

Abstract
The aim of this study is to explore the inhibitory effects of RNA interference (RNAi) targeting NET-1 or combined with sorafenib on HCC in vitro and in vivo and the possible underlying mechanisms. The expressions of NET-1 mRNA and protein were detected by RT-QPCR and western blot. The ability of proliferation was determined by CCK-8 assay. Apoptosis was examined by flow cytometry (FCM). Abilities of migration and invasion were measured by scratch-wound assay and transwell assay. MHCC97H cells with stable transfection of NET-1shRNA were injected subcutaneously to prepare nude mice model of HCC and Caspase-3, Caspase-8, and Caspase-9 mRNAs of tumor tissues in different groups were examined. NET-1 mRNA and protein were reduced sharply in MHCC97H cells transfected with NET-1shRNA. The abilities of proliferation and migration were inhibited and apoptosis was promoted in either NET-1shRNA or sorafenib as compared with untreated cells in vitro and in vivo (P < 0.05). The mRNA levels of caspase-3, caspase-8, and caspase-9 of tumor tissues were reduced in different treatment groups compared with untreated group, particularly in combination group. (P < 0.05). The combination NET-1shRNA with sorafenib dramatically enhanced the effects of sorafenib antitumor ,which may involve in blocking ras signaling pathway and stimulating apoptotic pathways simultaneously.
1. Introduction
Hepatocellular carcinoma (HCC) is the third leading cause of cancer deaths worldwide [1], with some certain geographic regions in developing countries where the incidence of HCC is 16–32 times higher than in developed countries. Gene therapy, a new and promising therapeutic strategy, has been used for many cancers including HCC. SiRNA-targeted silencing of the genes associated with tumor cell proliferation or metastasis, as one method of gene therapy, shows great potency on HCC treatment. New EST tetraspanin-1, also called NET-1(C4.8, Tspan-1, P503S), a member of the tetraspan superfamily (TM4SF) [2–4], seems to be rather expressed in most HCC than in normal adult liver tissues [5]. This attractive characteristic of tumor-specific expression could made NET-1 as potential therapeutic target for HCC.
Sorafenib, an oral multikinase inhibitor approved by the US Food and Drug Administration for the treatment of patients with advanced renal cell carcinoma (RCC) and those with refractory HCC. Recent years, in vivo and in vitro studies have shown that sorafenib could inhibit tumor growth and disrupts tumor microvasculature through antiproliferative, antiangiogenic, and/or proapoptotic effects. Sorafenib represents an important advance in the treatment of advanced HCC and is the first systemic therapy shown to prolong survival in advanced HCC. A number of trials examining the combined use of sorafenib plus chemotherapy agents (e.g., fluorouracil [6], gemcitabine [7], or capecitabine plus oxaliplatin [8]) or of sorafenib plus other molecularly targeted therapies (e.g., sirolimus [9]) are currently underway and are yielding promising results. However, studies about gene therapy using RNAi technology combination with sorafenib on HCC rarely have been reported. Therefore in the present study, we used NET-1shRNA combined with sorafenib to explore a novel strategy for treating HCC in vivo and in vitro.
2. Methods
2.1. Cell Culture
Human HCC cell line MHCC97H was kindly provided by Liver Cancer Institute, Zhongshan Hospital, Shanghai. Cells were cultured with Dulbecco's modified Eagle's medium (DMEM) (Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum (FBS) (Invitrogen, Carlsbad, CA) in humidified atmosphere containing 5% CO2 at 37°C. The present experiments were divided into four groups including untreated group, sorafenib group, NET-1shRNA group and the combination NET-1shRNA with sorafenib group.
2.2. Transfection of Plasmids
pSilencer4.1-CMVneo-NET-1shRNA (NET-1shRNA) and pSilencer4.1-CMVneo-control shRNA (control shRNA) plasmids were designed and synthesized by Biomics Biotechnologies Co., Ltd. (Nantong, Jiangsu, China). One day prior transfection, cells were cultured in medium without serum and antibiotics. After mixed gently and incubated for 20 minutes at room temperature, the transfection mixture of shRNA and Lipofectamine 2000 (Invitrogen) was added into culture plates. After 6 h, the transfection mixture was replaced by DMEM supplemented with 10% FBS. Cells were harvested at 48 h after transfection.
2.3. Real Time RT-QPCR
Total RNA was isolated with PicoPure RNA isolation Kit (Arcturus Bioscience, Mountain View, CA, USA) according to the manufacture's instructions. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was amplified as an internal control [10]. The PCR primers were designed by Premier Primer 5.0 software (Table 1). Primers (20 pmol/μL) were added into reaction buffer with a total volume of 25 μL together with template RNA 4 μL, Master Mix 12.5 μL, and SYBR Green I 0.5 μL. PCR was performed at the following conditions: 30 seconds at 94°C, 30 seconds at 62°C, and 40 seconds at 72°C for 50°C cycles. Real time RT-QPCR with SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA) was performed using a MiQ machine (Bio-Rad Laboratories, Hercules, CA). The fluorescent signals were collected during the extension phase, Ct values of the sample were calculated, and NET-1 transcript levels were analyzed by 2-ΔΔCt method.
Table 1.
