

Abstract
Obesity is a major worldwide public health issue. The physiological systems that regulate body weight are thus of great interest as targets for anti-obesity agents. Peptidergic systems are critical to the regulation of energy homeostasis by key regions in the hypothalamus and brainstem. A number of neuropeptide systems have therefore been investigated as potential treatments for obesity. Blocking orexigenic peptide signals such as neuropeptide Y, melanin-concentrating hormone, orexins, relaxin-3 and galanin-like peptide or stimulating anorectic signalling pathways used by peptides such as the melanocortins, ciliary neurotrophic factor and brain-derived neurotrophic factor, are approaches that have shown some promise, but which have also highlighted possible concerns. Manipulation of central peptidergic systems poses a number of therapeutic problems, including brain access and side effects. Given that the homeostatic defence of body weight may limit the effectiveness of any single-target therapy developed, a combination therapy approach may offer the best hope for the effective prevention and treatment of obesity.
LINKED ARTICLES
This article is part of a themed section on Neuropeptides. To view the other articles in this section visit http://dx.doi.org/10.1111/bph.2013.170.issue-7
Keywords: obesity, neuropeptides, hypothalamus, orexigenic, anorectic
Introduction
Obesity is a major public health issue worldwide. It is estimated that more than 1.5 billion individuals are overweight and more than 400 million adults are obese (World Health Organization, 2011). These figures are anticipated to rise, with predictions that by 2050, 60% of men, 50% of women and 25% of children under 16 in the UK will be obese (Foresight Report, 2007). Increased weight predisposes to and can aggravate many clinical conditions, including cardiovascular disease, type II diabetes mellitus, restrictive lung disease, certain cancers and infertility (Kopelman, 2000). Obesity is a multifactorial condition in which genetic, environmental, behavioural and socio-economic conditions all contribute to determine body weight, adipose tissue mass and distribution (Bray, 1992).
Current therapeutic options
For a new drug to be considered efficacious in the treatment of obesity by the US Food and Drug Administration (FDA), it must meet certain criteria in randomized controlled trials carried out over a minimum period of 1 year. The mean placebo-subtracted weight loss must be at least 5% from baseline, or alternatively, at least 35% of patients in the drug-treated group must lose more than 5% of baseline body weight (and the loss must be statistically significant and double that of the placebo group; FDA, 2007). The European Medicine Agency (EMEA) suggests that efficacy is demonstrated with a loss of at least 10% baseline body weight, statistically significant compared to placebo, following a minimum 1 year treatment period (EMEA, 2006).
Currently, the only approved drug for the long-term treatment of obesity in the UK is orlistat (Xenical), which reduces the absorption of dietary fat by inhibiting intestinal lipases (Hauptman et al., 1992; Sjöström et al., 1998). There are a number of anti-obesity drugs, which are internationally approved but which are not licensed for use in the UK. These include appetite-suppressing amphetamine analogues such as phentermine, which are only approved for short periods (less than 12 weeks) due to concerns regarding tolerance and dependency (Ioannides-Demos et al., 2011). Qsymia, a combination of phentermine and topiramate, and lorcaserin, a selective 5-HT2C receptor agonist (receptor nomenclature follows Alexander et al., 2011), have recently been approved for the treatment of obesity by the FDA (FDA, 2012). Two drugs previously approved for the treatment of obesity, which target the CNS, have been withdrawn in recent years due to their unacceptable side effects. Sibutramine (Reductil), a 5-HT and noradrenaline reuptake inhibitor was withdrawn in 2010 due to its cardiovascular side effects including increased blood pressure and heart rate associated with an increased risk of stroke and myocardial infarction (James et al., 2000; Kim et al., 2003). Rimonabant, an inverse agonist for the cannabinoid CB1 receptor was withdrawn in 2009 due to reports of severe depression and suicidal ideation (Van Gaal et al., 2005; Topol et al., 2010).
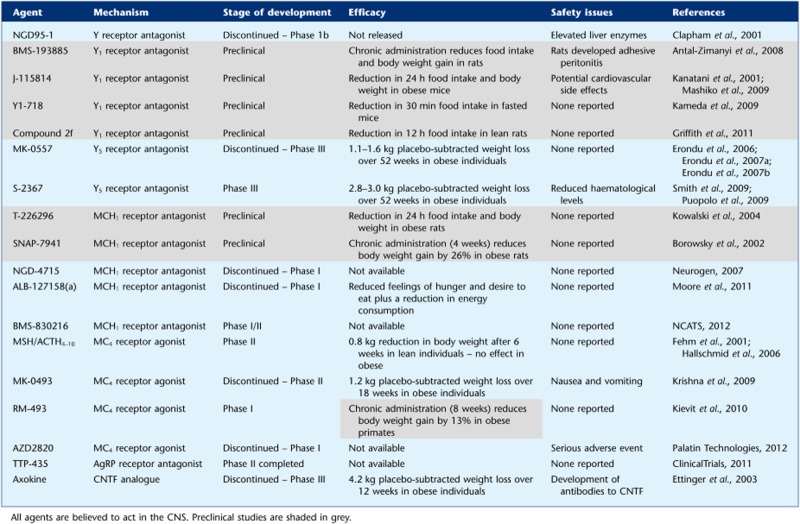
The withdrawal of these drugs illustrates the need for more selective drug targets in order to minimize adverse effects. Obesity is associated with numerous co-morbidities and any treatment for obesity is likely to be required long term; the therapeutic window of any new pharmacotherapy must therefore be closely scrutinized before approval. The classical neurotransmitter and endocannabinoid systems are widespread within the CNS, reflecting multiple physiological roles, and manipulation of these systems carries the risk of unwanted effects. Neuropeptides can act in the CNS as neuromodulators of the classical neurotransmitter systems. Typically expressed by discrete clusters of neurons, they project diffusely throughout the CNS, and modulate postsynaptic neurons in a way that can alter their responses to classical neurotransmitters. All CNS neurons that produce neuromodulators also release classical neurotransmitters, for example, neuropeptide Y (NPY) and galanin regulate central adrenergic transmission within the hypothalamus (Tsuda et al., 1989). Neuropeptides tend to be larger than classic neurotransmitters, and usually have more receptor recognition sites or motifs, leading to greater binding affinity and higher receptor selectivity, theoretically making pharmacological intervention less prone to side effects. Certain neuropeptidergic systems are more discretely localized than others and are thought to play more specific roles than classical neurotransmitters. Neuropeptides thus appear potentially more promising drug targets. However, despite the advances made over the past 40 years in our understanding of the neuropeptide systems controlling appetite, the numerous drug discovery programmes targeting these systems have not yielded an anti-obesity agent for clinical use (Table 1) (Elmquist et al., 1999; Gautron and Elmquist, 2011). The majority of these programmes, discontinued at a later stage, were abandoned due to safety issues or a lack of efficacy. In part, this may reflect the increased emphasis on safety for such agents over the period, and in some cases, the lack of efficacy is likely due to the lower doses being required because of safety concerns associated with higher doses (Greenway and Caruso, 2005). It may also reflect the difficulties in designing peptide-based agents, which can access the brain, and of developing small-molecule agonists for particular families of the GPCRs that mediate the effects of many of these neuropeptides (Miller et al., 2004). The problem of translating acute anorectic effects into long-term reductions in appetite and body weight may be due to tachyphylaxis and/or the tendency of other compensatory systems to act to oppose body weight loss. Thus, while stimulation of anorectic neuropeptide systems or blockade of orexigenic neuropeptide systems may be viable options for the design of new drugs, they have yet to prove their utility. Understanding of the complex pathways that regulate energy balance and the redundancy of these systems is critical to the identification of the most promising novel therapeutic targets and to identify potential safety concerns.
Table 1.
Agents targeting neuropeptidergic systems investigated for their potential as anti-obesity therapies
 |
The purpose of this article is to discuss CNS peptides and their therapeutic potential in the treatment of obesity. Brain-derived neurotrophic factor (BDNF), though a growth factor, has been included as there is evidence to suggest that its neuromodulatory properties regulate energy homeostasis (Nicholson et al., 2007). The effects of peripheral signals on the regulation of energy homeostasis, including those which act centrally to modulate appetite and energy expenditure, are not within the scope of this review, but are described in detail elsewhere (Murphy et al., 2006). We will examine the neuropeptide systems identified as potential targets for anti-obesity agents, reflect on the outcomes of clinical trials of agents that targeting these systems and consider the possible safety implications of manipulating these neuropeptidergic systems.
Central appetite regulation
Despite wide variation in daily food intake and energy expenditure, most individuals are able to maintain a remarkably stable body weight (Leibel, 1990). This homeostatic regulation is achieved by hypothalamic centres that receive signals from other CNS regions and from the periphery (summarized in Figure 1). Such peripheral signals can reflect body fat stores and nutritional state, and include nutrients, adipose tissue-, pancreas- and gut-derived peptides, such as insulin, leptin, ghrelin, glucagon-like peptide-1, peptide YY and cholecystokinin. The hypothalamus integrates and processes signals of energy homeostasis and induces appropriate changes in appetite and energy expenditure.
Figure 1.

Schematic diagram illustrating the neuropeptidergic signalling pathways regulating energy homeostasis. Hypothalamic nuclei integrate and process signals of energy homeostasis from the periphery and other CNS regions and induce appropriate changes in appetite and energy expenditure. Peptides coloured blue are orexigenic, promoting food intake, while peptides coloured red are anorexigenic, promoting satiety. Receptors are in white boxes. Purple indicates neurons, which can be either orexigenic or anorexigenic. Non-peptidergic neurotransmitters are not included in this diagram for clarity. The peripheral factors in the diagram are reviewed elsewhere (Murphy et al., 2006). CNTF-R, ciliary neurotrophic factor receptor; GalR1/2/3, galanin receptors 1,2 and 3; GHS-R, growth hormone secretagogue receptor; INS-R, insulin receptor; MC3/4-R, melanocortin 3/4 receptors; NAc, nucleus accumbens; OB-R, leptin receptor; RXFP3, relaxin/insulin-like family peptide receptor 3; TRkB-R, tyrosine-related kinase B receptor; VTA, ventral tegmental area; Y1/Y5-R, neuropeptide Y1/Y5 receptors.
The hypothalamus comprises several discrete nuclei, which communicate with each other, and with other parts of the brain via release of specific neuropeptides. These neuropeptide systems are potential targets for anti-obesity therapies. The hypothalamic nuclei important in the regulation of energy homeostasis include the arcuate nucleus (ARC), the paraventricular nucleus (PVN), the dorsomedial and ventromedial nuclei (DMN and VMN) and the lateral hypothalamic area (LHA; Figure 1).
The hypothalamus also contains and receives signals from neuronal circuits utilizing non-peptidergic neurotransmitters, including noradrenergic, dopaminergic, serotonergic, histaminergic and endocannabinoid signals. It is these systems that have predominantly been targeted in the treatment of obesity to date. Extra-hypothalamic brain regions important in regulating energy homeostasis, and which communicate with the hypothalamus, include the brainstem (which itself receives neural signals from the gut via the vagus nerve and hormonal signals), the ventral tegmental area within the midbrain and the nucleus accumbens in the striatum (Contreras et al., 1984; Hommel et al., 2006; Figure 1). The latter two brain regions are thought to be important in non-homeostatic regulation of feeding behaviour, modulating motivated behaviour, addiction and reward.
The hypothalamic ARC is thought to be pivotal to the integration and interpretation of signals of energy balance. Located at the base of the hypothalamus next to the median eminence with its incomplete blood–brain barrier, the ARC is able to respond to circulating hormones such as leptin, insulin and ghrelin (Banks, 2006). There are two well characterized neuronal populations within the ARC known to be involved in energy homeostasis (Cone et al., 2001). One population expresses the orexigenic peptides NPY and agouti-related peptide (AgRP; Broberger et al., 1998; Hahn et al., 1998). The second population produces the anorexigenic peptides α-melanocyte-stimulating hormone (α-MSH), cleaved from the pro-opiomelanocortin (POMC) precursor molecule and a peptide encoded by cocaine and amphetamine regulated transcript (CART; Elias et al., 1998a). The expression of NPY and AgRP mRNA in the ARC is increased by fasting (Hahn et al., 1998), and the activity of these neurons is directly inhibited by leptin and insulin (Sipols et al., 1995; Stephens et al., 1995) and stimulated by ghrelin (Kohno et al., 2003; Figure 1). POMC/CART expressing neurons are also regulated by leptin and insulin (Cheung et al., 1997; Kristensen et al., 1998), and their selective ablation causes hyperphagia and obesity (Xu et al., 2005). The expression of POMC and CART mRNA in the ARC is decreased by fasting (Kristensen et al., 1998; Smith and Funder, 1988). Administration of leptin directly into the ARC inhibits feeding (Satoh et al., 1997) and lesions of the ARC attenuate the anorectic effect of peripheral leptin administration (Tang-Christensen et al., 1999), suggesting that the ARC is critical in mediating leptin's effects on energy homeostasis.
