogenesis. Journal of Immunology. 2002;169(1):424–433. doi: 10.4049/jimmunol.169.1.424. [DOI] [PubMed] [Google Scholar]
- 11.Ngo VN, Tang HL, Cyster JG. Epstein-Barr virus-induced molecule 1 ligand chemokine is expressed by dendritic cells in lymphoid tissues and strongly attracts naive T cells and activated B cells. Journal of Experimental Medicine. 1998;188(1):181–191. doi: 10.1084/jem.188.1.181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Stein JV, Rot A, Luo Y, et al. The CC chemokine thymus-derived chemotactic agent 4 (TCA-4, secondary lymphoid tissue chemokine, 6Ckine, exodus-2) triggers lymphocyte function-associated antigen 1-mediated arrest of rolling T lymphocytes in peripheral lymph node high endothelial venules. Journal of Experimental Medicine. 2000;191(1):61–75. doi: 10.1084/jem.191.1.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Warnock RA, Campbell JJ, Dorf ME, Matsuzawa A, McEvoy LM, Butcher EC. The role of chemokines in the microenvironmental control of T versus B cell arrest in Peyer’s patch high endothelial venules. Journal of Experimental Medicine. 2000;191(1):77–88. doi: 10.1084/jem.191.1.77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gunn MD, Kyuwa S, Tam C, et al. Mice lacking expression of secondary lymphoid organ chemokine have defects in lymphocyte homing and dendritic cell localization. Journal of Experimental Medicine. 1999;189(3):451–460. doi: 10.1084/jem.189.3.451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Nakano H, Gunn MD. Gene duplications at the chemokine locus on mouse chromosome 4: multiple strain-specific haplotypes and the deletion of secondary lymphoid-organ chemokine and EBI-1 ligand chemokine genes in the plt mutation. Journal of Immunology. 2001;166(1):361–369. doi: 10.4049/jimmunol.166.1.361. [DOI] [PubMed] [Google Scholar]
- 16.Côté SC, Pasvanis S, Bounou S, Dumais N. CCR7-specific migration to CCL19 and CCL21 is induced by PGE2 stimulation in human monocytes: involvement of EP2/EP4 receptors activation. Molecular Immunology. 2009;46(13):2682–2693. doi: 10.1016/j.molimm.2008.08.269. [DOI] [PubMed] [Google Scholar]
- 17.Scandella E, Men Y, Gillessen S, Förster R, Groettrup M. Prostaglandin E2 is a key factor for CCR7 surface expression and migration of monocyte-derived dendritic cells. Blood. 2002;100(4):1354–1361. doi: 10.1182/blood-2001-11-0017. [DOI] [PubMed] [Google Scholar]
- 18.Scandella E, Men Y, Legler DF, et al. CCL19/CCL21-triggered signal transduction and migration of dendritic cells requires prostaglandin E2. Blood. 2004;103(5):1595–1601. doi: 10.1182/blood-2003-05-1643. [DOI] [PubMed] [Google Scholar]
- 19.Luft T, Jefford M, Luetjens P, et al. Functionally distinct dendritic cell (DC) populations induced by physiologic stimuli: prostaglandin E2 regulates the migratory capacity of specific DC subsets. Blood. 2002;100(4):1362–1372. doi: 10.1182/blood-2001-12-0360. [DOI] [PubMed] [Google Scholar]
- 20.Allaire M, Coté S, Dumais N. Involvement of the MAPK and RhoA/ROCK pathways in PGE2-mediated CCR7-dependent monocyte migration. Immunology Letters. 2012;146(1-2):70–73. doi: 10.1016/j.imlet.2012.05.002. [DOI] [PubMed] [Google Scholar]
- 21.Genois N, Robichaud GA, Tremblay MJ. Mono Mac 1: a new in vitro model system to study HIV-1 infection in human cells of the mononuclear phagocyte series. Journal of Leukocyte Biology. 2000;68(6):854–864. [PubMed] [Google Scholar]
- 22.Ziegler-Heitbrock HWL, Thiel E, Futterer A, Herzog V, Wirtz A, Riethmuller G. Establishment of a human cell line (Mono Mac 6) with characteristics of mature monocytes. International Journal of Cancer. 1988;41(3):456–461. doi: 10.1002/ijc.2910410324. [DOI] [PubMed] [Google Scholar]
