

Abstract
NF-κB plays central roles in regulation of diverse biological processes, including innate and adaptive immunity and inflammation. HSV-1 is the archetypal member of the alphaherpesviruses, with a large genome encoding over 80 viral proteins, many of which are involved in virus-host interactions and show immune modulatory capabilities. In this study, we demonstrated that the HSV-1 ICP0 protein, a viral E3 ubiquitin ligase, was shown to significantly suppress tumor necrosis factor alpha (TNF-α)-mediated NF-κB activation. ICP0 was demonstrated to bind to the NF-κB subunits p65 and p50 by coimmunoprecipitation analysis. ICP0 bound to the Rel homology domain (RHD) of p65. Fluorescence microscopy demonstrated that ICP0 abolished nuclear translocation of p65 upon TNF-α stimulation. Also, ICP0 degraded p50 via its E3 ubiquitin ligase activity. The RING finger (RF) domain mutant ICP0 (ICP0-RF) lost its ability to inhibit TNF-α-mediated NF-κB activation and p65 nuclear translocation and degrade p50. Notably, the RF domain of ICP0 was sufficient to interact with p50 and abolish NF-κB reporter gene activity. Here, it is for the first time shown that HSV-1 ICP0 interacts with p65 and p50, degrades p50 through the ubiquitin-proteasome pathway, and prevents NF-κB-dependent gene expression, which may contribute to immune evasion and pathogenesis of HSV-1.
INTRODUCTION
The innate immune response to viruses involves activation of pattern recognition receptors (1–3) and transcriptional induction of type I interferons (IFNs) and proinflammatory cytokines (4). The transcription factor NF-κB plays a pivotal role in many cellular events, such as innate and adaptive immunity and inflammation (5–8).
The mammalian NF-κB family comprises five members: p65/RelA, RelB, p50/NF-κB1, p52/NF-κB2, and c-Rel. All family members share a structurally conserved N-terminal region, named the Rel homology domain (RHD), which is critical for protein dimerization, DNA binding, interaction with IκB (an inhibitor of NF-κB), and nuclear translocation (9, 10). Rel proteins (p65/RelA, RelB, c-Rel) contain a C-terminal transactivation domain, which is lacking in p50 and p52. The predominant form of NF-κB is a heterodimer of p65 and p50 subunits (11, 12).
Tumor necrosis factor alpha (TNF-α) is a multifunctional proinflammatory cytokine involved in protecting the host from pathogen infections by induction and regulation of host innate and adaptive immune responses (13). The pathway of TNF-α-induced NF-κB activation is that TNF-α binds to its receptor, TNFR1, resulting in recruitment of the adaptor protein TNF receptor death domain (TRADD), and then TRADD recruits TNFR-associated factor 2 (TRAF2) and receptor-interacting protein 1 (RIP1) to the receptor complex and TRAF2 mediates K63-linked polyubiquitination of RIP1. Further, ubiquitinated RIP1 further recruits TGF-β-activated kinase 1 (TAK1) and subsequently activates the IκB kinase (IKK) complex, leading to phosphorylation and degradation of IκBα and, at last, activation of NF-κB (14).
Herpes simplex virus 1 (HSV-1) is a large DNA virus known to encode several gene products that enable viral evasion of the host innate immune response (15, 16). Several studies have shown that HSV-1 encodes proteins to disturb the NF-κB pathway. ICP27, an immediate early gene product of HSV-1, has been shown to antagonize NF-κB signaling (17). The γ134.5 protein, an HSV-1-encoded late-gene product, inhibits activation of NF-κB in CD8+ dendritic cells (DCs) (18). Vhs, a tegument protein, blocks the early replication-independent activation of NF-κB in HSV-1-infected DCs (19). HSV-1 VP16 blocks the activation of the NF-κB promoter induced by SeV or TNF-α treatment and expression of NF-κB-dependent genes through interaction with p65 (20). UL42, a DNA polymerase processivity factor of HSV-1, is a novel antagonism of the canonical NF-κB signaling pathway (21).
Protein ubiquitination plays an essential role in the positive and negative regulation of the TNF-α-mediated NF-κB signal transduction pathway (22). Ubiquitin contains seven lysines, which can be attached to another ubiquitin in a highly processive reaction to form a polyubiquitin chain. Typically, two types of linkages of polyubiquitin chains, K48 and K63, have been extensively investigated so far. In most cases, K48-linked polyubiquitin chains target their substrates for proteasome-dependent degradation (23).
HSV-1 ICP0 is a multifunctional and immediate early protein that plays a pivotal role during lytic and latent infections (24–26). ICP0 has an E3 ubiquitin ligase activity that promotes degradation of certain host proteins, and the interaction of ICP0 with the ubiquitin-proteasome system is well documented. For example, it mediates the degradation of several cellular proteins (27, 28), induces conjugation of ubiquitin (29), and sequesters proteasomes in the nucleus (30). The RING finger (RF) domain is required for many of ICP0's known functions and has E3 ubiquitin ligase activity (31–33). Several studies have demonstrated that HSV-1 infection activates interferon signaling in various cell types, and ICP0 is responsible for dampening the production of IFN-β and interferon-stimulated genes (ISGs) during infection (34–39). ICP0 also inhibited Toll-like receptor (TLR)-driven inflammatory cytokine response following viral infection by targeting the TLR2/NF-κB pathway (40, 41).
HSV-1 infection can induce proinflammatory and inflammatory cytokines, including TNF-α, one of the stimuli in the NF-κB signaling pathway. However, the role of HSV-1 in the TNF-α-mediated activation of NF-κB has yet to be addressed. In this study, we demonstrated that ectopic expression of ICP0 significantly downregulated TNF-α-induced NF-κB promoter activity, and the RF domain of ICP0 was indispensable for the inhibitory activity. Additionally, ICP0 bound to the RHD of p65 and abolished the nuclear translocation of p65 upon TNF-α-stimulation. We also found that ICP0 degraded p50 via its E3 ubiquitin ligase activity. For the first time, we found that ICP0 interacted with p50 and promoted it for degradation. Notably, the RF domain of the ICP0 was sufficient for interacting with p50 and preventing reporter gene activity. Our results expanded our knowledge about the molecular mechanisms by which HSV-1 counteracts the host innate antiviral immunity to ensure its replication and spread.
MATERIALS AND METHODS
Cells, viruses, antibodies, reagents, and cytokines.
HEK 293T cells, Vero cells, and U2OS cells were grown in Dulbecco's modified Eagle medium (DMEM) (Gibco-BRL) supplemented with 10% fetal bovine serum (FBS) and 100 U/ml of penicillin and streptomycin. HeLa cells were maintained in Eagle's minimum essential medium (MEM) (Gibco-BRL) supplemented with 10% FBS.
The wild-type (WT) HSV-1 F strain was propagated in Vero cells, and the titer was determined as described previously (42). The HSV-1 F strain ICP0-RF mutant virus (R8513) was kindly provided by Haidong Gu and Bernard Roizman. It was propagated in U2OS cells, and the titer was determined as described previously (43).
Mouse anti-Myc (isotype IgG1), anti-Flag (isotype IgG2b), and antihemagglutinin (anti-HA) (isotype IgG2b) monoclonal antibodies (MAbs) were purchased from ABmart (Shanghai, China). Mouse monoclonal IgG1 and IgG2b isotype control antibodies were purchased from eBioscience Inc. (San Diego, CA). Rabbit polyclonal anti-p65 and rabbit polyclonal anti-p50 were purchased from Proteintech (Wuhan, China). Rabbit anti-ICP0 polyclonal antibody (pAb) was purchased from GL Biochem, and mouse anti-β-actin MAb was purchased from Santa Cruz Biotechnology (Santa Cruz, CA). The proteasome inhibitor MG132 was purchased from Sigma (St. Louis, MO). Human TNF-α was purchased from BioVision (San Francisco, CA). Lipofectamine 2000 (Lipo2000) was purchased from Invitrogen.
Plasmid construction.