Hypothalamic orexigenic neuropeptides
Neuropeptide Y
NPY is a 36 amino acid neuropeptide, which belongs to the pancreatic polypeptide (PP) fold family of proteins, together with the gut hormones peptide YY and pancreatic polypeptide (Glover et al., 1984). The PP fold peptides bind to members of the Y receptor family (Y1 to Y5; Blomqvist and Herzog, 1997). NPY is one of the most abundant neuropeptides in the CNS (Allen et al., 1984). In the hypothalamus, it is synthesized by several hypothalamic nuclei but the NPY neurons in the ARC are best characterized and known to project to the PVN, the DMN and the LHA (Bai et al., 1985; Chronwall et al., 1985). NPY is the most potent orexigenic neuropeptide known (Woods et al., 1998). Injection of NPY into the hypothalamus increases food intake acutely in rats (Clark et al., 1984; Stanley and Leibowitz, 1984), while chronic administration results in hyperphagia and obesity (Stanley et al., 1986; Zarjevski et al., 1993). Hypothalamic NPY synthesis and release is increased in response to fasting, and is regulated by peripheral signals that reflect body nutritional status, such as ghrelin, insulin and leptin, suggesting that NPY plays a physiological role in the regulation of feeding (Sahu et al., 1988; 1995; Schwartz et al., 1992; Kamegai et al., 2001). NPY binds preferentially to the Y1 and Y5 receptors, and administration of Y1 or Y5 receptor antagonists attenuates feeding in fasted rats (Kanatani et al., 1996; Criscione et al., 1998). Surprisingly, NPY knockout mice do not display any profound alterations in their food intake and body weight, and respond normally to leptin, suggesting developmental compensation by other neuronal pathways (Erickson et al., 1996; Qian et al., 2002). In addition, Y1 and Y5 receptor knockout mice exhibit only slight alterations in energy homeostasis developing late onset mild obesity (Marsh et al., 1998; Pedrazzini et al., 1998). However, post-natal ablation of ARC NPY/AgRP neurones leads to hypophagia and weight loss in mice, and knockdown of NPY expression in the adult rat ARC reduces food intake and body weight gain, confirming an important physiological role for this neuronal population in energy homeostasis (Bewick et al., 2005; Gardiner et al., 2005; Gropp et al., 2005; Luquet et al., 2005).
Given the role of Y1 and Y5 receptors in mediating the orexigenic effects of NPY, many specific antagonists of these receptors have been developed as putative anti-obesity agents (see Yulyaningsih et al., 2011). NGD95-1, an early non-selective NPY receptor antagonist reached clinical trial stage but the trial was suspended following the detection of elevated levels of liver enzymes in the blood of subjects (Clapham et al., 2001).
Selective Y1 receptor antagonists have been investigated in animal studies but have not yet progressed to clinical trials in humans. BMS-193885 is a potent and selective Y1 receptor antagonist, which reduces food intake, and which results in an 8.5% reduction in body weight following 44 days of administration in rats. However, it lacks oral bioavailability as it is poorly absorbed by the intestine. BMS-193885 exhibited no behavioural side effects but a significant number of rats developed adhesive peritonitis following chronic administration, thought to result from poor tissue absorption of the compound (Antal-Zimanyi et al., 2008). J-115814, a 2,4-diaminopyridine-based selective Y1 receptor antagonist, reduced 24 hour food intake by 50–90% and body weight at 24 hours by approximately 5% in both leptin-resistant (db/db) and diet-induced obese (DIO) mice following peripheral administration (Kanatani et al., 2001; Mashiko et al., 2009). It has been suggested that this compound may be associated with cardiovascular side effects due to its interaction with hERG channels (Fermini and Fossa, 2003) and no further data has been released, suggesting its development has been terminated. A similar molecule, Y1-718, was subsequently designed to eliminate activity at these channels (Kameda et al., 2009) and has been used to validate a PET ligand in studies examining Y1 receptor distribution in the monkey brain (Hostetler et al., 2011). However, there are currently no suggestions of its use as a potential therapeutic agent. More recently, a series of pyrazolo[1,5-a]pyrimidine derivatives with high binding affinity and selectivity at Y1 receptors has been designed. Peripheral administration of compound 2f of this series resulted in a 12% reduction in nocturnal food intake, 12 hours after administration in lean rats (Griffith et al., 2011). The effects of chronic administration have not yet been reported.
Many Y5 receptor antagonists have been designed and have shown efficacy at reducing food intake and body weight in rodent models of obesity (Criscione et al., 1998; Daniels et al., 2002; Kakui et al., 2006; Mashiko et al., 2008; Sato et al., 2008; Omori et al., 2012). However, of these, only two (MK-0557 and S-2367) have progressed to clinical trial stage, and MK-0557 was recently discontinued due to its modest effect. MK-0557, an orally available Y5 antagonist, reduced body weight gain by 40% in mouse models of obesity after chronic peripheral administration for 35 days (Erondu et al., 2006). Clinical trials demonstrated that MK-0557 induced weight loss of 2 kg in obese individuals over a 12 week period and was generally safe and well tolerated. A longer-term study lasting 52 weeks in a cohort of 1661 overweight and obese individuals resulted in only a 1.1 kg placebo-subtracted weight loss (Erondu et al., 2006). Furthermore, MK-0557 treatment in combination with a hypocaloric diet for 52 weeks or with co-administration of sibutramine or orlistat for a period of 24 weeks failed to produce weight loss of greater than 1.6 kg over and above the effects of energy restriction or sibutramine/orlistat alone (Erondu et al., 2007a; 2007b). S-2367 (Velneperit), another orally active Y5 receptor antagonist, which reduced body weight in DIO mice (Yukioka et al., 2006) induced a 2.8 kg placebo-subtracted reduction in body weight and reduced waist and hip circumference in obese subjects after 52 weeks of treatment. The efficacy of S-2367 was enhanced to a 3 kg placebo-subtracted weight loss after 52 weeks when administered to subjects following a low-calorie diet (Puopolo et al., 2009; Smith et al., 2009). S-2367 was well tolerated with no significant safety issues. However, there are reports of a small reduction in the levels of erythrocyte count, haemoglobin and haematocrit in the majority of the trial subjects, raising a potential risk of anaemia that requires further investigation (Flach et al., 2007).
Considering the potent effect of NPY on appetite and energy balance, the results of clinical trials have been relatively disappointing. Poor oral bioavailability and brain penetrability, toxicity and lack of long-term effects have prevented the progress of many of these compounds into a clinical setting. It is apparent that NPY-induced feeding is mediated via more than one receptor subtype and that redundancy of such a critical system is likely to exist between Y1 and Y5 receptor signalling, whereby blockade at individual Y receptors may be overcome by increased activity at other Y receptors. Simultaneous targeting of more than one Y receptor with the aim of synergistic effects may therefore prove more successful in treating obesity, though to date, no dual Y1/Y5 receptor antagonists have reached a clinical trial stage. It is also important to consider the other roles of the NPY system, including the modulation of cardiovascular function, anxiety, seizures, memory, circadian rhythm, bone density and sexual functions (Brothers and Wahlestedt, 2010), which may be also be affected by long-term administration of NPY receptor antagonists.
Melanin-concentrating hormone
Melanin-concentrating hormone (MCH) is produced by neurons of the LHA and the zona incerta. MCH neurons project to other hypothalamic nuclei and to limbic areas reflecting its involvement in feeding, arousal and sensorimotor integration (Bittencourt et al., 1992). MCH expression is increased with fasting (Qu et al., 1996) and chronic MCH administration leads to hyperphagia and obesity in rodents (Della-Zuana et al., 2002; Gomori et al., 2003). Transgenic mice overexpressing MCH have an obese hyperphagic phenotype (Ludwig et al., 2001) and conversely, targeted deletion of MCH results in a hypophagic, lean phenotype with increased metabolic rate (Shimada et al., 1998). MCH binds to the MCH1 receptor, which is widely distributed throughout the rodent brain, and highly expressed in hypothalamic nuclei (Marsh et al., 2002). A second MCH receptor exists in humans (MCH2), although its function remains relatively obscure, as it is absent in rodents (Tan et al., 2002). Selective MCH1 receptor antagonists were first developed in 2002 and have demonstrated efficacy at reducing food intake and body weight in animal models of obesity (Borowsky et al., 2002; Takekawa et al., 2002).
Several pharmaceutical companies have subsequently created MCH1 receptor antagonists, which have been assessed for their efficacy and safety in human clinical trials (reviewed in Luthin, 2007). NGD-4715, a small molecule MCH1 receptor antagonist developed by Neurogen reportedly displayed anorectic effects in animal studies and completed phase I clinical trials in humans (Neurogen, 2007). While it was reported to be safe and well tolerated, details regarding its efficacy have not been released and no further trials have been pursued (Luthin, 2007). ALB-127158(a), a selective high-affinity MCH1 receptor antagonist developed by Albany Molecular Research, Inc. has been shown to reduce food intake by up to 30% and body weight by 18% following 28 days of administration in DIO mice (Guzzo et al., 2010). This molecule has recently completed phase I trials in humans and has been reported to be safe and well tolerated (Moore et al., 2011). Subjects reported an approximately 30% reduction in feelings of hunger and desire to eat, and a reduction in test meal consumption was observed after 14 days of treatment (Moore et al., 2011). A later report suggested that as higher doses than those originally predicted would be required, further studies with this molecule would not be carried out. It is possible that similar molecules with better CNS penetrance, and thus requiring lower doses, will be developed (AMRI, 2011). BMS-830216 is an orally bioavailable pro-drug of BMS-819881, a selective and potent antagonist of the MCH1 receptor developed by Bristol-Myers-Squibb. Preclinical studies demonstrated a reduction in food intake and a 6% reduction in body weight in both lean and DIO rats (NCATS, 2012). This molecule recently underwent phase I/II trials in humans and while details of its efficacy are awaited, reports released by the company suggest that it is generally safe and well tolerated (NCATS, 2012).
While the development of MCH1 receptor antagonists for the treatment of obesity has received a great deal of interest, the majority of drug programs targeting this system have been discontinued. It is important to consider the potential effects of manipulation of the MCH system. Although MCH mRNA has rather limited expression in the brain, MCH-containing fibres are widely distributed within the CNS, and MCH has been attributed a diverse range of functions (Luthin, 2007). Antagonism at the MCH1 receptor may therefore potentially cause numerous adverse effects. Most significantly, MCH has been demonstrated to reduce the incidence of seizures (Knigge and Wagner, 1997) and although there have been no reports of any pro-seizure effects of MCH1 receptor antagonists in human trials, this is a concern for chronic use. Antagonism at the MCH1 receptor has also been associated with a profound drop in blood pressure in animal studies (Kym et al., 2006; Lynch et al., 2006) but again, this has not been reported in human trials. MCH also has complex effects on several hypothalamo-pituitary axes (Saito and Nagasaki, 2008) and it will be important to assess whether small molecule MCH1 receptor antagonists influence the hypothalamic-pituitary-gonadal, -thyroid or -adrenal axes in clinical trials. Other concerns involve roles of MCH in learning and memory, and in the maintenance of bone density (Luthin, 2007).
Orexins
Orexin A and orexin B are hypothalamic neuropeptides synthesized in cell bodies within the LHA (de Lecea et al., 1998; Sakurai et al., 1998)and orexin-containing fibres project widely throughout the brain (Elias et al., 1998b; Date et al., 1999). The orexins bind to the orexin receptors OX1 and OX2; the OX1 receptor binds orexin A with greater affinity than orexin B, while the OX2 receptor binds both orexins with equal affinity (Sakurai et al., 1998). Central injection of either of the orexins stimulates food intake in rats (Edwards et al., 1999), and prepro-orexin mRNA expression is up-regulated in fasted rats (Sakurai et al., 1998). The orexins are also involved in arousal; orexin and OX2 receptor deficient mice exhibit a phenotype similar to human narcolepsy (Chemelli et al., 1999; Tokita et al., 2001), which itself is characterized by a deficiency in orexin production (Nishino et al., 2000).
Orexin receptor antagonists have been developed and progressed through clinical trials for the treatment of sleep disorders. Almorexant is a non-selective orexin receptor antagonist (Owen et al., 2009), which completed phase III clinical trials (Hoever et al., 2012). Further development of this drug was discontinued recently following a review of data from additional clinical studies conducted to further establish the clinical profile of almorexant, including the tolerability profile (Actelion, 2011). MK-4305 (Suvorexant), a second orexin receptor antagonist has also completed phase III trials for its effects on sleep disorders (Cox et al., 2010). There is no data currently available regarding the effects of these drugs on body weight in trial subjects, but it will be interesting to see if these compounds have anti-appetite actions.
Orexins have also been suggested to modulate pleasure/reward pathways in the brain (Harris et al., 2005; Aston-Jones et al., 2010). It seems unlikely that targeting the orexin system will result in a viable therapy for obesity due to the powerful centrally mediated behavioural effects of this system.
Relaxin-3
Relaxin-3 (INSL-7) is a member of the insulin superfamily, a group of structurally related peptide hormones that includes insulin, insulin-like growth factors, relaxins and insulin-like peptides (Bathgate et al., 2002). Relaxin-3 mRNA is predominantly expressed in the brainstem in neurons that project to brains areas including the hypothalamic nuclei involved in energy homeostasis (Burazin et al., 2002; Tanaka et al., 2005). Relaxin-3 binds to RXFP3 (GPCR135) and also to two other receptors: LGR7 and GPCR142 (Liu et al., 2003a,b; Sudo et al., 2003). RXFP3 is highly expressed in the PVN and the supraoptic nucleus of the hypothalamus, and relaxin-3 neurons also extend to these regions (Chen et al., 2005; Smith et al., ). Central injection of relaxin-3 for 14 days increases food intake threefold and increases body weight gain by approximately 15% in rats, an effect blocked by pretreatment with the selective RXFP3 antagonist (McGowan et al., 2005; 2006; Hida et al., 2006; Kuei et al., 2007). Relaxin-3 knockout mice exhibit a lean phenotype (Sutton et al., 2009). These results suggest therapeutic potential for RXFP3 antagonists to reduce body weight gain in obesity. However, an RXFP3 antagonist given alone had no effect on food intake in satiated rats (Sutton et al., 2009). The discrete localization of the relaxin-3/RXFP3 system in the brain holds the promise of highly selective antagonists that can reduce body weight, although relaxin-3 signalling also modulates an array of circuits involved in arousal, stress responses, affective state and cognition (see van der Westhuizen et al., 2008). A more complete understanding of the biological roles of the relaxin family peptides and their receptors will be useful in determining their utility as therapeutic targets.
