

Abstract
Tumour cells exploit both genetic and adaptive means to survive and proliferate in hypoxic microenvironments, resulting in the outgrowth of more aggressive tumour cell clones. Direct measurements of tumour oxygenation, and surrogate markers of the hypoxic response in tumours (for instance, hypoxia inducible factor-1α, carbonic anhydrase 9 and glucose transporter-1) are well-established prognostic markers in solid cancers. However, individual markers do not fully capture the complex, dynamic and heterogeneous hypoxic response in cancer. To overcome this, expression profiling has been employed to identify hypoxia signatures in cohorts or models of human cancer. Several of these hypoxia signatures have demonstrated prognostic significance in independent cancer datasets. Nevertheless, individual hypoxia markers have been shown to predict the benefit from hypoxia-modifying or anti-angiogenic therapies. This review aims to discuss the clinical impact of translational work on hypoxia markers and to explore future directions for research in this area.
Keywords: cancer, hypoxia, microarray, pathology, prognosis
Introduction
Accurate prognostic information is invaluable to both cancer patients and clinicians. It permits clinicians to make evidence-based recommendations on the incremental survival benefit of specific treatments or to recommend a palliative approach where appropriate. Many patients value accurate prognoses, which may help them to plan their future and make informed decisions regarding their medical care. The increasing choice of different chemotherapy regimens, including expensive targeted biological therapies, has placed renewed emphasis on predicting subsets of patients who are most or least likely to benefit. Tumour morphology, histopathological stage and/or grade are the prognostic gold standard for most cancer types. However, there are a number of instances where staging provides inadequate prognostic information. For example, the 5-year survival of patients with stage Ia lung cancer is only 40%[1]; histopathology is unable to predict which of these patients will relapse. The current TNM staging criteria for oesophageal cancer fails to adequately segregate patients into discrete prognostic groups, with no significant difference in survival between stages 0 and IIa in one study (P= 0.52) [2]. The predicted survival of patients with pancreatic endocrine cancers is dependent on their metastatic potential [3]. Unfortunately, prognosis and future metastases are not accurately predicted by routine histopathology [3]. Such deficiencies provide settings to develop and test other prognostic markers that may better inform clinical decisions.
Chronic hypoxia applies a selective pressure to neoplastic cells that limits the growth of early lesions (to within 180 μm of blood vessels) [4], until the tumour is able to activate the ‘angiogenic switch’ by secreting pro-angiogenic factors [5]. For example, vascular endothelial growth factor, which is secreted secondary to oncogenic signalling (e.g. Wnt, Ras and members of the epidermal growth factor receptor family) [6–9] and hypoxia-dependent transcription [10–15]. Tumour angiogenesis is poorly coordinated compared to neoangiogenesis during embryonic development [16]. As a consequence, large regions of the rich tumour vasculature are often non-functional, resulting in acutely hypoxic microenvironments within most tumours [17]. Acute hypoxia is often transient, with subsequent reperfusion generating reactive oxygen free-radicals that may damage cells in the affected tissue [18]. Hypoxic cancer cells use genetic and adaptive changes to permit survival and proliferation under these conditions (reviewed in [19]). Indeed, preclinical research and mathematical modelling suggests that the increased cellular stress observed in these hypoxic regions applies a selective pressure, which drives the clonal outgrowth of more aggressive cancers [20–23]. The consequence of increased angiogenesis secondary to hypoxia may also drive tumour growth [24] and facilitate metastasis [25]. A hypothesis follows that the more hypoxic a tumour, the more aggressive it will be and the worse the prognosis for the patient. Furthermore, there is clinical evidence to suggest that hypoxia and surrogate markers of hypoxia may mediate and therefore predict resistance to anti-cancer therapies, including radiotherapy [26], hypoxia-modifying therapy [27–31] and anti-angiogenic drugs [32].
A growing body of translational research has tested these hypotheses in exploratory retrospective subset analyses of patient cohorts and clinical trials [33]. The aim of this review is to discuss the evidence supporting the use of pathological markers of hypoxia in predicting patient survival and response to therapy targeting hypoxic tumour cells. In addition, the emerging role of expression profiling to define hypoxia signatures with prognostic significance will be considered.
Pathological markers of hypoxia
Hypoxia is defined as a partial pressure of oxygen below that required for normal cellular functions [34]. This is a physiological definition, which can only truly be assessed in patients by the invasive monitoring of cancers before they are resected. For example, the pO2 in cervical cancer has been measured using an oxygen electrode [35, 36]. Data from four of five independent studies suggest that a pO2≤5 or ≤10 mmHg is associated with a reduced likelihood of disease-free survival in cervical cancer [35, 37–41] (the one dissenting study found no evidence that hypoxia affects patient survival) [42]. However, this invasive method may be unpleasant for patients and it is not technically feasible for tumours that are not readily accessible (e.g. pancreatic cancer). The variable nature of tumour hypoxia, with well-perfused microenvironments co-existing alongside severe hypoxia, may not be accurately captured using a large single electrode (perhaps explaining the heterogeneity among studies of hypoxia in cervical cancer). These limitations have prompted research into in situ surrogate markers of hypoxia.
Necrosis
Rapidly growing cancers often exhibit regions of central necrosis, as they outgrow their blood supply, and necrosis is more frequently observed in solid tumours with increasing distance from a well-vascularized stroma [4]. Our colleagues have described associations between the presence of a fibrotic focus, hypoxia, angiogenesis and survival in breast [43] and pancreatic carcinoma [44]. Yet, necrosis and fibrosis are only observed in areas of extreme hypoxic-stress or anoxia. In areas of the tumour where hypoxia is not as severe, hypoxic stress may not be readily apparent morphologically. Morphology alone, therefore, tends to underestimate the frequency of hypoxic microenvironments in human cancer. Moreover, the presence of hypoxia in tumours may only be part of the story. It is the way in which hypoxia influences the tumour cells, their ability to survive, adapt and exploit hypoxia to develop aggressive treatment-resistant clones that is biologically and clinically important. As a consequence, markers of the tumour cells’ response to hypoxia have been investigated, to better understand the interaction between cancer and its microenvironment and how this influences the clinical behaviour of tumours.
Hypoxia inducible factor (HIF) targets
The hypoxic response involves genes concerned with numerous different cellular functions that contribute to the hallmarks of cancer [45], e.g. self-sufficiency in growth signals (IGF-2) [46, 47], evading apoptosis (IAP-2) [48], sustained angiogenesis (VEGF) [11, 12, 14], limitless replicative potential (telomerase) [49], tissue invasion (carbonic anhydrase 9) [50, 51] and metastasis (lysyl oxidase) [52, 53]. The expression of many of these genes is controlled at the transcriptional level by key transcription factors such as HIF-1 and HIF-2 [54] (discussed elsewhere in this issue) [55]. However, the hypoxic response may also limit tumour progression; high expression of HIF-1α and HIF-2α may reduce the clonogenic survival of hypoxic breast cancer cells in vitro[11], and many tumour suppressors are induced by hypoxia (e.g. BCL-interacting killer [56, 57] and DEC1 [58, 59]). Therefore, the biological behaviour of tumours (and the clinical implications of this) depends on the balance between the oncogenic and tumour suppressive effects of the hypoxic response within different microenvironments.
To date prognostic significance has been reported for numerous individual hypoxia markers, with a range of cellular functions, in numerous solid tumour types, including breast adenocarcinoma, head and neck cancer, cervical carcinoma and soft-tissue sarcomas
[33]. Global hypoxia signatures are associated with a worse outcome and, invariably, expression of individual hypoxia markers is reported to be associated with shorter survival [33, 60]. Indeed, in challenging settings where pathological staging performs poorly (such as early stage lung cancer [61] and pancreatic endocrine cancer [62]), markers of hypoxia have demonstrated that they can outperform pathological stage or provide discriminatory prognostic information within a single pathological stage.