All enzymes, except for the T4 DNA ligase (New England BioLabs, Massachusetts), used for cloning procedures were purchased from TaKaRa (Dalian, China). To construct ICP0-Flag and ICP0-Myc, the open reading frame (ORF) of ICP0 was amplified from plasmid ICP0-enhanced yellow fluorescent protein (EYFP) as previously described (44) and cloned into the BamHI and EcoRI sites of the pCMV-Flag and pCMV-Myc vectors (Beyotime, Shanghai, China). To construct Flag-p65 RHD, the ORF of the p65 RHD was amplified from plasmid pFlag-p65 and cloned into the BamHI and EcoRI sites of the pCMV-Flag. Commercial reporter plasmids include NF-κB-Luc (Stratagene, La Jolla, CA) and pRL-TK plasmid (Promega). Gift plasmids include the following: pEFIRES-HA-Ub, pEFIRES-HA-Ub(K48), and pEFIRES-HA-Ub(K63), obtained from Yossi Yarden (Weizmann Institute, Israel); pcDNA3-NIK (where NIK is NF-κB-inducing kinase) (45); pFlag-p65 (46); pFlag-p50 (47); pFlag-IKKβ (48); pHA-IKKα (49); pGL3-p-IL-6-Luc (where IL-6 is interleukin 6); and pXP2-p-IL-8-Luc (50).
Transfection and dual-luciferase reporter (DLR) assay.
HEK 293T cells were cotransfected with reporter plasmid NF-κB-Luc and internal control plasmid pRL-TK, with or without expression plasmids as indicated by standard calcium phosphate precipitation (51, 52). At 24 h posttransfection, cells were mock treated or treated with TNF-α (10 ng/ml) for 6 h. To detect NF-κB activity under HSV-1 infection, HEK 293T cells were cotransfected with reporter plasmid NF-κB-Luc and internal control plasmid pRL-TK by Lipo2000 according to the manufacturer's instructions. At 24 h posttransfection, the cells were infected with HSV-1 as indicated. Luciferase assays were performed as previously described with a luciferase assay kit (Promega, Madison, WI) (53).
RNA isolation, semiquantitative RT-PCR, and quantification of gel image.
Total RNA was extracted from HEK 293T cells with TRIzol (Invitrogen, California) according to the manufacturer's manual. Samples were digested with DNase I and subjected to reverse transcription. The cDNA was used as a template for semiquantitative PCR to investigate the accumulation of human IL-6 and IL-8 mRNA and viral mRNA. The detailed protocols have been previously described (53). The primer sets used for each gene of interest were as follows: GAPDH, 5′-TGACCTCAACTACATGGTTTACATGT-3′ and 5′-AGGGATCTCGCTCCTGGAA-3′; IL-6, 5′-GGATTCAATGAGGAGACTTGCC-3′ and 5′-ACAGCTCTGGCTTGTTCCTCAC-3′; IL-8, 5′-CAACACAGAAATTATTGTAAAGCTTTCT-3′ and 5′-GAATTCTCAGCCCTCTTCAAAAA-3′; ICP0, 5′-GGTCGCCCTGTCGCCTTA-3′ and 5′-GGTCGCCATGTTTCCCGT-3′; UL42, 5′-CCAACGCCAAGACGGTGTA-3′ and 5′-GGGGGTCGTGAGGAAGAAC-3′; UL46, 5′-CGAGACGGTGAGCGAGGA-3′ and 5′-GGGGGGAAAATAGGGCG-3′.
Coimmunoprecipitation assay and Western blot analysis.
Coimmunoprecipitation (co-IP) assays were performed as previously described (42). Briefly, HEK 293T cells were cotransfected with 10 μg of each of the indicated expression plasmids carrying Flag, Myc, or HA tags. Transfected cells were harvested at 36 h posttransfection and lysed on ice with 700 μl of lysis buffer. For virus infection, the cells were then infected with HSV-1 for an additional 16 h at 20 h posttransfection and lysed on ice with 700 μl of lysis buffer. For each IP, a 0.5-ml aliquot of lysate was incubated with 0.5 μg of the anti-HA MAb, anti-Myc MAb, or anti-Flag MAb or nonspecific mouse monoclonal antibody (IgG isotype matched with anti-Myc MAb; IgG2b isotype matched with anti-HA and anti-Flag MAbs) and 30 μl of a 1:1 slurry of protein A/G Plus-agarose (Santa Cruz, California) for at least 4 h or overnight at 4°C. The beads were washed four times with 1 ml of lysis buffer containing 500 mM NaCl and then subjected to Western blot (WB) analysis. All co-IP assays were repeated at least two times, similar data were obtained, and a typical blot was shown.
WB analysis was performed as previously described (42). Briefly, whole-cell extracts were subjected to 10% SDS-PAGE and transferred to polyvinylidene difluoride (PVDF) membranes, followed by blocking with 5% nonfat milk in Tris-buffered saline–Tween (TBST) and being probed with the indicated primary antibodies at 37°C for 2 h. After being washed with TBST, the membrane was incubated with alkaline phosphatase (AP)-conjugated goat anti-rabbit IgG or goat anti-mouse IgG. Protein bands specific to the antibody were developed by 5-bromo-4-chloro-3-indolylphosphate (BCIP)–nitroblue tetrazolium (NBT) and terminated by distilled water.
Immunofluorescence assay.
Immunofluorescence assays were performed as described previously (54). In brief, HeLa cells were transfected with the indicated plasmids for 24 h and then fixed in 4% paraformaldehyde. Cells were incubated with rabbit anti-p65 pAb (diluted 1:500) or with mouse anti-Flag MAb and anti-Myc MAb (diluted 1:2,000), followed by incubation with tetramethyl rhodamine isocyanate (TRITC)-conjugated goat anti-rabbit IgG (Pierce) and fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG (Sigma-Aldrich). After each incubation step, cells were washed extensively with phosphate-buffered saline (PBS). Samples were analyzed with fluorescence microscopy (Zeiss, Germany).
RESULTS
HSV-1 ICP0 dampens TNF-α-mediated NF-κB activation.
The canonical NF-κB signal pathway can be initiated by various stimuli, including TNF-α, a key cytokine regulating immune functions as well as inflammatory responses. In order to verify whether ICP0 affects NF-κB activity, we performed dual-luciferase reporter gene assays with cells treated with TNF-α. Flag-tagged ICP0 was coexpressed in HEK 293T cells in the presence of NF-κB-Luc reporter genes. TNF-α treatment resulted in strong induction of NF-κB activity (Fig. 1A). In contrast, ectopic expression of ICP0 significantly inhibited TNF-α-mediated activation of NF-κB promoter activity (Fig. 1A). Additionally, ICP0 inhibited NF-κB promoter activity in a dose-dependent manner (Fig. 1B).
Fig 1.
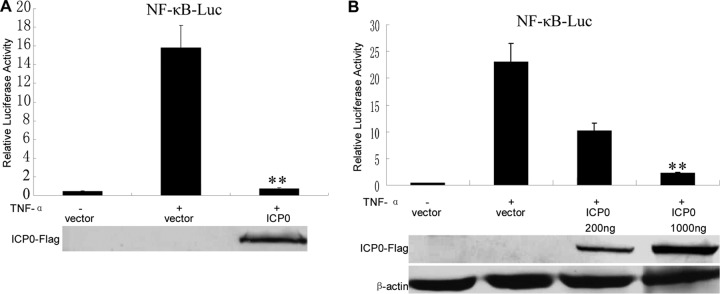
HSV-1 ICP0 inhibits TNF-α-mediated NF-κB activation. (A) HEK 293T cells were transfected with 500 ng of NF-κB promoter reporter plasmid NF-κB-luc, together with Renilla luciferase plasmid pRL-TK (50 ng) and pCMV-Flag control vector or plasmids encoding the indicated viral proteins (500 ng). At 24 h posttransfection, cells were treated with or without 10 ng/ml recombinant human TNF-α and incubated for an additional 6 h, followed by cell lysis. NF-κB-driven luciferase activity was determined by a dual-luciferase assay. (B) As described for panel A, except an increased amount of ICP0-Flag expression plasmid as indicated was used. The expression of ICP0 was analyzed by Western blotting using anti-Flag and anti-β-actin (as a control) monoclonal antibodies. The data represent means plus standard deviations for three replicates. Statistical analysis was performed using Student's t test. **, P < 0.01.