Galanin-like peptide
Galanin-like peptide (GALP) is a member of the galanin peptide family originally discovered as another endogenous ligand for the galanin receptors (Ohtaki et al., 1999). The distribution of GALP mRNA is restricted in the CNS to the ARC and the median eminence (Larm and Gundlach, 2000; Jureus et al., 2001; Takatsu et al., 2001). Central administration of GALP increases 2 hour food intake by up to 300% in both lean and DIO rats (Lawrence et al., 2002; Matsumoto et al., 2002; Tan et al., 2005). Counter-intuitively for an orexigenic peptide, GALP mRNA expression is decreased in response to fasting and is positively regulated by insulin and leptin, suggesting the actions of GALP on energy homeostasis are complex (Jureus et al., 2000; Fraley et al., 2004; Johansson et al., 2008). The increase in GALP expression in response to leptin may reflect a role in neuroendocrine regulation rather than in energy homeostasis. In addition to its effects on energy homeostasis, GALP also has a role in the regulation of the hypothalamo-pituitary gonadal axis (see Lang et al., 2007). There is an interest in the development of GALP as a therapeutic agent for the treatment of obesity due to its restricted distribution in the CNS and relatively specific biological activity (Shiba et al., 2010). However, GALP is known to bind to several receptors, increasing the chance of side effects, and it may not be possible to separate the effects of GALP on food intake from its effects on the reproductive axis. Until the biology of the galanin peptide family is completely understood, pharmacologically manipulating the GALP system remains problematic.
Hypothalamic anorectic neuropeptides
Melanocortins
The hypothalamic melanocortin system plays a critical role in energy homeostasis. The melanocortins are peptide products of the POMC polypeptide precursor, and include adrenocorticotrophic hormone (ACTH) and α, β and γ-MSH (Bertagna, 1994). The melanocortins mediate their effects via five GPCRs (MC1-MC5), though only MC3 and MC4 receptors are thought to regulate energy homeostasis. POMC mRNA is expressed in the ARC and the brainstem and is decreased in response to fasting (Smith and Funder, 1988). POMC and MC4 receptor knockout mice are hyperphagic and obese, suggesting a critical physiological role for the melanocortin circuitry in the regulation of energy homeostasis (Huszar et al., 1997; Yaswen et al., 1999).
α-MSH is an agonist at both the MC3 and MC4 receptors (Adan et al., 1994), and inhibits feeding after central injection in food-deprived rats (Tsujii and Bray, 1989). Unusually, the MC3 and MC4 receptors also have an endogenous antagonist, AgRP (Fong et al., 1997). Central injection of AgRP increases food intake by blocking the anorectic effect of α-MSH at MC4 receptors (Rossi et al., 1998), and transgenic mice overexpressing AgRP have a similar phenotype to MC4 receptor knockout mice (Graham et al., 1997; Ollmann et al., 1997). Melanotan-II, a melanocortin receptor agonist with increased potency at MC4 receptors reduces food intake in mice, an effect which is attenuated in MC4 receptor knockout mice (Chen et al., 2000). Furthermore, MC3 receptor knockout mice do not have altered body weight, but do have increased adipose mass (Butler et al., 2000). These studies suggest that it is the MC4 receptors that mediate an anorectic tone important in the restriction of food intake. The physiology of the melanocortin system is similar in humans to rodents, and individuals with genetic mutations in POMC or the MC3 and MC4 receptors are hyperphagic and obese (Vaisse et al., 1998; Yeo et al., 1998; Biebermann et al., 2003). Indeed, mutations in the MC3 and MC4 receptors are the most common known monogenetic causes of human obesity, although only with a prevalence of 4% in a population of morbidly obese patients (Vaisse et al., 2000).
Preclinical studies with synthetic melanocortin ligands have demonstrated potent effects on food intake and energy expenditure in animal models. However, few of these compounds have been investigated for their safety and efficacy in humans. Intranasal administration of the melanocortin core fragment MSH/ACTH4–10, an agonist at MC4 receptors, reduced body weight by 0.8 kg and body fat by 1.7 kg in normal weight individuals following 6 weeks of treatment, but caused no significant reduction in body weight or body fat in overweight subjects or in patients with mutations in the POMC gene (Fehm et al., 2001; Krude et al., 2003; Hallschmid et al., 2006). A potent and selective MC4 receptor agonist MK-0493 reached phase II clinical trials but was associated with only a non-significant, 1.2 kg placebo-subtracted weight reduction in obese subjects after 18 weeks of treatment and further studies are not planned (Krishna et al., 2009). These two studies with negligible effects on weight loss suggest that obesity may be associated with melanocortin resistance.
RM-493 (formerly BIM 22493) is a small peptide agonist with high selectivity for MC4 receptors. In preclinical studies, obese primates treated with RM-493 for 8 weeks displayed reduced food intake, a 13% reduction in body weight and improved glucose tolerance. This study also showed improvements in both heart rate and blood pressure, probably attributable to the improved body weight (Kievit et al., 2010). RM-493 is currently undergoing a phase I clinical trial as a potential new treatment for obesity (Rhythm Pharmaceuticals, 2011). Recently, AZD2820, an MC4 receptor partial agonist administered by s.c. injection, entered phase I trials. The trial was halted due to a serious adverse event suspected to be an allergic reaction after a first dose (Palatin Technologies, ). A small molecule inhibitor of AgRP, TTP-435 has been developed and has completed phase II clinical trials (ClinicalTrials.gov, 2011) but information regarding its efficacy has not yet been released.
Despite a considerable effort spent on the development of selective MC4 receptor agonists to treat obesity, clinical trials have yet to demonstrate efficacy. There are a number of issues relating to the development of drugs targeting the melanocortin system, including the need for a full agonist at MC4 receptors with receptor subtype selectivity. Although POMC is discretely expressed within the CNS, it still has multiple roles. Furthermore, the MC4 receptor is widely distributed within the CNS and agonism has been demonstrated to increase blood pressure and sympathetic function (Greenfield, 2011). It is possible that the adverse effects associated with MC4 receptor agonism may be avoided by new agents selectively targeting the appetite regulating pathways, activated by MC4 receptors.
Ciliary neurotrophic factor
Ciliary neurotrophic factor (CNTF) is a 22kD protein expressed in Schwann cells and astrocytes, originally described as a trophic factor important for motor neuron survival (Lin et al., 1990; Sendtner et al., 1991). Investigations into its neuroprotective properties in the management of motor neurone disease revealed a significant 2.9% placebo-subtracted body weight loss in the non-obese participants (The ALS CNTF Treatment Study Group, 1996). Further studies demonstrated that CNTF and leptin act through a common intracellular signalling molecule:,the STAT3 transcription factor (Peterson et al., 2000). In addition, the CNTF receptor is expressed in the hypothalamic nuclei known to be involved in appetite regulation (Gloaguen et al., 1997). CNTF causes a 10–20% placebo-subtracted body weight loss in both leptin-deficient ob/ob mice and also in mouse models of DIO in which leptin administration had little effect (Gloaguen et al., 1997; Lambert et al., 2001). This mouse study reflects the disappointing results of leptin administration in obese humans, in whom circulating leptin levels are already elevated, suggesting a leptin-resistant state (Heymsfield et al., 1999). The potential for CNTF to bypass this resistance was recognized and a modified version of CNTF, axokine, proceeded to clinical trials. Subjects receiving axokine in a phase III trial demonstrated a 4.2 kg placebo-subtracted weight loss over 12 weeks; 25.1% of those receiving axokine lost greater than 5% of their baseline body weight. However, approximately 70% of recipients developed antibodies to the drug (Ettinger et al., 2003). While this was not associated with a loss of efficacy, concerns regarding the potential for antibodies to interfere with the neuroprotective role of endogenous CNTF led to Regeneron abandoning development of axokine as a potential anti-obesity agent.
Brain-derived neurotrophic factor (BDNF)
BDNF is a member of the neurotrophin family of growth factors (Leibrock et al., 1989). There is strong evidence that BDNF can regulate energy homeostasis and it is possible that BDNF possesses neuromodulatory properties in addition to its role as a growth factor (Nicholson et al., 2007). The biologically active form is mature BDNF, which mediates its effects via the tropomyosin-related kinase B (TrkB) receptor (Klein et al., 1991). BDNF is widely expressed throughout the CNS, including in several hypothalamic nuclei known to be involved in energy homeostasis, the ARC, PVN, VMN and DMN (Conner et al., 1997). Expression of BDNF mRNA in the VMH is positively regulated by leptin (Komori et al., 2006). Central injection of BDNF for 3 weeks reduces cumulative food intake by approximately 30% and causes a 15–20% body weight loss in rats, in addition to increasing locomotor activity and metabolic rate (Pelleymounter et al., 1995; Wang et al., 2007; 2010). Knockout of BDNF is embryonically lethal, but postnatal deletion of brain BDNF in mice is not and leads to an obese and hyperactive phenotype (Rios et al., 2001), and mice expressing significantly reduced levels of TrkB are hyperphagic (Xu et al., ). It has been postulated that BDNF lies downstream of MC4R signalling, and in accord with this, central BDNF administration was able to ameliorate obesity in MC4 receptor knockout mice (Xu et al., ).
Human studies also support a critical role of the BDNF/TrkB pathway in energy homeostasis. A human mutation causing loss of function of the TrkB receptor leads to hyperphagia and obesity (Yeo et al., 2004), and this phenotype is also seen in humans with haploinsufficiency for the BDNF gene (Gray et al., 2006; Han et al., 2008). Furthermore, genome-wide association studies have identified single nucleotide polymorphisms in the BDNF gene associated with a higher body mass index (Speliotes et al., 2010). A number of studies have demonstrated a role for BDNF/TrkB in hedonic feeding, regulating motivation and the reward (see Cordeira and Rios, 2011). Disruptions in BDNF signalling in this circuitry may also contribute to obesity.
BDNF also has an important role in neuronal plasticity and neurogenesis and a phase III clinical trial has been carried out investigating the clinical efficacy of BDNF in the treatment of motor neurone disease. Safety and tolerability were demonstrated in these studies; however, there was no significant survival advantage in subjects receiving BDNF (The BDNF Study Group, 1999). No effects of BDNF on body weight of trial participants were reported.
Further preclinical studies are required to elucidate the precise role of BDNF/TrkB signalling in hypothalamic neuronal populations. Additionally, the role of BDNF elsewhere in the brain influencing eating behaviour warrants further investigation. However, given that mutations that interfere with BDNF signalling cause obesity within the human population, BDNF is an important potential target in the treatment of obesity, and clinical trials are highly anticipated.
Pipeline therapies
Currently, the majority of the new drugs seeking approval for the treatment of obesity are based on classical neurotransmitter systems, despite the widespread nature of these systems leading to concerns regarding their potential side effects. Lorcaserin (Belviq) a selective 5-HT2C receptor agonist very recently received FDA approval for use in the treatment of obesity in select groups of obese patients. Placebo-subtracted weight loss of greater than 5% baseline was achieved by 27.2% and 68% of patients maintained that weight loss following 2 years of treatment (Smith et al., 2010b; Fidler et al., 2011). The mechanism behind the effects of lorcaserin on body weight is not completely understood, but may be mediated by hypothalamic POMC activation (Halford et al., 2007). Clinical trials revealed only mild side effects with lorcaserin treatment, most commonly headache (Smith et al., 2010b).
Contrave is a combination of bupropion and naltrexone, which is currently seeking FDA approval. Phase III trials demonstrated that Contrave, when given in conjunction with a diet and exercise programme over 56 weeks, was associated with a 4.8% placebo-subtracted body weight loss and 39–48% of subjects receiving Contrave lost greater than 5% of their baseline body weight (Greenway et al., 2010). Bupropion is a dopamine and noradrenaline reuptake inhibitor and is already used in the treatment of depression and smoking cessation, while naltrexone is an opioid receptor antagonist used to treat opiate and alcohol addiction. Both drugs have individually shown evidence of weight loss, with placebo-subtracted weight losses of 1.4% with naltrexone and 5.9% with bupropion (Greenway et al., 2009), and it is hypothesized that their combination will have a synergistic effect due to the different mechanisms by which they mediate weight loss. Bupropion stimulates hypothalamic POMC neurons, while naltrexone blocks the auto-inhibition of the POMC neurons by endogenous β-endorphins (Plodkowski et al., 2009). A large-scale study of the long-term cardiovascular effects of Contrave is required by the FDA before approval will be considered.
Qsymia (formerly Qnexa) is a combination of phentermine and topiramate, which has been shown to lead to 9.3% placebo-subtracted weight loss in overweight and obese subjects when combined with a diet and lifestyle modification programme (Gadde et al., 2011; Allison et al., 2012). Phentermine is a noradrenaline reuptake inhibitor, which has been shown to cause placebo-subtracted weight loss of 3.6 kg; topiramate an anticonvulsant, has been associated with a placebo-subtracted weight loss of 6.5% (Li et al., 2005). Concerns have been raised about dangerous side effects of Qsymia including suicidal thoughts, heart palpitations, memory lapses and birth defects (VIVUS, 2012); however, it has recently been approved by the FDA for the long-term management of weight loss (FDA, 2012).