HIF-1 versus HIF-2
Each tumour type has an individual hypoxic response [60], and the prognostic significance of specific markers varies widely among them. Nevertheless, the transcriptional regulators of hypoxia, HIF-1 and HIF-2, and well described target genes, such as carbonic anhydrase 9 (a HIF-1, but not HIF-2, target) [63, 64] and glucose transporter-1 (a HIF-1 and/or HIF-2 target) [64, 65], are reported to have prognostic significance in numerous tumour types [33]. The transcriptional response to hypoxia mediated by HIF-1 is distinct from that mediated by HIF-2 [66], and this is reflected in the heterogeneous hypoxic response in cancer. For example, in breast cancer, the hypoxic response is predominantly mediated through HIF-1 with a minor contribution from HIF-2 [64]. Nuclear HIF-1α expression is widely reported to be a significant adverse prognostic factor in breast cancer [67–72] and the strength of the prognostic impact of a hypoxia expression signature is largely a feature of HIF-1 (not HIF-2)-mediated transcription [60]. In contrast, Helczynska and colleagues report that HIF-2α, not HIF-1α, has prognostic significance in breast cancer [73]. This might be explained by heterogeneity among studies (including patient demographics, the methods used to identify markers, scoring, sub-group cut-offs and statistical analyses), or publication bias. To investigate this, we performed a meta-analysis of reported hazard ratios for HIF-1α in multivariate analyses of disease-free survival in breast cancer from six published studies that included these data [68–72, 74] (unfortunately the dissenting study [73] did not include these data in their published manuscript). Overall, nuclear HIF-1α expression was a significant adverse prognostic factor, associated with a clinically significant hazard ratio for disease-free survival of 1.80 (95% confidence intervals, 1.32 to 2.47; Fig. 1A). A funnel plot suggests there may be publication bias in the reported prognostic significance of HIF-1α in breast cancer, with a one-sided P-value for publication bias of 0.066 (Begg and Mazumdar rank correlation test; Fig. 1B). Although, the classic fail-safe N estimates an additional 42 null studies would need to be included in the meta-analysis for the P-value to exceed 0.05. Despite this, scientists and clinicians must consider the widely reported variations in the significance of hypoxia markers depending on the stage of disease, patient characteristics and the methods used. Validation of a standardized approach is essential if these findings are to be used in clinical practice.
Fig 1.

(A) A forest plot of the prognostic significance of HIF-1α in published multivariate analyses of disease free survival in breast cancer [68–72, 74]. (B) A funnel plot of the studies described in (A).
Early versus late stage cancer
To confuse matters further, certain hypoxia markers have been reported to have different prognostic significance in pre-invasive versus invasive lesions; BNIP3 (a HIF-1 target gene and a pro-apoptotic member of the Bcl-2 gene family) [75], is reported to be a good prognostic factor in ductal carcinoma in situ, but a poor prognostic factor in invasive ductal carcinoma [76]. In cervical cancer, HIF-1α expression is associated with a trend towards a good prognosis in large tumours, but towards a poor prognosis in small tumours [77]. Perhaps these observations are a reflection of the growth inhibitory effects of HIF-1αin vitro (due to direct effects on proliferation and cell death) [11] as opposed to the growth promoting effects of HIF-1αin vivo (due to effects on angiogenesis and metastasis) [78]. Alternatively, they may be due to differences in the relative contribution of acute hypoxia (due to transient ischemia, secondary to dysfunctional angiogenesis; frequently co-existing with reperfusion injury) versus chronic hypoxia (due to cells existing at the limits of oxygen diffusion) in driving HIF-1α expression. There is biological evidence to suggest that acute hypoxia has a greater effect on promoting tumour metastasis than chronic hypoxia [79]. Moreover, the presence of HIF-1α in acute hypoxia provides a selective advantage to neoplastic cells, but in chronic hypoxia HIF-1α expression may promote necrosis of the tumour [80]. In another preclinical model, prolonged hypoxia was associated with increased radiosensitivity (versus normoxia), as opposed to the resistance to radiotherapy conferred by intermittent hypoxia [81]. One may hypothesize that this difference is due to modulation of the HIF-1 transcriptional program in acute versus chronic hypoxia; perhaps due to differences in microenvironmental conditions, such as pH [82].
Intratumour heterogeneity
Hypoxia and expression of hypoxia markers, is heterogeneous throughout tumours (Fig. 2). Therefore, it is crucial that the appropriate regions of whole sections are assessed for hypoxia and tissue microarrays are designed with sufficient core redundancy to represent hypoxic and perinecrotic areas [83]. Vleugel and colleagues have reported that perinecrotic HIF-1α expression has a greater adverse prognostic impact, than diffuse HIF-1α expression [84]. Similarly, our own data on head and neck cancer suggest that certain hypoxia markers only have prognostic significance when assessed in perinecrotic regions (Dr. Adrian Jubb, unpublished data; Fig. 3A, B). Perhaps this is because markers of hypoxia may also be up-regulated by oncogenic signalling or cellular stress that is not directly related to the hypoxic response [85]; the context in which expression is observed and the pattern of expression are, therefore, critical. For instance, diffuse expression of carbonic anhydrase 9 (a HIF-1 target) in clear-cell renal carcinomas is often secondary to constitutive activation of HIF-1α due to genetic inactivation of the VHL tumour suppressor (Fig. 4A) [86]. By contrast, the patchy expression of carbonic anhydrase 9 in renal papillary carcinomas with wild-type VHL is secondary to hypoxia (Fig. 4B) [86].
Fig 2.

Intratumour heterogeneity of immunohistochemical reactivity for carbonic anhydrase 9, showing that expression (brown diaminobenzidine staining) is limited to a minority of the cancer cells at the periphery of a head and neck cancer (arrows). Bar = 1 mm.
Fig 3.

Kaplan–Meier disease-free survival curves for a novel hypoxia marker scored in tissue microarray cores from random regions of head and neck cancer (n= 82 patients) (A) or tissue microarray cores from perinecrotic areas of head and neck cancer (B).
Fig 4.
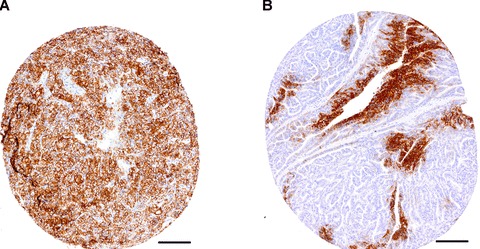
Diffuse expression of carbonic anhydrase 9 (brown diaminobenzidine staining) in a clear cell renal carcinoma with an inactivating VHL mutation (A), compared to hypoxic membranous expression of carbonic anhydrase 9 (brown) in a papillary renal carcinoma with wild-type VHL (B). Bar = 100 μm.
The time taken to induce and turnover hypoxia markers influences their clinical significance. Winter and colleagues have reported that HIF-1α, but not its transcriptional target carbonic anhydrase 9, has prognostic significance in a series of head and neck cancers [87]. By contrast, Generali and colleagues have reported that carbonic anhydrase 9, but not HIF-1α has prognostic significance in a series of breast cancers [68]. This might be due to HIF-1α-independent regulation of carbonic anhydrase 9 [88], but their expression was positively associated [68, 87]. Similarly, another group has reported disparity in the prognostic significance of hypoxia and several hypoxia-inducible proteins [89]. These results may be a reflection of the speed of induction and turnover (i.e. half-life) of these markers in response to hypoxia. HIF-1α protein stabilization is rapidly turned on and off, resulting in a short half-life measured in minutes [90]. By contrast carbonic anhydrase 9 is slow to accumulate and slow to turnover, with a half-life measured in hours [90]. Therefore, carbonic anhydrase 9 is a more sensitive marker of tissue hypoxia (it is more strongly associated with necrosis) [90, 91], but is not as accurate a reflection of transient fluctuations in tumour oxygenation as HIF-1α[90]. For instance, all cells of a clear-cell renal carcinoma may express carbonic anhydrase 9, under control of HIF-1, but nuclear HIF-1α may only be evident in 50% of cells (Fig. 5A, B). Similarly, HIF-1α may be rapidly and strongly induced adjacent to early necrotic lesions, before expression of carbonic anhydrase 9 has been fully induced (Fig. 5C) [84]. This has implications for study design and interpretation, but is rarely considered in the methods or discussion of hypoxia marker studies.
Fig 5.

