

Abstract
Painful degenerative disc diseases have been targeted by different biological treatment approaches. Nucleus pulposus (NP) cells play a central role in intervertebral disc (IVD) maintenance by orchestrating catabolic, anabolic and inflammatory factors that affect the extracellular matrix. IVD degeneration is associated with imbalances of these factors, resulting in a catabolic inflammatory metabolism. Therefore, accurate knowledge about their quantity and quality with regard to matrix synthesis is vital for a rational gene therapeutic approach. NP cells were isolated from 63 patients operated due to lumbar disc herniation (mean age 56 / range 29 - 84 years). Then, three-dimensional culture with low-glucose was completed in a collagen type I scaffold for four weeks. Subsequently cell proliferation evaluation was performed using 3-(4, 5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide and intracellular concentration of 28 endogenously expressed anabolic, catabolic, inflammatory factors and relevant matrix proteins was determined by enzyme-linked immunosorbent assay. Specimen-related grades of degeneration were confirmed by preoperative magnetic resonance imaging. Independent from gender, age and grade of degeneration proliferation rates remained similar in all groups of NP cells. Progressive grades of degeneration, however, showed a significant influence on accumulation of selective groups of factors such as disintegrin and metalloproteinase with thrombospondin motifs 4 and 5, matrix metalloproteinase 3, metalloproteinase inhibitor 1 and 2, interleukin-1β and interleukin-1 receptor. Along with these changes, the key NP matrix proteins aggrecan and collagen II decreased significantly. The concentration of anabolic factors bone morphogenetic proteins 2, 4, 6 and 7, insulin-like growth factor 1, transforming growth factor beta 1 and 3, however, remained below the minimal detectable quantities. These findings indicate that progressive degenerative changes in NP may be problematic with regard to biologic treatment strategies. Hence, gene therapeutic interventions regulating relevant bioactive factors identified in this work might contribute to the development of regenerative treatment approaches for degenerative disc diseases.
Introduction
Painful degenerative disc diseases, frequently caused by genetic predisposition, have been targeted by different biological treatment approaches [1,2]. These include the administration of growth factors, the application of autologous or allogenic cells, gene therapy, in situ therapy and the introduction of biomaterials or a combination thereof.
Nucleus pulposus (NP) cells play a central role in intervertebral disc (IVD) matrix maintenance by orchestrating several catabolic, anabolic and inflammatory factors that affect the extracellular matrix [3,4]. IVD degeneration is associated with imbalances of these factors, resulting in a catabolic inflammatory metabolism. Consequently, accurate knowledge about their quantity as well as their quality with regard to matrix synthesis is vital for a rational gene therapeutic approach. In the current experimental literature, the number of cells and the concentration of gene therapeutic factors applied for regeneration of NP in animal models fluctuate in part enormously, which indicates a deficit in data about cell proliferation rates and concentrations of target proteins in NP cells [5-11].
With this work, we intended a screening for the identification of potential target proteins for gene therapeutic approaches in a tissue engineering setting with human nucleus pulposus cells in a three-dimensional collagen type I scaffold. Therefore, we analyzed cell proliferation and intracellular protein concentration of 28 endogenously expressed anabolic, catabolic and inflammatory factors as well as matrix proteins in relation with age, gender and grade of degeneration by using 3-(4, 5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide (MTT) assay and enzyme-linked immunosorbent assay (ELISA) respectively. Specimen-related grades of degeneration were confirmed by preoperative magnetic resonance imaging.
Previous data on expressions profiles of NP cells were determined either qualitatively by histological and immunohistological methods or quantitatively by RNA techniques [12-17].
The advantage of the relative mass value evaluation method is the ability to perform precise quantifications of protein concentrations. To our knowledge this method has not been applied to analyze a broad array of endogenously expressed bioactive factors in NP cells from a larger number of specimens.
Here, we determined the relative mass value of 28 endogenously expressed target protein in NP cells isolated from 63 tissue specimens and analysed the unfavorable phenotypic alternations that might restrain NP matrix regeneration. Progressive grades of degeneration, showed a significant influence on accumulation of selective bioactive factors. The results of this study might contribute to the development of regenerative treatment approaches for degenerative disc diseases.
Materials and Methods
Tissue samples
Experimental studies of human spinal disc specimens were approved by the local research ethics committee (Heidelberg University, University Medical Center Mannheim: project 2009-217N-MA). Human NP tissues were obtained during surgery with informed consents of the patients. Participants provided their written informed consent to participate in this study. Scoring of spinal disc degeneration was based on magnetic resonance imaging (MRI) findings according to the Pfirrmann scoring system [18]. Representative images of disc degeneration grades III, IV, and V with lumbar disc herniation are shown in Figure 1. The study involved 63 lumbar discs of degeneration grade III, IV and V with a mean age of 56 years (range 29 - 84 years, 27 women and 36 men). More information is presented in table 1. Patient NP specimens were recruited from NP compartment during surgical procedure conducted on individuals with intervertebral disc herniation. Inclusion criteria for surgery were radiographically determined intervertebral disc herniations with nerve root compression on MRI, which correlated to primary symptoms that remained unresponsive to non-operative treatment for six weeks or demonstrated progressive neurological deterioration in the face of conservative treatment. NP specimens were immediately brought to the lab in sterile phosphate buffered saline solution (PBS) (Sigma-Aldrich) for instant cell isolation.
Figure 1. MRI showing representative images of disc degeneration grades (DDG) III, IV and V with lumbar disc herniation.
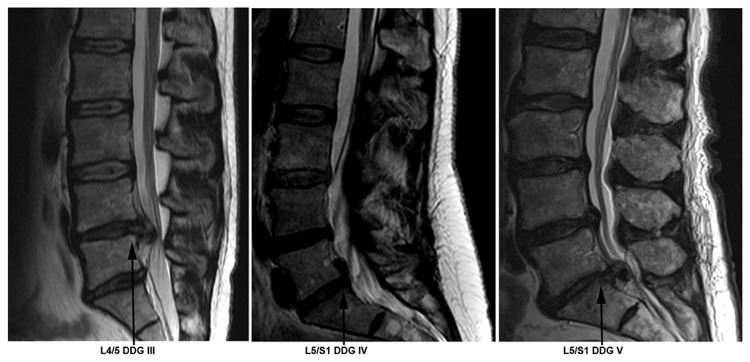
Table 1. Details of NP samples with disc level, disc degeneration grade (DDG), age and gender.