Primer sequences of NET-1, caspase 3, caspase 8, caspase 9, and GAPDH.
| Primer | Sequence | Product (bp) |
|---|---|---|
| NET-1-F′ | 5′-GTGGCTTCACCAACTATACG-3′ | 191 |
| NET-1-R′ | 5′-GACTGCATTAGTTCGGATGT-3′ | |
| Caspase-3-F′ | 5′-AGAACTGGACTGTGGCATTGAG-3′ | 191 |
| Caspase-3-R′ | 5′-GCTTGTCGGCATACTGTTTCAG-3′ | |
| Caspase-8-F′ | 5′-CATCCAGTCACTTTGCCAGA-3′ | 128 |
| Caspase-8-R′ | 5′-GCATCTGTTTCCCCATGTTT-3′ | |
| Caspase-9-F′ | 5′-TTCCCAGGTTTTGTTTCCTG-3′ | 143 |
| Caspase-9-R′ | 5′-CCTTTCACCGAAACAGCATT-3′ | |
| GAPDH-F′ | 5′-TGATGACATCAAGAAGGTGGTGAAG-3′ | 240 |
| GAPDH-R′ | 5′-TCCTTGGAGGCCATGTGGGCCAT-3′ |
2.4. Western Blot Analysis
MHCC97H cells were harvested at 48 h after transfection. Cells were lysed with buffer containing 0.1 mol/L Tris-HCl (pH6.8), 4% SDS, 20% glycerine, 0.1% BPB, and 5% β-mercaptoethanol. The complex was heated in boiling water for 5 minutes. Proteins were separated by 10% polyacrylamide gel electrophoresis and then transferred onto polyvinylidene fluoride (PVDF) membranes (Millipore, USA) at 350 mA for 2 h, which was later soaked for 2 h on a blocking solution (Tris-buffered saline containing 5% nonfat dry milk and 0.01% vol/vol Tween-20), and incubated for one hour at room temperature in the presence of Anti-NET-1 rabbit polyclonal antibody [11, 12], or anti-GAPDH mouse monoclonal antibody (Sigma, USA) used as internal control, then incubated at 4°C overnight. After incubation, the membrane was washed 3 times, and peroxidase-conjugated goat anti-rabbit or goat anti-mouse secondary antibodies (ICN Laboratories, Irvine, CA; diluted 1 : 10,000) were added and incubated for an additional one hour. Reaction was visualized by the ECL chemiluminescence detection system (Pierce, USA) on radiographic films (Koda, USA). The molecular weight of NET-1 and GAPDH [13] were 76 kDa and 37 kDa, respectively. The results were analyzed using Image J software.
2.5. Cell Counting Kit-8 (CCK-8) Assays
2.5.1. IC50 Determination
Sorafenib was purchased from Bayer Pharmaceutical Corporation, dissolved in sterile DMSO, and stored frozen under light-protected conditions at −20°C. Stock solutions of sorafenib were diluted in DMEM culture medium, and equal aliquots were added to individual wells so that the final concentrations were 0, 3, 9, 18, 30, 60, and 100 μM. To determine the 50% inhibitory concentration (IC50) for sorafenib, the CCK-8 (Dojindo, Kumamoto, Japan) was used. MHCC97H cells were seeded at 5.0 × 103 cells/well in 100 μL of DMEM in 96-well microplates and incubated overnight at 37°C in a humidified atmosphere with 5% CO2. After the cultures were incubated for 0 h, 24 h, 48 h, and 72 h, we exchanged fresh DMEM gently and 10 μL of CCK-8 solution was added to each well, and the plates were incubated for 2 h at 37°C. We measured the absorbance at 450 nm using an optical density (Microplate Reader 550; Bio-Rad, Tokyo, Japan) and calculated the IC50 concentration of sorafenib by the intersection of the plotted line (Figure 2). The IC50 of sorafenib was used in the growth, apoptosis, scratch-wound, and transwell assays.
Figure 2.
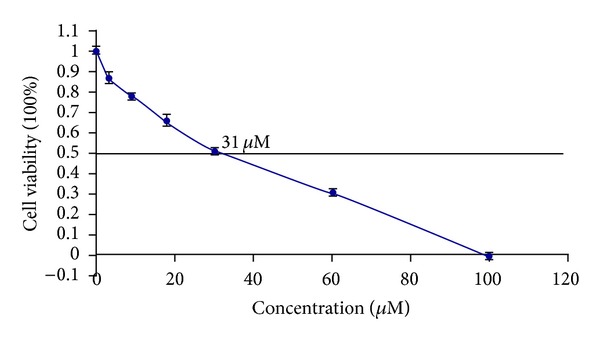
Dose-related cytotoxicity following 72 h exposured to sorafenib. The IC50 of sorafenib was indicated by the intersection of the plotted line. Sorafenib was 31 μM.
2.5.2. Growth Assays
5 × 103 cells in each group were seeded in 96-well plates and cultured for 0, 24, 48, and 72 h, the optical density at 450 nm wavelength was measured through an automated plate reader and then cell growth curves were drawn.
2.6. Flow Cytometry (FCM)
Cellular apoptosis was determined using the Annexin V-FITC Apoptosis Detection Kit I (Clontech Laboratories Inc., USA) according to the manufacturer's protocol. Cells were seeded in 6-well plates at the density of 2 × 105/mL and harvested by trypsinization then washed with cold PBS, centrifuged at 1000 rpm, resuspended in 400 μL 1 × binding buffer, centrifuged again and removed supernatant. Cells were resuspended in 200 μL 1 × binding buffer and transferred to a sterile FCM glass tube. 3 μL Annexin V-FITC and 3 μL propidium iodide were added and then incubated in the dark at room temperature. Cells were analyzed by FCM (FACSCalibur, Becton-Dickinson, USA) at 488 nm. The distribution of cells was analyzed using CellQuest software (Becton-Dickinson) in the FCM within 1 hour of staining. Data from 10,000 cells was collected for each data file. Apoptotic cells were identified as Annexin V-FITC-positive and P-negative cells.
2.7. Scratch-Wound Assay
Cells were seeded at a density of 5 × 105 in 24 well plates and incubated overnight. The day after, the surfaces of the dishes were mildly scratched with a yellow P200 pipette tip (Fisher) and images were taken under a Carl Zeiss Axiovert 25 (Thornwood New York, USA) inverted microscope with the use of a Cannon PowerShot G9 digital camera using a 40X objective, which were totally observed continuously for 72 h.
