

Abstract
Video game skills transfer to other tasks, but individual differences in performance and in learning and transfer rates make it difficult to identify the source of transfer benefits. We asked whether variability in initial acquisition and of improvement in performance on a demanding video game, the Space Fortress game, could be predicted by variations in the pretraining volume of either of 2 key brain regions implicated in learning and memory: the striatum, implicated in procedural learning and cognitive flexibility, and the hippocampus, implicated in declarative memory. We found that hippocampal volumes did not predict learning improvement but that striatal volumes did. Moreover, for the striatum, the volumes of the dorsal striatum predicted improvement in performance but the volumes of the ventral striatum did not. Both ventral and dorsal striatal volumes predicted early acquisition rates. Furthermore, this early-stage correlation between striatal volumes and learning held regardless of the cognitive flexibility demands of the game versions, whereas the predictive power of the dorsal striatal volumes held selectively for performance improvements in a game version emphasizing cognitive flexibility. These findings suggest a neuroanatomical basis for the superiority of training strategies that promote cognitive flexibility and transfer to untrained tasks.
Keywords: basal ganglia, caudate nucleus, cognitive flexibility, nucleus accumbens, procedural learning
Introduction
The pervasiveness of video game use across cultures, coupled with evidence that video game experts outperform novices on many basic measures of attention and perception, has increased interest in using these games for instructional and training purposes. Playing video games for as few as 10–20 h has been shown to improve performance on a number of attentional and perceptual tasks (Green and Bavelier 2003, 2006, 2007) and on tasks requiring executive control (Basak et al. 2008). Such evidence has led to the development of video games that purportedly improve memory, attention, processing speed, and performance in daily life. The assumption underlying these assertions is that skills acquired through training on one task (i.e., the video game) transfer to other untrained tasks, including complex real world tasks (Boot et al. 2008).
The evidence that video game training improves performance on untrained tasks is promising. Several studies have reported transfer of training from video games to untrained behaviors and tasks (Frederickson and White 1989; Green and Bavelier 2003, 2006, 2007; Basak et al. 2008). For example, training on the Space Fortress video game, a video game specifically designed to study the cognitive effects of training (Donchin et al. 1989), predicted the success of Israeli Air Force flight school cadets in learning flight control (Gopher et al. 1994). However, others have found that 20-plus hours of action video game practice provided no specific benefits for novice video gamers across a wide battery of cognitive tasks, even though expert gamers outperformed the novices on many of the same tasks (Boot et al. 2008). These contrasting findings raised the possibility that other factors contribute to the differences in performance between expert video gamers and nongamers, including self-selection effects originating from preexisting individual differences (Boot et al. 2008).
If preexisting individual differences contribute to a self-selection effect, they might also predict variability in learning rates. In the experiments reported here, we reasoned that such preexisting differences might include differences in brain structures important for procedural learning and cognitive flexibility, and that it might therefore be possible to predict learning rates on complex tasks such as video games by analyzing preexisting differences in brain structures. We focused on the striatum, a key node in procedural learning circuits (Doyon et al. 2003; Graybiel 2005, 2008; Yin and Knowlton 2006; Balleine et al. 2009). The caudate nucleus and putamen, which together make up the dorsal striatum, have been convincingly implicated in procedural and habit learning and in the execution of learned behaviors. These regions are also activated while performing tasks that require cognitive flexibility such as task-switching and transfer to untrained tasks (Ragozzino et al. 2002; Cools et al. 2004; Meiran et al. 2004; Dahlin et al. 2008; Kimchi and Laubach 2009). The nucleus accumbens, in the ventral striatum, is part of the limbic circuitry related to reinforcement and motivation (Belin and Everitt 2008; Graybiel 2008; De Martino et al. 2009), and evidence suggests that the nucleus accumbens is recruited during the early stages of learning (Hernandez et al. 2002; O'Doherty et al. 2004; Atallah et al. 2007). Positron emission tomography studies in humans have shown that dopamine release and binding are increased in both of these striatal regions when subjects play a video game, and that greater dopamine binding is associated with better performance (Koepp et al. 1998).