- 23.Daigneault M, Preston JA, Marriott HM, Whyte MKB, Dockrell DH. The identification of markers of macrophage differentiation in PMA-stimulated THP-1 cells and monocyte-derived macrophages. PloS ONE. 2010;5(1):p. e8668. doi: 10.1371/journal.pone.0008668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sokol RJ, Hudson G, James NT. Human macrophage development: a morphometric study. Journal of Anatomy. 1987;151:27–35. [PMC free article] [PubMed] [Google Scholar]
- 25.McCullough KC, Basta S, Knötig S, et al. Intermediate stages in monocyte-macrophage differentiation modulate phenotype and susceptibility to virus infection. Immunology. 1999;98(2):203–212. doi: 10.1046/j.1365-2567.1999.00867.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Le Naour R, Clayette P, Henin Y, et al. Infection of human macrophages with an endogenous tumour necrosis factor-α (TNF-α)-independent human immunodeficiency virus type 1 isolate is unresponsive to the TNF-α synthesis inhibitor RP 55778. Journal of General Virology. 1994;75(6):1379–1388. doi: 10.1099/0022-1317-75-6-1379. [DOI] [PubMed] [Google Scholar]
- 27.Ziegler-Heitbrock HWL, Ulevitch RJ. CD14: cell surface receptor and differentiation marker. Immunology Today. 1993;14(3):121–125. doi: 10.1016/0167-5699(93)90212-4. [DOI] [PubMed] [Google Scholar]
- 28.Gordon S, Taylor PR. Monocyte and macrophage heterogeneity. Nature Reviews Immunology. 2005;5(12):953–964. doi: 10.1038/nri1733. [DOI] [PubMed] [Google Scholar]
- 29.van Furth R, Cohn ZA. The origin and kinetics of mononuclear phagocytes. Journal of Experimental Medicine. 1968;128(3):415–435. doi: 10.1084/jem.128.3.415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Van Furth R, Diesselhoff Den Dulk M, Mattie H. Quantitative study on the production and kinetics of mononuclear phagocytes during an acute inflammatory reaction. Journal of Experimental Medicine. 1973;138(6):1314–1330. doi: 10.1084/jem.138.6.1314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Banchereau J, Briere F, Caux C, et al. Immunobiology of dendritic cells. Annual Review of Immunology. 2000;18:767–811. doi: 10.1146/annurev.immunol.18.1.767. [DOI] [PubMed] [Google Scholar]
- 32.Robbiani DF, Finch RA, Jäger D, Muller WA, Sartorelli AC, Randolph GJ. The leukotriene C4 transporter MRP1 regulates CCL19 (MIP-3β, ELC)-dependent mobilization of dendritic cells to lymph nodes. Cell. 2000;103(5):757–768. doi: 10.1016/s0092-8674(00)00179-3. [DOI] [PubMed] [Google Scholar]
- 33.Serhan CN. Resolution phase of inflammation: novel endogenous anti-inflammatory and proresolving lipid mediators and pathways. Annual Review of Immunology. 2007;25:101–137. doi: 10.1146/annurev.immunol.25.022106.141647. [DOI] [PubMed] [Google Scholar]
- 34.Bellingan GJ, Caldwell H, Howie SEM, Dransfield I, Haslett C. In vivo fate of the inflammatory macrophage during the resolution of inflammation: inflammatory macrophages do not die locally, but emigrate to the draining lymph nodes. Journal of Immunology. 1996;157(6):2577–2585. [PubMed] [Google Scholar]
- 35.Kolaczkowska E, Koziol A, Plytycz B, Arnold B. Inflammatory macrophages, and not only neutrophils, die by apoptosis during acute peritonitis. Immunobiology. 2010;215(6):492–504. doi: 10.1016/j.imbio.2009.07.001. [DOI] [PubMed] [Google Scholar]
- 36.Kunkel EJ, Kim CH, Lazarus NH, et al. CCR10 expression is a common feature of circulating and mucosal epithelial tissue IgA Ab-secreting cells. Journal of Clinical Investigation. 2003;111(7):1001–1010. doi: 10.1172/JCI17244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bowman EP, Kuklin NA, Youngman KR, et al. The intestinal chemokine thymus-expressed chemokine (CCL25) attracts IgA antibody-secreting cells. Journal of Experimental Medicine. 2002;195(2):269–275. doi: 10.1084/jem.20010670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hauser AE, Debes GF, Arce S, et al. Chemotactic responsiveness toward ligands for CXCR3 and CXCR4 is regulated on plasma blasts during the time course of a memory immune response. Journal of Immunology. 2002;169(3):1277–1282. doi: 10.4049/jimmunol.169.3.1277. [DOI] [PubMed] [Google Scholar]