| Sample | Disc level | DDG | Year/Sex | Sample | Disc level | DDG | Year/Sex | |
|---|---|---|---|---|---|---|---|---|
| 1 | L4/5 | III | 29/F | 33 | L5/S1 | IV | 47/M | |
| 2 | L4/5 | III | 30/M | 34 | L3/4 | IV | 49/F | |
| 3 | L5/S1 | III | 32/M | 35 | L4/5 | IV | 57/M | |
| 4 | L4/5 | III | 33/F | 36 | L4/5 | IV | 58/M | |
| 5 | L4/5 | III | 36/F | 37 | L5/S1 | IV | 59/M | |
| 6 | L5/S1 | III | 37/M | 38 | L3/4 | IV | 61/F | |
| 7 | L5/S1 | III | 39/F | 39 | L4/5 | IV | 67/F | |
| 8 | L5/S1 | III | 39/F | 40 | L4/5 | IV | 71/M | |
| 9 | L4/5 | III | 42/M | 41 | L4/5 | IV | 73/F | |
| 10 | L4/5 | III | 44/F | 42 | L4/5 | V | 61/F | |
| 11 | L5/S1 | III | 44/F | 43 | L4/5 | V | 62/M | |
| 12 | L5/S1 | III | 45/M | 44 | L4/5 | V | 63/F | |
| 13 | L4/5 | III | 47/M | 45 | L4/5 | V | 65/M | |
| 14 | L4/5 | III | 47/M | 46 | L5/S1 | V | 66/F | |
| 15 | L5/S1 | III | 49/M | 47 | L4/5 | V | 66/M | |
| 16 | L5/S1 | III | 50/F | 48 | L3/4 | V | 67/M | |
| 17 | L5/S1 | III | 51/M | 49 | L4/5 | V | 67/M | |
| 18 | L5/S1 | III | 52/M | 50 | L5/S1 | V | 67/M | |
| 19 | L5/S1 | III | 52/F | 51 | L5/S1 | V | 67/M | |
| 20 | L5/S1 | III | 54/M | 52 | L5/S1 | V | 67/F | |
| 21 | L4/5 | III | 54/M | 53 | L4/5 | V | 69/F | |
| 22 | L3/4 | III | 56/M | 54 | L5/S1 | V | 69/F | |
| 23 | L5/S1 | III | 59/M | 55 | L5/S1 | V | 71/M | |
| 24 | L4/5 | III | 62/M | 56 | L4/5 | V | 72/F | |
| 25 | L2/3 | III | 67/M | 57 | L2/3 | V | 72/F | |
| 26 | L4/5 | III | 76/M | 58 | L3/4 | V | 74/M | |
| 27 | L5/S1 | IV | 32/F | 59 | L5/S1 | V | 79/M | |
| 28 | L5/S1 | IV | 41/M | 60 | L4/5 | V | 79/F | |
| 29 | L5/S1 | IV | 43/M | 61 | L5/S1 | V | 79/F | |
| 30 | L3/4 | IV | 44/M | 62 | L5/S1 | V | 84/F | |
| 31 | L5/S1 | IV | 44/F | 63 | L4/5 | V | 84/F | |
| 32 | L 4/5 | IV | 44/M |
Samples from NP compartment were obtained from patients undergoing surgery due to spinal disc herniation.
Isolation and monolayer expansion of NP cells
IVD tissues were washed twice in PBS (1000 x g, 2 min) and NP tissues were carefully separated from AF (anulus fibrosus) on the basis of their macroscopic morphology (identification of the innermost lamellar ring of the AF) and finely minced into small fragments of approximately 2 mm3. Samples were directly digested with 0.02 % w/v pronase (Sigma-Aldrich) (1 h, 37 °C, 5 % CO2) in 20 ml DMEM (Dulbecco's Modified Eagle's Medium) containing 1 % penicillin/streptomycin, 1 % glucose and 1 % FCS (fetal calf serum) (Sigma-Aldrich). Samples were filtered through sterile 75 gm nylon mesh filters (Sigma-Aldrich) and supernatants were centrifuged (1000 x g, 2 min). Pellets were resuspended again in 20 ml DMEM and digested with 0.02 % w/v collagenase II (Sigma-Aldrich) and 100U hyaluronidase (Sigma-Aldrich) (3 h, 37 °C, 5 % CO2). Samples were filtered through sterile 75 gm nylon mesh filters and supernatants were centrifuged (1000 x g, 2 min). Pellets were resuspended in 10 ml DMEM containing 1 % penicillin/streptomycin, 1 % glucose and 10 % FCS and cultured in 75 cm2 tissue culture flask (Sigma-Aldrich) (2 weeks, 37 °C, 5 % CO2) by changing the culture medium every two days. Cells were cryopreserved at -196 °C in culture medium containing 30 % FCS and 15 % dimethyl sulfoxide (DMSO) (Sigma-Aldrich).
Three dimensional NP cell culture in collagen I scaffold
The scaffold collagen cell carrier (CCC) (Viscofan Bioengineering) was used for three dimensional (3D) cell cultures. Each well of the 6-well plates was loaded with 1.5 ml PBS (pH 7.3 without Ca2+ / Mg2+) to attach the 6-well format scaffold to the bottoms. Each scaffold was placed on to the PBS and incubated at room temperature for 20 min. After removing the remaining PBS, the plates were left in the operating laminar flow hood overnight. Prior to cell seeding, the scaffold was equilibrated by incubation with 1.5 ml pre-warmed culture medium (10 min, 37 °C, 5 % CO2). The medium was removed and an aliquot (4 x 105 cells/1.5 ml) of the cells was seeded onto the scaffold. Cells were cultured (37 °C, 5 % CO2) by changing the culture medium every two days and harvested after four weeks for analysis of cell proliferation and protein expression.
Isolation of NP cells from collagen I scaffold
Following four weeks of 3D cell culture, the scaffold was digested in 1 ml culture medium with 0.02 % w/v collagenase II (1 h, 37 °C, 5 % CO2). Samples were filtered through sterile 75 gm nylon mesh filters and supernatants were centrifuged (1000 x g, 2 min). Pellets were washed twice in 5 ml PBS (1000 x g, 2 min) and processed for assays of cell proliferation and protein expression.
NP cell proliferation assay
After culturing of 4 x 105 cells in the scaffold for four weeks, cell proliferation assay was determined by using 3-(4, 5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide (MTT) (Molecular Probes). NP cells from the scaffold were suspended in 0.5 ml culture medium and duplicates of 100 µl were plated into flat-bottomed 96 well plates. Duplicate control wells of medium alone were added to provide the blanks for absorbance readings. Cells were incubated (24 h, 37 °C, 5 % CO2) to recover from handling. After adding 10 µl MTT Reagent to each well, cells were again incubated (3 h, 37 °C, 5 % CO2) and 100 µl of the SDS-HCl solution was added for further incubation (4 h, 37 °C, 5 % CO2). The absorbance in each well was measured at 570 nm in a microtiter plate reader Infinite 200 (TECAN). The average value of the blank duplicate readings was subtracted from the average values of the sample duplicate readings and cell concentration was calculated from the standard curve.
Isolation and quantification of protein from NP cells
Protein isolation was performed with NP cells harvested from the scaffold using radio-immunoprecipitation assay (RIPA) buffer (Sigma-Aldrich). Cell pellets were washed twice in cold PBS (2,500 x g, 5 min), resuspended with 300 ml cold RIPA buffer containing protease inhibitor and phosphatase inhibitor cocktails (Sigma-Aldrich) and sonicated (30 sec, 50 % pulse). The Mixture was shacked gently on ice (15 min) and centrifuged (14,000 x g, 4 °C and 15 min). Supernatants were transferred to new tubes for protein quantification. Determination of protein concentration in samples was performed according to the instruction manual (Pierce Micro BCA Protein Assay Protocol) (Thermo Scientific).
Enzyme-linked immunosorbent assay
Enzyme-linked immunosorbent assay (ELISA) was applied to determine the presence and concentration of target proteins in 100 µg of total protein extracts of NP cells. For measuring the concentration of target proteins ELISA kits were purchased from kits producers R & D Systems (United Kingdom) or Uscn Life Science Inc. (USA). Assays were performed according to the instruction manuals of kits producers. The investigated 28 target proteins include: catabolic factors ADAMTS-4, ADAMTS-5 (a disintegrin and metalloproteinase with thrombospondin motifs), MMP-1, MMP-2, MMP-3, MMP-7, MMP-8, MMP-9, MMP-10 and MMP-13 (matrix metalloproteinase), anti-catabolic factors TIMP-1, TIMP-2, TIMP-3 and TIMP-4 (metalloproteinase inhibitor), inflammatory cytokines IL-1β (interleukin-1β), IL-1 R1 (interleukin-1 receptor), TNF- α (tumornekrosefaktor-α) and TNF-R1(tumornekrosefaktor receptor R1); anabolic factors BMP-2, BMP-4, BMP-6, BMP-7 (bone morphogenetic proteins), IGF-1 (insulin-like growth factor 1), TGF-β1 and TGF-β3 (transforming growth factor betas) as well as matrix proteins aggrecan, collagen I and collagen II.