To test directly whether the differential size of these striatal regions could be used to predict learning on an unfamiliar video game, we used magnetic resonance imaging (MRI)-based brain volumetry to measure striatal volumes of subjects with little previous video game experience before they received training on the classic Space Fortress video game. As a control brain structure, we analyzed the volume of the hippocampus, implicated in declarative memory formation (Squire et al. 2004). We used 2 different training strategies, one version emphasizing cognitive flexibility (variable priority training) and the other version not (fixed priority training). Variable priority training procedures periodically shift the emphasis of training from one task component to another, enhance learning rates and retention (Fabiani et al. 1989), and induce transfer to untrained tasks more effectively than procedures that employ a fixed priority training strategy (Kramer et al. 1999; Bherer et al. 2008). Thus, we reasoned that training emphasizing task-shifting might capitalize on the circuitry of the striatum more than training emphasizing overall performance on the entire task. We also compared the predictive value of the brain measures not only for the entire training periods, but also for different phases of learning including the initial acquisition period when performance was lowest but performance gains were highest.
We show here that striatal volumes, but not hippocampal volumes, predict learning improvements on a video game. Moreover, we found a dissociation between the predictive power of the dorsal striatal volumes and that of the ventral striatal volumes. The volume of the dorsal striatum positively predicted performance improvements for those individuals trained with strategies promoting cognitive flexibility, whereas the volume of the ventral striatum did not. But during early learning stages, both the volume of the ventral striatum and the volume of the dorsal striatum positively predicted performance improvements. These findings suggest that individual structural differences in the striatum are effective predictors of procedural learning and cognitive flexibility and are sensitive indicators of ventral-to-dorsal differences in striatal recruitment during learning.
Materials and Methods
Participants
Forty-two participants (age: 18–28 years, 12 males) were recruited from the Urbana-Champaign community either through flyers posted in campus buildings and businesses or through advertisements posted to online bulletin boards. Individuals responding to these advertisements were then asked to complete a survey of their video game habits and to return this survey via e-mail. All chosen participants reported normal or corrected-to-normal visual acuity, normal color vision, and normal hearing, were right-handed, reported not being on any medications, and met all criteria for participating in an MRI study including no previous head trauma, no previous head or neck surgery, no diagnosis of diabetes, no neuropsychiatric or neurological condition including brain tumors, and no metallic implants that could interfere with or cause injury due to the magnetic field. All signed an informed consent approved by the University of Illinois Internal Review Board. Finally, subjects were chosen for the study only if they reported playing less than 3 h of video games a week during the 2 years prior to the study. Of the original 42 individuals accepted, 39 completed the study. Three were excluded from the data analysis due to errors in processing of the MRI data. Participants were paid $15 an hour for testing and training. These subjects were randomly assigned to receive either fixed priority or variable priority training procedures (18 in each group: 6 males in variable priority and 4 males in fixed priority).
Apparatus
Ten networked computers were used to collect performance data. The Space Fortress game was displayed on 19-inch LCD monitors, and participants made game responses with the computer mouse and a Logitech Attack 3 Joystick.
Stimuli and Procedures
Participants were given instructions about the Space Fortress game and then, before game training, completed an MRI session. They were then given 20 h of Space Fortress game training (ten 2-h training sessions).
The Space Fortress game (see Fig. 2) requires players to navigate their ship with precise control using a joystick. The ship moves in a frictionless environment, and players can rotate the ship by moving the joystick left or right or by applying a thrust by pushing forward on the joystick. The ship has no braking system, so that in order for players to slow or stop the ship, they must rotate it so that it faces the direction opposite to its current direction of motion and apply a thrust. This requirement makes control of the ship a challenging and demanding task.
Figure 2.