HIF-1α expression in ∼50% of tumour cell nuclei (brown diaminobenzidine staining) from a clear cell renal carcinoma (A), in contrast to membranous carbonic anhydrase 9 (brown) expression in ∼100% of tumour cells in a serial section (B). Double staining for nuclear HIF-1α (brown) and membranous carbonic anhydrase 9 (blue) adjacent necrosis in a breast cancer (C). Bar = 100 μm. C, 40×. (C) was reproduced with permission from reference [84].
Quality of reports and analyses
The confusion in the literature may be explained, in part, by limitations in the conduct of tumour marker prognosis studies (many of which were published before the REMARK criteria were enforced) [92] and, in part, by the limitations of using a single marker to inform a complex, dynamic and heterogeneous disease process. Furthermore, assays are not done to GLP standards and criteria for staining and scoring vary widely. To overcome these limitations, expression signatures corresponding with specific tumour types and subtypes have been sought, to provide a more comprehensive picture of the hypoxic response, taking into account the variation in individual response to hypoxia with a more quantitative and reproducible technology.
Molecular signatures of hypoxia
Gene expression profiling has been used extensively to predict outcome in cancer patients. Initially, research groups used a supervised analysis to identify prognostic signatures, i.e. genes were selected that were most strongly associated with clinical outcome [93, 94]. However, if signatures are associated with specific phenotypes, they have the potential to be not only prognostic but also to predict benefit from specific interventions. Expression profiling of mRNA transcript levels has been exploited to identify a molecular signature associated with hypoxia and prognosis in several different model systems.
Comparison of tumour with in vitro hypoxic response profiles
Chi and colleagues examined the variation in the transcriptional response to hypoxia among different cell types and how this variation might relate to tissue-specific diseases using cDNA-microarrays [60]. A wide variation in the hypoxic response was observed between cell types and tissue types. However, they identified a 253-gene signature, including genes with common changes in response to hypoxia in epithelial cells; specifically, human mammary epithelial cells and renal proximal tubule epithelial cells. This signature showed coordinated variation in several human cancers, and it was a strong predictor of clinical outcome in breast and ovarian cancer [60]. The authors were able to stratify human cancers according to their hypoxic response and showed that a group of breast cancers, characterized by high expression of genes in the hypoxia signature, had a significantly worse overall and metastasis-free survival. This signature was also an independent prognostic factor in a multivariate analysis including clinicopathological variables and a previously derived wound signature [60].
A re-analysis of the experimental data from the above study focused on the time dependency of hypoxia-regulated gene expression by human mammary epithelial cells [95]. The early response to hypoxia was characterized by genes concerned with growth, apoptosis, the insulin pathway, oestrogen receptor signalling and transcription factors such as ATF3 (a regulator of the unfolded protein response) [96] and BACH1 (a hypoxia-inducible repressor of hemoxygenase-1) [97]. The late response to hypoxia was similar to the signature identified by Chi and colleagues, and included genes involved in angiogenesis, glucose transport, proliferation, metabolism and apoptosis [95]. The prognostic potential of early (1–6 hrs) and late (12–24 hrs) hypoxia signatures was tested in analyses of disease-specific survival in breast cancer. The authors reported that the early response signatures were prognostic in univariate analyses (P= 0.004 for signatures defined on exposure to 0% oxygen and P= 0.034 for 2% oxygen); late hypoxia signatures were not prognostic [95]. However, this significance was not maintained in multivariate analyses. Moreover, another group could not confirm the prognostic significance of Chi and colleagues’ hypoxia signature in predicting the local recurrence of breast cancer [98]. These data suggest that hypoxia signatures defined in vitro may not adequately reflect the complexity of the hypoxic response in human cancer and more detailed analyses are required.
To better understand this complexity, Chen and colleagues attempted to dissect transcriptional responses to lactic acidosis and hypoxia in a gene expression study of primary human mammary epithelial cells [99]. While the hypoxia signature was a statistically significant adverse prognostic marker in several publicly available breast cancer datasets, high expression of the lactic acidosis response signature was associated with good prognosis, independent of the hypoxia signature [99]. The authors hypothesized that this may relate to the role of lactic acidosis in directing energy generation towards aerobic respiration, and utilization of other energy sources, via repression of glycolysis and Akt inhibition [99].
In vivo clustering with seed genes
A data-mining approach to define a hypoxia signature in cancer in vivo has been explored by Winter and colleagues [100]. They derived an in vivo hypoxia metagene in head and neck squamous cell carcinoma (HNSCC) by clustering around the gene expression of a small set of hypoxia-regulated genes (e.g. carbonic anhydrase 9, glucose transporter-1 and VEGF) that have been well validated [19]. To minimize random association and false-positive findings, new genes were included in the metagene only if they clustered with several hypoxia-regulated genes. This approach was chosen to identify genes that are missed in in vitro experiments, but may play an important role in the hypoxic response in vivo, where the situation is more complex, requiring consideration of multiple cell types, microenvironmental changes and three-dimensional complexity. For example, VEGF was not present in the hypoxia signature derived by Chi and colleagues in epithelial cells [60], but it is a well-validated hypoxia-regulated gene [14] and plays a major role in cancer [101]. The 99-gene metagene identified by Winter and colleagues was prognostic for relapse-free survival in independent HNSCC and breast cancer datasets [100]. In addition, the identification of novel hypoxia-regulated genes has provided new avenues for research to better understand the hypoxic response in cancer.
A recent study by Murat and colleagues [102] used unsupervised clustering methods to identify a hypoxia-related signature in glioblastoma. This included several known hypoxia-inducible genes involved in angiogenesis and inflammation such as VEGF [14] and BIRC3(cIAP-2) [103], respectively. Furthermore, the hypoxic-regulation of several new genes comprised in this signature was experimentally confirmed in glioma cell lines and primary monocytes exposed to hypoxia [102]. Within the signature, several independent components were identified; among them, the relationship between hypoxia-modulated angiogenic genes and inflammatory genes was the only component associated with outcome in glioblastoma treated with chemoradiotherapy [102].
MicroRNAs
MicroRNAs are small non-coding RNA sequences that are thought to regulate cellular processes by mechanisms including mRNA turnover, inhibition of translation, promoter activation and epigenetic silencing [104]. Recently, several hypoxia-inducible microRNAs have been identified [105, 106]. For example, Camps and colleagues profiled changes in microRNA expression in MCF7 breast cancer cells exposed to hypoxia to identify several novel hypoxia-inducible microRNAs [105]. Of these genes, has-miR-210 was reported to be an independent prognostic factor in breast cancer of a clinically significant margin (hazard ratio for overall survival 11.38, 95% confidence intervals 4.1 to 31.7) [105]. Thus, investigating the micro-RNA response to hypoxia in cancer, and its relation to clinical outcome, might provide useful complementary information to gene expression studies.
In summary, molecular signatures of the hypoxic response have shown in statistically and clinically significant prognostic potential (Table 1). This is encouraging, but large discrepancies are present between signatures. In part, they might be due to a lack of standardization; although, standards such as MIAME [107] are continuously improving the reproducibility and transferability of results, and studies such as MACQC [108] have shown good reproducibility between different array technologies. Alternatively, discrepancies might reflect real biological differences in the hypoxic response in different tissues types, as some of these studies have already demonstrated. Furthermore, different components of the transcriptional response to hypoxia might play different roles in conferring resistance to specific treatments. This needs to be addressed in predictive gene expression studies where a specific interaction of the hypoxia transcriptional response with a given treatment is tested.
Table 1.
