

Abstract
Variants in the CDH13 gene have been identified as determinants of blood levels of adiponectin, an insulin-sensitizing adipokine. However, their association with other metabolic risk factors remains unclear. We examined variants at CDH13 in relation to total and high-molecular-weight (HMW) adiponectin using data from a genome-wide association study performed in 2,434 Singaporean Chinese with replication in up to 3,290 Japanese and 1,610 Koreans. The top signal rs4783244 in CDH13 showed strong associations with total adiponectin (standardized β [β] = −0.34, 95% CI −0.38 to −0.30, P = 2.0 × 10−70), HMW adiponectin (β = −0.40, 95% CI −0.43 to −0.36, P = 1.1 × 10−117), and the HMW-to-total adiponectin ratio (β = −0.44, 95% CI −0.49 to −0.40, P = 3.2 × 10−83). In the replication study, this single nucleotide polymorphism explained 4.1% of total and 6.5% of HMW adiponectin levels. No association was observed between rs4783244 and metabolic traits associated with insulin resistance before adjustment for HMW adiponectin levels. After adjustment for HMW adiponectin levels, the minor allele was associated with lower BMI (β = −0.15, 95% CI −0.19 to −0.11, P = 3.5 × 10−14), homeostasis model assessment-insulin resistance index (β = −0.16, 95% CI −0.20 to −0.12, P = 9.2 × 10−16), and triglycerides (β = −0.16, 95% CI −0.19 to −0.12, P = 1.3 × 10−16) and with higher HDL (β = 0.16, 95% CI 0.12 to 0.19, P = 2.1 × 10−17). CDH13 variants strongly influence plasma total and HMW adiponectin levels in East Asian populations but appear to alter adiponectin sensitivity, resulting in better metabolic health than expected based on circulating adiponectin levels.
Adiponectin is a protein abundantly secreted by adipose tissue with anti-inflammatory (1) and insulin-sensitizing properties (2). Blood levels of adiponectin are inversely associated with obesity (3), insulin resistance (4), and risk of type 2 diabetes (5). Genetic determinants account for a substantial proportion of the variation in plasma adiponectin (6), and genome-wide association studies (GWAS) have identified several loci associated with plasma adiponectin (7–10). Adiponectin exists in several forms in the blood. High-molecular-weight (HMW) adiponectin has shown stronger associations with insulin sensitivity and suppression of hepatic glucose production than other forms of adiponectin (11). Few existing GWAS have included both total and HMW adiponectin and compared the associations. This may be pertinent because CDH13 has been identified to code for T-cadherin, a specific receptor for hexameric and HMW adiponectin (12). In addition, the effects of variants at the CDH13 locus on insulin resistance and other metabolic risk factors remain unclear.
We therefore conducted a GWAS of total and HMW adiponectin in a Singaporean Chinese population. With an extension to other East Asian populations, we also examined the effects of CDH13 variants in relation to insulin resistance and associated metabolic traits.
RESEARCH DESIGN AND METHODS
Study populations.
Participants from several studies conducted in East Asians were used for the analyses presented here. These included 2,282 Chinese living in Singapore from the Singapore Prospective Study Program (SP2) (13), 3,290 Japanese from the Nomura study (14) and the Ehime University Hospital Antiaging Center (AAC) study (15), and 1,610 Koreans from the Yangpyeong Cohort Study (16). Detailed descriptions of these studies are included in the Supplementary Data.
Laboratory analyses and genotyping.
Fasting blood samples were obtained in all studies, and concentrations of total adiponectin, HMW adiponectin, and metabolic variables were measured with acceptable coefficients of variation. Details of the methods used are included in the Supplementary Data. Insulin resistance and β-cell function were calculated using homeostasis model assessment-insulin resistance (HOMA-IR) and HOMA–β-cell function (HOMA-B) indices.
Genotyping in SP2 was done on three different arrays (Illumina HumanHap 550, 610 Quad, and 1Mduov3 BeadChips; http://www.illumina.com). Details on genotyping and quality control measures are included in the Supplementary Data. For replication, blood-derived genomic DNA of a Japanese and Korean sample was used. The CDH13 single nucleotide polymorphism (SNP) rs4783244 was analyzed by a TaqMan probe assay (Applied Biosystems Co., Ltd., Foster City, CA) using commercially available primers and probes purchased from the Assay-on-Demand system (C_10076301_10).