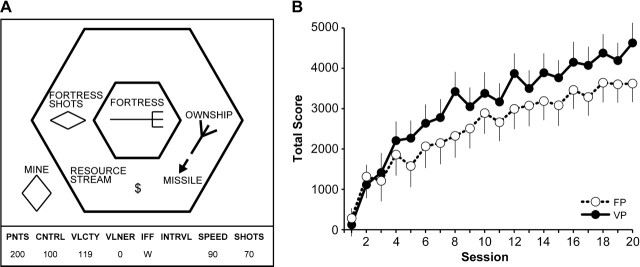
(A) Schematic diagram of the Space Fortress display seen by the participants. (B) Change in Total score for the fixed priority (FP) training group (open circles and dashed line) and for the variable priority (VP) training group (solid black circles and solid line). Both groups showed significant improvements in performance on the Space Fortress game.
The main goal of the Space Fortress game is for the players to destroy the fortress (at the center of the screen) as many times as possible while avoiding damage to their own ship. To destroy the fortress, players must hit the fortress with missiles by aiming toward it and pushing the fire button on the joystick. For the fortress to become vulnerable to destruction, it must first be hit with 10 missiles. The intervals between missiles must be at least 250 ms, so timing is an important component of successful performance. After 10 missile hits, the fortress can be destroyed by hitting it with a rapid double shot (2 missile hits separated by less than 250 ms). If participants hit the fortress with a double shot before it is vulnerable, the vulnerability of the fortress is reset to zero, and the player must start over again accumulating hits. To make the task even more challenging, the fortress rotates and shoots back at the player's ship, so that the player needs to keep his own ship in constant motion to avoid damage.
At regular intervals, mines appear on the screen. These objects can also damage the player's ship if they come in contact with it. Mines actively pursue the ship. Importantly, the fortress cannot be damaged or destroyed as long as a mine is on the screen, so mines must be dealt with as soon as possible. Each mine has a letter associated with it, and the letters are shown on an instrument panel displayed at the bottom of the screen. The letter identifies each mine as friend or foe, and at the beginning of each game, participants are asked to remember 3 letters that represent foe mines, with all other letters identifying friendly mines. If the mine that appears is a friendly mine, the player can shoot it, and the friendly mine will transfer this damage to the fortress. However, if it is a foe mine, it must be flagged as such using the mouse and then destroyed with a missile. Responding to mines incorrectly (identifying a friend as a foe or vice versa) has negative consequences in terms of the total score, and thus participants must be careful to remember which letters represent foe mines.
Finally, there is a constant monitoring task embedded in the Space Fortress game. A stream of symbols appears below the fortress. Whenever a dollar sign symbol appears twice in a row, players can use the mouse to select either bonus points or bonus missiles (which are a limited resource). However, if players incorrectly identify the first dollar symbol as the second, they miss their opportunity to obtain a bonus when the second symbol appears. Thus, participants are always encouraged to monitor this information.
Points are awarded to players based on their performance of the Space Fortress game. They are given a Total score and also sub-scores that reflect different aspects of their performance. Different actions add to, or subtract from, different sub-scores, and these are displayed in the instrument panel located at the base of the screen during the game. For example, participants are asked to keep their ship in the space between the 2 hexagons on the screen (see Fig. 2). Doing so increases the Control sub-score. Flying the ship outside of the large hexagon or leaving the screen entirely (going into hyperspace) subtracts from the Control sub-score. The Velocity sub-score increases when the players move the ship at slow speeds and decreases when they move it at high speeds. The Speed sub-score rewards/punishes participants for how quickly they deal with mines, and the Points sub-score increases when players shoot and destroy the fortress, but points are subtracted for damage and destruction of the player's ship.
Training Procedure
Before the MRI session, each subject was first familiarized with the Space Fortress game by completing the Aiming Task. In this task, subjects used the joystick to rotate their ship (which was in a fixed location at the center of the screen) and the fire button to destroy mines that appeared randomly on the screen. The objective was to destroy as many mines as possible. Participants completed 3 trials of the Aiming Task, with each trial lasting about 1 min. Next, participants watched a movie that demonstrated all the rules of the Space Fortress game and were given printed instructions to refer to. Then participants were given 1.5 h of Space Fortress experience, in which they completed twenty-four 3-min games of Space Fortress and were instructed to try to obtain the highest Total score.