2.8. Transwell Assay
Uncoated or Matrigel-coated transwells containing 8 μm pores were used for the assays (Costar, Corning, NY). Cells were seeded into the upper chamber in serum-free DMEM media. DMEM media containing 10% FBS was added to the lower chamber. Cells were fixed in 100% methanol 20 h later and stained with 0.2% concentration of crystal violet for 15 min at room temperature. Cells remaining on the upper side of the filter were removed with a cotton swab. The filters were then mounted onto cover slips and images were taken at 40X magnification. From these images, the number of migratory or invasive cells was counted.
2.9. G418 Selection
G418, an aminoglycoside antibiotic, is the most commonly antistable transfection reagents for screening in molecular genetic testing. Cells were seeded at the density of 2 × 105 cells/well in 24-well plates and grew overnight. The medium was replaced with complete medium without FBS. NET-1shRNA and control shRNA were transfected into MHCC97H cells using Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. The medium was replaced with a fresh medium of bovine serum (150 mL/L) after 6 h transfection. One day later, the transfected cells were selected by G418 (400 μg/mL) (Huamei Biotechnology Company, Beijing, China) until positive clones were discovered after 14 days. The cells were cultured and finally selected by G418 (200 μg/mL) for a further 14 days. Single clones were selected to build a stable transfected cell line.
2.10. Animals and Establishment of Tumor Model
2.10.1. Animals
Nude BALB/c mice, female, 4–6 weeks old and weighing 18∼20 g, were obtained from Shanghai Slac Laboratory Animal (China) and housed under pathogen-free conditions according to the recommendations of NIH guidelines for care and use of laboratory. This study has been approved by the Animal Ethical Committee of Nantong University.
2.10.2. Tumor Model
Nude mice were inoculated subcutaneously at the right anterior axilla with 1 × 107 stably transfected NET-1 MHCC97H cells in 200 μL PBS and at left anterior axilla with equal untreated cells, as autocontrol. The shortest axis (a) and the longest axis (b) of tumor were measured by caliper every day. The tumor volumes were calculated with the formula: volumes = a 2 × b × π/6. When the tumor volume reached 100 mm3 at least, the tumor-bearing nude mice model was established successfully.
The experiments composed of 12 nude mice, which were randomly assigned to four experimental groups. The four experimental groups were as follows: untreated MHCC97H cells group, stably transfected NET-1shRNA MHCC97H cells group, sorafenib treating MHCC97H cells group, and sorafenib treating stably transfected NET-1shRNA MHCC97H cells group. In sorafenib treatment group, the mice were given 100 mg/kg sorafenib in 100 μL by peritoneal injection after tumor implantation established. All control mice received an equal volume of carrier solution by peritoneal injection.
At the 3 weeks of treatments, all treated mice were sacrificed, livers were excised and weighed, and one part was used for pathological examination and the others were stored at −80°C for detecting Caspase-3, Caspase-8, and Caspase-9 mRNA by real time RT-QPCR; the primer sequences were in Table 1.
2.11. Statistical Analysis
All experiments were performed in triplicate and the results were expressed as mean ± standard errors. All data were analyzed with SPSS13.0 statistical software using student's t-test. Two-tailed P values of <0.05 were considered statistically significant.
3. Results
3.1. NET-1 Expression in MHCC97H Cells
RT-QPCR and western blot analysis was performed to determine whether transfection with NET-1shRNA resulted in a reduction of the expressions of NET-1 mRNA and protein. As compared with control shRNA, there were 49% and 51% reduction of NET-1 mRNA and protein levels in cells transfected with NET-1shRNA, and no significant reduction of NET-1 expression was found in cells transfected with either control shRNA or untreated group (Figure 1).
Figure 1.
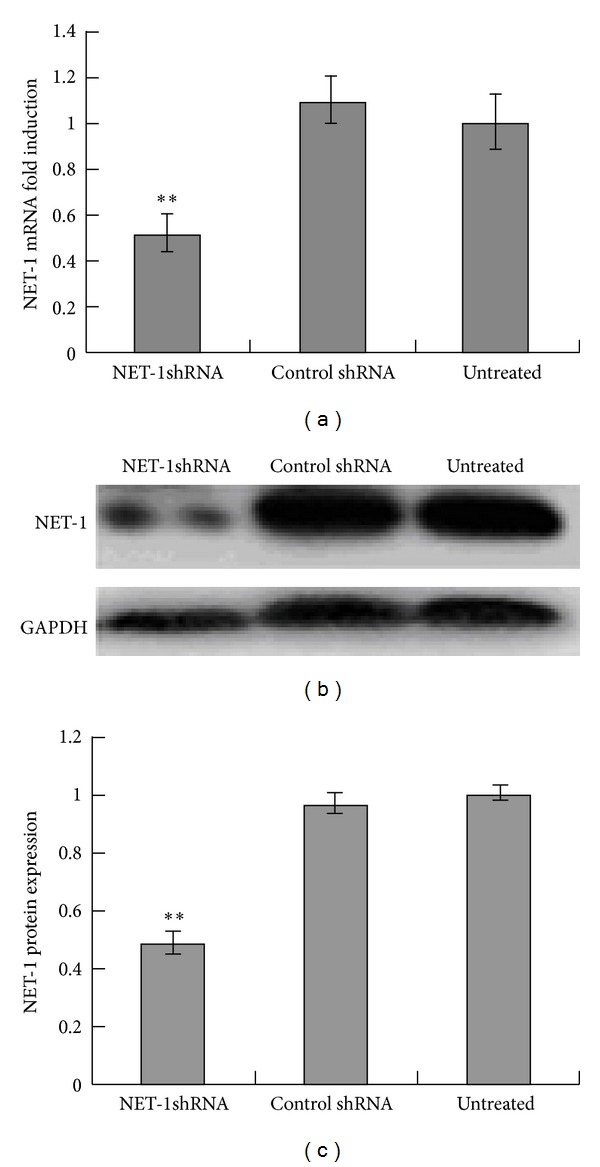
Inhibition of NET-1 mRNA and protein expression by NET-1shRNA in HCC cells line MHCC97H cells. (a) RT-QPCR showed NET-1shRNA resulted in 49% reduction of NET-1 mRNA levels, when compared with untreated group. (b) Western blot showed the intensities of NET-1 protein and GAPDH protein in these three groups. (c) Image J analyzed NET-1shRNA resulted in 51% reduction of NET-1 protein levels, when compared with untreated group (**P < 0.01).