Statistical analysis.
We standardized adiponectin and other metabolic variables to mean of 0 and variance of 1 to facilitate cross-study comparisons. Multiple linear regression analysis, based on additive and general genetic models, was used with different adiponectin forms and metabolic risk factors (log-transformed if necessary) as the dependent variable, and genotype, age, and sex were used as independent variables. In addition, for metabolic traits, multivariable models that also included HMW adiponectin (or total adiponectin) and BMI were evaluated. Results across studies were combined by a fixed-effect meta-analysis. Population structure in Singaporean Chinese was assessed by principal component analysis. A Bonferroni-corrected threshold α ≤ 5 × 10−8 was considered genome-wide significant, and α ≤ 0.005 was used as a cutoff for the tests on rs4783244 and 10 metabolic variables (calculated as 0.05/10). All tests were two-sided.
RESULTS
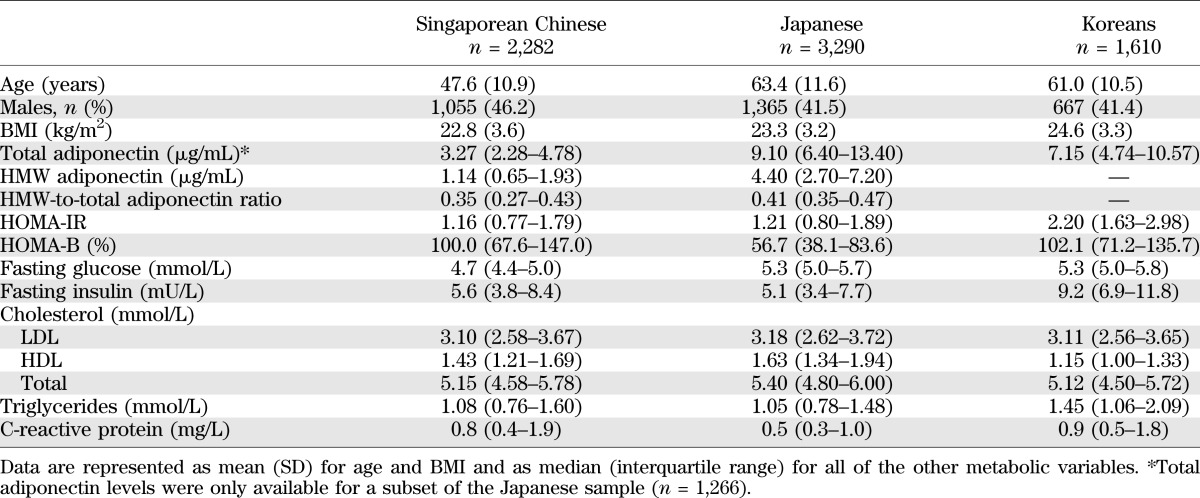
Characteristics of the participants in each study are summarized in Table 1. Singaporeans were generally younger than Japanese and Korean participants, whereas Koreans had a higher BMI, higher triglyceride levels, higher HOMA-IR, and lower HDL levels than the other populations. The substantial differences in adiponectin levels among study populations may be partly due to differences in laboratory methods and have been addressed by standardization of adiponectin levels in the data analysis. As expected, blood adiponectin levels were inversely correlated with insulin resistance (measured by HOMA-IR or fasting insulin), fasting glucose, triglycerides, and C-reactive protein and directly correlated with HDL in our study populations (Supplementary Table 1).
TABLE 1.
Characteristics of the study populations
In the GWAS in Singaporean Chinese, signals reaching genome-wide significance (5 ×10−8) mapped exclusively to the CDH13 and ADIPOQ gene (Supplementary Figs. 1 and 2). Associations for selected SNPs from these two genes and SNPs from other genes previously reported to be associated with adiponectin levels are listed in Supplementary Table 2. The strongest signal in CDH13 was rs4783244, located in the intron region. With regard to other previously reported loci, associations with total and HMW adiponectin levels reached genome-wide significance for ADIPOQ (rs10937273), and we observed nominally significant associations for GPR109A (rs601339), CMIP (rs2925979), and PEPD (rs731839) (Supplementary Table 2). The principal components were not correlated with total or HMW adiponectin levels (Supplementary Fig. 3 and Supplementary Table 3). For subsequent analyses, we focused on the top hit CDH13 SNP rs4783244.