Statistical data analysis
The reliability of the MRI evaluations was predicted using agreement percentage and kappa statistics between two raters (interobserver reliability), and frequency of disagreement was calculated for each degeneration grade [18,19]. Statistical analysis was performed using GraphPad Prism version 6.0b for Mac OS X, GraphPad Software, La Jolla California USA, www.graphpad.com. For cell proliferation and protein expression levels 1-way ANOVA with Brown-Forsythe test and multiple comparisons were applied. Cell proliferation rates and concentration of target proteins were analyzed separately for each experiment. Correlation analysis was performed using the software IBM SPSS Statistics 20, Armonk New York USA to calculate the significance of the changes in cell proliferation and protein concentration as a function of degeneration grade, age and gender. Significance in all cases was set at P < 0.05.
Using the correspondence address all raw data are available at the Department of Neurosurgery, Innsbruck Medical University, Anichstrasse 35, A-6020 Innsbruck, Austria.
Results
Interobserver reliability
Interobserver reliability agreement for rating intervertebral disc degeneration with two observers was rated with a reliability coefficient kappa of 0.9031 and the calculated frequency of disagreements for Pfirrmann score of degeneration grade III, IV and V were 9.68 %, 15.00 % and 0.00 % respectively
Proliferation rates of degenerative NP cells
Three dimensional culture of 4 x 105 adult NP cell in collagen I scaffold for four weeks resulted in similar proliferation rates for all tested samples of degeneration grade III, IV and V with mean values of 1,081 x 106 (+ 19182), 1,083 x 106 (+ 13121) and 1,056 x 106 (+ 13857) cells respectively (P = 0.0014). Only about 2 % growth rate difference was confirmed between degeneration grade III - V (table 2 and Figure 2a). Neither age nor gender appears to play a role in influencing cell proliferation potential in adult NP (data not shown).
Table 2. Proliferation rate of degenerative NP cells in collagen I scaffold.
| DDG | Minimum | Maximum | Range % | Mean | SD |
Mean difference
|
Mean fold | P value | |
|---|---|---|---|---|---|---|---|---|---|
| III | 1048696 | 1116433 | 6.06 | 1081847 | 19182 | III - V | 21912 | 1,020 | 0.0014 |
| IV | 1054062 | 1099726 | 4.15 | 1083120 | 13121 | III - IV | -1273 | 0,999 | 0.0014 |
| V | 1043674 | 1094825 | 4.67 | 1059934 | 13857 | IV- V | 23185 | 1,021 | 0.0014 |
Sixty three samples of degenerative IVD tissues were obtained from patients undergoing surgery due to spinal disc herniation. 4 x 105 cells from each sample were grown for four weeks in collagen I scaffold. Cell proliferation data (MTT assay) were statistically analyzed on the bases of disc degeneration grade (DDG).
Figure 2. Proliferation rate of adult NP cells and expression levels of ADAMTS-4 and -5.

NP cells were isolated from 63 samples of degenerative IVDs and 4 x 105 cells from each sample were grown for four weeks in collagen I scaffold. Cell proliferation data (MTT assay) and data of ADAMTS-4 and -5 protein concentration (ELISA) from 100 µg total protein extracts were statistically analyzed on the bases of disc degeneration grade (DDG). Box plots with whiskers min. to max. show NP cell proliferation rates (Figure 2a), ADAMTS-4 protein expression levels (Figure 2b) and ADAMTS-5 protein expression levels (Figure 2c).
Levels of catabolic, anti-catabolic and inflammatory proteins in degenerative NP cells
Three dimensional culture of 4 x 105 NP cell in collagen I scaffold for four weeks demonstrated high and increasing expression levels of the catabolic factor ADAMTS-4 with mean expression values of 1697 + 169 pg/ml, 3196 + 176 pg/ml and 5884 + 175 pg/ml for degeneration grades III, IV and V respectively (P < 0.0001). It shows about 3.5 fold increased mean expression value between degeneration grades III - V. Also high but similar expression levels of ADAMTS-5 with mean expression values of 3952 + 196 pg/ml, 4150 + 141 pg/ml and 4097 + 192 pg/ml (P < 0.0025) were recorded (table 3 and Figure 2b-c).
Table 3. Expression levels of catabolic proteins in adult NP cells.
| Catabolic proteins | DDG | Min. pg/ml | Max. pg/ml | Range % | Mean pg/ml | SD |
Mean difference
|
Mean fold | P value | |
|---|---|---|---|---|---|---|---|---|---|---|
| DDG | pg/ml | |||||||||
| ADAMTS-4 | III | 1556 | 2283 | 31.84 | 1697 | 169 | V - III | 4186 | 3.466 | < 0.0001 |
| IV | 2735 | 3366 | 18.74 | 3196 | 176 | V - IV | 2687 | 1.840 | < 0.0001 | |
| V | 5407 | 6203 | 12.83 | 5884 | 175 | IV- III | 1499 | 1.883 | < 0.0001 | |
| ADAMTS-5 | III | 3600 | 4337 | 16.99 | 3952 | 196 | V - III | 145 | 1.036 | < 0.0025 |
| IV | 3914 | 4330 | 9.6 | 4150 | 141 | V - IV | -52 | 0.987 | < 0.0025 | |
| V | 3786 | 4390 | 13.75 | 4097 | 192 | IV- III | 197 | 1.050 | < 0.0025 | |
| MMP-1 | III | 96 | 133 | 27.81 | 112 | 11.95 | V - III | 176 | 2.567 | < 0.0001 |
| IV | 243 | 279 | 12.90 | 260 | 10.25 | V - IV | 29 | 1.111 | < 0.0001 | |
| V | 268 | 319 | 15.98 | 289 | 15.67 | IV- III | 147 | 2.309 | < 0.0001 | |
| MMP-2 | III | 62 | 80 | 22.50 | 70 | 6.142 | V - III | 3.129 | 1.044 | 0.3295 |
| IV | 61 | 79 | 22.78 | 70 | 7.281 | V - IV | 3.271 | 1.046 | 0.3295 | |
| V | 64 | 81 | 20.98 | 73 | 5.483 | IV- III | -0.142 | 0.997 | 0.3295 | |
| MMP-3 | III | 8453 | 8748 | 3.37 | 8700 | 54,52 | V - III | 63 | 1.007 | 0.001 |
| IV | 8678 | 8981 | 3.37 | 8835 | 93.39 | V - IV | -72 | 0.991 | 0.001 | |
| V | 8443 | 8939 | 5.54 | 8763 | 153.4 | IV- III | 135 | 1.015 | 0.001 | |
| MMP-7 | III | 335 | 352 | 4.82 | 343 | 4.502 | V - III | -119 | 0.651 | < 0.0001 |
| IV | 259 | 276 | 6.15 | 268 | 4.694 | V - IV | -45 | 0.831 | < 0.0001 | |
| V | 212 | 238 | 10.92 | 223 | 6.460 | IV- III | -74 | 0.782 | < 0.0001 | |
| MMP-13 | III | 430 | 462 | 6.92 | 436 | 7.167 | V - III | 6.564 | 1.015 | 0.0233 |
| IV | 434 | 460 | 5.65 | 442 | 6.472 | V - IV | 1.348 | 1.003 | 0.0233 | |
| V | 432 | 482 | 10.37 | 443 | 10.66 | IV- III | 5.215 | 1.011 | 0.0233 | |
MMP-8, MMP-9 and MMP-10 expression levels were below the minimum detectable dose (MDD) of our detection system. MDD of MMP-8 < 20 pg/ml, MMP-9 < 156 pg/ml and MMP-10 < 4 pg/ml.