A summary of molecular signatures of hypoxia and cancer prognosis
| Reference | Signature derived from | Size of gene signature | Cancer tissue-type | Univariate analyses | Multivariate analyses |
|---|---|---|---|---|---|
| Chi et al.[60] | Mammary and renal tubular epithelial cells in vitro (hypoxia [2% or 0% O2] and normoxia, 12 hour time-points) | 253 genes | Breast cancer (2 independent datasets), ovarian cancer | Breast: overall survival, relapse-free survival, time to recurrence Ovarian: overall survival, relapse-free survival | Breast: overall survival, metastasis at first event |
| Seigneuric et al.[95] | Chi et al.[60] and human mammary epithelial cells in vitro (Hypoxia [2% or 0.02% O2] and normoxia, early 1–6 hour and late 12–24 hour time-points) | Chi: 253 genes Early: 15 genes Late: 93 genes | Breast cancer | Chi: not disease-specific survival Early: disease-specific survival Late: not disease-specific survival | Early: not disease-specific survival |
| Nuyten et al.[98] | Chi et al.[60] | 253 genes | Breast cancer | Not local recurrence | |
| Chen et al.[99] | Human mammary epithelial cells in vitro (hypoxia [2% O2]± 25 mM lactic acidosis [pH 6.7]) | 1585 genes for lactic acidosis 217 genes for hypoxia | Breast cancer (4 independent datasets) | Hypoxia: overall survival Lactic acidosis: overall survival | Hypoxia: overall survival Lactic acidosis: overall survival |
| Winter et al.[100] | 59 head and neck squamous cell carcinomas; clustering around known hypoxia-regulated genes | 99 genes | Head and neck cancer, breast cancer | Head and neck cancer: recurrence-free survival Breast cancer: overall survival, metastasis-free survival | Head and neck cancer: recurrence-free survival Breast cancer: overall survival, metastasis-free survival |
| Murat et al.[102] | 80 glioblastomas and 4 non-neoplastic brain tissues; unsupervised clustering | 52 genes | Glioblastoma (4 datasets, including the test-set) | Overall survival |
Markers of hypoxia and response to treatment
Clinical data have shown a strong correlation between the degree and extent of tumour hypoxia and resistance to radiotherapy in certain tumour types (e.g. head and neck cancer, cervical cancer and soft-tissue sarcoma; reviewed in [109]). Preclinical data suggest that radio-resistance is due, in part, to a preferentially pro-survival transcriptional response to HIF-1 expression by hypoxic cells [81, 110]. The hypothesis that hypoxia limits the curability of irradiated human cancers is difficult to prove, but there is emerging evidence that hypoxia markers can identify patients who are relatively radioresistant. For example, in head and neck cancer, the CHART (continuous hyperfractionated accelerated radiotherapy) clinical trial randomized 918 patients to receive accelerated radiotherapy (54 Gy over 12 days) versus conventional radiotherapy (66 Gy over 6–7 weeks) [111]. Unfortunately, the primary clinical end-point (an improvement in locoregional control) was not met (hazard ratio of 0.96, with 95% confidence intervals of 0.79 to 1.17, for CHART versus conventional radiotherapy) and the two treatment arms were considered equivalent [111]. However, retrospective subset analyses suggest that hypoxic tumours (identified by nuclear HIF-2α and/or membranous carbonic anhydrase 9 immunoreactivity) in this clinical trial were more resistant to CHART than conventional radiotherapy (interaction analysis, one-tailed P= 0.05) [112]. This is perhaps because CHART does not permit time for sufficient tumour re-oxygenation between doses, reducing radiation-induced cell death [112].
Furthermore, data from three independent trials suggest that hypoxic tumours experience the greatest benefit from hypoxia-modifying therapy. The first study showed that the level of pimonidazole binding (a hypoxia marker) in head and neck tumours predicted the likely benefit from hypoxia-modifying ARCON (accelerated radiotherapy plus carbogen [a hyperoxic gas] and nicotinamide [a vitamin B3 derivative], which are believed to improve tumour oxygenation), with survival rates of ∼60% and ∼18% for hypoxic tumours receiving ARCON compared to conventional radiotherapy, respectively [27, 28]. The second study involved a retrospective subset analysis of a randomized-controlled head and neck cancer trial (DAHANCA5, The Danish Head and Neck Cancer Group), which showed that the addition of hypoxia-modifying nimorazole (an oxygen mimic with a high affinity for electrons, that renders hypoxic cells radiosensitive) to conventional radiotherapy was associated with an increase in locoregional control (49%versus 33%, respectively) and overall survival (26%versus 16%, respectively) [29]. Patients in the DAHANCA5 trial with high plasma osteopontin levels (associated with tumour hypoxia) [113] were most likely to benefit from nimorazole [30]. Disease-specific survival rates were 51% and 21% for patients with high osteopontin levels undergoing hypoxia-modifying versus radiotherapy alone [30]. A third study showed that patients with hypoxic tumours (identified using 18F-FMISO positron-emission tomography) had an improved outcome following chemoradiotherapy plus the bioreductive agent tirapazamine (a cytotoxic agent with selective activity in hypoxic cells), compared with hypoxic tumours that received chemoradiotherapy alone (100%versus 39% locoregional control rate) [31]. These data suggest that hypoxia may be used to individualize cancer patient treatment, and work is ongoing in our laboratory to investigate this further.
In addition, hypoxia may play an important role in predicting the efficacy of anti-angiogenic drugs. The anti-VEGF receptor small molecule inhibitor vatalanib only significantly improved the median progression-free survival in two trials of first/second-line therapy for metastatic colorectal cancer, if the patients had high circulating levels of the hypoxia marker lactate dehydrogenase [32]. Similar analyses were negative in prospectively-planned subset analyses of the pivotal trial of the anti-VEGF monoclonal antibody bevacizumab in first-line metastatic colorectal cancer [114]. However, carbonic anhydrase 9 significantly predicted survival outcome in a single-arm phase II study of bevacizumab plus irinotecan in malignant astrocytoma [90]. Further work is necessary to determine whether this is a prognostic or a predictive association, and whether this observation applies to other cancer types.
Concluding remarks
Individual hypoxia markers have demonstrated independent prognostic significance in common tumour types, such as breast cancer [33], and in settings where routine histopathological staging has failed to accurately inform the prognosis [61, 62]. Nevertheless, hypoxia is heterogeneous throughout tumours, the adaptive and genetic consequences of hypoxia vary between and within tumour types, and hypoxia is a dynamic process that is in a constant state of flux. Expression profiling has been employed to better understand this complex process and several groups have identified novel hypoxic signatures with prognostic significance in solid human cancers [60, 100]. Nonetheless, hypoxia markers and expression signatures associated with hypoxia have yet to enter routine clinical practice. It is important to note that several new imaging agents are available to analyse hypoxia [31, 115], which can indicate the hypoxic volume of a tumour and may be used in radiotherapy planning. However, these modalities can indicate the degree of hypoxia, but not how individual tumours respond to hypoxia. How these two variables intersect with therapy will be a critical issue to assess in prospective randomized controlled trials. Recent studies of hypoxia-modifying and anti-angiogenic therapy have emphasized the potential utility of clinically significant hypoxia markers in guiding patient therapy [29, 30, 32]. Current and emerging evidence support a critical role for hypoxia markers in guiding individualized cancer therapies in the future.
Acknowledgments
To all members of the Harris Laboratory who contributed to the work reviewed here, and NHS consultant pathologists and surgeons involved in tumour sample collection and analysis. This study was supported by Cancer Research UK, the Experimental Cancer Medicine Centre Network, the European Union Metoxia Framework and the NIHR Biomedical Research Centre, Oxford.