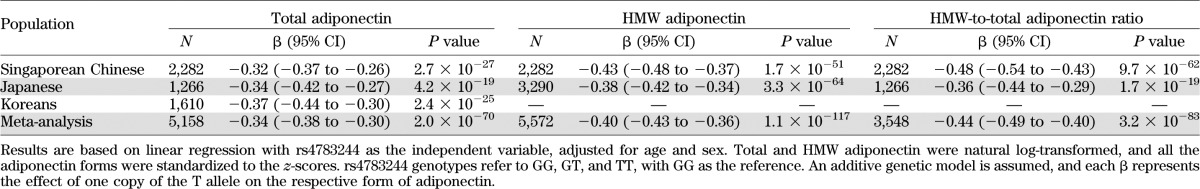
In the combined data from Singaporean Chinese, Japanese, and Korean cohorts, total adiponectin levels significantly decreased by 0.34 SD on the log scale for each additional T allele rs4783244 in CDH13 (95% CI −0.38 to −0.30, P = 2.0 × 10−70; Table 2). This CDH13 variant was even more strongly associated with HMW adiponectin levels (standardized β [β] = −0.40, 95% CI −0.43 to −0.36, P = 1.1 × 10−117) and the HMW-to-total adiponectin ratio (β = −0.44, 95% CI −0.49 to −0.40, P = 3.2 × 10−83) based on the Singaporean Chinese and Japanese data. Adjustment for BMI did not substantially affect these effect estimates, and similar results were obtained for the general genetic model (Supplementary Table 4). The CDH13 rs4783244 variant explained more than 4% of variation in total adiponectin (Singapore: 4.5%, Korea: 5.5%, Japan: 4.1%) and more than 6% of variation in HMW adiponectin levels (Singapore: 8.3%, Japan: 6.5%).
TABLE 2.
Association between rs4783244 in CDH13 and different forms of adiponectin
At a Bonferroni-corrected threshold of P ≤ 0.005, no significant association between rs4783244 in CDH13 and metabolic risk factors was observed in Singaporean Chinese or in Japanese (model 1; Table 3). Because CDH13 is known to code for a receptor for HMW adiponectin, we reassessed these associations after adjustment for HMW adiponectin levels (model 2; Table 3). In a meta-analysis of the Singaporean Chinese and Japanese samples, the minor allele T in rs4783244 was significantly associated with lower BMI (β = −0.15, 95% CI −0.19 to −0.11, P = 3.5 × 10−14), lower HOMA-IR (β = −0.16, 95% CI −0.20 to −0.12, P = 9.2 × 10−16), higher HDL (β = 0.16, 95% CI 0.12 to 0.19, P = 2.1 × 10−17), and lower triglycerides (β = −0.16, 95% CI −0.19 to −0.12, P = 1.3 × 10−16) after adjustment for HMW adiponectin levels (Fig. 1A). After further adjusting for BMI, associations were weaker but remained significant for HOMA-IR (β = −0.09, 95% CI −0.12 to −0.05, P = 5.4 × 10−7), HDL (β = 0.12, 95% CI 0.09 to 0.16, P = 5.4 × 10−12), and triglycerides (β = −0.12, 95% CI −0.16 to −0.09, P = 2.2 × 10−11; Fig. 1B). These associations were not driven by population admixture in Singaporean Chinese (Supplementary Table 5) and were significant but weaker when we adjusted for total adiponectin instead of HMW adiponectin (results not shown). A sensitivity analysis found minimal difference in effect estimates between fixed- and random-effect meta-analysis, and the estimates retained genome-wide significant with random-effect analysis.
TABLE 3.