Conclusions
There are many neuropeptide systems that influence food intake and energy homeostasis. Manipulation of several of these systems has been shown to reduce food intake and body weight in preclinical studies (Table 1). However, to date, there has been a disappointing lack of anti-obesity agents reaching clinical practice. Possible reasons for this paucity include the difficulties of such agents accessing the CNS, unacceptable side effects or a lack of efficacy. Although the ARC is incompletely isolated from the circulation by the blood brain barrier, targeting central neuropeptide systems in other regions of the CNS requires that drugs must cross the blood–brain barrier. Often a high dose of a drug is required to achieve sufficient penetrance of the CNS to mediate reductions in body weight, and this can lead to an increased risk of side effects.
While neuropeptide systems tend to be more discretely distributed and more restricted in their physiological function than the widespread neurotransmitter circuits, some, such as the NPY system, are equally widely distributed and are associated with a diverse range of functions. Drugs targeting widespread neuropeptide systems often have a limited therapeutic window, and targeting, for example, specific receptors within the NPY and melanocortin systems, or, the more discrete neuropeptide systems such as the GALP or relaxin-3 systems may represent more promising targets for future therapies. However, despite the discrete expression of POMC, GALP and relaxin-3, these systems have still been reported to be associated with multiple roles, and manipulation, as has been demonstrated with agents targeting the MC4R, may well be associated with side effects (Greenfield, 2011).
The development of therapeutic agents based on neuropeptide systems has been hampered by a lack of efficacy. This may in part be due to redundancy and adaptation to chronic administration protocols within the neuropeptide systems regulating energy balance. The lack of efficacy in humans of many of the drugs undergoing development contrasts with preclinical data from rodent models. Using animal models to examine the role of specific signals in energy homeostasis is necessary, but has limitations. Many of the side effects one might expect from higher doses of anorectic agents include nausea or changes in motivation, which can be difficult to measure in animals.
Peptide hormones are difficult to mimic with small molecules and therefore some of the drugs with clinical efficacy are not orally active. Injection of a drug is possible but the inconvenience and pain associated are more likely to lead to failures of compliance and complications associated with injection sites in patients. The use of long-acting injectable agents with reduced frequency of dosing may ameliorate the disadvantages associated with injectable therapy.
As with any drug based on a peptide, there is a chance that antibodies may develop against the drug. Where the endogenous peptide has important effects other than those relating to energy balance, this may lead to potentially disastrous effects. In addition, neuropeptide systems are often associated with redundancy, which may lead to a lack of efficacy of any single-target therapy. A combination therapy approach may therefore provide the best chance of an effective treatment for obesity.
It is therefore unclear whether neuropeptides can be used to treat obesity. However, despite the significant failure rate of novel anti-obesity agents, there are a number of potential therapies, which target hypothalamic neuropeptide systems that are currently progressing through clinical trials. It remains to be seen whether these agents have greater efficacy and/or more favourable side-effect profiles than those targeting neurotransmitter systems. The early results of these studies have shown promising weight-reducing effects and these agents may prove useful in the future clinical management of obesity.
All nomenclature in this review conforms to BJP's Guide to Receptors and Channels.
Acknowledgments
The section is funded by grants from the MRC, BBSRC, NIHR, an Integrative Mammalian Biology (IMB) Capacity Building Award, an FP7-HEALTH-2009-241592 EuroCHIP grant, and is supported by the NIHR Imperial Biomedical Research Centre Funding Scheme. Kevin G. Murphy is supported by project grants from the NIHR and BBSRC.
Glossary
- α-MSH
α-melanocyte-stimulating hormone
- ACTH
adrenocorticotropic hormone
- AgRP
agouti-related peptide
- ARC
arcuate nucleus
- BDNF
brain-derived neurotrophic factor
- CART
cocaine and amphetamine regulated transcript
- CNTF
ciliary neurotrophic factor
- DMN
dorsomedial nucleus
- EMEA
European Medicines Agency
- FDA
Food and Drugs Administration
- GALP
galanin-like peptide
- LHA
lateral hypothalamic area
- MCH
melanin-concentrating hormone
- MSH
melanocyte-stimulating hormone
- NPY
neuropeptide Y
- POMC
pro-opiomelanocortin
- PP
pancreatic polypeptide
- PVN
paraventricular nucleus
- VMN
ventromedial nucleus
Conflict of interest
The authors report no conflict of interests.
References
- Actelion. 2011. Actelion and GSK discontinue clinical development of Almorexant [press release], 28 January 2011. Available at: http://www.actelion.com/en/investors/media-releases/index.page?newsId=1483135 (accessed 15 August 2012)
- Adan RA, Oosterom J, Ludvigsdottir G, Brakkee JH, Burbach JP, Gispen WH. Identification of antagonists for melanocortin MC3, MC4 and MC5 receptors. Eur J Pharmacol. 1994;269:331–337. doi: 10.1016/0922-4106(94)90041-8. [DOI] [PubMed] [Google Scholar]
- Alexander SPH, Mathie A, Peters JA. Guide to receptors and channels (GRAC), 5th edition (2011) Br J Pharmacol. 2011;164(Suppl. 1):S1–S324. doi: 10.1111/j.1476-5381.2011.01649_1.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen JM, Yeats JC, Adrian TE, Bloom SR. Radioimmunoassay of neuropeptide Y. Regul Pept. 1984;8:61–70. doi: 10.1016/0167-0115(84)90029-6. [DOI] [PubMed] [Google Scholar]
- Allison DB, Gadde KM, Garvey WT, Peterson CA, Schwiers ML, Najarian T, et al. Controlled-release phentermine/topiramate in severely obese adults: a randomized controlled trial (EQUIP) Obesity (Silver Spring) 2012;20:330–342. doi: 10.1038/oby.2011.330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- AMRI. 2011. Available pipeline programs – obesity: MCH-1 antagonist program. [online] Available at: http://www.amriglobal.com/products_and_services/products_detail.cfm?prodID=7&subServID=67 (accessed 15 August 2012)
- Antal-Zimanyi I, Bruce MA, LeBoulluec KL, Iben LG, Mattson GK, McGovern RT, et al. Pharmacological characterization and appetite suppressive properties of BMS-193885, a novel and selective neuropeptide Y1 receptor antagonist. Eur J Pharmacol. 2008;590:224–232. doi: 10.1016/j.ejphar.2008.06.032. [DOI] [PubMed] [Google Scholar]
- Aston-Jones G, Smith RJ, Sartor GC, Moorman DE, Massi L, Tahsili-Fahadan P, et al. Lateral hypothalamic orexin/hypocretin neurons: a role in reward-seeking and addiction. Brain Res. 2010;1314:74–90. doi: 10.1016/j.brainres.2009.09.106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bai FL, Yamano M, Shiotani Y, Emson PC, Smith AD, Powell JF, et al. An arcuato-paraventricular and -dorsomedial hypothalamic neuropeptide Y-containing system which lacks noradrenaline in the rat. Brain Res. 1985;331:172–175. doi: 10.1016/0006-8993(85)90730-9. [DOI] [PubMed] [Google Scholar]
- Banks WA. The blood–brain barrier as a regulatory interface in the gut–brain axes. Physiol Behav. 2006;89:472–476. doi: 10.1016/j.physbeh.2006.07.004. [DOI] [PubMed] [Google Scholar]
- Bathgate RAD, Samuel CS, Burazin TCD, Layfield S, Claasz AA, Reytomas IGT, et al. Human relaxin gene 3 (H3) and the equivalent mouse relaxin (M3) gene. J Biol Chem. 2002;277:1148–1157. doi: 10.1074/jbc.M107882200. [DOI] [PubMed] [Google Scholar]
- Bertagna X. Proopiomelanocortin-derived peptides. Endocrinol Metab Clin North Am. 1994;23:467–485. [PubMed] [Google Scholar]
- Bewick GA, Gardiner JV, Dhillo WS, Kent AS, White NE, Webster Z, et al. Postembryonic ablation of AgRP neurons in mice leads to a lean, hypophagic phenotype. FASEB J. 2005;19:1680–1682. doi: 10.1096/fj.04-3434fje. [DOI] [PubMed] [Google Scholar]
- Biebermann H, Krude H, Elsner A, Chubanov V, Gudermann T, Grüters A. Autosomal-dominant mode of inheritance of a melanocortin-4 receptor mutation in a patient with severe early-onset obesity is due to a dominant-negative effect caused by receptor dimerization. Diabetes. 2003;52:2984–2988. doi: 10.2337/diabetes.52.12.2984. [DOI] [PubMed] [Google Scholar]
- Bittencourt JC, Presse F, Arias C, Peto C, Vaughan J, Nahon JL, et al. The melanin-concentrating hormone system of the rat brain: an immuno- and hybridization histochemical characterization. J Comp Neurol. 1992;319:218–245. doi: 10.1002/cne.903190204. [DOI] [PubMed] [Google Scholar]
- Blomqvist AG, Herzog H. Y-receptor subtypes – how many more? Trends Neurosci. 1997;20:294–298. doi: 10.1016/s0166-2236(96)01057-0. [DOI] [PubMed] [Google Scholar]
- Borowsky B, Durkin MM, Ogozalek K, Marzabadi MR, DeLeon J, Heurich R, et al. Antidepressant, anxiolytic and anorectic effects of a melanin-concentrating hormone-1 receptor antagonist. Nat Med. 2002;8:825–830. doi: 10.1038/nm741. [DOI] [PubMed] [Google Scholar]
- Bray GA. Pathophysiology of obesity. Am J Clin Nutr. 1992;55:488S–494S. doi: 10.1093/ajcn/55.2.488s. [DOI] [PubMed] [Google Scholar]
- Broberger C, Johansen J, Johansson C, Schalling M, Hökfelt T. The neuropeptide Y/agouti gene-related protein (AGRP) brain circuitry in normal, anorectic, and monosodium glutamate-treated mice. Proc Natl Acad Sci U S A. 1998;95:15043–15048. doi: 10.1073/pnas.95.25.15043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brothers SP, Wahlestedt C. Therapeutic potential of neuropeptide Y (NPY) receptor ligands. EMBO Mol Med. 2010;2:429–439. doi: 10.1002/emmm.201000100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burazin TCD, Bathgate RAD, Macris M, Layfield S, Gundlach AL, Tregear GW. Restricted, but abundant, expression of the novel rat gene-3 (R3) relaxin in the dorsal tegmental region of brain. J Neurochem. 2002;82:1553–1557. doi: 10.1046/j.1471-4159.2002.01114.x. [DOI] [PubMed] [Google Scholar]
- Butler AA, Kesteson RA, Khong K, Cullen MJ, Pelleymounter MA, Dekoning J, et al. A unique metabolic syndrome causes obesity in the melanocortin-3 receptor-deficient mouse. Endocrinology. 2000;141:3518–3521. doi: 10.1210/endo.141.9.7791. [DOI] [PubMed] [Google Scholar]
- Chemelli RM, Willie JT, Sinton CM, Elmquist JK, Scammell T, Lee C, et al. Narcolepsy in orexin knockout mice: molecular genetics of sleep regulation. Cell. 1999;98:437–451. doi: 10.1016/s0092-8674(00)81973-x. [DOI] [PubMed] [Google Scholar]
- Chen AS, Metzger JM, Trumbauer ME, X-m G, Yu H, Frazier EG, et al. Role of the melanocortin-4 receptor in metabolic rate and food intake in mice. Transgenic Res. 2000;9:145–154. doi: 10.1023/a:1008983615045. [DOI] [PubMed] [Google Scholar]
- Chen J, Kuei C, Sutton SW, Bonaventure P, Nepomuceno D, Eriste E, et al. Pharmacological characterization of relaxin-3/INSL7 receptors GPCR135 and GPCR142 from different mammalian species. J Pharmacol Exp Ther. 2005;312:83–95. doi: 10.1124/jpet.104.073486. [DOI] [PubMed] [Google Scholar]
- Cheung CC, Clifton DK, Steiner RA. Proopiomelanocortin neurons are direct targets for leptin in the hypothalamus. Endocrinology. 1997;138:4489–4492. doi: 10.1210/endo.138.10.5570. [DOI] [PubMed] [Google Scholar]
- Chronwall BM, DiMaggio DA, Massari VJ, Pickel VM, Ruggiero DA, O'Donohue TL. The anatomy of neuropeptide-y-containing neurons in rat brain. Neuroscience. 1985;15:1159–1181. doi: 10.1016/0306-4522(85)90260-x. [DOI] [PubMed] [Google Scholar]
- Clapham JC, Arch JRS, Tadayyon M. Anti-obesity drugs: a critical review of current therapies and future opportunities. Pharmacol Ther. 2001;89:81–121. doi: 10.1016/s0163-7258(00)00105-4. [DOI] [PubMed] [Google Scholar]
