

Abstract
Enzymes called SENPs catalyze both the maturation of small ubiquitin-like modifier (SUMO) precursors and removal of SUMO modifications, which regulate essential cellular functions such as cell cycle progression, DNA damage response and intracellular trafficking. Some members, such as SENP1, are potential targets for developing cancer therapeutics. We searched for small molecule inhibitors of SENPs using in-silico screening in conjunction with biochemical assays, and identified a new chemotype of small molecule inhibitors that non-covalently inhibit SENPs. The inhibitors confer the non-competitive inhibitory mechanism, as shown by nuclear magnetic resonance (NMR) and quantitative enzyme kinetic analysis. The NMR data also provided evidence for substrate-assisted inhibitor binding, which indicates the need for caution in using artificial substrates for compound screening, as the inhibitory effects could be significantly different from using the physiological substrates. This finding also suggests the possibility of designing inhibitors for this class of enzymes that are tuned for substrate-specificity.
Keywords: SUMO, SENP, inhibitor, NMR, enzyme kinetics
Post-translational modifications with the small ubiquitin-like modifiers (SUMO) are initiated and removed by the activities of SUMO-specific proteases (SENPs) (1). Unlike ubiquitylation, which has one modifier (i.e., ubiquitin) and one dominant role, namely protein degradation, SUMOylation involves three modifiers (SUMO-1, -2, and -3) and affects diverse cellular functions (2, 3). There are six SENPs, organized into three families based on sequence similarity: SENP1 and 2 that catalyze maturation of SUMO precursors and removal of SUMO-1 and SUMO-2/3 conjugates; SENP3 and 5 that preferentially remove SUMO-2/3 conjugates; and SENP6 and 7 that appear to be mainly involved in editing poly-SUMO-2/3 chains (4, 5). Recently, another de-SUMOylase has been discovered that does not share sequence similarity with the SENPs (6).
Small molecule inhibitors of SENPs are still in early development (7–11) and only one such inhibitor has been demonstrated to inhibit SENP in cells (9). SENP inhibitors with cellular activity would be advantageous for elucidating the role of SUMOylation in cellular regulation and for validating SENPs as therapeutic targets. SENP1 and SENP3 are potential targets for developing new therapeutic agents for cancer. They regulate the stability of hypoxia-inducible factor 1α (HIF1α), which is a key player in the formation of new blood vessels to support tumor growth (12–14). SENP1 is also highly expressed in human prostate cancer specimens and regulates androgen receptor (AR) activities (15–17). Androgen induces rapid and dynamic conjugation of SUMO-1 to AR, while SENP1 promotes AR-dependent transcription by cleaving SUMO-1-modified AR. SENP1 overexpression induces transformation of normal prostate gland tissue and facilitates the onset of high-grade prostatic intraepithelial neoplasia. Therefore, at least some members of the SENPs are potential targets for developing new cancer therapies.
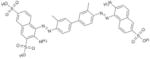
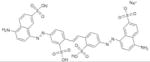
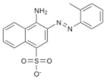
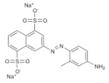