Association between rs4783244 in CDH13 and metabolic traits with and without adjustment for HMW adiponectin levels
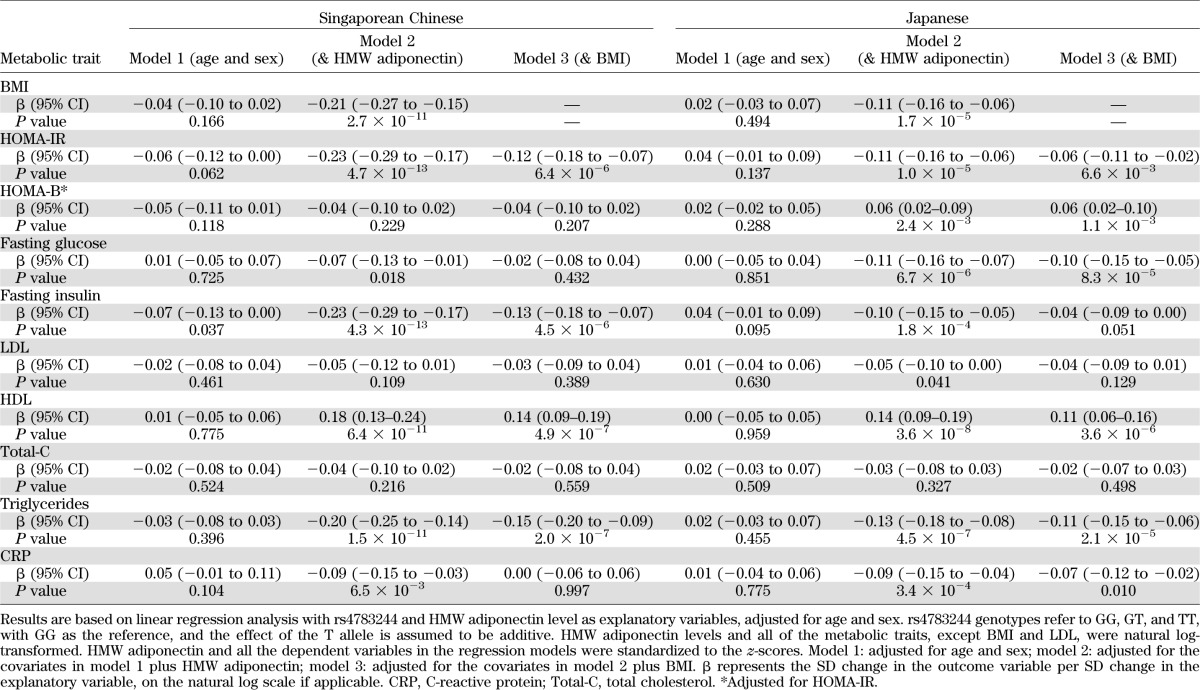
FIG. 1.

Effect estimates of rs4783244 in CDH13 on selected metabolic traits across studies adjusted for age, sex, and HMW adiponectin (A) and adjusted for age, sex, HMW adiponectin, and BMI (B). HMW adiponectin and levels of HOMA-IR, HDL, and triglycerides were natural log-transformed. HMW adiponectin and all the dependent variables in the regression models were standardized to the z-scores. β represents the SD change in the outcome variable per SD change in the explanatory variable, on the natural log scale if applicable. The solid squares denote the mean difference, the horizontal lines represent the 95% CIs, and the diamond denotes the weighted mean differences.
DISCUSSION
Our study replicates previously reported associations of variants at the CDH13, ADIPOQ, GPR109A, CMIP, and PEPD loci with blood adiponectin levels (10). Furthermore, we found that rs4783244 at the CDH13 locus, which encodes a receptor for hexameric and HMW adiponectin, was more strongly associated with HMW adiponectin than total adiponectin, explaining more than 6% of the variation in HMW adiponectin levels in East Asians.
However, rs4783244 at the CDH13 locus was not associated with other metabolic traits, which would be expected based on its association with adiponectin levels, if circulating adiponectin causally influences insulin resistance.