- Clark JT, Kalra PS, Crowley WR, Kalra SP. Neuropeptide Y and human pancreatic polypeptide stimulate feeding behavior in rats. Endocrinology. 1984;115:427–429. doi: 10.1210/endo-115-1-427. [DOI] [PubMed] [Google Scholar]
- ClinicalTrials.gov. 2011. Safety study of the inhibition of Agouti-related Protein (AgRP) for the management of obesity and weight loss. [online] Available at: http://clinicaltrials.gov/ct2/results?term=TTP-435 (accessed 15 August 2012)
- Cone RD, Cowley MA, Butler AA, Fan W, Marks DL, Low MJ. The arcuate nucleus as a conduit for diverse signals relevant to energy homeostasis. Int J Obes Relat Metab Disord. 2001;25(Suppl. 5):S63–S67. doi: 10.1038/sj.ijo.0801913. [DOI] [PubMed] [Google Scholar]
- Conner JM, Lauterborn JC, Yan Q, Gall CM, Varon S. Distribution of brain-derived neurotrophic factor (BDNF) protein and mRNA in the normal adult rat CNS: evidence for anterograde axonal transport. J Neurosci. 1997;17:2295–2313. doi: 10.1523/JNEUROSCI.17-07-02295.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Contreras RJ, Kosten T, Bird E. Area postrema: part of the autonomic circuitry of caloric homeostasis. Fed Proc. 1984;43:2966–2968. [PubMed] [Google Scholar]
- Cordeira J, Rios M. Weighing in the role of BDNF in the central control of eating behavior. Mol Neurobiol. 2011;44:441–448. doi: 10.1007/s12035-011-8212-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cox CD, Breslin MJ, Whitman DB, Schreier JD, McGaughey GB, Bogusky MJ, et al. Discovery of the dual orexin receptor antagonist [(7R)-4-(5-chloro-1,3-benzoxazol-2-yl)-7-methyl-1,4-diazepan-1-yl][5-methyl-2-(2H-1,2,3-triazol-2-yl)phenyl]methanone (MK-4305) for the treatment of Insomnia. J Med Chem. 2010;53:5320–5332. doi: 10.1021/jm100541c. [DOI] [PubMed] [Google Scholar]
- Criscione L, Rigollier P, Batzl-Hartmann C, Rüeger H, Stricker-Krongrad A, Wyss P, et al. Food intake in free-feeding and energy-deprived lean rats is mediated by the neuropeptide Y5 receptor. J Clin Invest. 1998;102:2136–2145. doi: 10.1172/JCI4188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daniels AJ, Grizzle MK, Wiard RP, Matthews JE, Heyer D. Food intake inhibition and reduction in body weight gain in lean and obese rodents treated with GW438014A, a potent and selective NPY-Y5 receptor antagonist. Regul Pept. 2002;106:47–54. doi: 10.1016/s0167-0115(02)00034-4. [DOI] [PubMed] [Google Scholar]
- Date Y, Ueta Y, Yamashita H, Yamaguchi H, Matsukura S, Kangawa K, et al. Orexins, orexigenic hypothalamic peptides, interact with autonomic, neuroendocrine and neuroregulatory systems. Proc Natl Acad Sci U S A. 1999;96:748–753. doi: 10.1073/pnas.96.2.748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Della-Zuana O, Presse F, Ortola C, Duhault J, Nahon JL, Levens N. Acute and chronic administration of melanin-concentrating hormone enhances food intake and body weight in Wistar and Sprague-Dawley rats. Int J Obes Relat Metab Disord. 2002;26:1289–1295. doi: 10.1038/sj.ijo.0802079. [DOI] [PubMed] [Google Scholar]
- Edwards CM, Abusnana S, Sunter D, Murphy KG, Ghatei MA, Bloom SR. The effect of the orexins on food intake: comparison with neuropeptide Y, melanin-concentrating hormone and galanin. J Endocrinol. 1999;160:R7–12. doi: 10.1677/joe.0.160r007. [DOI] [PubMed] [Google Scholar]
- Elias CF, Lee C, Kelly J, Aschkenasi C, Ahima RS, Couceyro PR, et al. Leptin activates hypothalamic CART neurons projecting to the spinal cord. Neuron. 1998a;21:1375–1385. doi: 10.1016/s0896-6273(00)80656-x. [DOI] [PubMed] [Google Scholar]
- Elias CF, Saper CB, Maratos-Flier E, Tritos NA, Lee C, Kelly J, et al. Chemically defined projections linking the mediobasal hypothalamus and the lateral hypothalamic area. J Comp Neurol. 1998b;402:442–459. [PubMed] [Google Scholar]
- Elmquist JK, Elias CF, Saper CB. From lesions to leptin: hypothalamic control of food intake and body weight. Neuron. 1999;22:221–232. doi: 10.1016/s0896-6273(00)81084-3. [DOI] [PubMed] [Google Scholar]
- EMEA. 2006. Guideline on clinical investigation of medicinal products used in weight control. [online] Available at: http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2009/09/WC500003213.pdf (accessed 15 August 2012)
- Erickson JC, Clegg KE, Palmiter RD. Sensitivity to leptin and susceptibility to seizures of mice lacking neuropeptide Y. Nature. 1996;381:415–421. doi: 10.1038/381415a0. [DOI] [PubMed] [Google Scholar]
- Erondu N, Gantz I, Musser B, Suryawanshi S, Mallick M, Addy C, et al. Neuropeptide Y5 receptor antagonism does not induce clinically meaningful weight loss in overweight and obese adults. Cell Metab. 2006;4:275–282. doi: 10.1016/j.cmet.2006.08.002. [DOI] [PubMed] [Google Scholar]
- Erondu N, Addy C, Lu K, Mallick M, Musser B, Gantz I, et al. NPY5R antagonism does not augment the weight loss efficacy of orlistat or sibutramine. Obesity. 2007a;15:2027–2042. doi: 10.1038/oby.2007.242. [DOI] [PubMed] [Google Scholar]
- Erondu N, Wadden T, Gantz I, Musser B, Nguyen AM, Bays H, et al. Effect of NPY5R antagonist MK-0557 on weight regain after very-low-calorie diet-induced weight loss. Obesity. 2007b;15:895–905. doi: 10.1038/oby.2007.620. [DOI] [PubMed] [Google Scholar]
- Ettinger MP, Littlejohn TW, Schwartz SL, Weiss SR, McIlwain HH, Heymsfield SB, et al. Recombinant variant of ciliary neurotrophic factor for weight loss in obese adults: a randomized, dose-ranging study. JAMA. 2003;289:1826–1832. doi: 10.1001/jama.289.14.1826. [DOI] [PubMed] [Google Scholar]
- FDA. 2007. Guidance for industry – developing products for weight management. [online] Available at: http://www.fda.gov/downloads/Drugs/…/Guidances/ucm071612.pdf (accessed 15 August 2012)
- FDA. 2012. FDA approves weight-management drug Qsymia. [press release], 17 July 2012. Available at: http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm312468.htm (accessed 15 August 2012)
- Fehm HL, Smolnik R, Kern W, McGregor GP, Bickel U, Born J. The melanocortin melanocyte-stimulating hormone/adrenocorticotropin4–10 decreases body fat in humans. J Clin Endocrinol Metab. 2001;86:1144–1148. doi: 10.1210/jcem.86.3.7298. [DOI] [PubMed] [Google Scholar]
- Fermini B, Fossa AA. The impact of drug-induced QT interval prolongation on drug discovery and development. Nat Rev Drug Discov. 2003;2:439–447. doi: 10.1038/nrd1108. [DOI] [PubMed] [Google Scholar]
- Fidler MC, Sanchez M, Raether B, Weissman NJ, Smith SR, Shanahan WR, et al. A one-year randomized trial of lorcaserin for weight loss in obese and overweight adults: the BLOSSOM trial. J Clin Endocrinol Metab. 2011;96:3067–3077. doi: 10.1210/jc.2011-1256. [DOI] [PubMed] [Google Scholar]
- Flach S, Williams E, Leonard L, Kochan R, Palleja S, Paul R, et al. 2007. Clinical safety and tolerability of S-2367 at doses up to 9600mg/day administered orally once a day for 7 days with an FDA high-fat caloric equivalent breakfast in healthy overweight and obese male and female subjects. The Obesity Society's 2007 Annual Scientific Meeting, October 20–24, New Orleans, Louisiana.
- Fong TM, Mao C, MacNeil T, Kalyani R, Smith T, Weinberg D, et al. ART (protein product of agouti-related transcript) as an antagonist of MC-3 and MC-4 receptors. Biochem Biophys Res Commun. 1997;237:629–631. doi: 10.1006/bbrc.1997.7200. [DOI] [PubMed] [Google Scholar]
- Foresight Report. 2007. Tackling obesities: future choices. [online] Available at: http://www.bis.gov.uk/foresight/our-work/projects/published-projects/tackling-obesities/reports-and-publications (accessed 15 August 2012)
- Fraley GS, Scarlett JM, Shimada I, Teklemichael DN, Acohido BV, Clifton DK, et al. Effects of diabetes and insulin on the expression of galanin-like peptide in the hypothalamus of the rat. Diabetes. 2004;53:1237–1242. doi: 10.2337/diabetes.53.5.1237. [DOI] [PubMed] [Google Scholar]
- Gadde KM, Allison DB, Ryan DH, Peterson CA, Troupin B, Schwiers ML, et al. Effects of low-dose, controlled-release, phentermine plus topiramate combination on weight and associated comorbidities in overweight and obese adults (CONQUER): a randomised, placebo-controlled, phase 3 trial. Lancet. 2011;377:1341–1352. doi: 10.1016/S0140-6736(11)60205-5. [DOI] [PubMed] [Google Scholar]
- Gardiner JV, Kong WM, Ward H, Murphy KG, Dhillo WS, Bloom SR. AAV mediated expression of anti-sense neuropeptide Y cRNA in the arcuate nucleus of rats results in decreased weight gain and food intake. Biochem Biophys Res Commun. 2005;327:1088–1093. doi: 10.1016/j.bbrc.2004.12.113. [DOI] [PubMed] [Google Scholar]
- Gautron L, Elmquist JK. Sixteen years and counting: an update on leptin in energy balance. J Clin Invest. 2011;121:2087–2093. doi: 10.1172/JCI45888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gloaguen I, Costa P, Demartis A, Lazzaro D, Di Marco A, Graziani R, et al. Ciliary neurotrophic factor corrects obesity and diabetes associated with leptin deficiency and resistance. Proc Natl Acad Sci U S A. 1997;94:6456–6461. doi: 10.1073/pnas.94.12.6456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glover ID, Barlow DJ, Pitts JE, Wood SP, Tickle IJ, Blundell TL, et al. Conformational studies on the pancreatic polypeptide hormone family. Eur J Biochem. 1984;142:379–385. doi: 10.1111/j.1432-1033.1984.tb08298.x. [DOI] [PubMed] [Google Scholar]
- Gomori A, Ishihara A, Ito M, Mashiko S, Matsushita H, Yumoto M, et al. Chronic intracerebroventricular infusion of MCH causes obesity in mice. Am J Physiol Endocrinol Metab. 2003;284:E583–E588. doi: 10.1152/ajpendo.00350.2002. [DOI] [PubMed] [Google Scholar]
- Graham M, Shutter JR, Sarmiento U, Sarosi I, Stark KL. Overexpression of Agrt leads to obesity in transgenic mice. Nat Genet. 1997;17:273–274. doi: 10.1038/ng1197-273. [DOI] [PubMed] [Google Scholar]
- Gray J, Yeo GS, Cox JJ, Morton J, Adlam AL, Keogh JM, et al. Hyperphagia, severe obesity, impaired cognitive function, and hyperactivity associated with functional loss of one copy of the brain-derived neurotrophic factor (BDNF) gene. Diabetes. 2006;55:3366–3371. doi: 10.2337/db06-0550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greenfield JR. Melanocortin signalling and the regulation of blood pressure in human obesity. J Neuroendocrinol. 2011;23:186–193. doi: 10.1111/j.1365-2826.2010.02088.x. [DOI] [PubMed] [Google Scholar]
- Greenway FL, Caruso MK. Safety of obesity drugs. Expert Opin Drug Saf. 2005;4:1083–1095. doi: 10.1517/14740338.4.6.1083. [DOI] [PubMed] [Google Scholar]
- Greenway FL, Dunayevich E, Tollefson G, Erickson J, Guttadauria M, Fujioka K, et al. Comparison of combined bupropion and naltrexone therapy for obesity with monotherapy and placebo. J Clin Endocrinol Metab. 2009;94:4898–4906. doi: 10.1210/jc.2009-1350. [DOI] [PubMed] [Google Scholar]
- Greenway FL, Fujioka K, Plodkowski RA, Mudaliar S, Guttadauria M, Erickson J, et al. Effect of naltrexone plus bupropion on weight loss in overweight and obese adults (COR-I): a multicentre, randomised, double-blind, placebo-controlled, phase 3 trial. Lancet. 2010;376:595–605. doi: 10.1016/S0140-6736(10)60888-4. [DOI] [PubMed] [Google Scholar]
- Griffith DA, Hargrove DM, Maurer TS, Blum CA, De Lombaert S, Inthavongsay JK, et al. Discovery and evaluation of pyrazolo[1,5-a]pyrimidines as neuropeptide Y1 receptor antagonists. Bioorg Med Chem Lett. 2011;21:2641–2645. doi: 10.1016/j.bmcl.2010.12.116. [DOI] [PubMed] [Google Scholar]
- Gropp E, Shanabrough M, Borok E, Xu AW, Janoschek R, Buch T, et al. Agouti-related peptide-expressing neurons are mandatory for feeding. Nat Neurosci. 2005;8:1289–1291. doi: 10.1038/nn1548. [DOI] [PubMed] [Google Scholar]
- Guzzo P, Surman M, Luche M, Hadden M, Henderson A, Jiang XW, et al. 2010. Pharmacology of ALB-127158(a), an antagonist of the MCH1 receptor for the treatment of obesity. Obesity 2010 28th Annual Scientific Meeting; San Diego, CA.