Sixty three samples of degenerative IVD tissues were obtained from patients undergoing surgery due to spinal disc herniation. 4 x 105 cells from each sample were grown for four weeks in collagen I scaffold. The protein concentration data (ELISA) of catabolic factors were statistically analyzed from 100 µg total protein extracts on the bases of disc degeneration grade (DDG).
Among all tested catabolic factors MMP-3 was expressed at highest levels in degeneration grade III, IV and V with similar mean expression values of 8700 + 54.52 pg/ml, 8835 + 93.39 pg/ml and 8763 + 153 pg/ml (P = 0.001) respectively. In comparison with MMP-3 expression, extremely lower and constant expression levels of MMP-13 with mean expression values 436 + 7.167 pg/ml, 442 + 6.472 pg/ml and 443 + 10.66 pg/m (P = 0.0233), and decreasing expression levels of MMP-7 with mean expression values 343 + 4.502 pg/ml, 268 + 4.694 pg/ml and 223 + 6.460 pg/m (P < 0.0001), as well as increasing expression levels of MMP-1 with mean expression values 112 + 11.95 pg/ml, 260 + 10.25 pg/ml and 289 + 15.65 pg/ml (P < 0.0001) and equivalent expression levels of MMP-2 with mean expression values less than 74 pg/ml were measured (table 3 and Figure 3). The expression levels of MMP-8, MMP-9 and MMP-10 were below the minimum detectable dose (MDD) of our detection system (MDD of MMP-8 < 20 pg/ml, MMP-9 < 156 pg/ml and MMP-10 < 4 pg/ml).
Figure 3. Expression levels of catabolic proteins MMPs in degenerative NP cells.

NP cells were isolated from 63 samples of degenerative IVDs and 4 x 105 cells from each sample were cultured for four weeks in collagen I scaffold. Concentration data (ELISA) of MMPs from 100 µg total protein extracts were statistically analyzed on the bases of disc degeneration grade (DDG). Box plots with whiskers min. to max. show MMP-1 expression levels (Figure 3a), MMP-2 expression levels (Figure 3b), MMP-3 expression levels (Figure 3c), MMP-7 expression levels (Figure 3d) and MMP-13 expression levels (Figure 3e).
The high expression levels of MMP-3 were counteracted by higher and increasing expression levels of TIMP-1 and TIMP-2 with mean expression values of 14189 + 348 pg/ml, 17095 + 336 pg/ml and 18578 + 322 pg/ml for TIMP-1 (P < 0.0001) and 10966 + 248 pg/ml, 13640 + 156 pg/ml and 16757 + 264 pg/ml (P < 0.0001) for TIMP-2. In contrast, considerably lower but slightly increasing TIMP-3 expression levels were recorded with mean expression values of 843 + 67 pg/ml, 939 + 45 pg/ml and 973 + 46 pg/ml (P < 0.0001). TIMP-4 had the lowermost expression levels with mean expression values of 176 + 7.072 pg/ml, 192 + 9.963 pg/ml and 215 + 6.167 pg/ml (P < 0.0001) respectively (table 4 and Figure 4).
Table 4. Expression levels of anti-catabolic and inflammatory proteins in adult NP cells.
| Anti-catabolic proteins | DDG | Min. pg/ml | Max. pg/ml | Range % | Mean pg/ml | SD |
Mean difference
|
Mean fold | P value | |
|---|---|---|---|---|---|---|---|---|---|---|
| DDG | pg/ml | |||||||||
| TIMP-1 | III | 13621 | 15187 | 10.31 | 14189 | 348 | V - III | 4388 | 1.309 | < 0.0001 |
| IV | 16531 | 17648 | 6.32 | 17095 | 336 | V - IV | 1482 | 1.086 | < 0.0001 | |
| V | 18036 | 18941 | 4.77 | 18578 | 322 | IV- III | 2906 | 1.204 | < 0.0001 | |
| TIMP-2 | III | 10399 | 11356 | 8.42 | 10966 | 248 | V - III | 5790 | 1.528 | < 0.0001 |
| IV | 13290 | 13839 | 3.96 | 13640 | 156 | V - IV | 3117 | 1.228 | < 0.0001 | |
| V | 16103 | 17042 | 5.50 | 16757 | 264 | IV- III | 2673 | 1.243 | < 0.0001 | |
| TIMP-3 | III | 765 | 936 | 18.26 | 843 | 67 | V - III | 129 | 1.153 | < 0.0001 |
| IV | 879 | 1000 | 12.10 | 939 | 45 | V - IV | 34 | 1.036 | < 0.0001 | |
| V | 902 | 1028 | 12.25 | 973 | 46 | IV- III | 95 | 1.113 | < 0.0001 | |
| TIMP-4 | III | 167 | 189 | 15.22 | 176 | 7.072 | V - III | 39 | 1.223 | < 0.0001 |
| IV | 174 | 208 | 24.34 | 192 | 9.963 | V - IV | 23 | 1.122 | < 0.0001 | |
| V | 201 | 226 | 11.06 | 215 | 6.167 | IV- III | 15 | 1.089 | < 0.0001 | |
| Inflammatory proteins | DDG | Min. pg/ml | Max. pg/ml | Range % | Mean pg/ml | SD |
Mean difference
|
Mean fold | P value | |
| DDG | pg/ml | |||||||||
| IL-1β | III | 101 | 113 | 10,61 | 110 | 3.770 | V - III | 29.362 | 1.264 | < 0.0001 |
| IV | 112 | 127 | 11.81 | 121 | 6.326 | V - IV | 18.315 | 1.150 | < 0.0001 | |
| V | 132 | 143 | 7.69 | 140 | 2.895 | IV- III | 11.047 | 1.099 | < 0.0001 | |
| IL-1 R | III | 119 | 140 | 15.00 | 128 | 5.944 | V - III | 36.998 | 1.288 | < 0.0001 |
| IV | 133 | 158 | 15.85 | 146 | 8.229 | V - IV | 18.670 | 1.127 | < 0.0001 | |
| V | 150 | 176 | 14.77 | 165 | 7.827 | IV- III | 18.328 | 1.142 | < 0.0001 | |
| TNF-α | III | 84 | 101 | 16.83 | 93 | 4.666 | V - III | 24.314 | 1.259 | < 0.0001 |
| IV | 97 | 114 | 14.91 | 105 | 6.259 | V - IV | 12.366 | 1.116 | < 0.0001 | |
| V | 110 | 125 | 12.00 | 118 | 4.486 | IV- III | 11.948 | 1.127 | < 0.0001 | |
| TNF-α R1 | III | 82 | 88 | 6.81 | 85 | 1.856 | V - III | 1.859 | 1.021 | 0.0002 |
| IV | 83 | 90 | 7.77 | 87 | 1.796 | V - IV | -0.227 | 0.997 | 0.0002 | |
| V | 84 | 89 | 5.61 | 86 | 1.426 | IV- III | 2.087 | 1.024 | 0.0002 | |
Sixty three samples of degenerative IVD tissues were obtained from patients undergoing surgery due to spinal disc herniation. 4 x 105 cells from each sample were grown for four weeks in collagen I scaffold. The protein concentration data (ELISA) of anti-catabolic and inflammatory factors were statistically analyzed from 100 µg total protein extracts on the bases of disc degeneration grade (DDG).
Figure 4. Expression levels of anti-catabolic proteins TIMPs in degenerative NP cells.