In this study, we set out to identify small molecule inhibitors of SENPs through in-silico screening in conjunction with enzyme kinetic, nuclear magnetic resonance (NMR) and cellular analyses. We performed in silico screening using PDB accession codes 2IYC and 2IY1 (18) and by considering hydrogen bonding and hydrophobic interactions between the C-terminus of full-length SUMO-1 and SENP1. The GLIDE program (19) was used to search the 250,000 compound library provided by the Developmental Therapeutics Program (DTP) of the National Cancer Institute, using the E-model scoring function of Cvdw, which is the sum of the van der Waals (Evdw) and electrostatic interaction energy terms (Eelec). Among the top hits, the dominant scaffolds were peptidomimetics and compounds that contained 2-fold symmetry. Forty compounds (100 μM) representing the dominant scaffolds were tested for their inhibitory effects on SENP1 and SENP2 for maturation of SUMO-1 and SUMO-2 precursors. The most potent compounds contained sulfonyl-benzene groups. Additional analogues of this group were obtained from DTP, and NSC5068, hereafter referred to as SPI-01 (SUMO protease inhibitor), was found to have the highest potency (Table 1). Available analogs of SPI-01 were obtained from DTP. Five compounds in this group (Table 1, SPI-06 to SPI-10) are “half” of the other compounds (Table 1, SPI-01 to SPI-05) and allowed the exploration of the activity requirements of the two-fold symmetric structure of SPI-01 to SPI-05. None of these molecules have been reported as inhibitors of SENPs. The inhibitory activity of these compounds on SENP1 and SENP2 was characterized using substrates that contained precursor SUMO-1 or SUMO-2 (S) flanked by yellow fluorescent protein (Y) at the N-terminus and enhanced cyan fluorescent protein (E) at the C-terminus (YSE) (20). Although the cleavage of the substrates can be detected by fluorescence resonance energy transfer (FRET), FRET could not be used because many of these compounds interfere with the FRET signal. Therefore, a gel-based assay was used to determine the inhibitory effects of all compounds on SENP1 and 2 (representative data shown in Figure 1A & 1B), and the gel bands were quantified to determine the half maximum inhibitory concentrations (IC50) (Table 1). The inhibitory effects of the compounds on the endopeptidase activities were not only enzyme-dependent, but also substrate-dependent. For SENP1-mediated cleavage of SUMO-1 precursor, only four of the compounds (SPI-01 to SPI-04) had half maximal inhibitory concentrations (IC50) below 60 μM. The inhibitors were more potent for inhibiting SENP2 than SENP1 for cleavage of the SUMO-1 precursor. However, for cleavage of the SUMO-2 precursor, some compounds (i.e. SPI-01 and SPI-04) had similar potency for inhibiting SENP1 and SENP2, while others (i.e. SPI-07 and SPI-10) were more potent for inhibiting SENP1 than SENP2 or vice versa (i.e. SPI-06 and SPI-09) (Table 1). In addition to the differential effects on SENP1 and SENP2, SPI-01 had more than 10 fold less potency for inhibiting a de-ubiquitin enzyme isopeptidase T than inhibiting SENP2 (Figure S1).
Table 1.
Effect of inhibitors on inhibition of the maturation of SUMO precursors by SENP1 and SENP2
| Compounds | IC50(μM) - SUM01 | IC50(μM) - SUM02 | ||||
|---|---|---|---|---|---|---|
| Structure | Code† | NCI ID‡ | SENP1 | SENP2 | SENP1 | SENP2 |
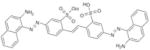
|
SPI-01 | NSC5068 | 32.8 ± 1.82 | 1.42 ± 3.0 | 1.88 ±2.2 | 1.1 ± 5.8 |
|
SPI-02 | NSC16224 | 26.5 ± 1.86 | 3.42 ± 1.6 | 2.08 ± 2.0 | 2.70 ± 2.1 |
|
|
SPI-03 | NSC8676 | 20.27 ± 2.47 | 5.17 ± 1.32 | 1.86 ± 2.3 | 3.0 ± 2.0 |
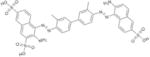
|
SPI-04 | NSC34933 | 11.2 ± 1.7 | 1.6 ± 2.5 | 2.32 ±2.6 | 2.15 ± 2.28 |
|
|
SPI-05 | NSC5067 | > 60 | 19.7 ± 1.47 | 7.5 ± 1.6 | 4.6 ± 1.65 |
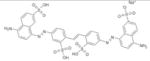
|
SPI-06 | NSC70551 | > 60 | 3.62 ± 1.98 | 43.2 ± 2.2 | 10.7 ± 1.6 |
|
SPI-07 | NSC58046 | > 60 | > 60 | 17.54 ± 4.9 | 28.06 ± 9.2 |
|
SPI-08 | NSC22940 | > 60 | 4.1 ± 3.0 | > 60 | 41.06 ± 5.2 |
|
SPI-09 | NSC42164 | > 60 | 23.36 ± 1.6 | > 60 | 26.96 ± 2.5 |
|
SPI-10 | NSC45551 | > 60 | 34.21 ± 1.9 | 11.1 ± 3.7 | 36.44 ± 5.7 |
Designation for our library of SUMO-protease inhibitors (SPI)
Designated by the National Cancer Institute
Figure 1. Identification and characterization of a new family of SENP inhibitors.