To evaluate the proliferation of MHCC97H cells treated by NET-1shRNA, sorafenib, and combination of them, cells growth curves were obtained through CCK-8 assay. There was a significant reduction of proliferation in all treated cells, and especially in combination NET-1shRNA with sorafenib as compared with untreated cells (each P < 0.01). However, there was no significant difference in cell proliferation rates between sorafenib and NET-1shRNA groups (Figure 3).
Figure 3.
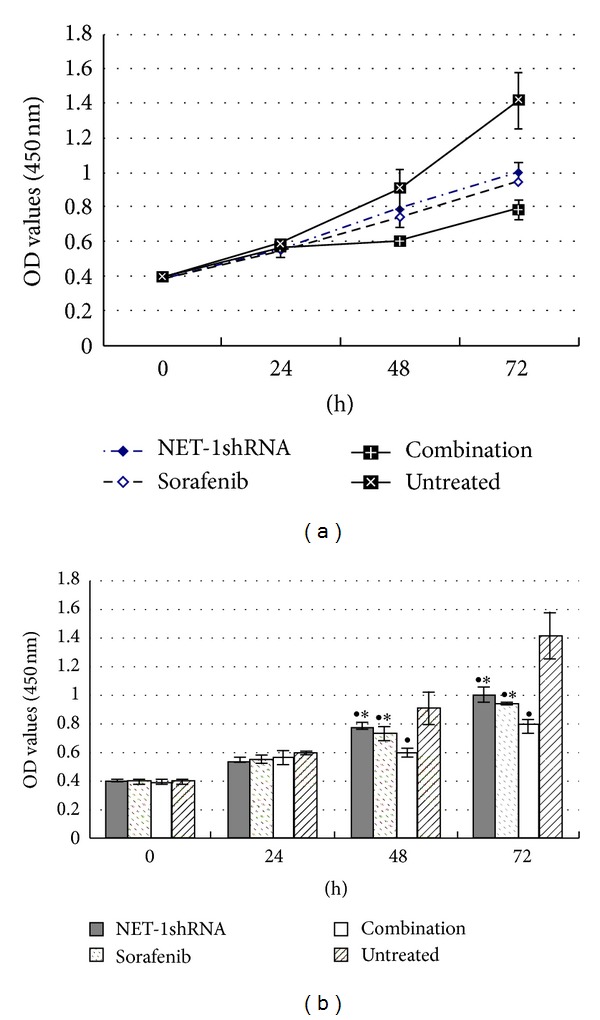
Growth curves showed the proliferation of MHCC97H cells until 72 h by CCK-8 assay. NET-1shRNA, sorafenib, and combination of NET-1shRNA with sorafenib resulted in 14.6%, 19.3%, and 33.3% reduction of cell proliferation at 48 h, in 46.5%, 50.6%, and 66.3% reduction of cell proliferation at 72 h. (● P < 0.05 compared with untreated group. *P < 0.05 compared with combination group).
3.2. Proliferation of MHCC97H Cells
MHCC97H cells in 96-well culture plates were exposed to different concentrations of sorafenib for 48 h. IC50 of sorafenib was calculated as 31 μM (Figure 2).
3.3. Apoptosis of MHCC97H Cells
FCM was used to examine the apoptosis of MHCC97H cells. The apoptosis rate rose up in all three treated groups compared with untreated group. Interestingly, combination NET-1shRNA with sorafenib notably advanced the apoptosis rate than either sorafenib or NET-1shRNA group (P < 0.05, resp.). But there was no difference between these two single groups (Figure 4).
Figure 4.
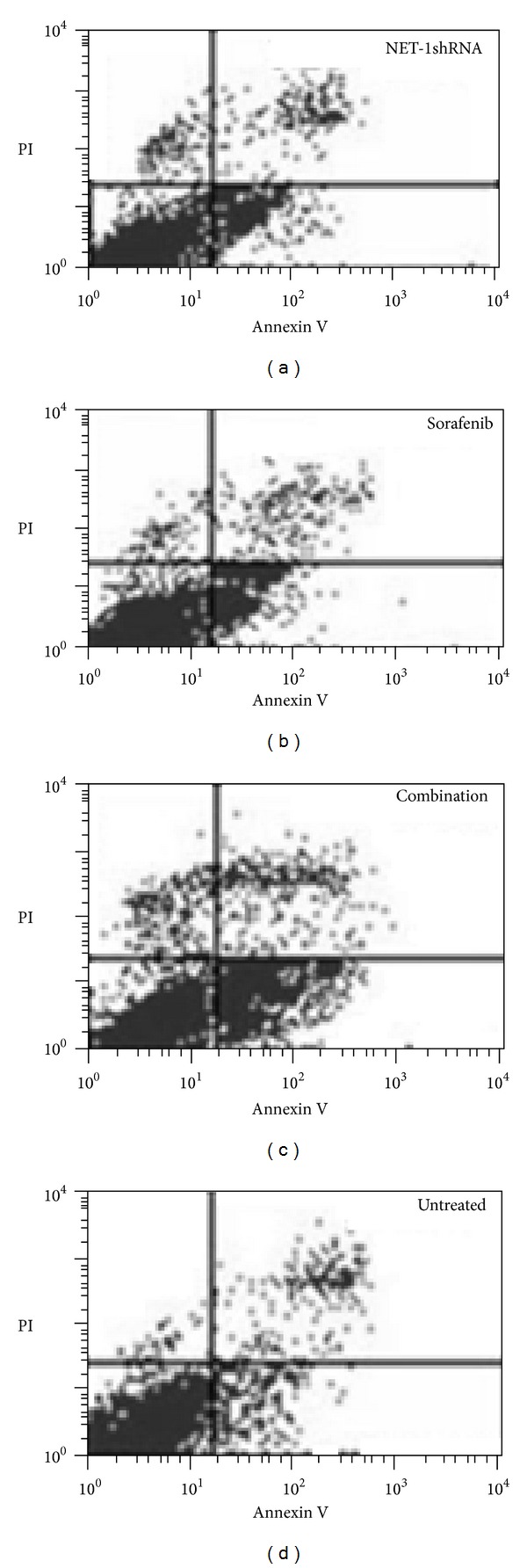
Percentage of apoptotic cells included both early- and late-stage apoptosis (AV+/PI− and AV+/PI+) was detected by FCM. The apoptosis rates of NET-1shRNA, sorafenib, combination, and untreated group were 24 ± 0.72%, 17.64 ± 0.46%, 51.34 ± 1.25%, and 6.3 ± 0.03%, respectively.