Results from previous studies also provided little support for an association between variants at the CDH13 locus and metabolic traits. In Filipino women, no significant associations with metabolic risk factors were detected for rs3865188 in CDH13 (linkage disequilibrium [LD] with rs4783244, r2 = 0.85) except for a nominal association (P = 0.042) with waist circumference (9). A Swedish study similarly reported that rs11646213, a SNP upstream of CDH13 in minimal LD with rs4783244 (r2 = 0.08), was not associated with metabolic risk factors (17). In a Taiwanese study, the significant associations between rs4783244 and waist circumference, glucose, and triglyceride levels did not remain after adjustment for BMI, although the adiponectin-lowering T allele was paradoxically still associated with a reduced risk for diabetes, the metabolic syndrome, and stroke (18).
After adjustment for total and HMW adiponectin levels, the CDH13 allele associated with lower blood adiponectin levels was associated with a better metabolic profile, including lower BMI, lower insulin resistance based on fasting insulin measures, lower triglyceride levels, and higher HDL levels. Japanese researchers recently reported an association between another CDH13 SNP (rs12051272) and BMI, fasting insulin, fasting glucose, HOMA-IR, and fasting triglycerides only after controlling for adiponectin levels (19). This SNP is close to and in moderate LD (r2 = 0.66) with rs4783244, which we studied. Together, the data suggest a complex relationship among variants at the CDH13 locus and metabolic traits that is only evident after controlling for their effects on blood adiponectin levels.
The association between the variants at CDH13 and plasma HMW adiponectin may be explained by the function of the T-cadherin receptor that it encodes. T-cadherin is a receptor for hexameric and HMW adiponectin that is expressed in the vasculature (20), cardiac myocytes (21), and epithelial cells in the lung (22). We believe that the T allele at rs4783244 is associated with increased binding of HMW adiponectin to the T-cadherin receptor, resulting in the sequestration of HMW adiponectin in these tissues and thus removing it from the blood. Consistent with this explanation, ablation of the T-cadherin receptor increased plasma adiponectin levels in mice (20–22). However, this does not explain the paradoxical observation that the T allele at rs4783244, which is associated with lower blood levels of HMW adiponectin, is associated with a more favorable metabolic profile than would be expected based on HMW adiponectin levels.
We hypothesize that the rs4783224 variant at the CDH13 locus may have an indirect effect on an individual’s sensitivity to circulating adiponectin. In this hypothesis, the chronically low levels of plasma adiponectin associated with the T allele may result in upregulation of adiponectin receptors AdipoR1/R2. Consistent with this proposed mechanism, chronic elevation of plasma adiponectin led to downregulation of AdipoR2 in adipose tissue in mice (23). Furthermore, the expression of AdipoR1/R2 was upregulated in insulin-resistant women with polycystic ovary syndrome (24), who would be expected to have low blood adiponectin levels. The greater expression of adiponectin receptors could counterbalance the low adiponectin levels, resulting in the lack of association between rs4783244 and the metabolic profile in unadjusted analyses. However, when the blood adiponectin levels are controlled for, then the greater “adiponectin sensitivity” results in an association between the T allele and a more favorable metabolic profile. Alternatively, effects of CDH13 on T-cadherin expression and receptor function may directly affect insulin sensitivity. A recent study identified the role of T-cadherin in regulating insulin action in the endothelium such that upregulation of T-cadherin promoted endothelial insulin resistance (25).
Strengths of our study include the relatively large sample size in a homogeneous Chinese population, replication in independent Japanese and Korean populations, and availability of HMW adiponectin and other metabolic variables in addition to total adiponectin. As a limitation, we are unable to elucidate the underlying biological pathways behind our epidemiological observations.
In summary, our study showed that a genetic variant in CDH13 explains a substantial part of variation in HMW adiponectin levels in East Asian populations. However, this effect of CDH13 on circulating HMW adiponectin levels did not appear to translate into effects on metabolic traits related to insulin resistance, suggesting that compensatory mechanisms exist that lead to greater “adiponectin sensitivity.” Further mechanistic studies on the complex interaction between CDH13, blood adiponectin levels, and metabolic traits are needed to better understand the physiologic significance of these observations.