(A) and (B) Representative Coomassie-stained gels showing cleavage of SUMO-1 and SUMO-2 by SENP1 and SENP2 in the presence of increasing concentrations of SPI-01 (A) and SPI-07 (B). YSE, fusion SUMO (S) precursors flanked by YFP (Y) and ECFP (E) at the N- and C-termini, respectively. (C) Effects of the panel of inhibitors (Table 1) at inhibiting SENP1, 2 and 7. In 96-well plates, SENPs (50–200 nM) were pre-treated with increasing concentrations of each compound, after which DUB-Glo (40 μM final concentration; Promega) was added as substrate. Experiments were performed in triplicate. The amount of cleaved product is proportional to the relative light unit (RLU), which is bioluminescence produced by luciferase catalyzed reaction of luciferin that was produced by SENP cleavage of DUB-Glo. (D) Accumulation of SUMO-2/3-modified proteins in HeLa cells upon treatment with increasing doses of SPI-01. (E) Retention of SUMOylated proteins during recovery of HeLa cells from heat shock in the presence of 60 μM SPI-01 and SPI-02.
To determine whether other SENPs can be inhibited by this family of inhibitors, a distant SENP member, SENP7, was tested in parallel with SENP1 and SENP2 using a pentapeptide substrate that contained the Gly-Gly motif and luciferin, known as DUB-Glo (Promega). Cleavage of luciferin by a SENP can be detected by a coupled bioluminescent assay using luciferase. The bioluminescent reporter was chosen instead of a fluorescent reporter to avoid interference by the compounds during detection. In addition, because SENP7 has different physiological substrates than SENP1 and SENP2 (4, 5), an advantage of DUB-Glo is that it can act as a common substrate for all SENPs, which enabled us to rule out substrate-specific effects. The dose-dependent inhibition of each SENP by the inhibitors was determined (Figure 1C), as was the IC50 for inhibition of SENP1, 2 and 7 of all the compounds (Table 2). Most compounds had more similar inhibitory effects on SENP1 and SENP2 than on SENP7, consistent with their amino acid sequence similarities. In addition, the compounds were more potent for inhibiting SENP1 when DUB-Glo was used as a substrate than when SUMO-1 precursor was used (Table 1 and 2). To rule out the possibility that these compounds used a promiscuous mechanism, we also tested them in SUMOylation and ubiquitination reactions, which also depend on enzymes containing catalytic Cys residues. The compounds were noninhibitory in these assays (Figure S2). Furthermore, comparison of the DUB-Glo and the SUMO maturation assays revealed that the effect of SENP inhibitors could be highly substrate-specific.
Table 2.
Inhibitory effect on SENP enzymatic activity using a bioluminescent peptide substrate
| Structure | Code† | NCI ID‡ | SENP1 IC50(μM) | SENP2 IC50(μM) | SENP7 IC50(μM) |
|---|---|---|---|---|---|
|
|
SPI-01 | NSC5068 | 5.9 ± 1.4 | 2.9 ± 1.6 | 3.5 ± 1.5 |
|
SPI-02 | NSC 16224 | 2.1 ± 1.9 | 2.0 ± 2.0 | 2.7 ± 1.8 |
|
|
SPI-03 | NSC 8676 | 3.8 ± 1.5 | 2.4 ± 1.8 | 4.8 ± 1.4 |
|
SPI-04 | NSC 34933 | 2.4 ± 1.8 | 2.3 ± 1.8 | 3.4 ± 1.5 |
|
|
SPI-05 | NSC 5067 | 13.3 ± 1.3 | 8.5 ± 1.3 | 4.6 ± 1.5 |
|
SPI-06 | NSC 70551 | 3.9 ± 1.4 | 3.7 ± 1.4 | 4.7 ± 1.7 |
|
SPI-07 | NSC 58046 | ≫60 | ≫60 | 1.9 ± 2.2 |
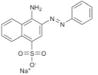
|
SPI-08 | NSC 22940 | 22.2 ± 1.5 | 17.2 ± 1.5 | 2.8 ± 1.6 |
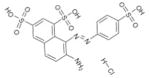
|
SPI-09 | NSC 42164 | > 60 | 6.8 ± 1.3 | 1.9 ± 2.1 |
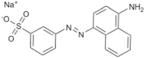
|
SPI-10 | NSC 45551 | 2.4 ± 1.8 | 2.5 ± 1.7 | 2.0 ± 2.0 |
Designation for our library of SUMO-protease inhibitors (SPI)