The RF domain of ICP0 is indispensable for the inhibition of TNF-α-mediated NF-κB activation.
ICP0 bears a C3H4 zinc RF domain in the N terminus and is a viral E3 ubiquitin ligase (Fig. 2A). The RF domain is conservative among alphaherpesvirus, and the role of ICP0 to suppress the host innate immunity is RF domain dependent (35). To determine whether the RF domain is also responsible for ICP0 inhibiting TNF-α-mediated activation of NF-κB, an RF domain mutant (ICP0-ΔRF) expression plasmid was generated by deletion of the amino acids (aa) 116 to 156 (Fig. 2A). We also constructed the ICP0(1-241aa) expression plasmid, which encompasses the E3 ubiquitin ligase domain of ICP0 (Fig. 2A). These two plasmids were transfected into HEK 293T cells and examined for their ability to inhibit TNF-α-induced NF-κB activation. In agreement with Fig. 1, WT ICP0 and ICP0(1-241aa) strongly inhibited TNF-α-induced NF-κB activation. However, ICP0-ΔRF failed to inhibit TNF-α-induced NF-κB activity (Fig. 2B). Additionally, ICP0(1-241aa) inhibited NF-κB promoter activity in a dose-dependent manner (Fig. 2C), but an increased amount of ICP0-ΔRF expression plasmid had no effect on NF-κB promoter activity (Fig. 2D). To test the role of the RF domain of ICP0 in inhibition of NF-κB in HSV-1-infected cells, we used WT HSV-1 and ICP0-RF mutant virus R8513 to infect HEK 293T cells at a multiplicity of infection (MOI) of 10. ICP0-RF mutant virus R8513 activated NF-κB reporter gene expression similarly to WT HSV-1 at 2 and 4 h (Fig. 2E), but it activated NF-κB reporter gene expression to higher levels than WT HSV-1 at 6 and 8 h (Fig. 2E). Taken together, these results suggested that the RF domain was critical for ICP0 to attenuate the NF-κB activation.
Fig 2.

The RF domain of ICP0 is indispensable for its inhibition of TNF-α-mediated NF-κB activation. (A) Schematic representation of the wild type, the RF deletion mutant of ICP0, and the N-terminal 241 aa of ICP0; (B) HEK 293T cells were cotransfected with pRL-TK plasmid and NF-κB-Luc reporter plasmid along with control vector or plasmids encoding ICP0 WT, the ICP0(1-241aa) mutant, or the ICP0-ΔRF mutant. At 24 h posttransfection, cells were mock treated or treated with recombinant human TNF-α (10 ng/ml), and luciferase activity was analyzed as described for Fig. 1A. (C and D) As described for panel B, except an increased amount of ICP0(1-241aa) and ICP0-ΔRF mutant expression plasmids as indicated was used. The expression of ICP0 and ICP0(1-241aa) and ICP0-ΔRF mutants was analyzed by Western blotting using anti-Flag, anti-Myc, and anti-β-actin (as a control) monoclonal antibodies. (E) HEK 293T cells were cotransfected with pRL-TK plasmid and NF-κB-Luc reporter plasmid by Lipo2000; at 24 h posttransfection, cells were infected with WT HSV-1 or ICP0-RF mutant virus R8513 at an MOI of 10. The infected cells were collected at the indicated time after infection, and luciferase activity was analyzed as described for Fig. 1A. The data represent means plus standard deviations for three replicates. Statistical analysis was performed using Student's t test. **, P < 0.01, *, P < 0.05.
ICP0 prevents the production of NF-κB-regulated proinflammatory cytokines.
To determine whether ICP0 was required to downregulate NF-κB-regulated proinflammatory cytokines, IL-6 and IL-8 reporter assays were performed using a reporter with an IL-6 or IL-8 promoter driving the expression of a luciferase gene. HEK 293T cells were treated with TNF-α, and the luciferase activities in both p-IL-6-Luc- and p-IL-8-Luc-transfected cells were unaffected by the control vector. However, ICP0 and ICP0(1-241aa) significantly inhibited IL-6 and IL-8 luciferase activity, but ICP0-ΔRF did not (Fig. 3A and B). To investigate whether the RF domain was important for inhibiting proinflammatory cytokine expression during HSV-1 infection, semiquantitative PCR was performed to measure the IL-6 and IL-8 mRNA expression in HEK 293T cells infected with WT HSV-1 or RF mutant virus R8513 at an MOI of 5. TNF-α induced a high level of IL-6 and IL-8 mRNA; however, WT HSV-1 infection induced only a trace amount of IL-6 and IL-8 mRNA. The RF mutant viruses R8513 induced significantly higher levels of IL-6 and IL-8 mRNA than WT HSV-1 did (Fig. 3C). And the accumulation of viral mRNAs of WT HSV-1 was similar to that of R8513 (Fig. 3C). Taken together, these results suggested that the inhibitory function of ICP0 on NF-κB-targeted proinflammatory genes was RF domain dependent.
Fig 3.

ICP0 inhibits the NF-κB-mediated proinflammatory cytokine expression. (A and B) HEK 293T cells were cotransfected with pCMV-Myc control vector, ICP0-Myc, ICP0(1-241aa)-Myc, or ICP0-ΔRF-Myc expression plasmid and p-x-Luc (where x is IL-6 or IL-8, as indicated) along with pRL-TK as an internal control. At 24 h posttransfection, the cells were treated with TNF-α (10 ng/ml) for 4 h, and luciferase activity was measured. (C) HEK 293T cells were infected with WT HSV-1 and ICP0-RF mutant virus R8513 at an MOI of 5; at 12 h postinfection, cells were mock treated or treated with recombinant human TNF-α (10 ng/ml) for 4 h. Semiquantitative RT-PCR analysis was then performed to detect the mRNA level of IL-6, IL-8, ICP0, UL42, and UL46. The data represent means plus standard deviations for three replicates. Statistical analysis was performed using Student's t test. **, P < 0.01.
ICP0 inhibits the NF-κB signaling pathway at the p65 level.
To determine at what level in the pathway ICP0 blocked NF-κB activity, increasing amounts of expression plasmids of Flag-tagged ICP0 and the canonical NF-κB signaling pathway components, including TRADD, TRAF2, TAK1, RIP1, NIK, IκB kinase alpha (IKKα), IKKβ, and p65, were cotransfected into HEK 293T cells. All expression constructs of the signaling pathway components resulted in a 10- to 100-fold induction of the NF-κB-Luc reporter activity (Fig. 4A to H). NF-κB promoter activation driven by all of the expression constructs was inhibited by ICP0 in a dose-dependent manner (Fig. 4A to H). The activation driven by TRADD, TRAF2, RIP1, TAK1, NIK, IKKα, and IKKβ was inhibited more than 80% (Fig. 4A to G), and the activation driven by p65 was inhibited up to 60% (Fig. 4H). These results indicated that ICP0 inhibits the NF-κB signaling at or downstream of p65.
Fig 4.

ICP0 targets p65 to inhibit NF-κB signaling. The HEK 293T cells were cotransfected with NF-κB-Luc, pRL-TK, and TRADD (A), TRAF2 (B), TAK1 (C), RIP1 (D), NIK (E), IKKα (F), IKKβ (G), and p65 (H) expression plasmids along with the indicated amount of ICP0 expression plasmid. Luciferase activity was analyzed as described for Fig. 1A. Cell lysates were analyzed by Western blotting with tag-specific Abs (bottom) to detect the expression of Myc-TRADD, Flag-TRAF2, Flag-TAK1, Flag-RIP1, HA-IKKα, Flag-IKKβ, and Flag-p65. The ICP0 expression was detected by mouse anti-Flag MAb. Western blotting of β-actin was used to verify equal loading of protein in each lane. The data represent means plus standard deviations for three replicates. Statistical analysis was performed using Student's t test. **, P < 0.01.
ICP0 interacts with endogenous p65 and p50.