- Hahn TM, Breininger JF, Baskin DG, Schwartz MW. Coexpression of Agrp and NPY in fasting-activated hypothalamic neurons. Nat Neurosci. 1998;1:271–272. doi: 10.1038/1082. [DOI] [PubMed] [Google Scholar]
- Halford JC, Harrold JA, Boyland EJ, Lawton CL, Blundell JE. Serotonergic drugs: effects on appetite expression and use for the treatment of obesity. Drugs. 2007;67:27–55. doi: 10.2165/00003495-200767010-00004. [DOI] [PubMed] [Google Scholar]
- Hallschmid M, Smolnik R, McGregor G, Born J, Fehm HL. Overweight humans are resistant to the weight-reducing effects of melanocortin4–10. J Clin Endocrinol Metab. 2006;91:522–525. doi: 10.1210/jc.2005-0906. [DOI] [PubMed] [Google Scholar]
- Han JC, Liu QR, Jones M, Levinn RL, Menzie CM, Jefferson-George KS, et al. Brain-derived neurotrophic factor and obesity in the WAGR syndrome. N Engl J Med. 2008;359:918–927. doi: 10.1056/NEJMoa0801119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris GC, Wimmer M, Aston-Jones G. A role for lateral hypothalamic orexin neurons in reward seeking. Nature. 2005;437:556–559. doi: 10.1038/nature04071. [DOI] [PubMed] [Google Scholar]
- Hauptman JB, Jeunet FS, Hartmann D. Initial studies in humans with the novel gastrointestinal lipase inhibitor Ro 18-0647 (tetrahydrolipstatin) Am J Clin Nutr. 1992;55:309S–313S. doi: 10.1093/ajcn/55.1.309s. [DOI] [PubMed] [Google Scholar]
- Heymsfield SB, Greenberg AS, Fujioka K, Dixon RM, Kushner R, Hunt T, et al. Recombinant leptin for weight loss in obese and lean adults: a randomized, controlled, dose-escalation trial. JAMA. 1999;282:1568–1575. doi: 10.1001/jama.282.16.1568. [DOI] [PubMed] [Google Scholar]
- Hida T, Takahashi E, Shikata K, Hirohashi T, Sawai T, Seiki T, et al. Chronic intracerebroventricular administration of relaxin-3 increases body weight in rats. J Recept Signal Transduct Res. 2006;26:147–158. doi: 10.1080/10799890600623373. [DOI] [PubMed] [Google Scholar]
- Hoever P, Dorffner G, Benes H, Penzel T, Danker-Hopfe H, Barbanoj MJ, et al. Orexin receptor antagonism, a new sleep-enabling paradigm: a proof-of-concept clinical trial. Clin Pharmacol Ther. 2012;91:975–985. doi: 10.1038/clpt.2011.370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hommel JD, Trinko R, Sears RM, Georgescu D, Liu Z-W, Gao X-B, et al. Leptin receptor signaling in midbrain dopamine neurons regulates feeding. Neuron. 2006;51:801–810. doi: 10.1016/j.neuron.2006.08.023. [DOI] [PubMed] [Google Scholar]
- Hostetler ED, Sanabria-Bohorquez S, Fan H, Zeng Z, Gantert L, Williams M, et al. Synthesis, characterization, and monkey positron emission tomography (PET) studies of [18F]Y1-973, a PET trace for the neuropeptide Y Y1 receptor. Neuroimaging. 2011;54:2635–2642. doi: 10.1016/j.neuroimage.2010.11.014. [DOI] [PubMed] [Google Scholar]
- Huszar D, Lynch CA, Fairchild-Huntress V, Dunmore JH, Fang Q, Berkemeier LR, et al. Targeted disruption of the melanocortin-4 receptor results in obesity in mice. Cell. 1997;88:131–141. doi: 10.1016/s0092-8674(00)81865-6. [DOI] [PubMed] [Google Scholar]
- Ioannides-Demos LL, Piccenna L, McNeil JJ. Pharmacotherapies for obesity: past, current and future therapies. J Obes. 2011 doi: 10.1155/2011/179674. 2011: 179674. DOI: 10.1155/2011/179674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- James WPT, Astrup A, Finer N, Hilsted J, Kopelman P, Rössner S, et al. Effect of sibutramine on weight maintenance after weight loss: a randomised trial. Lancet. 2000;356:2119–2125. doi: 10.1016/s0140-6736(00)03491-7. [DOI] [PubMed] [Google Scholar]
- Johansson A, Fredriksson R, Winnergren S, Hulting AL, Schioth HB, Lindblom J. The relative impact of chronic food restriction and acute food deprivation on plasma hormone levels and hypothalamic neuropeptide expression. Peptides. 2008;29:1588–1595. doi: 10.1016/j.peptides.2008.04.018. [DOI] [PubMed] [Google Scholar]
- Jureus A, Cunningham MJ, McClain ME, Clifton DK, Steiner RA. Galanin-like peptide (GALP) is a target for regulation by leptin in the hypothalamus of the rat. Endocrinology. 2000;141:2703–2706. doi: 10.1210/endo.141.7.7669. [DOI] [PubMed] [Google Scholar]
- Jureus A, Cunningham MJ, Li D, Johnson LL, Krasnow SM, Teklemichael DN, et al. Distribution and regulation of galanin-like peptide (GALP) in the hypothalamus of the mouse. Endocrinology. 2001;142:5140–5144. doi: 10.1210/endo.142.12.8542. [DOI] [PubMed] [Google Scholar]
- Kakui N, Tanaka J, Tabata Y, Asai K, Masuda N, Miyara T, et al. Pharmacological characterization and feeding-suppressive property of FMS586 [3-(5,6,7,8-Tetrahydro-9-isopropyl-carbazol-3-yl)-1-methyl-1-(2-pyridin-4-yl-ethyl)-urea Hydrochloride], a novel, selective, and orally active antagonist for neuropeptide Y Y5 receptor. J Pharmacol Exp Ther. 2006;317:562–570. doi: 10.1124/jpet.105.099705. [DOI] [PubMed] [Google Scholar]
- Kameda M, Kobayashi K, Ito H, Miyazoe H, Tsujino T, Nakama C, et al. Optimization of a series of 2,4-diaminopyridines as neuropeptide Y Y1 receptor antagonists with reduced hERG activity. Bioorg Med Chem Lett. 2009;19:4325–4329. doi: 10.1016/j.bmcl.2009.05.069. [DOI] [PubMed] [Google Scholar]
- Kamegai J, Tamura H, Shimizu T, Ishii S, Sugihara H, Wakabayashi I. Chronic central infusion of ghrelin increases hypothalamic neuropeptide Y and Agouti-related protein mRNA levels and body weight in rats. Diabetes. 2001;50:2438–2443. doi: 10.2337/diabetes.50.11.2438. [DOI] [PubMed] [Google Scholar]
- Kanatani A, Ishihara A, Asahi S, Tanaka T, Ozaki S, Ihara M. Potent neuropeptide Y Y1 receptor antagonist, 1229U91: blockade of neuropeptide Y-induced and physiological food intake. Endocrinology. 1996;137:3177–3182. doi: 10.1210/endo.137.8.8754736. [DOI] [PubMed] [Google Scholar]
- Kanatani A, Hata M, Mashiko S, Ishihara A, Okamoto O, Haga Y, et al. A typical Y1 receptor regulates feeding behaviors: effects of a potent and selective Y1 antagonist, J-115814. Mol Pharmacol. 2001;59:501–505. doi: 10.1124/mol.59.3.501. [DOI] [PubMed] [Google Scholar]
- Kievit P, Halem HA, Pranger L, Cowley MA, Marks DL, Grove KL, et al. A novel MC4R agonist reduces body weight and improves glucose homeostasis in high fat diet induced obese rhesus macaques. Endocr Rev. 2010;31(3 Suppl. 1):P2–P257. [Google Scholar]
- Kim SH, Lee YM, Jee SH, Nam CM. Effect of sibutramine on weight loss and blood pressure: a meta-analysis of controlled trials. Obes Res. 2003;11:1116–1123. doi: 10.1038/oby.2003.152. [DOI] [PubMed] [Google Scholar]
- Klein R, Nanduri V, Jing S, Lamballe F, Tapley P, Bryant S, et al. The trkB tyrosine protein kinase is a receptor for brain-derived neurotrophic factor and neurotrophin-3. Cell. 1991;66:395–403. doi: 10.1016/0092-8674(91)90628-c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knigge KM, Wagner JE. Melanin-concentrating hormone (MCH) involvement in pentylenetetrazole (PTZ)-induced seizure in rat and guinea pig. Peptides. 1997;18:1095–1097. doi: 10.1016/s0196-9781(97)00036-3. [DOI] [PubMed] [Google Scholar]
- Kohno D, Gao H-Z, Muroya S, Kikuyama S, Yada T. Ghrelin directly interacts with neuropeptide-Y-containing neurons in the rat arcuate nucleus. Diabetes. 2003;52:948–956. doi: 10.2337/diabetes.52.4.948. [DOI] [PubMed] [Google Scholar]
- Komori T, Morikawa Y, Nanjo K, Senba E. Induction of brain-derived neurotrophic factor by leptin in the ventromedial hypothalamus. Neuroscience. 2006;139:1107–1115. doi: 10.1016/j.neuroscience.2005.12.066. [DOI] [PubMed] [Google Scholar]
- Kopelman PG. Obesity as a medical problem. Nature. 2000;404:635–643. doi: 10.1038/35007508. [DOI] [PubMed] [Google Scholar]
- Kowalski TJ, Farley C, Cohen-Williams ME, Varty G, Spar BD. Melanin-concentrating hormone-1 receptor antagonism decreases feeding be reducing meal size. Eur J Pharmacol. 2004;497:41–47. doi: 10.1016/j.ejphar.2004.06.027. [DOI] [PubMed] [Google Scholar]
- Krishna R, Gumbiner B, Stevens C, Musser B, Mallick M, Suryawanshi S, et al. Potent and selective agonism of the melanocortin receptor 4 with MK-0493 does not induce weight loss in obese human subjects: energy intake predicts lack of weight loss efficacy. Clin Pharmacol Ther. 2009;86:659–666. doi: 10.1038/clpt.2009.167. [DOI] [PubMed] [Google Scholar]
- Kristensen P, Judge ME, Thim L, Ribel U, Christjansen KN, Wulff BS, et al. Hypothalamic CART is a new anorectic peptide regulated by leptin. Nature. 1998;393:72–76. doi: 10.1038/29993. [DOI] [PubMed] [Google Scholar]
- Krude H, Biebermann H, Schnabel D, Tansek MZ, Theunissen P, Mullis PE, et al. Obesity due to proopiomelanocortin deficiency: three new cases and treatment trials with thyroid hormone and ACTH4–10. J Clin Endocrinol Metab. 2003;88:4633–4640. doi: 10.1210/jc.2003-030502. [DOI] [PubMed] [Google Scholar]
- Kuei C, Sutton S, Bonaventure P, Pudiak C, Shelton J, Zhu J, et al. R3(BΔ23–27)R/I5 chimeric peptide, a selective antagonist for GPCR135 and GPCR142 over relaxin receptor LGR7. J Biol Chem. 2007;282:25425–25435. doi: 10.1074/jbc.M701416200. [DOI] [PubMed] [Google Scholar]
- Kym PR, Souers AJ, Campbell TJ, Lynch JK, Judd AS, Iyengar R, et al. Screening for cardiovascular safety: a structure−activity approach for guiding lead selection of melanin concentrating hormone receptor 1 antagonists. J Med Chem. 2006;49:2339–2352. doi: 10.1021/jm0512286. [DOI] [PubMed] [Google Scholar]
- Lambert PD, Anderson KD, Sleeman MW, Wong V, Tan J, Hijarunguru A, et al. Ciliary neurotrophic factor activates leptin-like pathways and reduces body fat, without cachexia or rebound weight gain, even in leptin-resistant obesity. Proc Natl Acad Sci U S A. 2001;98:4652–4657. doi: 10.1073/pnas.061034298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang R, Gundlach AL, Kofler B. The galanin peptide family: receptor pharmacology, pleiotropic biological actions, and implications in health and disease. Pharmacol Ther. 2007;115:177–207. doi: 10.1016/j.pharmthera.2007.05.009. [DOI] [PubMed] [Google Scholar]
- Larm JA, Gundlach AL. Galanin-like peptide (GALP) mRNA expression is restricted to arcuate nucleus of hypothalamus in adult male rat brain. Neuroendocrinology. 2000;72:67–71. doi: 10.1159/000054573. [DOI] [PubMed] [Google Scholar]
- Lawrence CB, Baudoin FM, Luckman SM. Centrally administered galanin-like peptide modifies food intake in the rat: a comparison with galanin. J Neuroendocrinol. 2002;14:853–860. doi: 10.1046/j.1365-2826.2002.00846.x. [DOI] [PubMed] [Google Scholar]
- de Lecea L, Kilduff TS, Peyron C, Gao X-B, Foye PE, Danielson PE, et al. The hypocretins: hypothalamus-specific peptides with neuroexcitatory activity. Proc Natl Acad Sci U S A. 1998;95:322–327. doi: 10.1073/pnas.95.1.322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leibel RL. Is obesity due to a heritable difference in ‘set point’ for adiposity? West J Med. 1990;153:429–431. [PMC free article] [PubMed] [Google Scholar]
- Leibrock J, Lottspeich F, Hohn A, Hofer M, Hengerer B, Masiakowski P, et al. Molecular cloning and expression of brain-derived neurotrophic factor. Nature. 1989;341:149–152. doi: 10.1038/341149a0. [DOI] [PubMed] [Google Scholar]
- Li Z, Maglione M, Tu W, Mojica W, Arterburn D, Shugarman LR, et al. Meta-analysis: pharmacologic treatment of obesity. Ann Intern Med. 2005;142:532–546. doi: 10.7326/0003-4819-142-7-200504050-00012. [DOI] [PubMed] [Google Scholar]
- Lin LF, Armes LG, Sommer A, Smith DJ, Collins F. Isolation and characterization of ciliary neurotrophic factor from rabbit sciatic nerves. J Biol Chem. 1990;265:8942–8947. [PubMed] [Google Scholar]
- Liu C, Chen J, Sutton S, Roland B, Kuei C, Farmer N, et al. Identification of relaxin-3/INSL7 as a ligand for GPCR142. J Biol Chem. 2003a;278:50765–50770. doi: 10.1074/jbc.M308996200. [DOI] [PubMed] [Google Scholar]
- Liu C, Eriste E, Sutton S, Chen J, Roland B, Kuei C, et al. Identification of relaxin-3/INSL7 as an endogenous ligand for the orphan G-protein-coupled receptor GPCR135. J Biol Chem. 2003b;278:50754–50764. doi: 10.1074/jbc.M308995200. [DOI] [PubMed] [Google Scholar]