Designated by the National Cancer Institute
We then tested the abilities of representative inhibitors to inhibit SENP in cells. HeLa cells were treated with increasing concentrations of SPI-01 for 48 h, after which we detected SUMOylated proteins in the cells by Western blots. SUMO-2/3 conjugates accumulated in cells and this accumulation correlated with inhibitor concentration, particularly at high molecular weights (Figure 1D). This result suggests that SPI-01 inhibits the isopeptidase activities of SENPs, particularly SENP6 and SENP7, which are required for SUMO chain editing. We observed less significant effects on the accumulation of SUMO-1 conjugates, possibly because most SENPs cleave SUMO-2/3-conjugates (4, 5). It is known that heat shock triggers a dramatic increase in global SUMO-2/3 conjugations and that during recovery, the SUMOylated proteins are removed, at least in part, due to the deSUMOylation activity of SENP1 (21). To further confirm that the inhibitors inhibited deSUMOylase activities, we treated HeLa cells with SPI-01 and SPI-02 for 2 h at 37 °C. Then, SPI-treated or untreated control HeLa cells was transferred to 42 °C for 30 min, followed by recovery for 4 h at 37 °C before processing for detection of global SUMO-2/3 levels. The inhibitor-treated cells had considerably higher levels of SUMOylated proteins than did the corresponding controls that did not receive heat shock or the mock-treated cells after the recovery period (Figure 1E). Thus, the results of the heat-shock experiments further confirmed that the SPI compounds had inhibitory effects on SENPs in cells.
We used NMR chemical shift perturbation (CSP) analysis to investigate whether this family of inhibitors binds the enzyme or the enzyme-substrate complex. CSP experiments were conducted using a 15N-labeled C603S mutant of the human SENP1 catalytic domain (SENP1-C603S, for which NMR chemical shift assignments have been obtained and deposited in BMRB with accession number 19083). Although the SENP1-C603S mutant is catalytically inactive (22), it retains binding activity for the precursor or mature SUMO paralogs or SUMOylated substrates (18). We observed that SPI-01 caused modest backbone amide CSP for a subset of SENP1-C603S residues. Of note, specific CSPs were observed at the canonical cysteine-protease catalytic triad residues (D550, H533, and C603), the proposed dynamic channel of conserved W465 and W534, and at several other residues located at or adjacent to the SENP catalytic center (W465, L466, G531, H533, W534, C535, M552, G554 and Q596) with only one residue located distal to this surface (E469) (Figure 2A). Interestingly, M552, G554, and Q596 are clustered at the SENP1 surface that contacts the C-terminal tail of SUMO-1. Supporting the importance of this surface in SENP catalytic activity, non-conservative point mutations of Q596 in SENP1 or the equivalent residue to SENP1 M552 in SENP2 (M497) perturb SUMO processing and deconjugation (23, 24). Residue E469 is positioned toward the binding surface for the structured region of SUMO-1, and its CSP may be due to an alternative interaction with the compound or long-range effects. These results indicate that SPI-01 binds the surface adjacent to the catalytic center that contacts the C-terminal portion of the SUMO precursors. The residues that showed CSP are highly conserved between SENP1 and SENP2, suggesting that SPI-01 can interact with the equivalent surface on SENP2.
Figure 2. Investigation of the inhibitory mechanism using NMR and enzyme kinetic analysis.