Supplementary Material
ACKNOWLEDGMENTS
The SP2 study was supported by the Biomedical Research Council (grant number 03/1/27/18/216) and the National Medical Research Council (grant numbers 0838/2004 and NMRC/CSI/0002/2005). The Japanese studies were supported by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan, a Science and Technology Incubation Program in Advanced Regions from the Japan Science and Technology Agency, a Grant-in-Aid for Scientific Research from the Japan Arteriosclerosis Prevention Fund, and a Research Promotion Award of Ehime University.
No potential conflicts of interest relevant to this article were reported.
H.G. formulated the proposal, performed data analysis, interpreted the results, and drafted the manuscript. Y.-M.K. performed the field survey, collected samples, and analyzed data for the Yangpyeong study. P.C., R.T.-H.O., and X.S. contributed to the SP2 study GWAS data analysis. M.I. and R.K. collected samples for the Japanese studies. M.K.K. supervised the field survey and analyzed data for the Yangpyeong study. K.K. and T.M. collected samples and supervised the study design for the Japanese studies. J.L. designed the SP2 study and supervised data collection. H.On. performed experiments for the Japanese studies. H.Os. supervised the statistical analysis for the Japanese studies. Y.Y.T. supervised the data analysis for the SP2 study. Y.T. collected samples, performed experiments, analyzed data, supervised the Japanese studies, and edited the manuscript. ES.T. designed the SP2 study, conceived the study design, interpreted the results, contributed to the writing of the manuscript, and supervised the project progress. R.M.v.D. conceived the study design, interpreted the results, contributed to the writing of the manuscript, and supervised the project progress. All the authors reviewed and approved the final version of the manuscript. R.M.v.D. is the guarantor of this work and, as such, had full access to all the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis.
Footnotes
This article contains Supplementary Data online at http://diabetes.diabetesjournals.org/lookup/suppl/doi:10.2337/db13-0129/-/DC1.
Y.T., E.S.T., and R.M.v.D. jointly directed this work.
REFERENCES
- 1.Tilg H, Moschen AR. Adipocytokines: mediators linking adipose tissue, inflammation and immunity. Nat Rev Immunol 2006;6:772–783 [DOI] [PubMed] [Google Scholar]
- 2.Kadowaki T, Yamauchi T, Kubota N, Hara K, Ueki K, Tobe K. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J Clin Invest 2006;116:1784–1792 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hotta K, Funahashi T, Arita Y, et al. Plasma concentrations of a novel, adipose-specific protein, adiponectin, in type 2 diabetic patients. Arterioscler Thromb Vasc Biol 2000;20:1595–1599 [DOI] [PubMed] [Google Scholar]
- 4.Lawlor DA, Davey Smith G, Ebrahim S, Thompson C, Sattar N. Plasma adiponectin levels are associated with insulin resistance, but do not predict future risk of coronary heart disease in women. J Clin Endocrinol Metab 2005;90:5677–5683 [DOI] [PubMed] [Google Scholar]
- 5.Li S, Shin HJ, Ding EL, van Dam RM. Adiponectin levels and risk of type 2 diabetes: a systematic review and meta-analysis. JAMA 2009;302:179–188 [DOI] [PubMed] [Google Scholar]
- 6.Cesari M, Narkiewicz K, De Toni R, Aldighieri E, Williams CJ, Rossi GP. Heritability of plasma adiponectin levels and body mass index in twins. J Clin Endocrinol Metab 2007;92:3082–3088 [DOI] [PubMed] [Google Scholar]