After this initial familiarization session, the training of the variable priority group and the fixed priority group diverged. All participants completed ten 2-h sessions consisting of thirty-six 3-min games. The block and trial structures were identical for the 2 groups. Within each session, performance was assessed twice (once at the beginning of the session and once at the end). Each session started and ended with 3 test game trials (baseline trials), in which participants were asked to maximize performance and focus on obtaining the highest Total score. This resulted in 20 actual assessments of performance with each one averaging performance over 6 games (3 before practice and 3 after practice). In between these baseline games, the subjects completed 30 practice games per session. For the fixed priority group, subjects were always asked to maximize the Total score during training and were reminded that Total score was the sum of the Control, Velocity, Speed, and Points sub-scores. This resulted in 20 assessments of performance over the training period. Participants in the fixed priority group were told to emphasize each of these subcomponents of the game equally. They completed 5 blocks of 6 trials each. For the variable priority group, participants were asked to focus their resources on improving and monitoring different sub-scores of the game during the 30 practice games. They were given 5 blocks of 6 trials each, in which they were asked to emphasize a particular aspect of the Space Fortress game, and this emphasis changed every 6 trials. Importantly, the subjects were instructed not to ignore other aspects of the game but just to put particular emphasis on one of them at any given time. On even-numbered sessions, participants completed emphasis blocks in the following order: Control, Velocity, Speed, Points, and Total. On odd-numbered sessions, they completed the same emphasis blocks in the reverse order. For both groups, the total training consisted of 360 games of Space Fortress. Participants completed 3–5 two-hour sessions per week.
MR Imaging Protocol and Image Processing
High resolution (1.3 × 1.3 × 1.3 mm) T1-weighted brain images were acquired using a 3D magnetization prepared rapid gradient echo imaging protocol with 144 contiguous slices collected in an ascending fashion. All images were collected on a 3-T Siemens Allegra scanner with an echo time = 3.87 ms, repetition time = 1800 ms, field of view = 256 mm, an acquisition matrix of 192 × 192 mm, and a flip angle of 8°.
For segmentation and volumetric analysis of the left and right striatum and hippocampus, we employed functional magnetic resonance imaging of the brain (FMRIB)'s Integrated Registration and Segmentation Tool (FIRST) in FMRIB's Software Library (FSL) version 4.0. FIRST is a semiautomated model-based segmentation tool utilizing a Bayesian framework based on shape and appearance models obtained from manually segmented images by the Center for Morphometric Analysis, Massachusetts General Hospital, Boston, MA. Structural and landmark information were obtained from 317 manually segmented and labeled T1-weighted images of the brain from normal children, adults, and clinical populations and were modeled as a point distribution model, in which the geometry and variation of the shape of the structure are submitted as priors. Volumetric labels are parameterized by a 3D deformation of a surface model based on multivariate Gaussian assumptions. FIRST then searches through linear combinations of shape modes of variation for the most probable shape given the intensity distribution in the T1-weighted image (Patenaude et al. 2007a, 2007b).
This method first runs a 2-stage affine registration to a standard space template (Montreal Neurological Institute space) with 1-mm resolution using 12 degrees of freedom and a subcortical mask to exclude voxels outside the subcortical regions. Second, the caudate nucleus, putamen, nucleus accumbens, and hippocampus are segmented with 30, 40, 50, and 30 modes of variation, respectively, for each structure. Finally, boundary correction takes place for each structure so that the boundary voxels are classified as belonging to the structure or not based on a statistical probability (z score > 3.00; P < 0.001). In the current study, the volume of each structure was measured in cm3. The segmentation results for each participant were visually inspected for any significant error that could have occurred during the segmentation process. No errors were noted.
Intracranial volume (ICV) is frequently used to adjust the regional volumes for gender and for height (Raz et al. 2005). Here, we calculated ICV as the sum of gray, white, and cerebrospinal fluid and adjusted the volume of each region by this measure using FMRIB's automated segmentation tool in FSL version 4.0 (Zhang et al. 2001; Smith et al. 2004). In accordance with other volumetric analyses, adjustment was performed for each region by an analysis of covariance approach: adjusted volume = raw volume – b × (ICV – mean ICV), where b is the slope of a regression of an region of interest volume on ICV (Raz et al. 2005; Erickson et al. 2009; Kennedy et al. 2009). Adjusted volume was used for all analyses described in this manuscript.