- Ludwig DS, Tritos NA, Mastaitis JW, Kulkarni R, Kokkotou E, Elmquist J, et al. Melanin-concentrating hormone overexpression in transgenic mice leads to obesity and insulin resistance. J Clin Invest. 2001;107:379–386. doi: 10.1172/JCI10660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luquet S, Perez FA, Hnasko TS, Palmiter RD. NPY/AgRP neurons are essential for feeding in adult mice but can be ablated in neonates. Science. 2005;310:683–685. doi: 10.1126/science.1115524. [DOI] [PubMed] [Google Scholar]
- Luthin DR. Anti-obesity effects of small molecule melanin-concentrating hormone receptor1 (MCHR1) antagonists. Life Sci. 2007;81:423–440. doi: 10.1016/j.lfs.2007.05.029. [DOI] [PubMed] [Google Scholar]
- Lynch JK, Freeman JC, Judd AS, Iyengar R, Mulhern M, Zhao G, et al. Optimization of chromone-2-carboxamide melanin concentrating hormone receptor 1 antagonists: assessment of potency, efficacy, and cardiovascular safety. J Med Chem. 2006;49:6569–6584. doi: 10.1021/jm060683e. [DOI] [PubMed] [Google Scholar]
- McGowan BMC, Stanley SA, Smith KL, White NE, Connolly MM, Thompson EL, et al. Central relaxin-3 administration causes hyperphagia in male Wistar rats. Endocrinology. 2005;146:3295–3300. doi: 10.1210/en.2004-1532. [DOI] [PubMed] [Google Scholar]
- McGowan BM, Stanley SA, Smith KL, Minnion JS, Donovan J, Thompson EL, et al. Effects of acute and chronic relaxin-3 on food intake and energy expenditure in rats. Regul Pept. 2006;136:72–77. doi: 10.1016/j.regpep.2006.04.009. [DOI] [PubMed] [Google Scholar]
- Marsh DJ, Hollopeter G, Kafer KE, Palmiter RD. Role of the Y5 neuropeptide Y receptor in feeding and obesity. Nat Med. 1998;4:718–721. doi: 10.1038/nm0698-718. [DOI] [PubMed] [Google Scholar]
- Marsh DJ, Weingarth DT, Novi DE, Chen HY, Trumbauer ME, Chen AS, et al. Melanin-concentrating hormone 1 receptor-deficient mice are lean, hyperactive, and hyperphagic and have altered metabolism. Proc Natl Acad Sci U S A. 2002;99:3240–3245. doi: 10.1073/pnas.052706899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mashiko S, Ishihara A, Iwaasa H, Moriya R, Kitazawa H, Mitobe Y, et al. Effects of a novel Y5 antagonist in obese mice: combination with food restriction or sibutramine. Obesity. 2008;16:1510–1515. doi: 10.1038/oby.2008.223. [DOI] [PubMed] [Google Scholar]
- Mashiko S, Moriya R, Ishihara A, Gomori A, Matsushita H, Egashira S, et al. Synergistic interaction between neuropeptide Y1 and Y5 receptor pathways in regulation of energy homeostasis. Eur J Pharmacol. 2009;615:113–117. doi: 10.1016/j.ejphar.2009.05.018. [DOI] [PubMed] [Google Scholar]
- Matsumoto Y, Watanabe T, Adachi Y, Itoh T, Ohtaki T, Onda H, et al. Galanin-like peptide stimulates food intake in the rat. Neurosci Lett. 2002;322:67–69. doi: 10.1016/s0304-3940(01)02515-0. [DOI] [PubMed] [Google Scholar]
- Miller KJ, Murphy BJ, Pelleymounter MA. Central G-protein coupled receptors (GPCR)s as molecular targets for the treatment of obesity: assets, liabilities and development status. Curr Drug Target CNS Neurol Disord. 2004;3:357–377. doi: 10.2174/1568007043337003. [DOI] [PubMed] [Google Scholar]
- Moore NA, Surman M, Guzzo P, Sargent BJ, Silverman MH, Blundell J, et al. 2011. MCH1 receptor antagonist, ALB-127158(a): phase I safety, tolerability and activity in lean and overweight volunteersAMRI, Albany NY, USA.
- Murphy KG, Dhillo WS, Bloom SR. Gut peptides in the regulation of food intake and energy homeostasis. Endocr Rev. 2006;27:719–727. doi: 10.1210/er.2006-0028. [DOI] [PubMed] [Google Scholar]
- NCATS. 2012. BMS-830216 [online] Available at: http://www.ncats.nih.gov/files/BMS-830216.pdf (accessed 15 August 2012)
- Neurogen. 2007. Neurogen announces results of first-in-human trial for new approach to treating obesity [press release], 2 May 2007, Available at: http://www.businesswire.com/news/home/20070502005240/en/Neurogen-Announces-Results-First-in-Human-Trial-Approach-Treating (Accessed 15 August 2012)
- Nicholson JR, Peter JC, Lecourt AC, Barde YA, Hofbauer KG. Melanocortin-4 receptor activation stimulates hypothalamic brain-derived neurotrophic factor release to regulate food intake, body temperature and cardiovascular function. J Neuroendocrinol. 2007;19:974–982. doi: 10.1111/j.1365-2826.2007.01610.x. [DOI] [PubMed] [Google Scholar]
- Nishino S, Ripley B, Overeem S, Lammers GJ, Mignot E. Hypocretin (orexin) deficiency in human narcolepsy. Lancet. 2000;355:39–40. doi: 10.1016/S0140-6736(99)05582-8. [DOI] [PubMed] [Google Scholar]
- Ohtaki T, Kumano S, Ishibashi Y, Ogi K, Matsui H, Harada M, et al. Isolation and cDNA cloning of a novel galanin-like peptide (GALP) from porcine hypothalamus. J Biol Chem. 1999;274:37041–37045. doi: 10.1074/jbc.274.52.37041. [DOI] [PubMed] [Google Scholar]
- Ollmann MM, Wilson BD, Yang Y-K, Kerns JA, Chen Y, Gantz I, et al. Antagonism of central melanocortin receptors in vitro and in vivo by agouti-related protein. Science. 1997;278:135–138. doi: 10.1126/science.278.5335.135. [DOI] [PubMed] [Google Scholar]
- Omori N, Kouyama N, Yukimasa A, Watanabe K, Yokota Y, Tanioka H, et al. Hit to lead SAR study on benzoxazole derivatives for an NPY Y5 antagonist. Bioorg Med Chem Lett. 2012;22:2020–2023. doi: 10.1016/j.bmcl.2012.01.027. [DOI] [PubMed] [Google Scholar]
- Owen RT, Castaner R, Bolos J, Estivill C. Almorexant. Drugs Future. 2009;34:5–10. [Google Scholar]
- Palatin Technologies. (2012). Palatin Technologies announces halting of phase I clinical trial of obesity compound in AstraZeneca Research Collaboration. [press release], 19 June 2012. Available at: http://www.palatin.com (accessed 15 August 2012)
- Pedrazzini T, Seydoux J, Kunstner P, Aubert JF, Grouzmann E, Beermann F, et al. Cardiovascular response, feeding behavior and locomotor activity in mice lacking the NPY Y1 receptor. Nat Med. 1998;4:722–726. doi: 10.1038/nm0698-722. [DOI] [PubMed] [Google Scholar]
- Pelleymounter MA, Cullen MJ, Wellman CL. Characteristics of BDNF-induced weight loss. Exp Neurol. 1995;131:229–238. doi: 10.1016/0014-4886(95)90045-4. [DOI] [PubMed] [Google Scholar]
- Peterson WM, Wang Q, Tzekova R, Wiegand SJ. Ciliary neurotrophic factor and stress stimuli activate the Jak-STAT pathway in retinal neurons and glia. J Neurosci. 2000;20:4081–4090. doi: 10.1523/JNEUROSCI.20-11-04081.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plodkowski RA, Nguyen Q, Sundaram U, Nguyen L, Chau DL, St Jeor S. Bupropion and naltrexone: a review of their use individually and in combination for the treatment of obesity. Expert Opin Pharmacother. 2009;10:1069–1081. doi: 10.1517/14656560902775750. [DOI] [PubMed] [Google Scholar]
- Puopolo A, Heshka S, Karmally W, Alvarado R, Kakudo S, Ochiai T, et al. Obesity Soc. Ann. Sci. Meeting. Washington DC: 2009. A yearlong study of the efficacy and safety of 0, 800, or 1,600 mg/day of velneperit (S-2367) with a reduced calorie diet (RCD) in obese subjects; p. 214. [Google Scholar]
- Qian S, Chen H, Weingarth D, Trumbauer ME, Novi DE, Guan X, et al. Neither agouti-related protein nor neuropeptide Y is critically required for the regulation of energy homeostasis in mice. Mol Cell Biol. 2002;22:5027–5035. doi: 10.1128/MCB.22.14.5027-5035.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qu D, Ludwig DS, Gammeltoft S, Piper M, Pelleymounter MA, Cullen MJ, et al. A role for melanin-concentrating hormone in the central regulation of feeding behaviour. Nature. 1996;380:243–247. doi: 10.1038/380243a0. [DOI] [PubMed] [Google Scholar]
- Rhythm Pharmaceuticals. 2011. RM-493: MC4R peptide therapeutic. [online] Available at: http://www.rhythmtx.com/PROGRAMS/RM493.html (accessed 15 August 2012)
- Rios M, Fan G, Fekete C, Kelly J, Bates B, Kuehn R, et al. Conditional deletion of brain-derived neurotrophic factor in the postnatal brain leads to obesity and hyperactivity. Mol Endocrinol. 2001;15:1748–1757. doi: 10.1210/mend.15.10.0706. [DOI] [PubMed] [Google Scholar]
- Rossi M, Kim MS, Morgan DG, Small CJ, Edwards CM, Sunter D, et al. A C-terminal fragment of Agouti-related protein increases feeding and antagonizes the effect of alpha-melanocyte stimulating hormone in vivo. Endocrinology. 1998;139:4428–4431. doi: 10.1210/endo.139.10.6332. [DOI] [PubMed] [Google Scholar]
- Sahu A, Kalra PS, Kalra SP. Food deprivation and ingestion induce reciprocal changes in neuropeptide Y concentrations in the paraventricular nucleus. Peptides. 1988;9:83–86. doi: 10.1016/0196-9781(88)90013-7. [DOI] [PubMed] [Google Scholar]
- Sahu A, Dube MG, Phelps CP, Sninsky CA, Kalra PS, Kalra SP. Insulin and insulin-like growth factor II suppress neuropeptide Y release from the nerve terminals in the paraventricular nucleus: a putative hypothalamic site for energy homeostasis. Endocrinology. 1995;136:5718–5724. doi: 10.1210/endo.136.12.7588328. [DOI] [PubMed] [Google Scholar]
- Saito Y, Nagasaki H. The melanin-concentrating hormone system and its physiological functions. Results Probl Cell Differ. 2008;46:159–179. doi: 10.1007/400_2007_052. [DOI] [PubMed] [Google Scholar]
- Sakurai T, Amemiya A, Ishii M, Matsuzaki I, Chemelli RM, Tanaka H, et al. Orexins and orexin receptors: a family of hypothalamic neuropeptides and g protein-coupled receptors that regulate feeding behavior. Cell. 1998;92:573–585. doi: 10.1016/s0092-8674(00)80949-6. [DOI] [PubMed] [Google Scholar]
- Sato N, Jitsuoka M, Shibata T, Hirohashi T, Nonoshita K, Moriya M, et al. (9S)-9-(2-hydroxy-4,4-dimethyl-6-oxo-1-cyclohexen-1-yl)-3,3-dimethyl-2,3,4,9-tetrahydro-1H-xanthen-1-one, a selective and orally active neuropeptide Y Y5 receptor antagonist. J Med Chem. 2008;51:4765–4770. doi: 10.1021/jm8003587. [DOI] [PubMed] [Google Scholar]
- Satoh N, Ogawa Y, Katsuura G, Hayase M, Tsuji T, Imagawa K, et al. The arcuate nucleus as a primary site of satiety effect of leptin in rats. Neurosci Lett. 1997;224:149–152. doi: 10.1016/S0304-3940(97)00163-8. [DOI] [PubMed] [Google Scholar]
- Schwartz MW, Sipols AJ, Marks JL, Sanacora G, White JD, Scheurink A, et al. Inhibition of hypothalamic neuropeptide Y gene expression by insulin. Endocrinology. 1992;130:3608–3616. doi: 10.1210/endo.130.6.1597158. [DOI] [PubMed] [Google Scholar]
- Sendtner M, Arakawa Y, Stockli KA, Kreutzberg GW, Thoenen H. Effect of ciliary neurotrophic factor (CNTF) on motoneuron survival. J Cell Sci Suppl. 1991;15:103–109. doi: 10.1242/jcs.1991.supplement_15.14. [DOI] [PubMed] [Google Scholar]
- Shiba K, Kageyama H, Takenoya F, Shioda S. Galanin-like peptide and the regulation of feeding behaviour and energy metabolism. FEBS J. 2010;277:5006–5013. doi: 10.1111/j.1742-4658.2010.07933.x. [DOI] [PubMed] [Google Scholar]
- Shimada M, Tritos NA, Lowell BB, Flier JS, Maratos-Flier E. Mice lacking melanin-concentrating hormone are hypophagic and lean. Nature. 1998;396:670–674. doi: 10.1038/25341. [DOI] [PubMed] [Google Scholar]
- Sipols AJ, Baskin DG, Schwartz MW. Effect of intracerebroventricular insulin infusion on diabetic hyperphagia and hypothalamic neuropeptide gene expression. Diabetes. 1995;44:147–151. doi: 10.2337/diab.44.2.147. [DOI] [PubMed] [Google Scholar]
- Sjöström L, Rissanen A, Andersen T, Boldrin M, Golay A, Koppeschaar HPF, et al. Randomised placebo-controlled trial of orlistat for weight loss and prevention of weight regain in obese patients. Lancet. 1998;352:167–172. doi: 10.1016/s0140-6736(97)11509-4. [DOI] [PubMed] [Google Scholar]
- Smith AI, Funder JW. Proopiomelanocortin processing in the pituitary, central nervous system, and peripheral tissues. Endocr Rev. 1988;9:159–179. doi: 10.1210/edrv-9-1-159. [DOI] [PubMed] [Google Scholar]
- Smith D, Heshka S, Karmally W, Doepner D, Narukawa Y, Ochiai T, et al. 2009. A yearlong study of the efficacy and safety of placebo or 1,600 mg/day of velneperit (S-2367) with an initial 6-week low calorie diet (LCD) in obese subjects. Obesity Soc. Ann. Sci. Meeting. pp 221-P.