In order to clarify the molecular mechanism of ICP0 to suppress NF-κB activation, the potential interaction between ICP0 and the NF-κB subunits p65 and p50 was investigated. ICP0-Myc and either Flag-p65 or Flag-p50 expression plasmids were cotransfected into HEK 293T cells, and co-IP/WB analysis was performed with anti-Myc and anti-Flag MAbs. Both p65 and p50 were efficiently co-IPed with ICP0 by anti-Myc MAbs (Fig. 5A and D). We further investigated the interaction between ICP0 and endogenous p65 or p50. ICP0-Flag expression plasmid was transfected into HEK 293T cells, and at 24 h posttransfection, the cells were treated with or without 10 ng/ml TNF-α for 8 h. Both endogenous p65 and p50 proteins were efficiently co-IPed with ICP0 by anti-Flag MAbs (Fig. 5B, C, E, and F) in the presence of TNF-α or not. Moreover, ICP0 interacts with p65 and p50 during viral infection. In WT HSV-1-infected cells, both p65 and p50 proteins were easily IPed by ICP0 using anti-ICP0 pAb (Fig. 5G and H, respectively) but not by control antibody IgG (Fig. 5G and H). Taken together, these results demonstrated that ICP0 interacts with endogenous p65 and p50 in the context of viral infection.
Fig 5.

ICP0 interacts with p65 and p50. The HEK 293T cells were cotransfected with ICP0-Myc and Flag-p65 expression plasmids (A) and ICP0-Myc and Flag-p50 expression plasmids (D). At 36 h after transfection, cells were harvested and lysed; the samples were then subjected to IP assays using anti-Myc MAb (IP:Myc) or nonspecific mouse monoclonal antibody (IgG). IP samples were separated by 10% SDS-PAGE, and proteins were transferred onto a PVDF membrane. WB were probed with the indicated Ab. (B, C, E, F) HEK 293T cells were transfected with Flag-ICP0 expression plasmid. At 24 h posttransfection, cells were mock treated (B and E) or treated (C and F) with TNF-α (10 ng/ml) and incubated for an additional 8 h, cells were harvested and lysed, and the samples were then subjected to IP assays using anti-Flag MAb (IP:Flag) or nonspecific mouse monoclonal antibody (IgG2b). HEK 293T cells were infected with WT HSV-1 at an MOI of 10 for 16 h. The cells were then lysed, and the extracts were subjected to IP using anti-ICP0 pAb (IP:ICP0) or control IgG. Precipitates were analyzed by Western blotting (G and H).
ICP0 degrades p50 through the ubiquitin-proteasome pathway.
ICP0 is an E3 ubiquitin ligase and induces the proteasome-dependent degradation of several cellular proteins and the accumulation of colocalizing conjugated ubiquitin in vivo (31). ICP0 interacted with p65 and p50, so we hypothesized that ICP0 could degrade p65 and p50 through the ubiquitin-proteasome pathway.
To test this, we coexpressed Flag-p65 or Flag-p50 with increasing amounts of ICP0-Myc plasmids. Expression of increasing amounts of Myc-ICP0 had no influence on p65 (Fig. 6A). However, coexpression of Myc-ICP0 with Flag-p50 resulted in a lower abundance of p50 in a dose-dependent manner (Fig. 6B). Next, we investigated whether ICP0-mediated p50 degradation was RF domain dependent. Coexpression of Myc-ICP0-ΔRF with Flag-p50 did not influence the abundance of p50 (Fig. 6C), suggesting that the RF domain is required for ICP0 to regulate the turnover of p50. Interestingly, the proteasome inhibitor MG132 blocked ICP0-mediated degradation of p50 (Fig. 6D). ICP0 also degraded endogenous p50 (Fig. 6E, F, and G). To investigate whether endogenous p50 was degraded during viral infection, HEK 293T cells were infected with WT HSV-1 or ICP0-RF mutant HSV-1 R8513 at an MOI of 1 and 10 for 16 h. The endogenous p50 was significantly degraded during WT HSV-1 infection at an MOI of 10 (Fig. 6H), and the proteasome inhibitor MG132 blocked the degradation of p50 (Fig. 6I). However, the ICP0-RF mutant HSV-1 could not degrade p50 (Fig. 6J). Taken together, all these results suggest that ICP0 degrades p50 via the RF domain-dependent mechanism.
Fig 6.

ICP0 degrades p50 through the ubiquitin-proteasome pathway. (A and B) HEK 293T cells were cotransfected with Flag-p65 or Flag-p50 expression plasmid and with increased concentrations of expression vector for ICP0-Myc. (C) HEK 293T cells transfected with Flag-p50 expression plasmid and increasing concentrations of expression vector for ICP0-ΔRF-Myc. (D) HEK 293T cells transfected with Flag-p50 expression plasmid and increasing concentrations of the expression vector for ICP0-Myc and cultured with MG132. (E and F) HEK 293T cells transfected with increasing concentrations of expression vectors for ICP0-Flag (E) or ICP0-ΔRF-Myc (F). (G) HEK 293T cells transfected with increasing concentrations of the expression vector for ICP0-Flag and cultured with MG132. (H and J) HEK 293T cells were infected with WT HSV-1 or ICP0-RF mutant HSV-1 at an MOI of 1 and 10 for 16 h. (I) HEK 293T cells were infected with WT HSV-1 strain F at an MOI of 1 and 10 and cultured with MG132 for 1 h. WB was probed with the indicated Ab.
p50 is targeted for K48-linked ubiquitination by ICP0.
The aforementioned results suggested that ICP0 degrades p50 through the ubiquitin-proteasome pathway. So we hypothesized that ICP0 could target p50 by K48-linked ubiquitination. To test this, Flag-p50 was coexpressed with HA-tagged Ub(WT), Ub(K48), or Ub(K63) into HEK 293T cells. At 24 h posttransfection, the cells were infected with WT HSV-1 or ICP0-RF mutant virus R8513. Polyubiquitinated forms of p50 were detected during WT HSV-1 infection when expressed with Ub(WT) (Fig. 7A, first lane). Notably, ICP0 catalyzed p50 polyubiquitination with expression of Ub(K48) (Fig. 7A, third lane) but not Ub(K63) (Fig. 7A, fifth lane).
Fig 7.
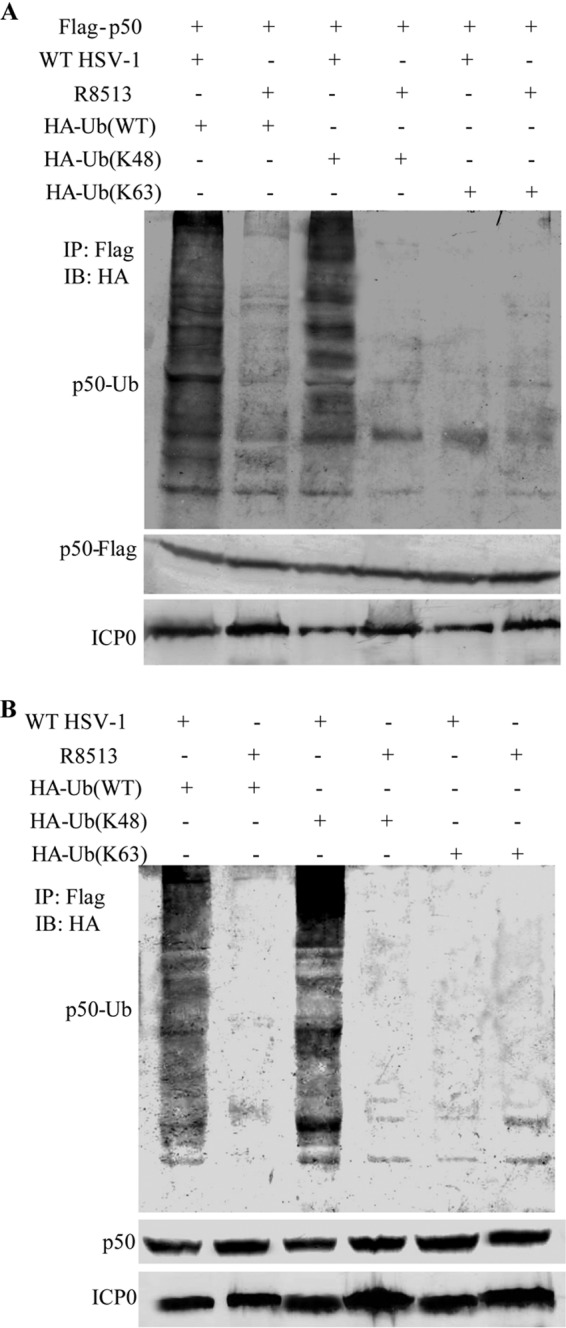
ICP0 targets p50 for K48-linked ubiquitination. ICP0 catalyzes K48-linked ubiquitination to degrade p50. (A) HEK 293T cells cotransfected with p50-Flag and HA-Ub(WT), HA-Ub(K48), or HA-Ub(K63). At 24 h posttransfection, the cells were infected with WT HSV-1 or ICP0-RF mutant virus R8513, followed by IP of lysates with anti-Flag and WB analysis with anti-HA. (B) HEK 293T cells transfected with HA-Ub(WT), HA-Ub(K48), or HA-Ub(K63). At 24 h posttransfection, the cells were infected with WT HSV-1 or ICP0-RF mutant virus R8513, followed by IP of lysates with anti-Flag and WB analysis with anti-HA.