To assure reliability of the segmentation algorithm for these structures, we ran an additional MRI scan on all subjects 2 weeks after the completion of the training program. We employed the same segmentation algorithm described above to these images and assessed the test–retest reliability of the segmentation algorithm. We found high reliability for all regions using Cronbach’s alpha. The hippocampus, caudate nucleus, and putamen had Cronbach α values greater than 0.94, and the values for the nucleus accumbens were at α = 0.90. Thus, the segmentation algorithm was able to identify the locations of the structures of interest across scans with high consistency.
Analyses
Behavioral performance on the Space Fortress game was analyzed by a repeated measures analysis of variance (ANOVA) with 2 factors: practice level (within-subjects) and training group (between-subjects). Total scores and sub-scores were analyzed individually.
We examined whether improvement on any measure of the Space Fortress game was associated with volumes of either the dorsal or ventral striatum or the hippocampus. We calculated the difference in performance between the first and last sessions to obtain a measure of improvement for the Total score and each of the sub-scores. These difference values were then entered into a series of multiple regression analyses with initial performance entered as a covariate and the difference in performance (performance improvement) entered as the dependent variable. Volume was used as the independent variable to predict performance. We calculated standardized beta values (β) and significance values. These multiple regression analyses were conducted in one omnibus analysis collapsing across training groups. Interaction terms were also included to determine whether one training group profited more from variation in volume of one or more of the sites examined (group × regional volume interaction).
To examine whether any brain region would be related more to learning rates in the early training sessions than later sessions, we divided the 20 two-hour training sessions into 4 and calculated improvements in performance by taking the difference between the first and last sessions in each phase. We then used these values in a multiple regression analysis to determine whether regional brain volume was related to performance improvements early in task acquisition. Initial performance for each phase was entered as a covariate in the model. Standardized beta values (β) and significance values are reported.
Results
Imaging of the Striatum and Hippocampus
We imaged the striatum and hippocampus with high-resolution MRI before the video game training but after familiarization with the video game (see Materials and Methods). To determine the location, size, shape, and boundaries of striatal and hippocampal regions, we used an automated segmentation algorithm that employs a point distribution model from manual tracing of defined regions (Fig. 1). We also normalized the measurements. After segmentation, the volume of each region was calculated based on voxel dimensions and adjusted for total ICV. Total ICV was used to adjust for variation in total brain and head size, accounting for sex differences, by multiple regression. The normalized volumes of the left and right caudate nucleus ranged from 3.80 to 7.43 cm3 (mean = 5.33; standard deviation [SD] = 0.85), those of the left and right putamen ranged from 3.61 to 6.81 cm3 (mean = 5.22; SD = 0.68), and those of the left and right nucleus accumbens ranged from 0.425 to 1.45 cm3 (mean = 0.77; SD = 0.22). The hippocampal volumes for the left and right sides ranged from 3.46 to 6.55 cm3 (mean = 5.10; SD = 0.63).
Figure 1.
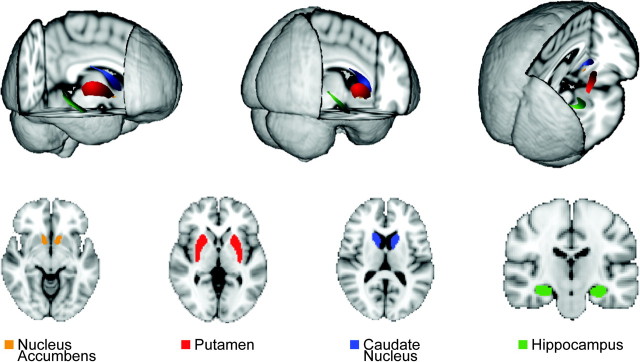
Regions studied. Display in 3D and 2D images for the segmentations used to identify the nucleus accumbens (orange), putamen (red), caudate nucleus (blue), and hippocampus (green).