NP cells were isolated from 63 samples of degenerative IVDs and 4 x 105 cells from each sample were cultured for four weeks in collagen I scaffold. Concentration data (ELISA) of TIMPs from 100 µg total protein extracts were statistically analyzed on the bases of disc degeneration grade (DDG). Box plots with whiskers min. to max. show TIMP-1 expression levels (Figure 4a), TIMP-2 expression levels (Figure 4b), TIMP-3 expression levels (Figure 4c) and TIMP-4 expression levels (Figure 4d).
Very low expression levels of inflammatory cytokines IL-1β, IL-1 R, TNF-α and TNF-α R1 were detected in degeneration grade III, IV and V. Compared to TNF-α and TNF-α R1, higher expression levels of IL-1β and IL-1 R were recorded. The mean expression values of IL-1β were 110 + 3.770 pg/ml, 121 + 6.326 pg/ml, 140 + 2.895 pg/ml (P < 0.0001), and of IL-1 R were 128 + 5.944 pg/ml, 146 + 8.229 pg/ml, 165 + 7.827 pg/ml (P < 0.0001). The mean expression values of TNF-α were 93 + 4.666 pg/ml, 105 + 6.259 pg/ml, 118 + 4.486 pg/ml (P < 0.0001) and of TNF-α R1 were 85 + 1.856 pg/ml, 87 + 1.796 pg/ml, 86 + 1.426 pg/ml (P = 0.0002) respectively. About 30 % increased mean expression value of IL-1β, IL-1 R and TNF-α were recorded between degeneration grade III - V , whereas the mean expression values of TNF-α R1 remained constant (table 4 and Figure 5).
Figure 5. Expression levels of inflammatory cytokines in degenerative NP cells.

NP cells were isolated from 63 samples of degenerative IVDs and 4 x 105 cells from each sample were cultured for four weeks in collagen I scaffold. Concentration data (ELISA) of inflammatory cytokines from 100 µg total protein extracts were statistically analyzed on the bases of disc degeneration grade (DDG). Box plots with whiskers min. to max. show IL-1β expression levels (Figure 5a), IL-1 R expression levels (Figure 5b), TNF-α expression levels (Figure 5c) and TNF-α R1 expression levels (Figure 5d).
The calculated significance of concentration changes as a function of age by decade showed increasing expression levels of MMP-1 (P < 0.0001) and TNF-α (P < 0.0001). Gender does not appear to play a role in influencing the expression levels of the analyzed catabolic, anti-catabolic and inflammatory proteins (data not shown).
Levels of anabolic and matrix proteins in degenerative NP cells
Matrix proteins aggrecan and collagen II were expressed in decreasing manner with respect to degeneration grade III, IV and V. Declining expression levels of aggrecan with mean expression values of 26371 + 1978 pg/ml, 14151 + 1284 pg/ml, 7970 + 718 pg/ml (P < 0.0001) and collagen II with mean expression values of 9766 + 414 pg/ml, 5821 + 517 pg/ml, 2673 + 283 pg/ml (P < 0.0001) were determined respectively. About 3.3 fold decreased mean expression values of aggrecan and similarly about 3.7 fold decreased mean expression values of collagen II were recorded between degeneration grade III - V (table 5 and Figure 6).
Table 5. Expression levels of anabolic factors and matrix proteins in adult NP cells.
| Anabolic proteins | DDG | Min. pg/ml | Max. pg/ml | Range % | Mean pg/ml | SD |
Mean difference
|
Mean fold | P value | |
|---|---|---|---|---|---|---|---|---|---|---|
| DDG | pg/ml | |||||||||
| Anabolic protein expression levels were below the minimum detectable dose (MDD). MDD of BMP-2 < 11 pg/ml, BMP-4 < 1 pg/ml, BMP-6 < 3 pg/ml, BMP-7 < 2 pg/ml, IGF-1 < 25 pg/ml, TGF-β1 and TGF-β3 < 5 pg/ml. | ||||||||||
| Matrix proteins | DDG | Min. pg/ml | Max. pg/ml | Range % | Mean pg/ml | SD |
Mean difference
|
Mean fold | P value | |
| DDG | pg/ml | |||||||||
| Aggrecan | III | 23663 | 29598 | 20.05 | 26371 | 1978 | III - V | 18400 | 3.308 | < 0.0001 |
| IV | 12556 | 16152 | 22.26 | 14151 | 1284 | III - IV | 12219 | 1.863 | < 0.0001 | |
| V | 6557 | 8793 | 25.42 | 7970 | 718 | IV- V | 6181 | 1.775 | < 0.0001 | |
| Collagen type II | III | 9037 | 10566 | 14.47 | 9766 | 414 | III – V | 7092 | 3.652 | < 0.0001 |
| IV | 5059 | 6390 | 20.82 | 5821 | 517 | III – IV | 3945 | 1.677 | < 0.0001 | |
| V | 2418 | 3269 | 26.03 | 2673 | 283 | IV- V | 3147 | 2.177 | < 0.0001 | |
Collagen I expression level was below MDD (MDD < 217 pg/ml).
Sixty three samples of degenerative IVD tissues were obtained from patients undergoing surgery due to spinal disc herniation. 4 x 105 cells from each sample were grown for four weeks in collagen I scaffold. The protein concentration data (ELISA) of anabolic factors and matrix proteins were statistically analyzed from 100 µg total protein extracts on the bases of disc degeneration grade (DDG).
Figure 6. Expression levels of matrix proteins in degenerative NP cells.

NP cells were isolated from 63 samples of degenerative IVDs and 4 x 105 cells from each sample were cultured for four weeks in collagen I scaffold. Concentration data (ELISA) of aggrecan and collagen II from 100 µg total protein extracts were statistically analyzed on the bases of disc degeneration grade (DDG). Box plots with whiskers min. to max. show aggrecan expression levels (Figure 6a) and collagen II expression levels (Figure 6b).
The protein expression level of collagen I was below the minimum detectable dose (MDD) of our detection system (MDD < 217 pg/ml). Neither control 3D culture in agarose gel nor 2D culture on TC dishes delivered additional result. Thus collagen I scaffold used for 3D culture did not inhibit collagen I expression. Although the minimum detectable dose of our detection system was extremely low for anabolic factors, the protein expression levels of all tested anabolic factors remained below the MDD. The recorded MDDs were for BMP-2 < 11 pg/ml, BMP-4 < 1 pg/ml, BMP-6 < 3 pg/ml, BMP-7 < 2 pg/ml, IGF-1 < 25 pg/ml, TGF-β1 and TGF-β3 < 5 pg/ml. The calculated significance of concentration changes as a function of age by decade showed decreasing expression levels of aggrecan (P < 0.0001) and collagen II (P < 0.0001). Gender does not appear to play a role in influencing the expression levels of the anabolic and matrix proteins (data not shown).
Discussion
NP cell density and IVD degeneration
Previous studies have shown that during childhood disc specimens exhibit considerably higher cell density than during adolescence [20-24]. During growth the cell environment within the disc changes harshly, as its size increases and blood supply as well as diffusion decrease, which results in decreased concentration of glucose and oxygen [25-29]. There are conflicting reports with regard to cell densities in different grades of degeneration in adulthood. Some studies reported a decline of NP cell densities with increasing degeneration [30-32]. A recent study, however, indicated comparable NP cell densities in adult age with different grades of IVD degeneration [20]. It may indicate that progressive degenerative changes in adult NP may not be caused by loss of NP cells but rather due to unfavorable phenotypic alternation of the NP cells. Similarly, our study showed comparable NP cell proliferation rates between the degeneration grades III, IV and V (table 2, Figure 2a). Neither age nor gender appears to play a role in influencing cell proliferation potential.