References
- 1.Buccheri G, Ferrigno D. Prognostic value of stage grouping and TNM descriptors in lung cancer. Chest. 2000;117:1247–55. doi: 10.1378/chest.117.5.1247. [DOI] [PubMed] [Google Scholar]
- 2.Rizk NP, Venkatraman E, Bains MS, et al. American Joint Committee on Cancer staging system does not accurately predict survival in patients receiving multimodality therapy for esophageal adenocarcinoma. J Clin Oncol. 2007;25:507–12. doi: 10.1200/JCO.2006.08.0101. [DOI] [PubMed] [Google Scholar]
- 3.Roldo C, Missiaglia E, Hagan JP, et al. MicroRNA expression abnormalities in pancreatic endocrine and acinar tumors are associated with distinctive pathologic features and clinical behavior. J Clin Oncol. 2006;24:4677–84. doi: 10.1200/JCO.2005.05.5194. [DOI] [PubMed] [Google Scholar]
- 4.Tomlinson R, Gray L. The histological structure of some lung cancers and the possible implications for radiotherapy. Br J Cancer. 1955;9:539–49. doi: 10.1038/bjc.1955.55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Almog N, Ma L, Raychowdhury R, et al. Transcriptional switch of dormant tumors to fast-growing angiogenic phenotype. Cancer Res. 2009;69:836–44. doi: 10.1158/0008-5472.CAN-08-2590. [DOI] [PubMed] [Google Scholar]
- 6.Zhang X, Gaspard JP, Chung DC. Regulation of vascular endothelial growth factor by the Wnt and K-ras pathways in colonic neoplasia. Cancer Res. 2001;61:6050–4. [PubMed] [Google Scholar]
- 7.Petit AM, Rak J, Hung MC, et al. Neutralizing antibodies against epidermal growth factor and ErbB-2/neu receptor tyrosine kinases down-regulate vascular endothelial growth factor production by tumor cells in vitro and in vivo: angiogenic implications for signal transduction therapy of solid tumors. Am J Pathol. 1997;151:1523–30. [PMC free article] [PubMed] [Google Scholar]
- 8.Casanova ML, Larcher F, Casanova B, et al. A critical role for ras-mediated, epidermal growth factor receptor-dependent angiogenesis in mouse skin carcinogenesis. Cancer Res. 2002;62:3402–7. [PubMed] [Google Scholar]
- 9.Rak J, Mitsuhashi Y, Bayko L, et al. Mutant ras oncogenes upregulate VEGF/VPF expression: implications for induction and inhibition of tumor angiogenesis. Cancer Res. 1995;55:4575–80. [PubMed] [Google Scholar]
- 10.Mizukami Y, Li J, Zhang X, et al. Hypoxia-inducible factor-1-independent regulation of vascular endothelial growth factor by hypoxia in colon cancer. Cancer Res. 2004;64:1765–72. doi: 10.1158/0008-5472.can-03-3017. [DOI] [PubMed] [Google Scholar]
- 11.Blancher C, Moore JW, Talks KL, et al. Relationship of hypoxia-inducible factor (HIF)-1alpha and HIF-2alpha expression to vascular endothelial growth factor induction and hypoxia survival in human breast cancer cell lines. Cancer Res. 2000;60:7106–13. [PubMed] [Google Scholar]
- 12.Liu Y, Cox SR, Morita T, et al. Hypoxia regulates vascular endothelial growth factor gene expression in endothelial cells. Identification of a 5′ enhancer. Circ Res. 1995;77:638–43. doi: 10.1161/01.res.77.3.638. [DOI] [PubMed] [Google Scholar]
- 13.Ryan HE, Lo J, Johnson RS. HIF-1 alpha is required for solid tumor formation and embryonic vascularization. EMBO J. 1998;17:3005–15. doi: 10.1093/emboj/17.11.3005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Shweiki D, Itin A, Soffer D, et al. Vascular endothelial growth factor induced by hypoxia may mediate hypoxia-initiated angiogenesis. Nature. 1992;359:843–5. doi: 10.1038/359843a0. [DOI] [PubMed] [Google Scholar]
- 15.Maxwell PH, Dachs GU, Gleadle JM, et al. Hypoxia-inducible factor-1 modulates gene expression in solid tumors and influences both angiogenesis and tumor growth. Proc Natl Acad Sci USA. 1997;94:8104–9. doi: 10.1073/pnas.94.15.8104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Skinner SA, Frydman GM, O’Brien PE. Microvascular structure of benign and malignant tumors of the colon in humans. Dig Dis Sci. 1995;40:373–84. doi: 10.1007/BF02065424. [DOI] [PubMed] [Google Scholar]
- 17.Brown JM, Giaccia AJ. The unique physiology of solid tumors: opportunities (and problems) for cancer therapy. Cancer Res. 1998;58:1408–16. [PubMed] [Google Scholar]
- 18.Li C, Wright MM, Jackson RM. Reactive species mediated injury of human lung epithelial cells after hypoxia-reoxygenation. Exp Lung Res. 2002;28:373–89. doi: 10.1080/01902140290092001. [DOI] [PubMed] [Google Scholar]
- 19.Harris AL. Hypoxia–a key regulatory factor in tumour growth. Nat Rev Cancer. 2002;2:38–47. doi: 10.1038/nrc704. [DOI] [PubMed] [Google Scholar]
- 20.Anderson AR, Weaver AM, Cummings PT, et al. Tumor morphology and phenotypic evolution driven by selective pressure from the microenvironment. Cell. 2006;127:905–15. doi: 10.1016/j.cell.2006.09.042. [DOI] [PubMed] [Google Scholar]
- 21.Graeber TG, Osmanian C, Jacks T, et al. Hypoxia-mediated selection of cells with diminished apoptotic potential in solid tumours. Nature. 1996;379:88–91. doi: 10.1038/379088a0. [DOI] [PubMed] [Google Scholar]
- 22.Kim CY, Tsai MH, Osmanian C, et al. Selection of human cervical epithelial cells that possess reduced apoptotic potential to low-oxygen conditions. Cancer Res. 1997;57:4200–4. [PubMed] [Google Scholar]
- 23.Kondo A, Safaei R, Mishima M, et al. Hypoxia-induced enrichment and mutagenesis of cells that have lost DNA mismatch repair. Cancer Res. 2001;61:7603–7. [PubMed] [Google Scholar]
- 24.Kim KJ, Li B, Winer J, et al. Inhibition of vascular endothelial growth factor-induced angiogenesis suppresses tumour growth in vivo. Nature. 1993;362:841–4. doi: 10.1038/362841a0. [DOI] [PubMed] [Google Scholar]
- 25.Claffey KP, Brown LF, Del Aguila LF, et al. Expression of vascular permeability factor/vascular endothelial growth factor by melanoma cells increases tumor growth, angiogenesis, and experimental metastasis. Cancer Res. 1996;56:172–81. [PubMed] [Google Scholar]
- 26.Wouters BG, Brown JM. Cells at intermediate oxygen levels can be more important than the “hypoxic fraction” in determining tumor response to fractionated radiotherapy. Radiat Res. 1997;147:541–50. [PubMed] [Google Scholar]
- 27.Kaanders JH, Wijffels KI, Marres HA, et al. Pimonidazole binding and tumor vascularity predict for treatment outcome in head and neck cancer. Cancer Res. 2002;62:7066–74. [PubMed] [Google Scholar]
- 28.Kaanders JH, Pop LA, Marres HA, et al. ARCON: experience in 215 patients with advanced head-and-neck cancer. Int J Radiat Oncol Biol Phys. 2002;52:769–78. doi: 10.1016/s0360-3016(01)02678-5. [DOI] [PubMed] [Google Scholar]
- 29.Overgaard J, Hansen HS, Overgaard M, et al. A randomized double-blind phase III study of nimorazole as a hypoxic radiosensitizer of primary radiotherapy in supraglottic larynx and pharynx carcinoma. Results of the Danish Head and Neck Cancer Study (DAHANCA) Protocol 5–85. Radiother Oncol. 1998;46:135–46. doi: 10.1016/s0167-8140(97)00220-x. [DOI] [PubMed] [Google Scholar]
- 30.Overgaard J, Eriksen JG, Nordsmark M, et al. Plasma osteopontin, hypoxia, and response to the hypoxia sensitiser nimorazole in radiotherapy of head and neck cancer: results from the DAHANCA 5 randomised double-blind placebo-controlled trial. Lancet Oncol. 2005;6:757–64. doi: 10.1016/S1470-2045(05)70292-8. [DOI] [PubMed] [Google Scholar]
