

Abstract
The tactoidal state in systems containing long, rod-like molecules consists of partially aligned solute molecules in equilibrium with and at a concentration not much higher than that in the conjugate isotropic phase. Under the liquid lattice model of Flory [Proc. R. Soc. London Ser. A, (1956) 234, 73-89], as well as under other models, tactoid formation by molecules of fixed axial ratio depends on nonideality induced by excluded volumes; the process is wholly entropy driven and requires no direct interactions between rods.
Many rod-like biological polymers exhibit reversible polymerization, so that axial ratio and length are not fixed. Polymerization and rod length will then not only induce nonideality, alignment, and phase separation, but will be affected by these. In this work these interrelations are treated under the model of Flory, modified to include a free energy of polymerization and to permit reversible changes in rod length. The primary conclusion is that, in contrast to the situation for fixed lengths, excluded volume-dependent nonideality alone does not suffice to induce a tactoidal phase separation. In the absence of attractions or repulsions between rods the anisotropic phase is highly concentrated. This phase only becomes tactoidal when a minimal level of repulsive interaction between rods is reached. Under this model, tactoid formation in systems such as deoxygenated hemoglobin S and tobacco mosaic virus depends on repulsive interactions or metastability or both. As a secondary result it is shown that rod length in the anisotropic phase is much greater than in the conjugate isotropic phase.
Keywords: entropy driven, polymer alignment, liquid crystals, length control, hemoglobin S
Full text
PDF
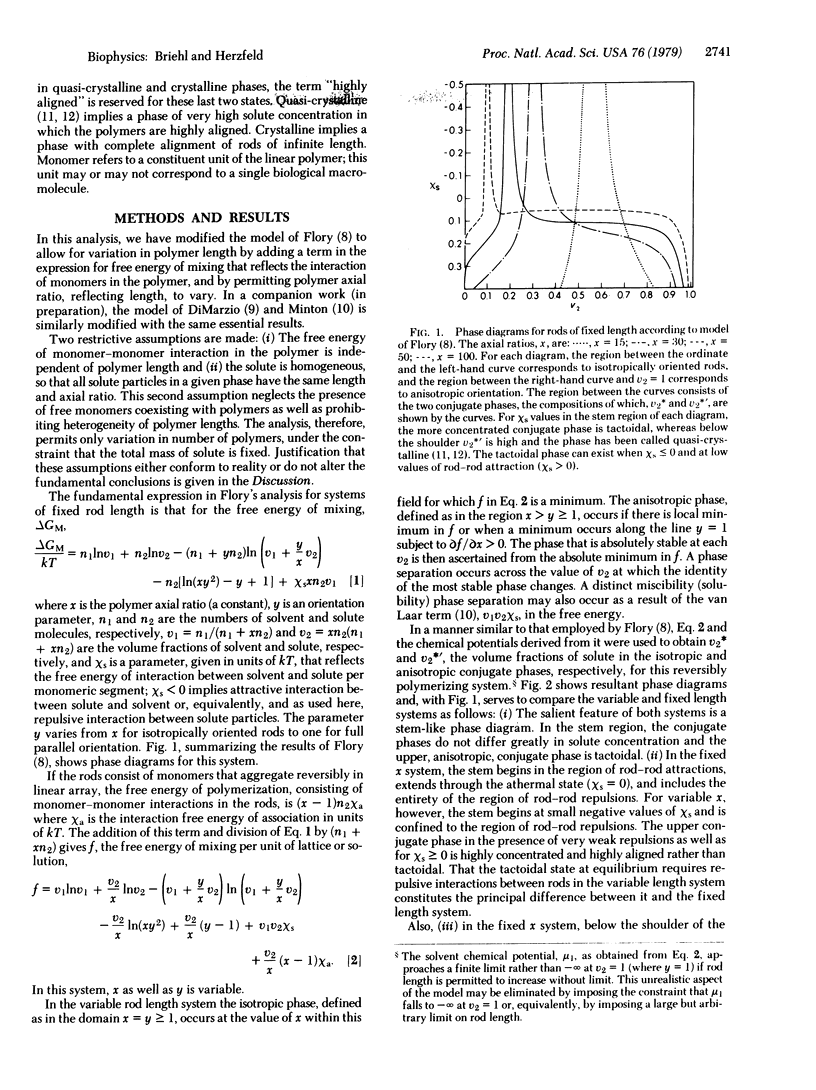


Selected References
These references are in PubMed. This may not be the complete list of references from this article.
- Cantor C. R. Kinetics of primer-dependent polynucleotide phosphorylase synthetic reactions. Biopolymers. 1968;6(3):369–384. doi: 10.1002/bip.1968.360060309. [DOI] [PubMed] [Google Scholar]
- HARRIS J. W. Studies on the destruction of red blood cells. VIII. Molecular orientation in sickle cell hemoglobin solutions. Proc Soc Exp Biol Med. 1950 Oct;75(1):197–201. doi: 10.3181/00379727-75-18144. [DOI] [PubMed] [Google Scholar]
- Minton A. P. Models for the gelling behavior of binary mixtures of hemoglobin variants. J Mol Biol. 1973 Apr 15;75(3):559–574. doi: 10.1016/0022-2836(73)90461-0. [DOI] [PubMed] [Google Scholar]
- Oosawa F. Size distribution of protein polymers. J Theor Biol. 1970 Apr;27(1):69–86. doi: 10.1016/0022-5193(70)90129-3. [DOI] [PubMed] [Google Scholar]
- Pumphrey J. G., Steinhardt J. Crystallization of sickle hemoglobin from gently agitated solutions--an alternative to gelation. J Mol Biol. 1977 May 25;112(3):359–375. doi: 10.1016/s0022-2836(77)80187-3. [DOI] [PubMed] [Google Scholar]
- Pumphrey J. G., Steinhardt J. Formation of needle-like aggregates in stirred solutions of hemoglobin S1. Biochem Biophys Res Commun. 1976 Mar 8;69(1):99–105. doi: 10.1016/s0006-291x(76)80278-1. [DOI] [PubMed] [Google Scholar]
- Scheele R. B., Schuster T. M. Kinetics of protein subunit interactions: simulation of a polymerization overshoot. Biopolymers. 1974;13(2):276–288. doi: 10.1002/bip.1974.360130204. [DOI] [PubMed] [Google Scholar]
- Scheele R. B., Schuster T. M. Letter: Hysteresis of proton binding to tobacco mosaic virus protein associated with metastable polymerization. J Mol Biol. 1975 May 25;94(3):519–525. doi: 10.1016/0022-2836(75)90218-1. [DOI] [PubMed] [Google Scholar]