(A) Superimposition of a section of the 2D1H-15N-HSQC spectra of the catalytically inactive C603S mutant of human SENP1 in the absence (black cross-peaks) and presence of SPI-01 (green cross-peaks) at 25 °C. Perturbed representative cross-peaks at or near the catalytic site of SENP1 are labeled. (B) Superimposition of a section of the 2D1H-15N-HSQC spectra of SUMO-1 precursor showing labeled peaks of the C-terminal residues when free (black) and bound to SENP1-C603S (red) or both SENP1-C603S and SPI-01 (green) at 35 °C. (C) All SPI-01 perturbed residues on SENP1 are labeled and colored in green on the surface representation of SENP1 in complex with SUMO-1 precursor (pdb ID: 2IY1). Perturbed residues that are located in the vicinity of the catalytic center of SENP1 or the C-terminus of precursor SUMO-1 are labeled in black and red, respectively. (D) Enzyme kinetic measurements for SPI-01 indicate a non-competitive mode of inhibition. The data were fit to obtain the indicated kinetic parameters (α, Ki and Km) using Graphpad Prism. Lineweaver-Burk plot analysis of the data also confirmed non-competitive inhibition.
We also investigated the binding of SPI-01 to the enzyme-substrate complex. We carried out CSP analysis on the 40 kDa complex of 15N-labeled full length precursor SUMO-1-GGHSTV (SUMO-1-FL) with unlabeled SENP1-C603S. An equimolar amount of SPI-01 was added to the 1:1 enzyme-substrate complex. The only observed CSP on the 15N-labeled precursor SUMO-1-FL was on the C-terminal residues S99 and V101 (Figure 2B and 2C, and Table S1) (25). This result indicates that SPI-01 binds the enzyme-substrate complex at the interface between SENP and the C-terminal tails of precursor SUMO-FL. X-ray crystal structures showed that the C-terminal tail of precursor SUMO sits in and projects out of the catalytic tunnel of SENPs (18). In the case of SENP1, the region that interacts with the projected C-terminus is predominantly acidic and favors the C-terminus of SUMO-1, which is polar and positively charged, over that of SUMO-2, whose C-terminus is mainly hydrophobic (18, 26). In addition, the more hydrophobic C-terminus of SUMO-2 may favor binding of aromatic inhibitors. These properties may account for the more potent inhibition of processing of the SUMO-2 precursor (Table 1).
To further investigate the inhibitory mechanism, enzyme kinetic experiments were conducted using the pentapeptide substrate DUB-Glo (Figure 2D). The data was fit to a mixed inhibition mechanism, as described by the kinetic equation:
in which the value of “α” indicates the mechanism of inhibition (27). For both SENP1 and SENP2, the “α” values indicated that the inhibitory mechanism is mainly noncompetitive and suggests that the inhibitor binds to the enzyme and the enzyme-substrate complex to inhibit chemical conversion. This finding is consistent with the NMR binding analysis indicating that the inhibitor binds both the enzyme and the enzyme-substrate complex as discussed above.
In conclusion, this study has identified a new chemotype of SENP inhibitors that do not covalently modify the catalytic Cys residue. This study has also provided the first mechanistic insights into how a small molecule inhibitor of SENPs that does not covalently modify the catalytic Cys can inhibit the enzymes. The substrate-assisted inhibitor binding indicates the need for caution in designing high throughput screening assays that use fluorogenic or chemiluminescent artificial substrates, as the results could be significantly different from using the physiological substrates. The substrate-dependent inhibitory effect suggests the possibility of designing SENP inhibitors that are tuned for substrate-specificity.
Methods
Protein Purification
The catalytic domains of SENP1, 2, and 7 were expressed as His-tagged protein in E. coli (DE3) and purified using nickel affinity chromatography (28). The pET11 expression plasmids for SENP1 and 2 contained a cDNA insert coding for the catalytic domain of human SENP1-WT (419–644) and SENP2-WT (364–589). The expression plasmid for the SENP1 active site point mutant C603S was generated using the QuikChange mutagenesis kit (Agilent Technologies). The expression plasmid for the catalytic domain of SENP7 was obtained from Dr. Salvesen’s laboratory through Addgene (29).