- 7.Richards JB, Waterworth D, O’Rahilly S, et al. GIANT Consortium A genome-wide association study reveals variants in ARL15 that influence adiponectin levels. PLoS Genet 2009;5:e1000768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Qi L, Menzaghi C, Salvemini L, De Bonis C, Trischitta V, Hu FB. Novel locus FER is associated with serum HMW adiponectin levels. Diabetes 2011;60:2197–2201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wu Y, Li Y, Lange EM, et al. Genome-wide association study for adiponectin levels in Filipino women identifies CDH13 and a novel uncommon haplotype at KNG1-ADIPOQ. Hum Mol Genet 2010;19:4955–4964 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Dastani Z, Hivert MF, Timpson N, et al. DIAGRAM+ Consortium. MAGIC Consortium. GLGC Investigators. MuTHER Consortium. DIAGRAM Consortium. GIANT Consortium. Global B Pgen Consortium. Procardis Consortium. MAGIC investigators. GLGC Consortium Novel loci for adiponectin levels and their influence on type 2 diabetes and metabolic traits: a multi-ethnic meta-analysis of 45,891 individuals. PLoS Genet 2012;8:e1002607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lara-Castro C, Luo N, Wallace P, Klein RL, Garvey WT. Adiponectin multimeric complexes and the metabolic syndrome trait cluster. Diabetes 2006;55:249–259 [PubMed] [Google Scholar]
- 12.Hug C, Wang J, Ahmad NS, Bogan JS, Tsao TS, Lodish HF. T-cadherin is a receptor for hexameric and high-molecular-weight forms of Acrp30/adiponectin. Proc Natl Acad Sci USA 2004;101:10308–10313 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Nang EE, Khoo CM, Tai ES, et al. Is there a clear threshold for fasting plasma glucose that differentiates between those with and without neuropathy and chronic kidney disease? the Singapore Prospective Study Program. Am J Epidemiol 2009;169:1454–1462 [DOI] [PubMed] [Google Scholar]
- 14.Tabara Y, Kohara K, Kita Y, et al. Global Blood Pressure Genetics Consortium Common variants in the ATP2B1 gene are associated with susceptibility to hypertension: the Japanese Millennium Genome Project. Hypertension 2010;56:973–980 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Tabara Y, Igase M, Kido T, Ochi N, Miki T, Kohara K. Composition of lower extremity in relation to a high ankle-brachial index. J Hypertens 2009;27:167–173 [DOI] [PubMed] [Google Scholar]
- 16.Yang YJ, Choi BY, Chun BY, et al. Dietary zinc intake is inversely related to subclinical atherosclerosis measured by carotid intima-media thickness. Br J Nutr 2010;104:1202–1211 [DOI] [PubMed] [Google Scholar]
- 17.Fava C, Danese E, Montagnana M, et al. A variant upstream of the CDH13 adiponectin receptor gene and metabolic syndrome in Swedes. Am J Cardiol 2011;108:1432–1437 [DOI] [PubMed] [Google Scholar]
- 18.Chung CM, Lin TH, Chen JW, et al. A genome-wide association study reveals a quantitative trait locus of adiponectin on CDH13 that predicts cardiometabolic outcomes. Diabetes 2011;60:2417–2423 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Morisaki H, Yamanaka I, Iwai N, et al. CDH13 gene coding T-cadherin influences variations in plasma adiponectin levels in the Japanese population. Hum Mutat 2012;33:402–410 [DOI] [PubMed] [Google Scholar]
- 20.Hebbard LW, Garlatti M, Young LJ, Cardiff RD, Oshima RG, Ranscht B. T-cadherin supports angiogenesis and adiponectin association with the vasculature in a mouse mammary tumor model. Cancer Res 2008;68:1407–1416 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Denzel MS, Scimia MC, Zumstein PM, Walsh K, Ruiz-Lozano P, Ranscht B. T-cadherin is critical for adiponectin-mediated cardioprotection in mice. J Clin Invest 2010;120:4342–4352 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhu M, Hug C, Kasahara DI, et al. Impact of adiponectin deficiency on pulmonary responses to acute ozone exposure in mice. Am J Respir Cell Mol Biol 2010;43:487–497 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bauche IB, Ait El Mkadem S, Rezsohazy R, et al. Adiponectin downregulates its own production and the expression of its AdipoR2 receptor in transgenic mice. Biochem Biophys Res Commun 2006;345:1414–1424 [DOI] [PubMed] [Google Scholar]
- 24.Tan BK, Chen J, Digby JE, Keay SD, Kennedy CR, Randeva HS. Upregulation of adiponectin receptor 1 and 2 mRNA and protein in adipose tissue and adipocytes in insulin-resistant women with polycystic ovary syndrome. Diabetologia 2006;49:2723–2728 [DOI] [PubMed] [Google Scholar]
- 25.Philippova M, Joshi MB, Pfaff D, et al. T-cadherin attenuates insulin-dependent signalling, eNOS activation, and angiogenesis in vascular endothelial cells. Cardiovasc Res 2012;93:498–507 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.