To investigate whether ICP0 degraded endogenous p50 by K48-linked ubiquitination during viral infection, the HEK 293T cells were infected with WT HSV-1 or ICP0-RF mutant virus R8513 at 24 h posttransfection of plasmid HA-tagged Ub(WT), Ub(K48), or Ub(K63). Polyubiquitinated forms of p50 were detected during WT HSV-1 infection with expression of Ub(WT) (Fig. 7B, first lane). Notably, ICP0 catalyzed p50 polyubiquitination with expression of Ub(K48) (Fig. 7B, third lane) but not Ub(K63) (Fig. 7B, fifth lane). Taken together, these results demonstrated that ICP0-mediated K48-linked ubiquitination promotes proteasomal degradation of p50 in an RF domain-dependent manner.
ICP0 interacts with the RHD of p65 and prevents nuclear translocation of p65.
To determine if ICP0 protein bound to the N-terminal RHD of p65, which is responsible for dimerization, DNA binding, and nuclear import (55), the fragment of p65 spanning amino acids 21 to 186 (the p65 RHD) was constructed. ICP0-Myc and Flag-p65 RHD expression plasmids were cotransfected into HEK 293T cells to investigate their interaction. As a result, ICP0 was efficiently co-IPed with the p65 RHD (Fig. 8A). To further determine their interaction during viral infection, the HEK 293T cells were infected with WT HSV-1 at 24 h posttransfection of the Flag-p65 RHD expression plasmid. As a result, the Flag-p65 RHD was efficiently co-IPed with ICP0 (Fig. 8B). Taken together, these results demonstrated that ICP0 interacts with the p65 RHD during viral infection.
Fig 8.

ICP0 interacts with the RHD of p65 and blocks TNF-α-induced p65 nuclear translocation. (A) The HEK 293T cells were cotransfected with ICP0-Myc and Flag-p65 RHD expression plasmids. At 36 h after transfection, cells were harvested and lysed, and the samples were then subjected to IP assays using anti-Flag MAb (IP:Flag) or nonspecific mouse monoclonal antibody (IgG2b). WB were probed with the indicated Ab. (B) The HEK 293T cells were transfected with Flag-p65 RHD expression plasmid. At 24 h after transfection, the cells were infected with WT HSV-1 at an MOI of 10; after 16 h postinfection, cells were harvested and lysed, and the samples were then subjected to IP assays using anti-Flag MAb (IP:Flag) or nonspecific mouse monoclonal antibody (IgG2b). WB were probed with the indicated Ab. (C) HeLa cells were transfected with control vector, Flag-tagged ICP0 WT, or Myc-tagged ICP0-ΔRF mutant expression plasmid. At 24 h posttransfection, cells were treated with TNF-α (10 ng/ml) or mock treated for 30 min as indicated. Cells were stained with mouse anti-Flag or mouse anti-Myc MAb and rabbit anti-p65 pAb. FITC-conjugated goat anti-rabbit (green) and TRITC-conjugated goat anti-mouse (red) were used as the secondary antibodies. Cell nuclei (blue) were stained with Hoechst 33258. The images were obtained by fluorescence microscopy using a 40× objective.
Nuclear translocation of p65 is crucial for the transcription of NF-κB. Immunofluorescence was carried out to investigate whether ICP0 prevented the nuclear translocation of p65 (Fig. 8C). In mock-infected HeLa cells, p65 localized exclusively to the cytoplasm; however, p65 translocated into the nuclei in TNF-α-stimulated cells. Ectopic expression of ICP0 prevented the nuclear translocation of p65 induced by TNF-α, whereas the ICP0-ΔRF mutant did not (Fig. 8C). These results demonstrated that the binding of ICP0 to the p65 RHD is sufficient to prevent the nuclear accumulation of NF-κB and thereby to preclude its transcriptional activity, and the RF domain of ICP0 is essential for this activity.
The RF domain of ICP0 is required for its interaction with p50.
ICP0 degraded p50 through the ubiquitin-proteasome pathway via its RF domain, so we reasoned that p50 might interact with the ICP0-RF domain directly. To verify this, HEK 293T cells were transfected with ICP0(1-241aa)-Flag or ICP0-ΔRF-Myc expression plasmid. As a result, ICP0(1-241aa) was efficiently co-IPed with endogenous p50, while the ICP0-ΔRF mutant did not (Fig. 9A to D). To determine whether p50 might interact with the ICP0-RF domain during viral infection, co-IP experiments were performed with endogenous p50. In ICP0-RF mutant HSV-1 R8513-infected cells, the RF mutant ICP0 did not IP with p50 using anti-p50 pAb and the control antibody IgG (Fig. 9E). These results, together with the results in Fig. 5, demonstrate that ICP0 interacts with endogenous p50 through its RF domain under physiological conditions during viral infection.
Fig 9.

The RF domain of ICP0 is required for its interaction with p50. The HEK 293T cells were transfected with ICP0(1-241aa)-Flag or ICP0-ΔRF-Myc expression plasmids. At 24 h after transfection, cells were mock treated (A and C) or treated (B and D) with TNF-α (10 ng/ml) and incubated for an additional 8 h. Cells were harvested and lysed, and the samples were then subjected to IP assays using anti-Flag or anti-Myc MAb (IP:Flag or Myc) or nonspecific mouse monoclonal antibody (IgG2b or IgG1). WB were probed with the indicated Ab. (E) HEK293T cells were infected with ICP0-RF mutant HSV-1 at an MOI of 10 for 16 h. The cells were then lysed, and the extracts were subjected to IP using anti-p50 pAb (IP:p50) or control IgG. Precipitates were analyzed by WB analysis with the indicated Ab.
DISCUSSION
NF-κB activation during viral infection is believed to be a protective response of the host to the viral pathogens. Therefore, many viruses have evolved distinct strategies to control the activation of NF-κB to evade the immune response. Known mechanisms of NF-κB suppression in virus-infected cells include inhibition of the IKK activity and IκB phosphorylation (56), inhibition of the NF-κB nuclear translocation (57), induction of the caspase-mediated cleavage of the p65 subunit (58), and inhibition of p65 transcriptional activation through protein-protein interaction (59).
The activation of NF-κB and IRF-3 and IRF-7, whether through TLR-dependent or -independent pathways, plays a major role in the immune response to virus infection. However, the molecular mechanism by which ICP0 interrupts innate immunity through TLR-independent pathways has not been reported. So we used TNF-α as a tool to study the TNFR-dependent NF-κB signaling pathway. As we know, these two innate pathways are important targets for immune evasion by HSV through its gene product ICP0, and ICP0 has been demonstrated to serve as one of several strategies used by HSV-1 to interrupt the innate immune responses. In this study, we have shown that the immediate early protein ICP0 encoded by HSV-1 dampens the TNF-α-mediated activation of the NF-κB signaling pathway.