In this study inclusion criteria for surgery were radiographically determined intervertebral disc herniations with nerve root compression on MRI, which correlated to primary symptoms that remained unresponsive to non-operative treatment for six weeks or demonstrated progressive neurological deterioration in the face of conservative treatment. Our findings may therefore be representative of degenerative discs from such patients.
Imbalance between catabolic and anti-catabolic factors in degenerative IVDs
In contrast to previous findings that showed the percentage of immunohistological ADAMTS-5 positive cells correlating with the age of the patients [33], we found age to be an independent factor with regard to ADAMTS-5, which was highly expressed in all grades of degeneration. Additionally, we presented high and increasing expression levels of ADAMTS-4 with 3.5 fold amplification between degeneration grades III and V (table 3, Figure 2b-c). In human osteoarthritic cartilage, ADAMTS-4 and ADAMTS-5 have been shown to cause aggrecan degradation [34,35]. In vitro inhibition of ADAMTS-4 and ADAMTS-5 by siRNA in rat chondrocytes could significantly increase the aggrecan and collagen II content [36,37]. Additionally, in vivo inhibition of ADAMTS-5 by siRNA in a rabbit annular needle-puncture model resulted in improved MRI scores with increased signal intensity and improved histological grade scores [38]. Hence, both ADAMTS-4 and ADAMTS-5 might represent attractive targets for biological treatment of human IVD degeneration.
The expression of MMPs and their counter parts TIMPs in NP cells has been controversy discussed. On one hand, the number of immunopositive cells for MMP-1, -3, -13 and ADAMTS-4 increased with the severity of degeneration and this was accompanied by increased number of immunopositive cells for TIMP-1 and -2 but not for TIMP-3 [14]. On the other hand, the most extensive immunohistochemical staining was seen for MMP-1, -2, -3, and -9 and much less for MMP-7 and -8, and this up-regulation was paralleled by greater expression of TIMP-2 and not TIMP-1[39]. Moreover, consistent and substantial up-regulated mRNA levels of MMP-3 and MMP-8 were observed and this up-regulation was paralleled by greater expression of TIMP-1 and not TIMP-2 [15]. In another study, immunohistological staining was negative for TIMP-1 in MMP-3 positive stained surgical specimens [40]. Our protein concentration data of MMP-1, -2, -3, -7, -8, -9, -10 and -13 showed by far the highest concentration levels for MMP-3 and its levels did not increase with the severity of degeneration. The mean expression values of MMP-3 was about 20 fold of MMP-13, 30 fold of MMP-1 and 120 fold of MMP-2 (Table 3 and Figure 3). The expression values of MMP-8, -9 and -10 were below the MDD (Table 3).
Contrary to the above observations, where TIMPs were expressed at lower levels than MMP-3, we found higher expression levels of TIMP-1 and -2. The high levels of MMP-3 were counteracted by higher and increasing expression levels of TIMP-1(2.1 fold of MMP-3) and TIMP-2 (1.9 fold of MMP-3). In comparison, considerably less TIMP-3 and even much less TIMP-4 expression levels were recorded. The mean expression values of TIMP-3 and -4 were about 5.2 % and 1.2 % of TIMP-1 respectively (table 4 and Figure 4).
According to our data, repression of MMP-3 as well as ectopic expression of TIMP-3, an inhibitor of ADAMTs, would potentially be more effective to enhance the regeneration potential of NP cells. As the expression levels of TIMP-1 and -2 are higher than MMP-3, it would be more interesting to focus on their posttranslational modifications than their expression. Nonetheless, adenoviral delivery of TIMP-1 to degenerative human NP cells in vitro resulted in increased proteoglycan synthesis in pellet cultures [41].
Implication of inflammatory cytokines in the pathogenesis of IVD degeneration
In non-degenerative, degenerative and herniated human disc cells the expressions of IL-1β, TNF-α and their receptors IL-1 R and TNF-α R1 were investigated using quantitative real time PCR and immunohistochemistry. Higher levels of IL-1β, TNF-α and IL-1 R expression were observed in degenerative or herniated IVDs compared to non-degenerative IVDs [16]. In another study the expression of IL-1β, IL-1 R and IL-1 Ra were shown in non-degenerative and degenerative NP and the expressions of IL-1β and IL-1 R increased with severity of degeneration [39]. In our study IL-1β and IL-1 R were expressed at higher levels than TNF-α and TNF-α R1. The protein expression levels of IL-1β, IL-1 R and TNF-α were increased with severity of degeneration. However the expression levels of TNF-α R1 remained steady throughout all degeneration grades (table 4 and Figure 5). Even though both IL-1β and TNF-α could be implicated in the pathogenesis of disc degeneration, IL-1β could be a better therapeutic target for IVD regeneration. Furthermore, treatment of human NP and AF cells with 10 ng/ml recombinant IL-1β has shown increased expression of MMP-3, MMP-13 and ADAMTS-4 as well as decreased expression of aggrecan, collagen II, collagen I, and SOX6 [42]. In addition, treatment of NP cells with TNF-α and IL-1β increased the NF-κΒ dependent expression of syndecan-4 (SDC4), which may cause ADAMTS-5 mediated degradation of aggrecan [43].
Expression of growth factors and matrix proteins in degenerative IVDs
Age and degeneration grade related changes in the extracellular matrix of NP and AF of human IVDs have been presented [44,45]. Several growth factors such as TGF-βs, BMPs, IGF-1, bFGF, GDF-5 and PDGF have been shown to be important biological components for the stimulation of matrix synthesis [46]. Furthermore, the therapeutic potential of growth factors TGF-β1, PRP, BMP-2, BMP-12 and growth factors combination (TGF- β1, IGF-1 and BMP-2) has been discussed comprehensively [2]. However, the endogenous expression levels of growth factors in IVDs have scarcely been published. Only a few number of publications presented endogenous expression of growth factors bFGF, TGF-β1, TGF-β2 and growth factor receptors TGFβ RII, FGF R3 and BMP RI in normal and degenerative IVDs using immunohistochemical analysis [17,47]. In contrast, although we applied a large amount of total protein extract from NP cells (100 µg) and the minimum detectable doses for tested growth factors were very low (1 - 25 pg/ml), the protein expression levels of all tested growth factors (BMP-2, -4, -6, -7, IGF-1, TGF-β1 and TGF-β3) remained below the minimum detectable doses (table 5). Hence, our data underline an imbalance between the expression levels of anabolic and catabolic proteins in degenerative NP cells. Furthermore, we measured with increasing degeneration decreasing expression levels of aggrecan and collagen II. About 3.3 fold decreased mean expression values of aggrecan and 3.7 fold decreased mean expression values of collagen II were recorded between grade III - V (table 5 and Figure 6).
Conclusions
Considering the protein concentration data of the analyzed degenerative and regenerative factors of IVD, repression of the catabolic factors ADAMTS-4, ADAMTS-5 and MMP-3 as well as the inflammatory cytokines IL-1β and IL-1 R could be a promising approach. The repression course could be combined with ectopic expression of the anabolic factors and the anti-catabolic factor TIMP-3. This combinatorial process could be a prospective objective to enhance the regeneration potential of degenerative NP cells.
Acknowledgments
We thank Prof. Lothar Schilling for providing laboratory equipment.