- 31.Rischin D, Hicks RJ, Fisher R, et al. Prognostic significance of [18F]-misonidazole positron emission tomography-detected tumor hypoxia in patients with advanced head and neck cancer randomly assigned to chemoradiation with or without tirapazamine: a substudy of Trans-Tasman Radiation Oncology Group Study 98.02. J Clin Oncol. 2006;24:2098–104. doi: 10.1200/JCO.2005.05.2878. [DOI] [PubMed] [Google Scholar]
- 32.Scott EN, Meinhardt G, Jacques C, et al. Vatalanib: the clinical development of a tyrosine kinase inhibitor of angiogenesis in solid tumours. Expert Opin Investig Drugs. 2007;16:367–79. doi: 10.1517/13543784.16.3.367. [DOI] [PubMed] [Google Scholar]
- 33.Vaupel P, Mayer A. Hypoxia in cancer: significance and impact on clinical outcome. Cancer Metastasis Rev. 2007;26:225–39. doi: 10.1007/s10555-007-9055-1. [DOI] [PubMed] [Google Scholar]
- 34.Hockel M, Vaupel P. Tumor hypoxia: definitions and current clinical, biologic, and molecular aspects. J Natl Cancer Inst. 2001;93:266–76. doi: 10.1093/jnci/93.4.266. [DOI] [PubMed] [Google Scholar]
- 35.Hockel M, Knoop C, Schlenger K, et al. Intratumoral pO2 predicts survival in advanced cancer of the uterine cervix. Radiother Oncol. 1993;26:45–50. doi: 10.1016/0167-8140(93)90025-4. [DOI] [PubMed] [Google Scholar]
- 36.Gatenby RA, Kessler HB, Rosenblum JS, et al. Oxygen distribution in squamous cell carcinoma metastases and its relationship to outcome of radiation therapy. Int J Radiat Oncol Biol Phys. 1988;14:831–8. doi: 10.1016/0360-3016(88)90002-8. [DOI] [PubMed] [Google Scholar]
- 37.Fyles A, Milosevic M, Hedley D, et al. Tumor hypoxia has independent predictor impact only in patients with node-negative cervix cancer. J Clin Oncol. 2002;20:680–7. doi: 10.1200/JCO.2002.20.3.680. [DOI] [PubMed] [Google Scholar]
- 38.Hockel M, Schlenger K, Aral B, et al. Association between tumor hypoxia and malignant progression in advanced cancer of the uterine cervix. Cancer Res. 1996;56:4509–15. [PubMed] [Google Scholar]
- 39.Hockel M, Schlenger K, Hockel S, et al. Hypoxic cervical cancers with low apoptotic index are highly aggressive. Cancer Res. 1999;59:4525–8. [PubMed] [Google Scholar]
- 40.Knocke TH, Weitmann HD, Feldmann HJ, et al. Intratumoral pO2-measurements as predictive assay in the treatment of carcinoma of the uterine cervix. Radiother Oncol. 1999;53:99–104. doi: 10.1016/s0167-8140(99)00139-5. [DOI] [PubMed] [Google Scholar]
- 41.Lyng H, Sundfor K, Trope C, et al. Disease control of uterine cervical cancer: relationships to tumor oxygen tension, vascular density, cell density, and frequency of mitosis and apoptosis measured before treatment and during radiotherapy. Clin Cancer Res. 2000;6:1104–12. [PubMed] [Google Scholar]
- 42.Nordsmark M, Loncaster J, Aquino-Parsons C, et al. The prognostic value of pimonidazole and tumour pO2 in human cervix carcinomas after radiation therapy: a prospective international multi-center study. Radiother Oncol. 2006;80:123–31. doi: 10.1016/j.radonc.2006.07.010. [DOI] [PubMed] [Google Scholar]
- 43.Colpaert CG, Vermeulen PB, Fox SB, et al. The presence of a fibrotic focus in invasive breast carcinoma correlates with the expression of carbonic anhydrase IX and is a marker of hypoxia and poor prognosis. Breast Cancer Res Treat. 2003;81:137–47. doi: 10.1023/A:1025702330207. [DOI] [PubMed] [Google Scholar]
- 44.Couvelard A, O’Toole D, Leek R, et al. Expression of hypoxia-inducible factors is correlated with the presence of a fibrotic focus and angiogenesis in pancreatic ductal adenocarcinomas. Histopathology. 2005;46:668–76. doi: 10.1111/j.1365-2559.2005.02160.x. [DOI] [PubMed] [Google Scholar]
- 45.Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100:57–70. doi: 10.1016/s0092-8674(00)81683-9. [DOI] [PubMed] [Google Scholar]
- 46.Christofori G, Naik P, Hanahan D. A second signal supplied by insulin-like growth factor II in oncogene-induced tumorigenesis. Nature. 1994;369:414–8. doi: 10.1038/369414a0. [DOI] [PubMed] [Google Scholar]
- 47.Feldser D, Agani F, Iyer NV, et al. Reciprocal positive regulation of hypoxia-inducible factor 1alpha and insulin-like growth factor 2. Cancer Res. 1999;59:3915–8. [PubMed] [Google Scholar]
- 48.Dong Z, Venkatachalam MA, Wang J, et al. Up-regulation of apoptosis inhibitory protein IAP-2 by hypoxia. Hif-1-independent mechanisms. J Biol Chem. 2001;276:18702–9. doi: 10.1074/jbc.M011774200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Seimiya H, Tanji M, Oh-hara T, et al. Hypoxia up-regulates telomerase activity via mitogen-activated protein kinase signaling in human solid tumor cells. Biochem Biophys Res Commun. 1999;260:365–70. doi: 10.1006/bbrc.1999.0910. [DOI] [PubMed] [Google Scholar]
- 50.Rofstad EK, Mathiesen B, Kindem K, Galappathi K. Acidic extracellular pH promotes experimental metastasis of human melanoma cells in athymic nude mice. Cancer Res. 2006;66:6699–707. doi: 10.1158/0008-5472.CAN-06-0983. [DOI] [PubMed] [Google Scholar]
- 51.Swietach P, Wigfield S, Cobden P, et al. Tumor-associated carbonic anhydrase 9 spatially coordinates intracellular pH in three-dimensional multicellular growths. J Biol Chem. 2008;283:20473–83. doi: 10.1074/jbc.M801330200. [DOI] [PubMed] [Google Scholar]
- 52.Erler JT, Bennewith KL, Cox TR, et al. Hypoxia-induced lysyl oxidase is a critical mediator of bone marrow cell recruitment to form the premetastatic niche. Cancer Cell. 2009;15:35–44. doi: 10.1016/j.ccr.2008.11.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Erler JT, Bennewith KL, Nicolau M, et al. Lysyl oxidase is essential for hypoxia-induced metastasis. Nature. 2006;440:1222–6. doi: 10.1038/nature04695. [DOI] [PubMed] [Google Scholar]
- 54.Mole DR, Blancher C, Copley RR, et al. Genome-wide association of hypoxia-inducible factor (HIF)-1{alpha} and HIF-2{alpha} DNA binding with expression profiling of hypoxia-inducible transcripts. J Biol Chem. 2009;284:16767–75. doi: 10.1074/jbc.M901790200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kim W. The VHL tumor suppressor protein, HIF and cancer. J Cell Mol Med. 2009 [Google Scholar]
- 56.Boyd JM, Gallo GJ, Elangovan B, et al. Bik, a novel death-inducing protein shares a distinct sequence motif with Bcl-2 family proteins and interacts with viral and cellular survival-promoting proteins. Oncogene. 1995;11:1921–8. [PubMed] [Google Scholar]
- 57.Koong AC, Denko NC, Hudson KM, et al. Candidate genes for the hypoxic tumor phenotype. Cancer Res. 2000;60:883–7. [PubMed] [Google Scholar]
- 58.Ivanova AV, Ivanov SV, Danilkovitch-Miagkova A, et al. Regulation of STRA13 by the von Hippel-Lindau tumor suppressor protein, hypoxia, and the UBC9/ubiquitin proteasome degradation pathway. J Biol Chem. 2001;276:15306–15. doi: 10.1074/jbc.M010516200. [DOI] [PubMed] [Google Scholar]
- 59.Nishiwaki T, Daigo Y, Kawasoe T, et al. Isolation and mutational analysis of a novel human cDNA, DEC1 (deleted in esophageal cancer 1), derived from the tumor suppressor locus in 9q32. Genes Chromosomes Cancer. 2000;27:169–76. [PubMed] [Google Scholar]