3.4. Motility of MHCC97H Cells
Scratch-wound and trans-well chamber assays were conducted to evaluate the migration of MHCC97H cells in the presence of sorafenib, NET-1shRNA, and both of them. Under these conditions, Scratch-wound (Figure 5) and trans-well chamber (Figure 6) showed MHCC97H cells display different migrating potential compared to untreated cells, respectively. Interestingly, there was no disparity between NET-1shRNA group and sorafenib group. Cells were observed under an inverted microscope.
Figure 5.
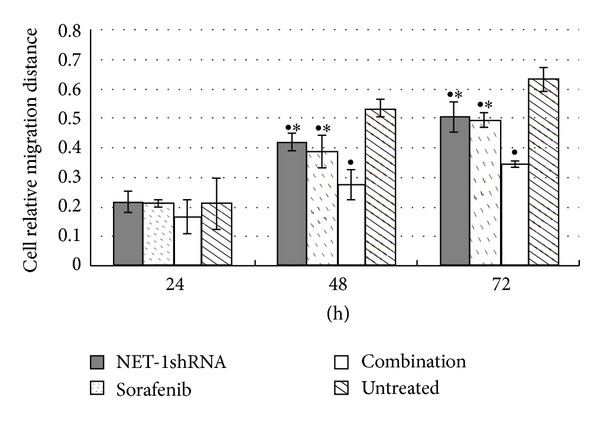
Cell relative migration distance at different time points in different groups. ● P < 0.05 compared with untreated group. *P < 0.05 compared with combination group.
Figure 6.
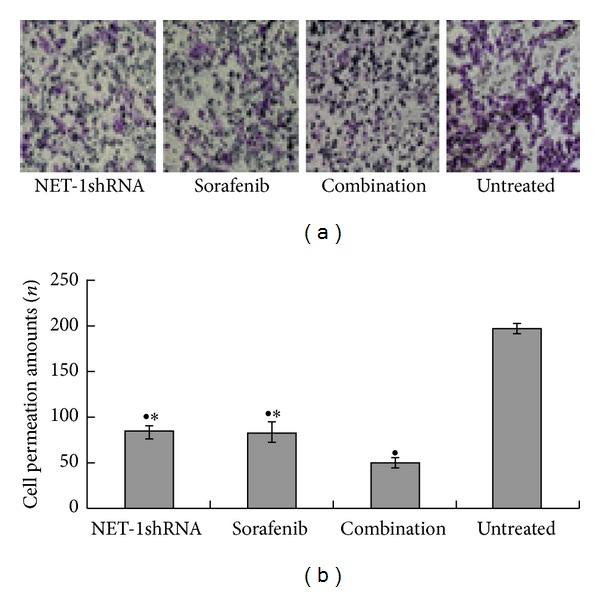
Motility of MHCC97H cells was inhibited by NET-1shRNA and sorafenib. (a) Trans-well chamber showed the permeation amounts of MHCC97H cells after 48 h of NET-1shRNA transfection or sorafenib treatment. (b) Trans-well chamber assay suggested the permeation amounts of MHCC97H cells in NET-1shRNA, sorafenib, and combination NET-1shRNA with sorafenib and untreated cells groups were 83.67 ± 6.03, 83.33 ± 10.01, 50.33 ± 4.73, and 197.00 ± 6.00, respectively. (● represents P < 0.05 compared with untreated group. * represents P < 0.05 compared with combination group).
3.5. Inhibition of HCC Model Growth by NET-1shRNA Alone or in Combination with Sorafenib
The tumor was used to mimic liver tumor growth in mice. The average tumor size presented statistical differences in these four groups. There was smaller tumor size in all treated groups than that in untreated group (P < 0.05, resp.). Notably, combination group in the inhibiting effect was superior to each single group (P < 0.05, resp.). However, no statistical difference was found between NET-1shRNA group and sorafenib group (Table 2).
Table 2.
Comparing HCC growth treated by NET-1shRNA alone and in combination with sorafenib.
| Groups | MHCC97H (cm3) |
|---|---|
| NET-1shRNA | 1.354 ± 0.042●★ |
| Sorafenib | 1.201 ± 0.152●★ |
| Combination | 0.558 ± 0.018● |
| Untreated | 3.459 ± 0.121 |
| Empty vector | 3.279 ± 0.084 |
The average tumor size in each group was 1.354 ± 0.042 cm3, 1.201 ± 0.152 cm3, 0.558 ± 0.018 cm3, and 3.459 ± 0.121 cm3, respectively, (● P < 0.05 compared with untreated group. ★ P < 0.05 compared with combination group).