SUMO Cleavage Assays
SUMO cleavage assays were performed by incubating SENPs with various concentrations of the inhibitor (0–60 μM) at room temperature for 10 min in assay buffer (50 mM Tris, pH 7.4, 100 mM NaCl, 10 mM DTT). SENP concentrations were 32–50 nM when 50 μg/ml of the final substrate YFP-SUMO-ECFP (YSE) fusion protein was added. The mixture was incubated (37 °C, 15 min), followed by SDS-PAGE and Coomassie staining for visualization. For cellular SENP inhibition experiments, HeLa cells cultured in DMEM plus 10% FBS, 100 units/ml penicillin, 100 mg/ml streptomycin, and 0.2 M glutamine were treated for 48 h with SPI compounds. For heat shock experiment, HeLa cells were treated with SPI compounds or mock treated (2 h, 37 °C), after which cells were transferred to 42 °C for 30 min. After heat shock, the cells were allowed to recover (4–5 h) before being harvested and lysed. Proteins were separated by SDS-PAGE and immunoblotted to determine global SUMO-2/3 levels.
DUB-Glo Assay
The luciferase substrate assay (DUB-Glo, Promega) was performed according to the manufacturer’s instructions. Briefly, SENPs (final concentration 50–100 nM) in Tris buffer (50 mM Tris, pH 8.0, 100 mM NaCl, 10 mM DTT) were pre-incubated (10 min, room temperature) with increasing concentrations of inhibitor (0–60 μM final concentration) followed by addition of the luciferase substrate. Luciferase output was recorded 30 min after addition of the luciferase substrate. Values are the averages of experiments performed in triplicate.
NMR Experiments
Samples used for NMR titration or chemical shift perturbation analyses were 15N or 15N/13C-labeled; the titrant protein or SPI-01 was not labeled. The 15N/13C SUMO-1-FL sample was used to extend the backbone assignments of mature SUMO-1 to the HSTV tail by using 2D-NH-HSQC, 3D-HNCA, 3D-HNCOCA, and 3D-HNCACB. Additionally, comparison of NH-HSQC between precursor and mature SUMO quickly identified the resonances of the HSTV tail. For SENP1 assignments, a full suite of triple-resonance NMR experiments were acquired on 15N/13C/2H or 15N/13C samples: HNCA, HNCOCA, HNCACB, HNCOCACB, HNCO, HNCACO, and NOESY-HSQC. All samples were dissolved in the NMR buffer: 20 mM sodium phosphate (pH 6.8), 10% D2O, 0.03% sodium azide and 10 mM d10-dithiothreitol. Purified perdeuterated SENP1 samples were unfolded and refolded into NMR buffer.
For titration of SENP1-C603S with SPI-01, 270 μM 15N-labeled sample was titrated with the inhibitor that was prepared by diluting a 10 mM stock in 100% DMSO-d6 to a concentration of 1.7 mM in the NMR buffer. The 2D 1H-15N-HSQC spectra of SENP1 were recorded at each incremental addition of 5 μl of SPI-01 into 250 μl of SENP1. The chemical shift perturbation (CSP) analysis compared the spectra of SENP1 in the absence or the presence of equimolar SPI-01. A separate DMSO control titration was performed to account for DMSO-induced CSP. NMR resonance assignments for SUMO samples at 35 °C were transferred from those obtained at 25 °C by spectral acquisition at 2.5 °C incremental increases. All data were acquired on a 600 MHz Bruker Avance NMR spectrometer equipped with a TXI Cryoprobe.
Supplementary Material
Acknowledgments
This work was supported by NIH Grants R01GM074748, R01GM086171 and GM102538 to Y Chen.
Abbreviations
- SUMO
small ubiquitin-like modifier
- SENP
SUMO-specific proteases
- SPI
SUMO protease inhibitor
- NMR
nuclear magnetic resonance
- CSP
chemical shift perturbation
Footnotes
Supporting Information
This material is available free of charge via the Internet http://pubs.acs.org.