The RF domain of ICP0 was very important for inhibiting IRF-3- and IRF-7-mediated activation of interferon-stimulated genes and TLR2-dependent NF-κB activity (35, 41), and also results obtained from another alphaherpesvirus, varicella-zoster virus (VZV), may yield some clues. The molecular mechanisms involved in the inhibition of the TNF-α-induced NF-κB activation by VZV have been well elucidated, which is dependent on the RF domain of ORF61, the homology protein of ICP0 (60). In this study, we demonstrated that the RF domain mutant ICP0 failed to inhibit the TNF-α-induced NF-κB activation, further demonstrating that ICP0 E3 ubiquitin ligase activity contributed to the inhibition of NF-κB activation.
ICP0 could inhibit TRADD-, TRAF2-, TAK1-, RIP1-, NIK-, IKKα-, IKKβ-, or p65-mediated activation of NF-κB-Luc promoter activity, implying that ICP0 might act at or downstream of the p50/p65 level. We hypothesized that ICP0 may interact with p65 or p50 to inhibit NF-κB promoter activity. To validate our hypothesis, co-IP analysis was employed to determine whether ICP0 could directly interact with p65 or p50 under physiological conditions. As expected, ICP0 interacted with both p65 and p50 during HSV-1 infection, and also ICP0 interacted with the RHD of p65 and abrogated the nuclear translocation of p65. ICP0 has been previously reported to be an inhibitor of TLR-dependent NF-κB responses, and expression of ICP0 reduces the levels of MyD88 and Mal and then blocks TLR2-driven responses at or downstream of the MyD88 adaptor and upstream of p65 (41). However, we found that ICP0 was able to suppress not only IKK-mediated NF-κB activity but also the NF-κB activity induced by ectopic expression of p65, which revealed a universal, downstream inhibitory mechanism by ICP0. We also found that ICP0 could inhibit NIK- and IKKα-induced NF-κB promoter activity, suggesting that ICP0 might be an antagonism of the noncanonical NF-κB signaling pathway.
p65/p50 is a crucial transcription factor in the NF-κB pathway and host innate immune system. Many viruses, including RNA and DNA viruses, antagonize the host proinflammation cytokine response at the p65/p50 level. For example, the West Nile virus NS1 protein, measles virus V protein, Hantaan virus nucleocapsid protein, and myxoma virus pyrin domain-containing protein M013 inhibit p65 nuclear translocation (47, 61–63). Variola virus G1R protein and myxoma virus pyrin domain-containing protein M013 protein were shown to be capable of interacting with NF-κB1/p105 and inhibiting the NF-κB signaling pathway (61, 64). In ectromelia virus, p105bp is encoded by Evm002 and is able to inhibit NF-κB1/p50 in cultured cells and reduce the speed and efficiency of NF-κB activation and then decrease transcription of several inflammatory genes (65).
In this study, we identified ICP0 as an antagonist of TNF-α-mediated NF-κB signaling, which could also bind to p65/p50 and inhibit downstream activation of the NF-κB signaling pathway. ICP0 bound to the RHD of p65 and inhibited its nuclear translocation, and also ICP0 bound to p50 via its RF domain. ICP0 degraded p50 by K48-linked ubiquitination. It was for the first time that we demonstrated that a viral protein could interact with p50 and promote its degradation.
The relationship between HSV-1 infection and the host NF-κB innate immune response is quite complex, and it is well known that the classical NF-κB pathway is significantly activated upon HSV-1 infections (66, 67) and was shown to be involved in upregulation of several host genes (68) and in promoting the progression of the virus replication cycle (66, 69). However, NF-κB signaling also promotes inflammatory cytokine expression to reduce viral infection. How could HSV-1 selectively promote sufficient NF-κB activity to promote its propagation and avoid the robust innate cytokine response driven by the same NF-κB activity? This apparent contradiction is reconciled by the fact that HSV-1 activates NF-κB through multiple signaling pathways. The early wave involves the UL31 and UL37 virion tegument protein (70, 71) and possibly virion glycoprotein D, glycoprotein B, glycoprotein H, and glycoprotein L (72, 73). UL37 activates NF-κB signaling through interaction with TRAF6, and glycoprotein D activates the TNF superfamily receptor herpesvirus entry mediator (HVEM) to drive both the canonical and alternative NF-κB pathways; glycoprotein B, glycoprotein H, and glycoprotein L are ligands to TLR2, and they initiate a signaling cascade which leads to NF-κB activation. Thus, HSV may transiently activate NF-κB to start its infection optimally and then inhibit the major part of the innate response using viral gene products expressed during infection. ICP0 might be a pivotal regulator in maintaining the balance between suppressing an antiviral response at a tolerable limit and keeping the sufficient activation of NF-κB for viral replication and consequently contribute to viral virulence.
In summary, this is the first study to demonstrate that the HSV-1 ICP0 protein suppresses the NF-κB-mediated antiviral response via interference with both p65 and p50. This function of ICP0 may benefit the regulation of the delicate balance between the replication process and host defense programs induced by the NF-κB pathway and may therefore provide an attractive target for the development of antiviral therapies.
ACKNOWLEDGMENTS
This work was supported by the National Natural Science Foundation of China (81171584) and the Program for Changjiang Scholars and Innovative Research Team in Soochow University (PCSIRT, IRT1075) and Jiangsu Provincial Innovative Research Team.
We thank Yossi Yarden (Weizmann Institute, Israel), Rao Anjana (Immune Disease Institute, Harvard Medical School), and Gangmin Hur (Chungnam National University) for providing plasmids and Haidong Gu and Bernard Roizman for providing the HSV-1 ICP0-RF mutant virus R8513.