To further investigate the mechanism about inhibiting tumor growth, we detected the mRNA levels of caspase-3, caspase-8, and caspase-9 of the tumor tissues from these four groups. The results of real time RT-QPCR showed caspase 3 in NET-1shRNA, sorafenib, and combination groups significantly increased, respectively, when compared with untreated group (each P < 0.05). Because Caspase 8 mRNA levels significantly increased in NET-1shRNA and combination groups, Caspase 9 mRNA levels significantly increased in sorafenib and combination groups, respectively, when compared with untreated group (each P < 0.05), so that increasing expressions of caspase-3, Caspase-8, and Caspase-9 mRNA in combination group were higher than that in each single group (P < 0.05, resp.) (Figure 7).
Figure 7.
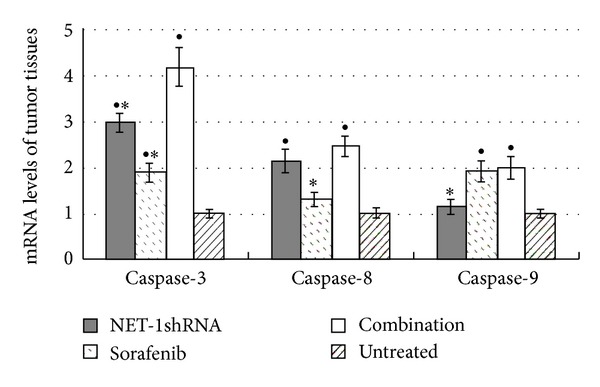
The expression mRNA levels of caspase 3, caspase 8, and caspase 9 of tumor tissues in NET-1shRNA, sorafenib, combination, and untreated groups. RT-QPCR showed that in NET-1shRNA, sorafenib, and combination groups caspase 3 mRNA levels increased by 2.99, 1.89, and 4.18 times, Caspase 8 mRNA levels increased by 2.14, 1.31 and 2.47 times, and Caspase 9 mRNA levels increased by 1.15, 1.91, and 1.99 times, compared with untreated group, respectively. (● P < 0.05 compared with untreated group, *P < 0.05 compared with combination group).
4. Discussion
NET-1 as a novel tumor relative gene is over-expressed in many malignant tumors including the breast, uterine cervix, colon, esophagus, liver, lung, ovary, pancreas, prostate, gastrointestinal, and skin [10, 14–28]. Recent studies have shown that NET-1 is involved in a variety of processes such as oncogenesis, cell cycle control, apoptosis, and migration. Wollscheid et al. [17] found that NET-1 gene expression correlated to cell proliferation and may be used as a marker for cervical cancer prognosis.
RNAi technology is a powerful approach to silence mammalian gene expression for studies of gene function and has the potential for gene therapy. Synthetic siRNA can trigger RNAi response in mammalian cells and induce specific inhibition of gene expression. FDA approval has been granted for an investigational new drug license to test the use of expressed RNA sequences against HBV [29], bringing a promise for RNAi treatment on HCC. The specific shRNA used in our experiments has been exploited by Chen et al. and confirmed effective in previous research [10]. In the present study, NET-1 mRNA and protein expressions were significantly reduced by 49% and 51%, respectively, using NET-1shRNA.
NET-1 was proved to be associated with neoplastic cell proliferation [20]. Our data indicated an inhibition of tumor cell proliferation by 46.5% after single-agent sorafenib treatment, and 50.6% after NET-1shRNA transfection, respectively. Combined treatment with sorafenib plus NET-1shRNA significantly enhanced the antiproliferative effect of sorafenib on HCC in vitro. Furthermore, combined treatment also inhibited tumor volume of the tumor-bearing nude mice model more assertively than treated with a single agent in vivo. NET-1 belongs to the member of the guanine nucleotide exchange factor (GEF) family, the latter helps small G protein mutual transition from GDP to GTP, thus activating Ras and Rho. So our findings implied that NET-1 may promote tumor proliferation and growth through activating Ras signal transduction pathway, which mediates tumor cell proliferation, differentiation, and survival. The antitumor effect of sorafenib plus downregulation NET-1 by RNAi may achieve a synergistic effect on the inhibition of cell growth by inhibiting several relevant pathways.
NET-1 has 30% identity with metastasis-associated tetraspans (e.g., Co-029 and Talla-1) [2]. Tetraspanins were implicated in regulation of signaling, motility, migration, and invasiveness [30]. Huang et al. showed a MK-induced NET-1 pathway contributing to migration/invasiveness of human head and neck squamous cell carcinoma cells then found RNAi silencing of NET-1 dramatically decreased MK-induced expression of MMP-2, which demonstrated the role of NET-1(TSPAN1) as one of various signaling components [31]. Our results revealed that migration and infiltration of MHCC97H cell were dramatically reduced after transfection of NET-1shRNA. Combination therapy with sorafenib and NET-1shRNA amplified the inhibition effects of sorafenib on tumor migration/invasiveness from 57.71% to 74.46%, compared with the untreated. Meanwhile, Sorafenib has been classified as a vascular endothelial growth factor receptor (VEGFR) inhibitor, which blocks tumor angiogenesis by decreasing microvessel density and circulating levels of VEGF. VEGF is one of the most potent angiogenic factors. MMP2 is a marker associated with angiogenesis as well as metastatic invasion [32]. Several studies demonstrated that the intensity of angiogenesis in HCC correlated with the risk of vascular invasion, metastasis, and patient prognosis [33, 34]. Therefore, these two agents (sorafenib and NET-1shRNA) in combination may boost the effect of each single agent.