References
- 1.Yeh ET. SUMOylation and De-SUMOylation: wrestling with life’s processes. The Journal of biological chemistry. 2009;284:8223–8227. doi: 10.1074/jbc.R800050200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Melchior F. SUMO—nonclassical ubiquitin. Annu Rev Cell Dev Biol. 2000;16:591–626. doi: 10.1146/annurev.cellbio.16.1.591. [DOI] [PubMed] [Google Scholar]
- 3.Hay RT. SUMO: a history of modification. Molecular cell. 2005;18:1–12. doi: 10.1016/j.molcel.2005.03.012. [DOI] [PubMed] [Google Scholar]
- 4.Kolli N, Mikolajczyk J, Drag M, Mukhopadhyay D, Moffatt N, Dasso M, Salvesen G, Wilkinson KD. Distribution and paralogue specificity of mammalian deSUMOylating enzymes. Biochemical Journal. 2010;430:335–344. doi: 10.1042/BJ20100504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Shen LN, Geoffroy MC, Jaffray EG, Hay RT. Characterization of SENP7, a SUMO-2/3-specific isopeptidase. The Biochemical journal. 2009;421:223–230. doi: 10.1042/BJ20090246. [DOI] [PubMed] [Google Scholar]
- 6.Shin EJ, Shin HM, Nam E, Kim WS, Kim JH, Oh BH, Yun Y. DeSUMOylating isopeptidase: a second class of SUMO protease. EMBO Rep. 2012 doi: 10.1038/embor.2012.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Qiao Z, Wang W, Wang L, Wen D, Zhao Y, Wang Q, Meng Q, Chen G, Wu Y, Zhou H. Design, synthesis, and biological evaluation of benzodiazepine-based SUMO-specific protease 1 inhibitors. Bioorganic & medicinal chemistry letters. 2011;21:6389–6392. doi: 10.1016/j.bmcl.2011.08.101. [DOI] [PubMed] [Google Scholar]
- 8.Xu Y, Li J, Zuo Y, Deng J, Wang LS, Chen GQ. SUMO-specific protease 1 regulates the in vitro and in vivo growth of colon cancer cells with the upregulated expression of CDK inhibitors. Cancer letters. 2011;309:78–84. doi: 10.1016/j.canlet.2011.05.019. [DOI] [PubMed] [Google Scholar]
- 9.Uno M, Koma Y, Ban HS, Nakamura H. Discovery of 1-[4-(N-benzylamino)phenyl]-3-phenylurea derivatives as non-peptidic selective SUMO-sentrin specific protease (SENP)1 inhibitors. Bioorganic & medicinal chemistry letters. 2012;22:5169–5173. doi: 10.1016/j.bmcl.2012.06.084. [DOI] [PubMed] [Google Scholar]
- 10.Albrow VE, Ponder EL, Fasci D, Bekes M, Deu E, Salvesen GS, Bogyo M. Development of small molecule inhibitors and probes of human SUMO deconjugating proteases. Chemistry & biology. 2011;18:722–732. doi: 10.1016/j.chembiol.2011.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Borodovsky A, Ovaa H, Meester WJ, Venanzi ES, Bogyo MS, Hekking BG, Ploegh HL, Kessler BM, Overkleeft HS. Small-molecule inhibitors and probes for ubiquitin- and ubiquitin-like-specific proteases. Chembiochem: a European journal of chemical biology. 2005;6:287–291. doi: 10.1002/cbic.200400236. [DOI] [PubMed] [Google Scholar]
- 12.Cheng J, Kang X, Zhang S, Yeh ET. SUMO-specific protease 1 is essential for stabilization of HIF1alpha during hypoxia. Cell. 2007;131:584–595. doi: 10.1016/j.cell.2007.08.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Xu Y, Zuo Y, Zhang H, Kang X, Yue F, Yi Z, Liu M, Yeh ET, Chen G, Cheng J. Induction of SENP1 in endothelial cells contributes to hypoxia-driven VEGF expression and angiogenesis. The Journal of biological chemistry. 2010 doi: 10.1074/jbc.M110.164236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Huang C, Han Y, Wang Y, Sun X, Yan S, Yeh ET, Chen Y, Cang H, Li H, Shi G, Cheng J, Tang X, Yi J. SENP3 is responsible for HIF-1 transactivation under mild oxidative stress via p300 de-SUMOylation. EMBO J. 2009;28:2748–2762. doi: 10.1038/emboj.2009.210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cheng J, Bawa T, Lee P, Gong L, Yeh ET. Role of desumoylation in the development of prostate cancer. Neoplasia. 2006;8:667–676. doi: 10.1593/neo.06445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bawa-Khalfe T, Cheng J, Lin SH, Ittmann MM, Yeh ET. SENP1 induces prostatic intraepithelial neoplasia through multiple mechanisms. The Journal of biological chemistry. 