- Smith CM, Shen P-J, Banerjee A, Bonaventure P, Ma S, Bathgate RAD, et al. Distribution of relaxin-3 and RXFP3 within arousal, stress, affective, and cognitive circuits of mouse brain. J Comp Neurol. 2010a;518:4016–4045. doi: 10.1002/cne.22442. [DOI] [PubMed] [Google Scholar]
- Smith SR, Weissman NJ, Anderson CM, Sanchez M, Chuang E, Stubbe S, et al. Multicenter, placebo-controlled trial of lorcaserin for weight management. N Engl J Med. 2010b;363:245–256. doi: 10.1056/NEJMoa0909809. [DOI] [PubMed] [Google Scholar]
- Speliotes EK, Willer CJ, Berndt SI, Monda KL, Thorleifsson G, Jackson AU, et al. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat Genet. 2010;42:937–948. doi: 10.1038/ng.686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stanley BG, Leibowitz SF. Neuropeptide Y: stimulation of feeding and drinking by injection into the paraventricular nucleus. Life Sci. 1984;35:2635–2642. doi: 10.1016/0024-3205(84)90032-8. [DOI] [PubMed] [Google Scholar]
- Stanley BG, Kyrkouli SE, Lampert S, Leibowitz SF. Neuropeptide Y chronically injected into the hypothalamus: a powerful neurochemical inducer of hyperphagia and obesity. Peptides. 1986;7:1189–1192. doi: 10.1016/0196-9781(86)90149-x. [DOI] [PubMed] [Google Scholar]
- Stephens TW, Basinski M, Bristow PK, Bue-Valleskey JM, Burgett SG, Craft L, et al. The role of neuropeptide Y in the antiobesity action of the obese gene product. Nature. 1995;377:530–532. doi: 10.1038/377530a0. [DOI] [PubMed] [Google Scholar]
- Sudo S, Kumagai J, Nishi S, Layfield S, Ferraro T, Bathgate RAD, et al. H3 relaxin is a specific ligand for LGR7 and activates the receptor by interacting with both the ectodomain and the exoloop 2. J Biol Chem. 2003;278:7855–7862. doi: 10.1074/jbc.M212457200. [DOI] [PubMed] [Google Scholar]
- Sutton SW, Shelton J, Smith C, Williams J, Yun S, Motley T, et al. Metabolic and neuroendocrine responses to RXFP3 modulation in the central nervous system. Ann N Y Acad Sci. 2009;1160:242–249. doi: 10.1111/j.1749-6632.2008.03812.x. [DOI] [PubMed] [Google Scholar]
- Takatsu Y, Matsumoto H, Ohtaki T, Kumano S, Kitada C, Onda H, et al. Distribution of galanin-like peptide in the rat brain. Endocrinology. 2001;142:1626–1634. doi: 10.1210/endo.142.4.8089. [DOI] [PubMed] [Google Scholar]
- Takekawa S, Asami A, Ishihara Y, Terauchi J, Kato K, Shimomura Y, et al. T-226296: a novel, orally active and selective melanin-concentrating hormone receptor antagonist. Eur J Pharmacol. 2002;438:129–135. doi: 10.1016/s0014-2999(02)01314-6. [DOI] [PubMed] [Google Scholar]
- Tan CP, Sano H, Iwaasa H, Pan J, Sailer AW, Hreniuk DL, et al. Melanin-concentrating hormone receptor subtypes 1 and 2: species-specific gene expression. Genomics. 2002;79:785–792. doi: 10.1006/geno.2002.6771. [DOI] [PubMed] [Google Scholar]
- Tan HM, Gundlach AL, Morris MJ. Exaggerated feeding response to central galanin-like peptide administration in diet-induced obese rats. Neuropeptides. 2005;39:333–336. doi: 10.1016/j.npep.2004.12.025. [DOI] [PubMed] [Google Scholar]
- Tanaka M, Iijima N, Miyamoto Y, Fukusumi S, Itoh Y, Ozawa H, et al. Neurons expressing relaxin 3/INSL 7 in the nucleus incertus respond to stress. Eur J Neurosci. 2005;21:1659–1670. doi: 10.1111/j.1460-9568.2005.03980.x. [DOI] [PubMed] [Google Scholar]
- Tang-Christensen M, Holst JJ, Hartmann B, Vrang N. The arcuate nucleus is pivotal in mediating the anorectic effects of centrally administered leptin. Neuroreport. 1999;10:1183–1187. doi: 10.1097/00001756-199904260-00005. [DOI] [PubMed] [Google Scholar]
- The ALS CNTF Treatment Study Group. A double-blind placebo-controlled clinical trial of subcutaneous recombinant human ciliary neurotrophic factor (rHCNTF) in amyotrophic lateral sclerosis. Neurology. 1996;46:1244–1249. doi: 10.1212/wnl.46.5.1244. [DOI] [PubMed] [Google Scholar]
- The BDNF Study Group. A controlled trial of recombinant methionyl human BDNF in ALS: (Phase III) Neurology. 1999;52:1427–1433. doi: 10.1212/wnl.52.7.1427. [DOI] [PubMed] [Google Scholar]
- Tokita S, Chemelli RM, Willie JT, Yanagisawa M. Behavioural characterization of orexin-2 receptor (OX2R) knockout mice [Abstract] Sleep. 2001;24(suppl):A20. [Google Scholar]
- Topol EJ, Bousser M-G, Fox KAA, Creager MA, Despres J-P, Easton JD, et al. Rimonabant for prevention of cardiovascular events (CRESCENDO): a randomised, multicentre, placebo-controlled trial. Lancet. 2010;376:517–523. doi: 10.1016/S0140-6736(10)60935-X. [DOI] [PubMed] [Google Scholar]
- Tsuda K, Yokoo H, Goldstein M. Neuropeptide Y and galanin in norepinephrine release in hypothalamic slices. Hypertension. 1989;14:81–86. doi: 10.1161/01.hyp.14.1.81. [DOI] [PubMed] [Google Scholar]
- Tsujii S, Bray GA. Acetylation alters the feeding response to MSH and beta-endorphin. Brain Res Bull. 1989;23:165–169. doi: 10.1016/0361-9230(89)90142-1. [DOI] [PubMed] [Google Scholar]
- Vaisse C, Clement K, Guy-Grand B, Froguel P. A frameshift mutation in human MC4R is associated with a dominant form of obesity. Nat Genet. 1998;20:113–114. doi: 10.1038/2407. [DOI] [PubMed] [Google Scholar]
- Vaisse C, Clement K, Durand E, Hercberg S, Guy-Grand B, Froguel P. Melanocortin-4 receptor mutations are a frequent and heterogeneous cause of morbid obesity. J Clin Invest. 2000;106:253–262. doi: 10.1172/JCI9238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Gaal LF, Rissanen AM, Scheen AJ, Ziegler O, Rössner S. Effects of the cannabinoid-1 receptor blocker rimonabant on weight reduction and cardiovascular risk factors in overweight patients: 1-year experience from the RIO-Europe study. Lancet. 2005;365:1389–1397. doi: 10.1016/S0140-6736(05)66374-X. [DOI] [PubMed] [Google Scholar]
- VIVUS. 2012. VIVUS provides update on timing of European decision for qnexa. [press release], 5 June 2012, Available at: http://ir.vivus.com/releasedetail.cfm?ReleaseID=679994 (accessed 15 August 2012)
- Wang C, Bomberg E, Billington C, Levine A, Kotz CM. Brain-derived neurotrophic factor in the hypothalamic paraventricular nucleus increases energy expenditure by elevating metabolic rate. Am J Physiol Regul Integr Comp Physiol. 2007;293:R992–R1002. doi: 10.1152/ajpregu.00516.2006. [DOI] [PubMed] [Google Scholar]
- Wang C, Godar RJ, Billington CJ, Kotz CM. Chronic administration of brain-derived neurotrophic factor in the hypothalamic paraventricular nucleus reverses obesity induced by high-fat diet. Am J Physiol Regul Integr Comp Physiol. 2010;298:R1320–R1332. doi: 10.1152/ajpregu.00844.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Westhuizen ET, Halls ML, Samuel CS, Bathgate RAD, Unemori EN, Sutton SW, et al. Relaxin family peptide receptors – from orphans to therapeutic targets. Drug Discov Today. 2008;13:640–651. doi: 10.1016/j.drudis.2008.04.002. [DOI] [PubMed] [Google Scholar]
- Woods SC, Figlewicz DP, Madden L, Porte D, Jr, Sipols AJ, Seeley RJ. NPY and food intake: discrepancies in the model. Regul Pept. 1998;75-76:403–408. doi: 10.1016/s0167-0115(98)00095-0. [DOI] [PubMed] [Google Scholar]
- World Health Organization. 2011. Obesity and overweight. [online] Available at: http://www.who.int/mediacentre/factsheets/fs311/en/index.html (accessed 15 August 2012)
- Xu B, Goulding EH, Zang K, Cepoi D, Cone RD, Jones KR, et al. Brain-derived neurotrophic factor regulates energy balance downstream of melanocortin-4 receptor. Nat Neurosci. 2003;6:736–742. doi: 10.1038/nn1073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu AW, Kaelin CB, Morton GJ, Ogimoto K, Stanhope K, Graham J, et al. Effects of hypothalamic neurodegeneration on energy balance. PLoS Biol. 2005;3:e415. doi: 10.1371/journal.pbio.0030415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yaswen L, Diehl N, Brennan MB, Hochgeschwender U. Obesity in the mouse model of pro-opiomelanocortin deficiency responds to peripheral melanocortin. Nat Med. 1999;5:1066–1070. doi: 10.1038/12506. [DOI] [PubMed] [Google Scholar]
- Yeo GS, Farooqi IS, Aminian S, Halsall DJ, Stanhope RG, O'Rahilly S. A frameshift mutation in MC4R associated with dominantly inherited human obesity. Nat Genet. 1998;20:111–112. doi: 10.1038/2404. [DOI] [PubMed] [Google Scholar]
- Yeo GSH, Hung CCC, Rochford J, Keogh J, Gray J, Sivaramakrishnan S, et al. A de novo mutation affecting human TrkB associated with severe obesity and developmental delay. Nat Neurosci. 2004;7:1187–1189. doi: 10.1038/nn1336. [DOI] [PubMed] [Google Scholar]
- Yukioka H, Kawanishi Y, Katsuura G, Takenaka H, Hara S. A potent and selective neuropeptide Y Y5 receptor antagonist, S-2367, attenuates the development of diet-induced obesity in mice. 2006. NAASO 2006 Annual Scientific Meeting.
- Yulyaningsih E, Zhang L, Herzog H, Sainsbury A. NPY receptors as potential targets for anti-obesity drug development. Br J Pharmacol. 2011;163:1170–1202. doi: 10.1111/j.1476-5381.2011.01363.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zarjevski N, Cusin I, Vettor R, Rohner-Jeanrenaud F, Jeanrenaud B. Chronic intracerebroventricular neuropeptide-Y administration to normal rats mimics hormonal and metabolic changes of obesity. Endocrinology. 1993;133:1753–1758. doi: 10.1210/endo.133.4.8404618. [DOI] [PubMed] [Google Scholar]