Sorafenib has recently been found to induce apoptosis in several human cancer lines. Although the mechanism through which sorafenib induces apoptosis has not been fully elucidated, Yu et al. found sorafenib induced apoptosis by downregulating myeloid cell leukemia-1(Mcl-1) [35], which was presumed to be associated with the release of cytochrome c from mitochondria into the cytosol, caspase activation, and apoptotic cell death. In vitro apoptosis assay proved this hypothesis: the apoptosis rate of sorafenib treated group was higher than untreated cells. Moreover, our results revealed that caspase 3 and caspase 9 mRNA levels changed significantly after sorafenib treatment. Interestingly, from the results, we could find that caspase 3 mRNA level increased dramatically and the caspase 8 mRNA increased in NET-1shRNA and combination groups. The increase of caspase 8 mRNA led by NET-1shRNA perhaps hinting that NET-1 was probably an apoptosis related gene associated with the death receptor pathway mediated by membrane receptor. In addition, in vitro results demonstrated that reduction of NET-1 by RNAi may also induce apoptosis. Under our deduction, it is easy to explain why caspase 3 mRNA levels increased greatly in combination group. As for the relationship between NET-1 and caspase 8, we still need to explore the mechanism of NET-1shRNA induced apoptosis through other effective methods.
Taken together, in this study NET-1shRNA has obviously suppressed proliferation, survival, and migration/invasiveness of HCC both in vitro and in vivo. NET-1shRNA manifested inhibitory effect on HCC as well as sorafenib. Furthermore, combination of sorafenib plus NET-1shRNA showed better antitumor effect than single-agent treatment. These findings would bring a new therapy for HCC.
Acknowledgments
This research was supported by funding from the Priority Academic Program Development of Jiangsu Higher Education Institutions, the Foundation of the Ministry of Health, Jiangsu province, China (no. H201052); the Science Foundation of Nantong City, Jiangsu province, China (nos. K2009060 and S2010018); and the Advanced Project Program of Nantong University. The authors are grateful to Biomics Biotechnologies Co., Ltd. (Nantong, Jiangsu, China) for kindly helping us with the synthesis of NET-1shRNA and technical guidance.
References
- 1.WHO. The Global Burden of Disease: 2004 Update. Geneva, Switzerland: World Health Organization; 2008. http://www.who.int/healthinfo/global_burden_disease/2004_report_update/en/index.html. [Google Scholar]
- 2.Serru V, Dessen P, Boucheix C, Rubinstein E. Sequence and expression of seven new tetraspans. Biochimica et Biophysica Acta. 2000;1478(1):159–163. doi: 10.1016/s0167-4838(00)00022-4. [DOI] [PubMed] [Google Scholar]
- 3.Claas C, Seiter S, Claas A, Savelyeva L, Schwab M, Zöller M. Association between the rat homologue of CO-029, a metastasis-associated tetraspanin molecule and consumption coagulopathy. Journal of Cell Biology. 1998;141(1):267–280. doi: 10.1083/jcb.141.1.267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Xu J, Stolk JA, Zhang X, et al. Identification of differentially expressed genes in human prostate cancer using subtraction and microarray. Cancer Research. 2000;60(6):1677–1682. [PubMed] [Google Scholar]
- 5.Li C, Dechun L, Yuanyuan Z. Expression and significance of NET-1 gene in liver cancer. Chinese Journal of Clinical and Experimental Pathology. 2003;19(5):488–491. [Google Scholar]
- 6.Italian Trial in Medical Oncology. Phase II study of sorafenib and infusional 5-fluorouracil in advanced hepatocellular carcinoma (HCC) [ClinicalTrials.gov identifier NCT00619541] US National Institutes of Health, 2008, http://www.clinicaltrials.gov/
- 7.Mahidol University. Efficacy and safety of sorafenib (Nexavar) in combination with gemcitabine in advanced hepatocellular carcinoma(HCC) [ClinicalTrials.gov identifier NCT00703365] US National Institutes of Health, 2008, http://www.clinicaltrials.gov/
- 8.The University of Hong Kong. Sorafenib with capecitabine and oxaliplatin for advanced or metastatic hepatocellular carcinoma (SECOX) [ClinicalTrials.gov identifier NCT00752063] US National Institutes of Health, 2008, http://www.clinicaltrials.gov/
- 9.Radboud University. Phase I study with sorafenib and sirolimus [ClinicalTrials.gov identifier NCT00509613] US National Institutes of Health, 2008, http://www.clinicaltrials.gov/
- 10.Chen L, Wang JL, Li H, Qin J, Wu Y. Functions of NET-1 gene in skin squamous cell carcinoma cell line (A431): a siRNA study. Zhonghua Bing Li Za Zhi. 2009;38(10):691–696. [PubMed] [Google Scholar]
- 11.Chen L, Li X, Wang G, Wang Y, Zhu Y, Zhu J. Clinicopathological significance of overexpression of TSPAN1, KI67 and CD34 in gastric carcinoma. Tumori. 2008;94(4):531–538. doi: 10.1177/030089160809400415. [DOI] [PubMed] [Google Scholar]
- 12.Wang GL, Chen L, Wei YZ, et al. The effect of NET-1 on the proliferation, migration and endocytosis of the SMMC-7721 HCC cell line. Oncology Reports. 2012;27(6):1944–1952. doi: 10.3892/or.2012.1698. [DOI] [PubMed] [Google Scholar]
- 13.Lott P, Amiott EA, Soto J, et al. Mitochondrial fusion and function in Charcot-Marie-Tooth type 2A patient fibroblasts with mitofusin 2 mutations. Experimental Neurology. 2008;211(1):115–127. doi: 10.1016/j.expneurol.2008.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Abba MC, Hu Y, Sun H, et al. Gene expression signature of estrogen receptor α status in breast cancer. BMC Genomics. 2005;6, article 37 doi: 10.1186/1471-2164-6-37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Nishidate T, Katagiri T, Lin M, et al. Genome-wide gene-expression profiles of breast-cancer cells purified with laser microbeam microdissection: identification of genes associated with progression and metastasis. International Journal of Oncology. 2004;25(4):797–819. [PubMed] [Google Scholar]