- 60.Chi JT, Wang Z, Nuyten DS, et al. Gene expression programs in response to hypoxia: cell type specificity and prognostic significance in human cancers. PLoS Med. 2006;3:395–409. doi: 10.1371/journal.pmed.0030047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kim SJ, Rabbani ZN, Vollmer RT, et al. Carbonic anhydrase IX in early-stage non-small cell lung cancer. Clin Cancer Res. 2004;10:7925–33. doi: 10.1158/1078-0432.CCR-04-0636. [DOI] [PubMed] [Google Scholar]
- 62.Couvelard A, O’Toole D, Turley H, et al. Microvascular density and hypoxia-inducible factor pathway in pancreatic endocrine tumours: negative correlation of microvascular density and VEGF expression with tumour progression. Br J Cancer. 2005;92:94–101. doi: 10.1038/sj.bjc.6602245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Loncaster JA, Harris AL, Davidson SE, et al. Carbonic anhydrase (CA IX) expression, a potential new intrinsic marker of hypoxia: correlations with tumor oxygen measurements and prognosis in locally advanced carcinoma of the cervix. Cancer Res. 2001;61:6394–9. [PubMed] [Google Scholar]
- 64.Sowter HM, Raval RR, Moore JW, et al. Predominant role of hypoxia-inducible transcription factor (Hif)-1alpha versus Hif-2alpha in regulation of the transcriptional response to hypoxia. Cancer Res. 2003;63:6130–4. [PubMed] [Google Scholar]
- 65.Chen C, Pore N, Behrooz A, et al. Regulation of glut1 mRNA by hypoxia-inducible factor-1. Interaction between H-ras and hypoxia. J Biol Chem. 2001;276:9519–25. doi: 10.1074/jbc.M010144200. [DOI] [PubMed] [Google Scholar]
- 66.Hu CJ, Wang LY, Chodosh LA, et al. Differential roles of hypoxia-inducible factor 1alpha (HIF-1alpha) and HIF-2alpha in hypoxic gene regulation. Mol Cell Biol. 2003;23:9361–74. doi: 10.1128/MCB.23.24.9361-9374.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Dales JP, Garcia S, Meunier-Carpentier S, et al. Overexpression of hypoxia-inducible factor HIF-1alpha predicts early relapse in breast cancer: retrospective study in a series of 745 patients. Int J Cancer. 2005;116:734–9. doi: 10.1002/ijc.20984. [DOI] [PubMed] [Google Scholar]
- 68.Generali D, Berruti A, Brizzi MP, et al. Hypoxia-inducible factor-1alpha expression predicts a poor response to primary chemoendocrine therapy and disease-free survival in primary human breast cancer. Clin Cancer Res. 2006;12:4562–8. doi: 10.1158/1078-0432.CCR-05-2690. [DOI] [PubMed] [Google Scholar]
- 69.Yamamoto Y, Ibusuki M, Okumura Y, et al. Hypoxia-inducible factor 1alpha is closely linked to an aggressive phenotype in breast cancer. Breast Cancer Res Treat. 2008;110:465–75. doi: 10.1007/s10549-007-9742-1. [DOI] [PubMed] [Google Scholar]
- 70.Trastour C, Benizri E, Ettore F, et al. HIF-1alpha and CA IX staining in invasive breast carcinomas: prognosis and treatment outcome. Int J Cancer. 2007;120:1451–8. doi: 10.1002/ijc.22436. [DOI] [PubMed] [Google Scholar]
- 71.Bos R, Van Der Groep P, Greijer AE, et al. Levels of hypoxia-inducible factor-1alpha independently predict prognosis in patients with lymph node negative breast carcinoma. Cancer. 2003;97:1573–81. doi: 10.1002/cncr.11246. [DOI] [PubMed] [Google Scholar]
- 72.Schindl M, Schoppmann SF, Samonigg H, et al. Overexpression of hypoxia-inducible factor 1alpha is associated with an unfavorable prognosis in lymph node-positive breast cancer. Clin Cancer Res. 2002;8:1831–7. [PubMed] [Google Scholar]
- 73.Helczynska K, Larsson AM, Holmquist Mengelbier L, et al. Hypoxia-inducible factor-2alpha correlates to distant recurrence and poor outcome in invasive breast cancer. Cancer Res. 2008;68:9212–20. doi: 10.1158/0008-5472.CAN-08-1135. [DOI] [PubMed] [Google Scholar]
- 74.Gruber G, Greiner RH, Hlushchuk R, et al. Hypoxia-inducible factor 1 alpha in high-risk breast cancer: an independent prognostic parameter? Breast Cancer Res. 2004;6:R191–8. doi: 10.1186/bcr775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Chen G, Ray R, Dubik D, et al. The E1B 19K/Bcl-2-binding protein Nip3 is a dimeric mitochondrial protein that activates apoptosis. J Exp Med. 1997;186:1975–83. doi: 10.1084/jem.186.12.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Tan EY, Campo L, Han C, et al. BNIP3 as a progression marker in primary human breast cancer; opposing functions in in situ versus invasive cancer. Clin Cancer Res. 2007;13:467–74. doi: 10.1158/1078-0432.CCR-06-1466. [DOI] [PubMed] [Google Scholar]
- 77.Hutchison GJ, Valentine HR, Loncaster JA, et al. Hypoxia-inducible factor 1alpha expression as an intrinsic marker of hypoxia: correlation with tumor oxygen, pimonidazole measurements, and outcome in locally advanced carcinoma of the cervix. Clin Cancer Res. 2004;10:8405–12. doi: 10.1158/1078-0432.CCR-03-0135. [DOI] [PubMed] [Google Scholar]
- 78.Liao D, Corle C, Seagroves TN, et al. Hypoxia-inducible factor-1alpha is a key regulator of metastasis in a transgenic model of cancer initiation and progression. Cancer Res. 2007;67:563–72. doi: 10.1158/0008-5472.CAN-06-2701. [DOI] [PubMed] [Google Scholar]
- 79.Rofstad EK, Galappathi K, Mathiesen B, et al. Fluctuating and diffusion-limited hypoxia in hypoxia-induced metastasis. Clin Cancer Res. 2007;13:1971–8. doi: 10.1158/1078-0432.CCR-06-1967. [DOI] [PubMed] [Google Scholar]
- 80.Ginouves A, Ilc K, Macias N, et al. PHDs overactivation during chronic hypoxia “desensitizes” HIFalpha and protects cells from necrosis. Proc Natl Acad Sci USA. 2008;105:4745–50. doi: 10.1073/pnas.0705680105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Martinive P, Defresne F, Bouzin C, et al. Preconditioning of the tumor vasculature and tumor cells by intermittent hypoxia: implications for anticancer therapies. Cancer Res. 2006;66:11736–44. doi: 10.1158/0008-5472.CAN-06-2056. [DOI] [PubMed] [Google Scholar]
- 82.Willam C, Warnecke C, Schefold JC, et al. Inconsistent effects of acidosis on HIF-alpha protein and its target genes. Pflugers Arch. 2006;451:534–43. doi: 10.1007/s00424-005-1486-3. [DOI] [PubMed] [Google Scholar]
- 83.Iakovlev VV, Pintilie M, Morrison A, et al. Effect of distributional heterogeneity on the analysis of tumor hypoxia based on carbonic anhydrase IX. Lab Invest. 2007;87:1206–17. doi: 10.1038/labinvest.3700680. [DOI] [PubMed] [Google Scholar]
- 84.Vleugel MM, Greijer AE, Shvarts A, et al. Differential prognostic impact of hypoxia induced and diffuse HIF-1alpha expression in invasive breast cancer. J Clin Pathol. 2005;58:172–7. doi: 10.1136/jcp.2004.019885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Li YM, Zhou BP, Deng J, et al. A hypoxia-independent hypoxia-inducible factor-1 activation pathway induced by phosphatidylinositol-3 kinase/Akt in HER2 overexpressing cells. Cancer Res. 2005;65:3257–63. doi: 10.1158/0008-5472.CAN-04-1284. [DOI] [PubMed] [Google Scholar]
- 86.Wykoff CC, Beasley NJ, Watson PH, et al. Hypoxia-inducible expression of tumor-associated carbonic anhydrases. Cancer Res. 2000;60:7075–83. [PubMed] [Google Scholar]