Funding Statement
The study is financed by University Medical Center Mannheim, Heidelberg University. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Hegewald AA, Ringe J, Sittinger M, Thome C (2008) Regenerative treatment strategies in spinal surgery. Front Biosci 13: 1507-1525. doi: 10.2741/2777. PubMed: 17981645. [DOI] [PubMed] [Google Scholar]
- 2. Mern DS, Beierfuß A, Thomé C, Hegewald AA (2012) Enhancing human nucleus pulposus cells for biological treatment approaches of degenerative intervertebral disc diseases: a systematic review. J Tissue Eng Regen Med doi: 10.1002/term 1583 [DOI] [PubMed] [Google Scholar]
- 3. Zhao CQ, Wang LM, Jiang LS, Dai LY (2007) The cell biology of intervertebral disc aging and degeneration. Ageing Res Rev 6: 247-261. doi: 10.1016/j.arr.2007.08.001. PubMed: 17870673. [DOI] [PubMed] [Google Scholar]
- 4. Kalb S, Martirosyan NL, Kalani MYS, Broc GG, Theodore N (2012) Genetics of the Degenerated Intervertebral Disc. World Neurosurg 77: 491-501. doi: 10.1016/j.wneu.2011.07.014. PubMed: 22120330. [DOI] [PubMed] [Google Scholar]
- 5. Nomura T, Mochida J, Okuma M, Nishimura K, Sakabe K (2001) Nucleus pulposus allograft retards intervertebral disc degeneration. Clin Orthop Relat Res 389: 94-101. doi: 10.1097/00003086-200108000-00015. PubMed: 11501830. [DOI] [PubMed] [Google Scholar]
- 6. Meisel HJ, Siodla V, Ganey T, Minkus Y, Hutton WC et al. (2007) Clinical experience in cell based therapeutics: disc chondrocyte transplantation a treatment for degenerated or damaged intervertebral disc. Biomol Eng 24: 5-21. doi: 10.1016/j.bioeng.2006.07.002. PubMed: 16963315. [DOI] [PubMed] [Google Scholar]
- 7. Sakai D, Mochida J, Yamamoto Y, Nomura T, Okuma M et al. (2003) Transplantation of mesenchymal stem cells embedded in Atelocollagen gel to the intervertebral disc: a potential therapeutic model for disc degeneration. Biomaterials 24: 3531-3541. doi: 10.1016/S0142-9612(03)00222-9. PubMed: 12809782. [DOI] [PubMed] [Google Scholar]
- 8. Revell PA, Damien E, Di Silvio L, Gurav N, Longinotti C et al. (2007) Tissue engineered intervertebral disc repair in the pig using injectable polymers. J Mater Sci Mater Med 18: 303-308. doi: 10.1007/s10856-006-0693-6. PubMed: 17323162. [DOI] [PubMed] [Google Scholar]
- 9. Nishida K, Kang JD, Gilbertson LG, Moon SH, Suh JK et al. (1999) Modulation of the biologic activity of the rabbit intervertebral disc by gene therapy: an in vivo study of adenovirusmediated transfer of the human transforming growth factor-β1 encoding gene. Spine 24: 2419-2425. doi: 10.1097/00007632-199912010-00002. PubMed: 10626303. [DOI] [PubMed] [Google Scholar]
- 10. Nishida K, Doita M, Takada T, Kakutani K, Miyamoto H et al. (2006) Sustained transgene expression in intervertebral disc cells in vivo mediated by microbubbleenhanced ultrasound gene therapy. Spine 31: 1415-1419. doi: 10.1097/01.brs.0000219945.70675.dd. PubMed: 16741448. [DOI] [PubMed] [Google Scholar]
- 11. Okuma M, Mochida J, Nishimura K, Sakabe K, Seiki K (2000) Reinsertion of stimulated nucleus pulposus cells retards intervertebral disc degeneration: an in vitro and in vivo experimental study. J Orthop Res 18: 988-997. doi: 10.1002/jor.1100180620. PubMed: 11192261. [DOI] [PubMed] [Google Scholar]
- 12. Sive JI, Baird P, Jeziorsk M, Watkins A, Hoyland JA et al. (2002) Expression of chondrocyte markers by cells of normal and degenerate intervertebral discs. Mol Pathol 55: 91-97. doi: 10.1136/mp.55.2.91. PubMed: 11950957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Millward-Sadler SJ, Costello PW, Freemont AJ, Hoyland JA (2009) Regulation of catabolic gene expression in normal and degenerate human intervertebral disc cells: implications for the pathogenesis of intervertebral disc degeneration. Arthritis Res Ther 11(3): R65. doi: 10.1186/ar2693. PubMed: 19435506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Le Maitre CL, Freemont AJ, Hoyland JA (2004) Localization of degradative enzymes and their inhibitors in the degenerate human intervertebral disc. J Pathol 204: 47-54. doi: 10.1002/path.1608. PubMed: 15307137. [DOI] [PubMed] [Google Scholar]
- 15. Bachmeier BE, Nerlich A, Mittermaier N, Weiler C, Lumenta C et al. (2009) Matrix metalloproteinase expression levels suggest distinct enzyme roles during lumbar disc herniation and degeneration. Eur Spine J 18: 1573-1586. doi: 10.1007/s00586-009-1031-8. PubMed: 19466462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Le Maitre CL, Hoyland JA, Freemont AJ (2007) Catabolic cytokine expression in degenerate and herniated human intervertebral discs: IL-1beta and TNF-alpha expression profile. Arthritis Res Ther 9: R77. doi: 10.1186/ar2275. PubMed: 17688691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Le Maitre CL, Richardson SM, Baird P, Freemont AJ, Hoyland JA (2005) Expression of receptors for putative anabolic growth factors in human intervertebral disc: implications for repair and regeneration of the disc. J Pathol 207: 445-452. doi: 10.1002/path.1862. PubMed: 16278806. [DOI] [PubMed] [Google Scholar]
- 18. Pfirrmann CW, Metzdorf A, Zanetti M, Hodler J, Boos N (2001) Magnetic resonance classification of lumbar intervertebral disc degeneration. Spine 26: 1873-1878. doi: 10.1097/00007632-200109010-00011. PubMed: 11568697. [DOI] [PubMed] [Google Scholar]
- 19. Landis RJ, Koch GG (1977) The measurement of observer agreement for categorical data. Biometrics 33: 159-174. doi: 10.2307/2529310. PubMed: 843571. [DOI] [PubMed] [Google Scholar]
- 20. Liebscher T, Haefeli M, Wuertz K, Nerlich AG, Boos N (2011) Age-related variation in cell density of human lumbar intervertebral discs. Spine 36: 153-159. doi: 10.1097/BRS.0b013e3181cd588c. PubMed: 20671592. [DOI] [PubMed] [Google Scholar]
- 21. Gruber HE, Hanley EN Jr (2002) Ultrastructure of the human intervertebral disc during aging and degeneration: comparison of surgical and control specimens. Spine 27: 798-805. doi: 10.1097/00007632-200204150-00004. PubMed: 11935100. [DOI] [PubMed] [Google Scholar]
- 22. Antoniou J, Steffen T, Nelson F, Winterbottom N, Hollander AP et al. (1996) The human lumbar intervertebral disc: evidence for changes in the biosynthesis and denaturation of the extracellular matrix with growth, maturation, ageing, and degeneration. J Clin Invest 98: 996-1003. doi: 10.1172/JCI118884. PubMed: 8770872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Antoniou J, Goudsouzian NM, Heathfield TF, Winterbottom N, Steffen T et al. (1996) The human lumbar endplate. Evidence of changes in biosynthesis and denaturation of the extracellular matrix with growth, maturation, aging, and degeneration. Spine 21: 1153-1163. doi: 10.1097/00007632-199605150-00006. PubMed: 8727189. [DOI] [PubMed] [Google Scholar]