2010;285:25859–25866. doi: 10.1074/jbc.M110.134874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kaikkonen S, Jaaskelainen T, Karvonen U, Rytinki MM, Makkonen H, Gioeli D, Paschal BM, Palvimo JJ. SUMO-specific protease 1 (SENP1) reverses the hormone-augmented SUMOylation of androgen receptor and modulates gene responses in prostate cancer cells. Mol Endocrinol. 2009;23:292–307. doi: 10.1210/me.2008-0219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Shen L, Tatham MH, Dong C, Zagorska A, Naismith JH, Hay RT. SUMO protease SENP1 induces isomerization of the scissile peptide bond. Nat Struct Mol Biol. 2006;13:1069–1077. doi: 10.1038/nsmb1172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Friesner RA, Banks JL, Murphy RB, Halgren TA, Klicic JJ, Mainz DT, Repasky MP, Knoll EH, Shelley M, Perry JK, Shaw DE, Francis P, Shenkin PS. Glide: a new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. Journal of medicinal chemistry. 2004;47:1739–1749. doi: 10.1021/jm0306430. [DOI] [PubMed] [Google Scholar]
- 20.Tatham MH, Hay RT. FRET-based in vitro assays for the analysis of SUMO protease activities. Methods Mol Biol. 2009;497:253–268. doi: 10.1007/978-1-59745-566-4_17. [DOI] [PubMed] [Google Scholar]
- 21.Nefkens I, Negorev DG, Ishov AM, Michaelson JS, Yeh ETH, Tanguay RM, Muller WEG, Maul GG. Heat shock and Cd2+ exposure regulate PML and Daxx release from ND10 by independent mechanisms that modify the induction of heat-shock proteins 70 and 25 differently. J Cell Sci. 2003;116:513–524. doi: 10.1242/jcs.00253. [DOI] [PubMed] [Google Scholar]
- 22.Xu Z, Chau SF, Lam KH, Chan HY, Ng TB, Au SWN. Crystal structure of the SENP1 mutant C603S-SUMO complex reveals the hydrolytic mechanism of SUMO-specific protease. Biochem J. 2006;398:345–352. doi: 10.1042/BJ20060526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Reverter D, Lima CD. Structural basis for SENP2 protease interactions with SUMO precursors and conjugated substrates. Nat Struct Mol Biol. 2006;13:1060–1068. doi: 10.1038/nsmb1168. [DOI] [PubMed] [Google Scholar]
- 24.Shen LN, Dong CJ, Liu HT, Naismith JH, Hay RT. The structure of SENP1-SUMO-2 complex suggests a structural basis for discrimination between SUMO paralogues during processing. Biochem J. 2006;397:279–288. doi: 10.1042/BJ20052030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Song J, Durrin LK, Wilkinson TA, Krontiris TG, Chen Y. Identification of a SUMO-binding motif that recognizes SUMO-modified proteins. Proceedings of the National Academy of Sciences of the United States of America. 2004;101:14373–14378. doi: 10.1073/pnas.0403498101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Shen LN, Dong C, Liu H, Naismith JH, Hay RT. The structure of SENP1-SUMO-2 complex suggests a structural basis for discrimination between SUMO paralogues during processing. The Biochemical journal. 2006;397:279–288. doi: 10.1042/BJ20052030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Segel IEK. Enzyme Kinetics. John Wiley & Sons; 1993. [Google Scholar]
- 28.Namanja AT, Li YJ, Su Y, Wong S, Lu J, Colson LT, Wu C, Li SS, Chen Y. Insights into High Affinity Small Ubiquitin-like Modifier (SUMO) Recognition by SUMO-interacting Motifs (SIMs) Revealed by a Combination of NMR and Peptide Array Analysis. The Journal of biological chemistry. 2012;287:3231–3240. doi: 10.1074/jbc.M111.293118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mikolajczyk J, Drag M, Bekes M, Cao JT, Ronai Z, Salvesen GS. Small ubiquitin-related modifier (SUMO)-specific proteases: profiling the specificities and activities of human SENPs. The Journal of biological chemistry. 2007;282:26217–26224. doi: 10.1074/jbc.M702444200. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.