- 87.Winter SC, Shah KA, Han C, et al. The relation between hypoxia-inducible factor (HIF)-1alpha and HIF-2alpha expression with anemia and outcome in surgically treated head and neck cancer. Cancer. 2006;107:757–66. doi: 10.1002/cncr.21983. [DOI] [PubMed] [Google Scholar]
- 88.Kaluz S, Kaluzova M, Chrastina A, et al. Lowered oxygen tension induces expression of the hypoxia marker MN/carbonic anhydrase IX in the absence of hypoxia-inducible factor 1 alpha stabilization: a role for phosphatidylinositol 3’-kinase. Cancer Res. 2002;62:4469–77. [PubMed] [Google Scholar]
- 89.Nordsmark M, Eriksen JG, Gebski V, et al. Differential risk assessments from five hypoxia specific assays: the basis for biologically adapted individualized radiotherapy in advanced head and neck cancer patients. Radiother Oncol. 2007;83:389–97. doi: 10.1016/j.radonc.2007.04.021. [DOI] [PubMed] [Google Scholar]
- 90.Sobhanifar S, Aquino-Parsons C, Stanbridge EJ, et al. Reduced expression of hypoxia-inducible factor-1alpha in perinecrotic regions of solid tumors. Cancer Res. 2005;65:7259–66. doi: 10.1158/0008-5472.CAN-04-4480. [DOI] [PubMed] [Google Scholar]
- 91.Ord JJ, Agrawal S, Thamboo TP, et al. An investigation into the prognostic significance of necrosis and hypoxia in high grade and invasive bladder cancer. J Urol. 2007;178:677–82. doi: 10.1016/j.juro.2007.03.112. [DOI] [PubMed] [Google Scholar]
- 92.McShane LM, Altman DG, Sauerbrei W, et al. REporting recommendations for tumour MARKer prognostic studies (REMARK) Br J Cancer. 2005;93:387–91. doi: 10.1038/sj.bjc.6602678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Van De Vijver MJ, He YD, Van’t Veer LJ, et al. A gene-expression signature as a predictor of survival in breast cancer. N Engl J Med. 2002;347:1999–2009. doi: 10.1056/NEJMoa021967. [DOI] [PubMed] [Google Scholar]
- 94.Wang Y, Klijn JG, Zhang Y, et al. Gene-expression profiles to predict distant metastasis of lymph-node-negative primary breast cancer. Lancet. 2005;365:671–9. doi: 10.1016/S0140-6736(05)17947-1. [DOI] [PubMed] [Google Scholar]
- 95.Seigneuric R, Starmans MH, Fung G, et al. Impact of supervised gene signatures of early hypoxia on patient survival. Radiother Oncol. 2007;83:374–82. doi: 10.1016/j.radonc.2007.05.002. [DOI] [PubMed] [Google Scholar]
- 96.Zhang C, Kawauchi J, Adachi MT, et al. Activation of JNK and transcriptional repressor ATF3/LRF1 through the IRE1/TRAF2 pathway is implicated in human vascular endothelial cell death by homocysteine. Biochem Biophys Res Commun. 2001;289:718–24. doi: 10.1006/bbrc.2001.6044. [DOI] [PubMed] [Google Scholar]
- 97.Kitamuro T, Takahashi K, Ogawa K, et al. Bach1 functions as a hypoxia-inducible repressor for the heme oxygenase-1 gene in human cells. J Biol Chem. 2003;278:9125–33. doi: 10.1074/jbc.M209939200. [DOI] [PubMed] [Google Scholar]
- 98.Nuyten DS, Kreike B, Hart AA, et al. Predicting a local recurrence after breast-conserving therapy by gene expression profiling. Breast Cancer Res. 2006;8:R62. doi: 10.1186/bcr1614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Chen JL, Lucas JE, Schroeder T, et al. The genomic analysis of lactic acidosis and acidosis response in human cancers. PLoS Genet. 2008;4:e1000293. doi: 10.1371/journal.pgen.1000293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Winter SC, Buffa FM, Silva P, et al. Relation of a hypoxia metagene derived from head and neck cancer to prognosis of multiple cancers. Cancer Res. 2007;67:3441–9. doi: 10.1158/0008-5472.CAN-06-3322. [DOI] [PubMed] [Google Scholar]
- 101.Hurwitz H, Fehrenbacher L, Novotny W, et al. Bevacizumab plus irinotecan, fluorouracil, and leucovorin for metastatic colorectal cancer. N Engl J Med. 2004;350:2335–42. doi: 10.1056/NEJMoa032691. [DOI] [PubMed] [Google Scholar]
- 102.Murat A, Migliavacca E, Hussain SF, et al. Modulation of angiogenic and inflammatory response in glioblastoma by hypoxia. PLoS One. 2009;4:e5947. doi: 10.1371/journal.pone.0005947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Walsh S, Gill C, O’Neill A, et al. Hypoxia increases normal prostate epithelial cell resistance to receptor-mediated apoptosis via AKT activation. Int J Cancer. 2009;124:1871–8. doi: 10.1002/ijc.24145. [DOI] [PubMed] [Google Scholar]
- 104.Calin GA, Croce CM. MicroRNA signatures in human cancers. Nat Rev Cancer. 2006;6:857–66. doi: 10.1038/nrc1997. [DOI] [PubMed] [Google Scholar]
- 105.Camps C, Buffa FM, Colella S, et al. hsa-miR-210 Is induced by hypoxia and is an independent prognostic factor in breast cancer. Clin Cancer Res. 2008;14:1340–8. doi: 10.1158/1078-0432.CCR-07-1755. [DOI] [PubMed] [Google Scholar]
- 106.Kulshreshtha R, Ferracin M, Wojcik SE, et al. A microRNA signature of hypoxia. Mol Cell Biol. 2007;27:1859–67. doi: 10.1128/MCB.01395-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Brazma A, Hingamp P, Quackenbush J, et al. Minimum information about a microarray experiment (MIAME)-toward standards for microarray data. Nat Genet. 2001;29:365–71. doi: 10.1038/ng1201-365. [DOI] [PubMed] [Google Scholar]
- 108.Shi L, Reid LH, Jones WD, et al. The MicroArray Quality Control (MAQC) project shows inter- and intraplatform reproducibility of gene expression measurements. Nat Biotechnol. 2006;24:1151–61. doi: 10.1038/nbt1239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Moeller BJ, Richardson RA, Dewhirst MW. Hypoxia and radiotherapy: opportunities for improved outcomes in cancer treatment. Cancer Metastasis Rev. 2007;26:241–8. doi: 10.1007/s10555-007-9056-0. [DOI] [PubMed] [Google Scholar]
- 110.Williams KJ, Telfer BA, Xenaki D, et al. Enhanced response to radiotherapy in tumours deficient in the function of hypoxia-inducible factor-1. Radiother Oncol. 2005;75:89–98. doi: 10.1016/j.radonc.2005.01.009. [DOI] [PubMed] [Google Scholar]
- 111.Dische S, Saunders M, Barrett A, et al. A randomised multicentre trial of CHART versus conventional radiotherapy in head and neck cancer. Radiother Oncol. 1997;44:123–36. doi: 10.1016/s0167-8140(97)00094-7. [DOI] [PubMed] [Google Scholar]
- 112.Koukourakis MI, Bentzen SM, Giatromanolaki A, et al. Endogenous markers of two separate hypoxia response pathways (hypoxia inducible factor 2 alpha and carbonic anhydrase 9) are associated with radiotherapy failure in head and neck cancer patients recruited in the CHART randomized trial. J Clin Oncol. 2006;24:727–35. doi: 10.1200/JCO.2005.02.7474. [DOI] [PubMed] [Google Scholar]
- 113.Zhu Y, Denhardt DT, Cao H, et al. Hypoxia upregulates osteopontin expression in NIH-3T3 cells via a Ras-activated enhancer. Oncogene. 2005;24:6555–63. doi: 10.1038/sj.onc.1208800. [DOI] [PubMed] [Google Scholar]
- 114.Fyfe GA, Hurwitz H, Fehrenbacher L, et al. Bevacizumab plus irinotecan/5-FU/leucovorin for treatment of metastatic colorectal cancer results in survival benefit in all pre-specified patient subgroups. J Clin Oncol. 2004;22:3617. [Google Scholar]
- 115.Minn H, Gronroos TJ, Komar G, et al. Imaging of tumor hypoxia to predict treatment sensitivity. Curr Pharm Des. 2008;14:2932–42. doi: 10.2174/138161208786404380. [DOI] [PubMed] [Google Scholar]