Footnotes
Published ahead of print 25 September 2013
REFERENCES
- 1.Akira S, Uematsu S, Takeuchi O. 2006. Pathogen recognition and innate immunity. Cell 124:783–801 [DOI] [PubMed] [Google Scholar]
- 2.Beutler B, Eidenschenk C, Crozat K, Imler JL, Takeuchi O, Hoffmann JA, Akira S. 2007. Genetic analysis of resistance to viral infection. Nat. Rev. Immunol. 7:753–766 [DOI] [PubMed] [Google Scholar]
- 3.Medzhitov R. 2007. Recognition of microorganisms and activation of the immune response. Nature 449:819–826 [DOI] [PubMed] [Google Scholar]
- 4.Koyama S, Ishii KJ, Coban C, Akira S. 2008. Innate immune response to viral infection. Cytokine 43:336–341 [DOI] [PubMed] [Google Scholar]
- 5.Bonizzi G, Karin M. 2004. The two NF-kappaB activation pathways and their role in innate and adaptive immunity. Trends Immunol. 25:280–288 [DOI] [PubMed] [Google Scholar]
- 6.Ghosh S, May MJ, Kopp EB. 1998. NF-kappa B and Rel proteins: evolutionarily conserved mediators of immune responses. Annu. Rev. Immunol. 16:225–260 [DOI] [PubMed] [Google Scholar]
- 7.Li Q, Verma IM. 2002. NF-kappaB regulation in the immune system. Nat. Rev. Immunol. 2:725–734 [DOI] [PubMed] [Google Scholar]
- 8.Oeckinghaus A, Ghosh S. 2009. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 1:a000034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gilmore TD. 2006. Introduction to NF-kappaB: players, pathways, perspectives. Oncogene 25:6680–6684 [DOI] [PubMed] [Google Scholar]
- 10.Hayden MS, Ghosh S. 2004. Signaling to NF-kappaB. Genes Dev. 18:2195–2224 [DOI] [PubMed] [Google Scholar]
- 11.Baeuerle PA, Baltimore D. 1996. NF-kappa B: ten years after. Cell 87:13–20 [DOI] [PubMed] [Google Scholar]
- 12.Baldwin AS., Jr 1996. The NF-kappa B and I kappa B proteins: new discoveries and insights. Annu. Rev. Immunol. 14:649–683 [DOI] [PubMed] [Google Scholar]
- 13.Herbein G, O'Brien WA. 2000. Tumor necrosis factor (TNF)-alpha and TNF receptors in viral pathogenesis. Proc. Soc. Exp. Biol. Med. 223:241–257 [DOI] [PubMed] [Google Scholar]
- 14.Chen ZJ. 2005. Ubiquitin signalling in the NF-kappaB pathway. Nat. Cell Biol. 7:758–765 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Leib DA. 2002. Counteraction of interferon-induced antiviral responses by herpes simplex viruses. Curr. Top. Microbiol. Immunol. 269:171–185 [DOI] [PubMed] [Google Scholar]
- 16.Paladino P, Mossman KL. 2009. Mechanisms employed by herpes simplex virus 1 to inhibit the interferon response. J. Interferon Cytokine Res. 29:599–607 [DOI] [PubMed] [Google Scholar]
- 17.Kim JC, Lee SY, Kim SY, Kim JK, Kim HJ, Lee HM, Choi MS, Min JS, Kim MJ, Choi HS, Ahn JK. 2008. HSV-1 ICP27 suppresses NF-kappaB activity by stabilizing IkappaBalpha. FEBS Lett. 582:2371–2376 [DOI] [PubMed] [Google Scholar]
- 18.Jin H, Ma Y, Yan Z, Prabhakar BS, He B. 2012. Activation of NF-kappaB in CD8+ dendritic cells ex vivo by the gamma134.5 null mutant correlates with immunity against herpes simplex virus 1. J. Virol. 86:1059–1068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cotter CR, Kim WK, Nguyen ML, Yount JS, Lopez CB, Blaho JA, Moran TM. 2011. The virion host shutoff protein of herpes simplex virus 1 blocks the replication-independent activation of NF-kappaB in dendritic cells in the absence of type I interferon signaling. J. Virol. 85:12662–12672 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Xing J, Ni L, Wang S, Wang K, Lin R, Zheng C. 2013. HSV-1 encoded tegument protein VP16 abrogates the production of interferon beta by inhibiting NF-kappaB activation and blocking IRF-3 to recruit its coactivator CBP. J. Virol. 87:9788–9801 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhang J, Wang S, Wang K, Zheng C. 2013. Herpes simplex virus 1 DNA polymerase processivity factor UL42 inhibits TNF-alpha-induced NF-kappaB activation by interacting with p65/RelA and p50/NF-kappaB1. Med. Microbiol. Immunol. 202:313–325 [DOI] [PubMed] [Google Scholar]
- 22.Ea CK, Deng L, Xia ZP, Pineda G, Chen ZJ. 2006. Activation of IKK by TNFalpha requires site-specific ubiquitination of RIP1 and polyubiquitin binding by NEMO. Mol. Cell 22:245–257 [DOI] [PubMed] [Google Scholar]
- 23.Chau V, Tobias JW, Bachmair A, Marriott D, Ecker DJ, Gonda DK, Varshavsky A. 1989. A multiubiquitin chain is confined to specific lysine in a targeted short-lived protein. Science 243:1576–1583 [DOI] [PubMed] [Google Scholar]
- 24.Everett RD. 2000. ICP0, a regulator of herpes simplex virus during lytic and latent infection. Bioessays 22:761–770 [DOI] [PubMed] [Google Scholar]
- 25.Hagglund R, Roizman B. 2004. Role of ICP0 in the strategy of conquest of the host cell by herpes simplex virus 1. J. Virol. 78:2169–2178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Roizman B, Gu H, Mandel G. 2005. The first 30 minutes in the life of a virus: unREST in the nucleus. Cell Cycle 4:1019–1021 [DOI] [PubMed] [Google Scholar]
- 27.Everett RD, Meredith M, Orr A, Cross A, Kathoria M, Parkinson J. 1997. A novel ubiquitin-specific protease is dynamically associated with the PML nuclear domain and binds to a herpesvirus regulatory protein. EMBO J. 16:1519–1530 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kawaguchi Y, Van Sant C, Roizman B. 1997. Herpes simplex virus 1 alpha regulatory protein ICP0 interacts with and stabilizes the cell cycle regulator cyclin D3. J. Virol. 71:7328–7336 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Everett RD. 2000. ICP0 induces the accumulation of colocalizing conjugated ubiquitin. J. Virol. 74:9994–10005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lopez P, Van Sant C, Roizman B. 2001. Requirements for the nuclear-cytoplasmic translocation of infected-cell protein 0 of herpes simplex virus 1. J. Virol. 75:3832–3840 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Boutell C, Sadis S, Everett RD. 2002. Herpes simplex virus type 1 immediate-early protein ICP0 and its isolated RING finger domain act as ubiquitin E3 ligases in vitro. J. Virol. 76:841–850 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Everett RD. 1988. Analysis of the functional domains of herpes simplex virus type 1 immediate-early polypeptide Vmw110. J. Mol. Biol. 202:87–96 [DOI] [PubMed] [Google Scholar]
- 33.Everett RD. 1987. A detailed mutational analysis of Vmw110, a trans-acting transcriptional activator encoded by herpes simplex virus type 1. EMBO J. 6:2069–2076 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Eidson KM, Hobbs WE, Manning BJ, Carlson P, DeLuca NA. 2002. Expression of herpes simplex virus ICP0 inhibits the induction of interferon-stimulated genes by viral infection. J. Virol. 76:2180–2191 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lin R, Noyce RS, Collins SE, Everett RD, Mossman KL. 2004. The herpes simplex virus ICP0 RING finger domain inhibits IRF3- and IRF7-mediated activation of interferon-stimulated genes. J. Virol. 78:1675–1684 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Melroe GT, DeLuca NA, Knipe DM. 2004. Herpes simplex virus 1 has multiple mechanisms for blocking virus-induced interferon production. J. Virol. 78:8411–8420 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Melroe GT, Silva L, Schaffer PA, Knipe DM. 2007. Recruitment of activated IRF-3 and CBP/p300 to herpes simplex virus ICP0 nuclear foci: potential role in blocking IFN-beta induction. Virology 360:305–321 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mossman KL, Saffran HA, Smiley JR. 2000. Herpes simplex virus ICP0 mutants are hypersensitive to interferon. J. Virol. 74:2052–2056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Paladino P, Collins SE, Mossman KL. 2010. Cellular localization of the herpes simplex virus ICP0 protein dictates its ability to block IRF3-mediated innate immune responses. PLoS One 5:e10428. 10.1371/journal.pone.0010428 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Daubeuf S, Singh D, Tan Y, Liu H, Federoff HJ, Bowers WJ, Tolba K. 2009. HSV ICP0 recruits USP7 to modulate TLR-mediated innate response. Blood 113:3264–3275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.van Lint AL, Murawski MR, Goodbody RE, Severa M, Fitzgerald KA, Finberg RW, Knipe DM, Kurt-Jones EA. 2010. Herpes simplex virus immediate-early ICP0 protein inhibits Toll-like receptor 2-dependent inflammatory responses and NF-kappaB signaling. J. Virol. 84:10802–10811 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Xing J, Wang S, Lin F, Pan W, Hu CD, Zheng C. Comprehensive characterization of interaction complexes of herpes simplex virus type 1 ICP22, UL3, UL4, and UL20.5. J. Virol. 85:1881–1886 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gu H, Roizman B. 2007. Herpes simplex virus-infected cell protein 0 blocks the silencing of viral DNA by dissociating histone deacetylases from the CoREST-REST complex. Proc. Natl. Acad. Sci. U. S. A. 104:17134–17139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Xing J, Wang S, Li Y, Guo H, Zhao L, Pan W, Lin F, Zhu H, Wang L, Li M, Zheng C. Characterization of the subcellular localization of herpes simplex virus type 1 proteins in living cells. Med. Microbiol. Immunol. 200:61–68 [DOI] [PubMed] [Google Scholar]