- 24. Gruber HE, Hanley EN Jr (2003) Recent advances in disc cell biology. Spine 28: 186-193. PubMed: 12544938. [DOI] [PubMed] [Google Scholar]
- 25. Nerlich AG, Schaaf R, Wächli B, Boos N (2007) Temporospatial distribution of blood vessels in human lumbar intervertebral discs. Eur Spine J 16: 547-555. doi: 10.1007/s00586-006-0213-x. PubMed: 16947015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Rajasekaran S, Babu JN, Arun R, Armstrong BR, Shetty AP et al. (2004) ISSLS prize winner: a study of diffusion in human lumbar discs: a serial magnetic resonance imaging study documenting the influence of the endplate on diffusion in normal and degenerate discs. Spine 29: 2654-2667. doi: 10.1097/01.brs.0000148014.15210.64. PubMed: 15564914. [DOI] [PubMed] [Google Scholar]
- 27. Bibby SR, Urban JP (2004) Effect of nutrient deprivation on the viability of intervertebral disc cells. Eur Spine J 13: 695-701. doi: 10.1007/s00586-003-0616-x. PubMed: 15048560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Selard E, Shirazi-Adl A, Urban JP (2003) Finite element Study of nutrient diffusion in the human intervertebral disc Spine 28: 1945-1953 [DOI] [PubMed] [Google Scholar]
- 29. Soukane DM, Shirazi-Adl A, Urban JP (2007) Computation of coupled diffusion of oxygen, glucose and lactic acid in an intervertebral disc. J Biomech 40: 2645-2654 [Google Scholar]
- 30. Hastreiter D, Ozuna RM, Spector M (2001) Regional variations in certain cellular characteristics in human lumbar intervertebral discs, including the presence of a-smooth muscle actin. J Orthop Res 19: 597-604. doi: 10.1016/S0736-0266(00)00069-3. PubMed: 11518268. [DOI] [PubMed] [Google Scholar]
- 31. Roberts S, Evans EH, Kletsas D, Jaffray DC, Eisenstein SM (2006) Senescence in human intervertebral discs. Eur Spine J 15 (suppl 3): 312-316. doi: 10.1007/s00586-006-0126-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Maroudas A, Stockwell RA, Nachemson A, Urban J (1975) Factors involved in the nutrition of the human lumbar intervertebral disc: cellularity and diffusion of glucose in vitro . J Anat 120: 113-130. PubMed: 1184452. [PMC free article] [PubMed] [Google Scholar]
- 33. Zhao CQ, Zhang YH, Jiang SD, Li H, Jiang LS et al. (2011) ADAMTS-5 and intervertebral disc degeneration: the results of tissue immunohistochemistry and in vitro cell culture. J Orthop Res 29: 718-725. doi: 10.1002/jor.21285. PubMed: 21437951. [DOI] [PubMed] [Google Scholar]
- 34. Malfait AM, Liu RQ, Ijiri K, Komiya S, Tortorella MD (2002) Inhibition of ADAM-TS4 and ADAM-TS5 prevents aggrecan degradation in osteoarthritic cartilage. J Biol Chem 277: 22201-22208. doi: 10.1074/jbc.M200431200. PubMed: 11956193. [DOI] [PubMed] [Google Scholar]
- 35. Naito S, Shiomi T, Okada A, Kimura T, Chijiiwa M et al. (2007) Expression of ADAMTS4 (aggrecanase-1) in human osteoarthritic cartilage. Pathol Int 57: 703-711. doi: 10.1111/j.1440-1827.2007.02167.x. PubMed: 17922681. [DOI] [PubMed] [Google Scholar]
- 36. Wang ZH, Yang ZQ, He XJ, Wang L, Li LX et al. (2008) Effects of RNAi-mediated inhibition of aggrecanase-1 and aggrecanase-2 on rat costochondral chondrocytes in vitro . Acta Pharmacol Sin 29: 1215-1226. doi: 10.1111/j.1745-7254.2008.00856.x. PubMed: 18817627. [DOI] [PubMed] [Google Scholar]
- 37. Wang ZH, Yang ZQ, He XJ, Kamal BE, Xing Z (2010) Lentivirus mediated knockdown of aggrecanase-1 and -2 promotes chondrocyte-engineered cartilage formation in vitro . Biotechnol Bioeng 107: 730-736. doi: 10.1002/bit.22862. PubMed: 20632367. [DOI] [PubMed] [Google Scholar]
- 38. Seki S, Asanuma-Abe Y, Masuda K, Kawaguchi Y, Asanuma K et al. (2009) Effect of small interference RNA (siRNA) for ADAMTS5 on intervertebral disc degeneration in the rabbit anular needle-puncture model. Arthritis Res Ther 11: R166. doi: 10.1186/ar2851. PubMed: 19889209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Roberts S, Caterson B, Menage J, Evans EH, Jaffray DC et al. (2000) Matrix metalloproteinases and aggrecanase: their role in disorders of the human intervertebral disc. Spine 25: 3005-3013. doi: 10.1097/00007632-200012010-00007. PubMed: 11145811. [DOI] [PubMed] [Google Scholar]
- 40. Kanemoto M, Hukuda S, Komiya Y, Katsuura A, Nishioka J (1996) Immunohistochemical study of matrix metalloproteinase-3 and tissue inhibitor of metalloproteinase-1 human intervertebral discs. Spine 21: 1-8. doi: 10.1097/00007632-199601010-00001. PubMed: 9122749. [DOI] [PubMed] [Google Scholar]
- 41. Wallach CJ, Sobajima S, Watanabe Y, Kim JS, Georgescu HI et al. (2003) Gene transfer of the catabolic inhibitor TIMP-1 increases measured proteoglycans in cells from degenerated human intervertebral discs. Spine 28: 2331-2337. doi: 10.1097/01.BRS.0000085303.67942.94. PubMed: 14560079. [DOI] [PubMed] [Google Scholar]
- 42. Le Maitre CL, Freemont AJ, Hoyland JA (2005) The role of interleukin-1 in the pathogenesis of human intervertebral disc degeneration. Arthritis Res Ther 7: R732-R745. doi: 10.1186/ar1732. PubMed: 15987475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Wang J, Markova D, Anderson DG, Zheng Z, Shapiro IM et al. (2011) TNF-α and IL-1β promote a disintegrin-like and metalloprotease with thrombospondin type I motif-5-mediated aggrecan degradation through syndecan-4 in intervertebral disc. J Biol Chem 286: 39738-39749. doi: 10.1074/jbc.M111.264549. PubMed: 21949132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Singh K, Masuda K, Thonar EJ, An HS, Cs-Szabo G (2009) Age-related changes in the extracellular matrix of nucleus pulposus and anulus fibrosus of human intervertebral disc. Spine 34: 10-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Nerlich AG, Schleicher ED, Boos N (1997) 1997 Volvo Award winner in basic science studies. Immunohistologic markers for age-related changes of human lumbar intervertebral discs. Spine 22: 2781-2795. doi: 10.1097/00007632-199712150-00001. PubMed: 9431614. [DOI] [PubMed] [Google Scholar]
- 46. Masuda K, Oegema TR Jr, An HS (2004) Growth factors and treatment of intervertebral disc degeneration. Spine 29: 2757-2769. Available online at: doi:10.1097/01.brs.0000146048.14946.af. PubMed: 15564925. [DOI] [PubMed] [Google Scholar]
- 47. Tolonen J, Grönblad M, Vanharanta H, Virri J, Guyer RD et al. (2006) Growth factor expression in degenerated intervertebral disc tissue. An immunohistochemical analysis of transforming growth factor beta, fibroblast growth factor and platelet-derived growth factor. Eur Spine J 15: 588-596. doi: 10.1007/s00586-005-0930-6. PubMed: 15980999. [DOI] [PMC free article] [PubMed] [Google Scholar]