- 39.Hauser AE, Muehlinghaus G, Manz RA, et al. Long-lived plasma cells in immunity and inflammation. Annals of the New York Academy of Sciences. 2003;987:266–269. doi: 10.1111/j.1749-6632.2003.tb06059.x. [DOI] [PubMed] [Google Scholar]
- 40.Muehlinghaus G, Cigliano L, Huehn S, et al. Regulation of CXCR3 and CXCR4 expression during terminal differentiation of memory B cells into plasma cells. Blood. 2005;105(10):3965–3971. doi: 10.1182/blood-2004-08-2992. [DOI] [PubMed] [Google Scholar]
- 41.Baggiolini M, Dewald B, Moser B. Human chemokines: an update. Annual Review of Immunology. 1997;15:675–705. doi: 10.1146/annurev.immunol.15.1.675. [DOI] [PubMed] [Google Scholar]
- 42.Hyun KK, De La Luz Sierra M, Williams CK, Gulino AV, Tosato G. G-CSF down-regulation of CXCR4 expression identified as a mechanism for mobilization of myeloid cells. Blood. 2006;108(3):812–820. doi: 10.1182/blood-2005-10-4162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Durafourt BA, Moore CS, Zammit DA, et al. Comparison of polarization properties of human adult microglia and blood-derived macrophages. GLIA. 2012;60(5):717–727. doi: 10.1002/glia.22298. [DOI] [PubMed] [Google Scholar]
- 44.Williams AJ, Cronstein BN. The effect of A2A adenosine receptor activation on C-C chemokine receptor 7 expression in human THP1 macrophages during inflammation. Inflammation. 2012;35(2):614–622. doi: 10.1007/s10753-011-9353-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Mburu YK, Wang J, Wood MA, Walker WH, Ferris RL. CCR7 mediates inflammation-associated tumor progression. Immunologic Research. 2006;36(1–3):61–72. doi: 10.1385/IR:36:1:61. [DOI] [PubMed] [Google Scholar]
- 46.Pugh CW, MacPherson GG, Steer HW. Characterization of nonlymphoid cells derived from rat peripheral lymph. Journal of Experimental Medicine. 1983;157(6):1758–1779. doi: 10.1084/jem.157.6.1758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Randolph GJ, Inaba K, Robbiani DF, Steinman RM, Muller WA. Differentiation of phagocytic monocytes into lymph node dendritic cells in vivo. Immunity. 1999;11(6):753–761. doi: 10.1016/s1074-7613(00)80149-1. [DOI] [PubMed] [Google Scholar]
- 48.Norbury CC, Malide D, Gibbs JS, Bennink JR, Yewdell JW. Visualizing priming of virus-specific CD8+ T cells by infected dendritic cells in vivo. Nature Immunology. 2002;3(3):265–271. doi: 10.1038/ni762. [DOI] [PubMed] [Google Scholar]
- 49.Cao C, Lawrence DA, Strickland DK, Zhang L. Aspecific role of integrin Mac-1 in accelerated macrophage efflux to the lymphatics. Blood. 2005;106(9):3234–3241. doi: 10.1182/blood-2005-03-1288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Randolph GJ. Emigration of monocyte-derived cells to lymph nodes during resolution of inflammation and its failure in atherosclerosis. Current Opinion in Lipidology. 2008;19(5):462–468. doi: 10.1097/MOL.0b013e32830d5f09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Tacke F, Alvarez D, Kaplan TJ, et al. Monocyte subsets differentially employ CCR2, CCR5, and CX3CR1 to accumulate within atherosclerotic plaques. Journal of Clinical Investigation. 2007;117(1):185–194. doi: 10.1172/JCI28549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Angeli V, Llodrá J, Rong JX, et al. Dyslipidemia associated with atherosclerotic disease systemically alters dendritic cell mobilization. Immunity. 2004;21(4):561–574. doi: 10.1016/j.immuni.2004.09.003. [DOI] [PubMed] [Google Scholar]
- 53.Llodrá J, Angeli V, Liu J, Trogan E, Fisher EA, Rendolph GJ. Emigration of monocyte-derived cells from atherosclerotic lesions characterizes regressive, but not progressive, plaques. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(32):11779–11784. doi: 10.1073/pnas.0403259101. [DOI] [PMC free article] [PubMed] [Google Scholar]