- 45.Jiang X, Takahashi N, Matsui N, Tetsuka T, Okamoto T. 2003. The NF-kappa B activation in lymphotoxin beta receptor signaling depends on the phosphorylation of p65 at serine 536. J. Biol. Chem. 278:919–926 [DOI] [PubMed] [Google Scholar]
- 46.Severa M, Coccia EM, Fitzgerald KA. 2006. Toll-like receptor-dependent and -independent viperin gene expression and counter-regulation by PRDI-binding factor-1/BLIMP1. J. Biol. Chem. 281:26188–26195 [DOI] [PubMed] [Google Scholar]
- 47.Schuhmann KM, Pfaller CK, Conzelmann KK. 2011. The measles virus V protein binds to p65 (RelA) to suppress NF-kappaB activity. J. Virol. 85:3162–3171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Mercurio F, Zhu H, Murray BW, Shevchenko A, Bennett BL, Li J, Young DB, Barbosa M, Mann M, Manning A, Rao A. 1997. IKK-1 and IKK-2: cytokine-activated IkappaB kinases essential for NF-kappaB activation. Science 278:860–866 [DOI] [PubMed] [Google Scholar]
- 49.Park KA, Byun HS, Won M, Yang KJ, Shin S, Piao L, Kim JM, Yoon WH, Junn E, Park J, Seok JH, Hur GM. 2007. Sustained activation of protein kinase C downregulates nuclear factor-kappaB signaling by dissociation of IKK-gamma and Hsp90 complex in human colonic epithelial cells. Carcinogenesis 28:71–80 [DOI] [PubMed] [Google Scholar]
- 50.Gao S, Song L, Li J, Zhang Z, Peng H, Jiang W, Wang Q, Kang T, Chen S, Huang W. 2012. Influenza A virus-encoded NS1 virulence factor protein inhibits innate immune response by targeting IKK. Cell Microbiol. 14:1849–1866 [DOI] [PubMed] [Google Scholar]
- 51.Jordan M, Schallhorn A, Wurm FM. 1996. Transfecting mammalian cells: optimization of critical parameters affecting calcium-phosphate precipitate formation. Nucleic Acids Res. 24:596–601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zhong B, Yang Y, Li S, Wang YY, Li Y, Diao F, Lei C, He X, Zhang L, Tien P, Shu HB. 2008. The adaptor protein MITA links virus-sensing receptors to IRF3 transcription factor activation. Immunity 29:538–550 [DOI] [PubMed] [Google Scholar]
- 53.Zhu H, Zheng C, Xing J, Wang S, Li S, Lin R, Mossman KL. Varicella-zoster virus immediate-early protein ORF61 abrogates the IRF3-mediated innate immune response through degradation of activated IRF3. J. Virol. 85:11079–11089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Xing J, Wu F, Pan W, Zheng C. Molecular anatomy of subcellular localization of HSV-1 tegument protein US11 in living cells. Virus Res. 153:71–81 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.May MJ, Ghosh S. 1997. Rel/NF-kappa B and I kappa B proteins: an overview. Semin. Cancer Biol. 8:63–73 [DOI] [PubMed] [Google Scholar]
- 56.Cook JL, Walker TA, Worthen GS, Radke JR. 2002. Role of the E1A Rb-binding domain in repression of the NF-kappa B-dependent defense against tumor necrosis factor-alpha. Proc. Natl. Acad. Sci. U. S. A. 99:9966–9971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Friess M, Engelhardt P, Dobbelaere D, Zurbriggen A, Grone A. 2005. Reduced nuclear translocation of nuclear factor (NF)-kappaB p65 in the footpad epidermis of dogs infected with distemper virus. J. Comp. Pathol. 132:82–89 [DOI] [PubMed] [Google Scholar]
- 58.Neznanov N, Chumakov KM, Neznanova L, Almasan A, Banerjee AK, Gudkov AV. 2005. Proteolytic cleavage of the p65-RelA subunit of NF-kappaB during poliovirus infection. J. Biol. Chem. 280:24153–24158 [DOI] [PubMed] [Google Scholar]
- 59.Morrison TE, Kenney SC. 2004. BZLF1, an Epstein-Barr virus immediate-early protein, induces p65 nuclear translocation while inhibiting p65 transcriptional function. Virology 328:219–232 [DOI] [PubMed] [Google Scholar]
- 60.Sloan E, Henriquez R, Kinchington PR, Slobedman B, Abendroth A. 2012. Varicella-zoster virus inhibition of the NF-kappaB pathway during infection of human dendritic cells: role for open reading frame 61 as a modulator of NF-kappaB activity. J. Virol. 86:1193–1202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Rahman MM, Mohamed MR, Kim M, Smallwood S, McFadden G. 2009. Co-regulation of NF-kappaB and inflammasome-mediated inflammatory responses by myxoma virus pyrin domain-containing protein M013. PLoS Pathog. 5:e1000635. 10.1371/journal.ppat.1000635 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Taylor SL, Frias-Staheli N, Garcia-Sastre A, Schmaljohn CS. 2009. Hantaan virus nucleocapsid protein binds to importin alpha proteins and inhibits tumor necrosis factor alpha-induced activation of nuclear factor kappa B. J. Virol. 83:1271–1279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wilson JR, de Sessions PF, Leon MA, Scholle F. 2008. West Nile virus nonstructural protein 1 inhibits TLR3 signal transduction. J. Virol. 82:8262–8271 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mohamed MR, Rahman MM, Rice A, Moyer RW, Werden SJ, McFadden G. 2009. Cowpox virus expresses a novel ankyrin repeat NF-kappaB inhibitor that controls inflammatory cell influx into virus-infected tissues and is critical for virus pathogenesis. J. Virol. 83:9223–9236 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Rubio D, Xu RH, Remakus S, Krouse TE, Truckenmiller ME, Thapa RJ, Balachandran S, Alcami A, Norbury CC, Sigal LJ. 2013. Crosstalk between the type 1 interferon and nuclear factor kappa B pathways confers resistance to a lethal virus infection. Cell Host Microbe 13:701–710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Amici C, Belardo G, Rossi A, Santoro MG. 2001. Activation of I kappa b kinase by herpes simplex virus type 1. A novel target for anti-herpetic therapy. J. Biol. Chem. 276:28759–28766 [DOI] [PubMed] [Google Scholar]
- 67.Patel A, Hanson J, McLean TI, Olgiate J, Hilton M, Miller WE, Bachenheimer SL. 1998. Herpes simplex type 1 induction of persistent NF-kappa B nuclear translocation increases the efficiency of virus replication. Virology 247:212–222 [DOI] [PubMed] [Google Scholar]
- 68.Taddeo B, Esclatine A, Roizman B. 2002. The patterns of accumulation of cellular RNAs in cells infected with a wild-type and a mutant herpes simplex virus 1 lacking the virion host shutoff gene. Proc. Natl. Acad. Sci. U. S. A. 99:17031–17036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Gregory D, Hargett D, Holmes D, Money E, Bachenheimer SL. 2004. Efficient replication by herpes simplex virus type 1 involves activation of the IkappaB kinase-IkappaB-p65 pathway. J. Virol. 78:13582–13590 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Liu X, Fitzgerald K, Kurt-Jones E, Finberg R, Knipe DM. 2008. Herpesvirus tegument protein activates NF-kappaB signaling through the TRAF6 adaptor protein. Proc. Natl. Acad. Sci. U. S. A. 105:11335–11339 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Roberts KL, Baines JD. 2011. UL31 of herpes simplex virus 1 is necessary for optimal NF-kappaB activation and expression of viral gene products. J. Virol. 85:4947–4953 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Leoni V, Gianni T, Salvioli S, Campadelli-Fiume G. 2012. Herpes simplex virus glycoproteins gH/gL and gB bind Toll-like receptor 2, and soluble gH/gL is sufficient to activate NF-kappaB. J. Virol. 86:6555–6562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Medici MA, Sciortino MT, Perri D, Amici C, Avitabile E, Ciotti M, Balestrieri E, De Smaele E, Franzoso G, Mastino A. 2003. Protection by herpes simplex virus glycoprotein D against Fas-mediated apoptosis: role of nuclear factor kappaB. J. Biol. Chem. 278:36059–36067 [DOI] [PubMed] [Google Scholar]