- 16.Chen L, Zhu YY, Zhang X, et al. TSPAN1 protein expression: a significant prognostic indicator for patients with colorectal adenocarcinoma. World Journal of Gastroenterology. 2009;15(18):2270–2276. doi: 10.3748/wjg.15.2270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wollscheid V, Kühne-Heid R, Stein I, et al. Identification of a new proliferation-associated protein NET-1/C4.8 characteristic for a subset of high-grade cervical intraepithelial neoplasia and cervical carcinomas. International Journal of Cancer. 2002;99(6):771–775. doi: 10.1002/ijc.10442. [DOI] [PubMed] [Google Scholar]
- 18.Brabender J, Marjoram P, Salonga D, et al. A multigene expression panel for the molecular diagnosis of Barrett’s esophagus and Barrett’s adenocarcinoma of the esophagus. Oncogene. 2004;23(27):4780–4788. doi: 10.1038/sj.onc.1207663. [DOI] [PubMed] [Google Scholar]
- 19.Chen L, Shen AG, Wang G, Lu P, Li X. Expression of NET-1 gene and protein in hepatocellular carcinoma and related tissues. Ai Zheng. 2006;25(3):320–325. [PubMed] [Google Scholar]
- 20.Chen L, Wang Z, Zhan X, Li DC, Zhu YY, Zhu J. Association of NET-1 gene expression with human hepatocellular carcinoma. International Journal of Surgical Pathology. 2007;15(4):346–353. doi: 10.1177/1066896907306083. [DOI] [PubMed] [Google Scholar]
- 21.Borczuk AC, Gorenstein L, Walter KL, Assaad AA, Wang L, Powell CA. Non-small-cell lung cancer molecular signatures recapitulate lung developmental pathways. The American Journal of Pathology. 2003;163(5):1949–1960. doi: 10.1016/S0002-9440(10)63553-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Euer NI, Kaul S, Deissler H, Möbus VJ, Zeillinger R, Weidle UH. Identification of L1CAM, Jagged2 and Neuromedin U as ovarian cancer-associated antigens. Oncology Reports. 2005;13(3):375–387. [PubMed] [Google Scholar]
- 23.Santin AD, Zhan F, Bellone S, et al. Gene expression profiles in primary ovarian serous papillary tumors and normal ovarian epithelium: identification of candidate molecular markers for ovarian cancer diagnosis and therapy. International Journal of Cancer. 2004;112(1):14–25. doi: 10.1002/ijc.20408. [DOI] [PubMed] [Google Scholar]
- 24.Buchholz M, Braun M, Heidenblut A, et al. Transcriptome analysis of microdissected pancreatic intraepithelial neoplastic lesions. Oncogene. 2005;24(44):6626–6636. doi: 10.1038/sj.onc.1208804. [DOI] [PubMed] [Google Scholar]
- 25.Iacobuzio-Donahue CA, Ashfaq R, Maitra A, et al. Highly expressed genes in pancreatic ductal adenocarcinomas: a comprehensive characterization and comparison of the transcription profiles obtained from three major technologies. Cancer Research. 2003;63(24):8614–8622. [PubMed] [Google Scholar]
- 26.Iacobuzio-Donahue CA, Maitra A, Olsen M, et al. Exploration of global gene expression patterns in pancreatic adenocarcinoma using cDNA microarrays. The American Journal of Pathology. 2003;162(4):1151–1162. doi: 10.1016/S0002-9440(10)63911-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ryu B, Jones J, Blades NJ, et al. Relationships and differentially expressed genes among pancreatic cancers examined by large-scale serial analysis of gene expression. Cancer Research. 2002;62(3):819–826. [PubMed] [Google Scholar]
- 28.Lee CH, Bang SH, Lee S, Song K, Lee I. Gene expression profiling reveals sequential changes in gastric tubular adenoma and carcinoma in situ. World Journal of Gastroenterology. 2005;11(13):1937–1945. doi: 10.3748/wjg.v11.i13.1937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Arbuthnot P, Thompson LJ. Harnessing the RNA interference pathway to advance treatment and prevention of hepatocellular carcinoma. World Journal of Gastroenterology. 2008;14(11):1670–1681. doi: 10.3748/wjg.14.1670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hemler ME. Tetraspanin proteins mediate cellular penetration, invasion, and fusion events and define a novel type of membrane microdomain. Annual Review of Cell and Developmental Biology. 2003;19:397–422. doi: 10.1146/annurev.cellbio.19.111301.153609. [DOI] [PubMed] [Google Scholar]
- 31.Huang Y, Sook-Kim M, Ratovitski E. Midkine promotes tetraspanin-integrin interaction and induces FAK-Stat1α pathway contributing to migration/invasiveness of human head and neck squamous cell carcinoma cells. Biochemical and Biophysical Research Communications. 2008;377(2):474–478. doi: 10.1016/j.bbrc.2008.09.138. [DOI] [PubMed] [Google Scholar]
- 32.Tutton MG, George ML, Eccles SA, Burton S, Swift RI, Abulafi AM. Use of plasma MMP-2 and MMP-9 levels as a surrogate for tumour expression in colorectal cancer patients. International Journal of Cancer. 2003;107(4):541–550. doi: 10.1002/ijc.11436. [DOI] [PubMed] [Google Scholar]
- 33.Poon RT, Ng IO, Lau C, et al. Tumor microvessel density as a predictor of recurrence after resection of hepatocellular carcinoma: a prospective study. Journal of Clinical Oncology. 2002;20(7):1775–1785. doi: 10.1200/JCO.2002.07.089. [DOI] [PubMed] [Google Scholar]
- 34.Yao DF, Wu XH, Zhu Y, et al. Quantitative analysis of vascular endothelial growth factor, microvascular density and their clinicopathologic features in human hepatocellular carcinoma. Hepatobiliary and Pancreatic Diseases International. 2005;4(2):220–226. [PubMed] [Google Scholar]
- 35.Yu C, Bruzek LM, Xue WM, et al. The role of Mcl-1 downregulation in the proapoptotic activity of the multikinase inhibitor BAY 43-9006. Oncogene. 2005;24(46):6861–6869. doi: 10.1038/sj.onc.1208841. [DOI] [PubMed] [Google Scholar]