Behavioral Learning during Training on the Space Fortress Video Game
Training reliably improved performance on the Space Fortress game (Fig. 2). The combined Total scores for subjects trained with the fixed priority and variable priority strategies improved significantly with training across the 4 phases (F3,102 = 153.45, P < 0.001), as did each of the sub-scores (Velocity, Points, Control, Speed; all P < 0.001). The degree of improvement on the Total scores ranged from 1100 to 7300 points (42% and 5500% improvement).
During the first 4 sessions (in phase 1), the slopes of the learning curves were similar for the fixed priority and variable priority groups, but the scores for the groups trained on these 2 versions diverged thereafter. As predicted from previous work, the variable priority training led to faster learning than did fixed priority training (Fabiani et al. 1989; Bherer et al. 2008). The group by training level interaction for the Total score was significant when examining all 20 sessions (F19,646 = 2.37, P < 0.001, Fig. 2B) and was marginally significant when splitting the data into 4 phases (F3,102 = 2.16, P < 0.09). By the end of training, the Total scores of participants trained in the variable priority protocol were 29% higher than those of the subjects trained with the fixed priority strategy. This difference in the Total scores for the 2 groups was mainly due to differential improvement by the variable training group on the Points and Velocity sub-scores. Improvements in performance were not correlated with initial performance for the Total score (r = 0.02), Points sub-score (r = 0.05), or the Velocity sub-score (r = −0.19), but they were correlated with initial performance for the Speed (r = −0.25) and the Control (r = −0.68) sub-scores. Variability in performance improvements for all sub-scores was statistically equivalent between the fixed priority and variable priority training groups as measured by Levene's test for homogeneity of variance with the one exception that the improvement in the Total score was more variable for variable priority training than for fixed priority training (P < 0.05). This result confirms that our data were appropriate for interrogation within the general linear model.
Predicting Initial Behavioral Performance Based on Brain Volume Measurements
The relationship between striatal volumes and improvements in game performance was not accounted for by individual differences in initial game performance. Out of 30 correlations between the 5 performance measures and the right and left hemisphere volumes of each brain region, including the caudate nucleus, nucleus accumbens, putamen, and hippocampus, only 2 were marginally significant: the Points sub-score was correlated with the left caudate nucleus volume (r = 0.33, P < 0.04) and marginally with the left nucleus accumbens volume (r = 0.30, P < 0.06). All other correlations between initial performance and volume were not significant (all P > 0.05). These correlations can be considered a liberal estimate of the relationship between initial performance and brain volume, as all were conducted without correction for multiple comparisons.
Hippocampal Volume Does Not Predict Performance Improvement across Training
Our main predictions were specific to the volume of the dorsal and ventral regions of the striatum. As a test of whether the correlations that we found were generalized or not, we examined whether the hippocampal volumes were also predictors of learning on the Space Fortress task. We found that hippocampal volumes were not predictive of performance, or improvements in performance (left: F2,33 = 1.40; not significant [n.s.]; right: F2,33 = 1.41; n.s.), for either the fixed priority or the variable priority versions (Fig. 3D). Left and right hippocampal volumes accounted for less than 2% of the variance in performance improvements for both training groups (all effects P > 0.05). By contrast, striatal volumes predicted changes in performance with training and did so in patterns specific both for the different striatal regions analyzed and for the type of training protocol experienced by the subjects.
Figure 3.

Scatterplots comparing the improvement in Total score in relation to the mean volume of the 4 different regions in each subject. Values for individuals trained in the variable priority (VP) version of the Space Fortress game are shown by black circles, and the solid line represents the linear fit of these data for the VP group. Values for the fixed priority (FP) group subjects are shown by open circles, and the dashed line represents the linear fit of these FP data. Correlations are shown separately for the volumes of the putamen (A), caudate nucleus (B), nucleus accumbens (C), and hippocampus (D). The only significant correlations between volumes and Total scores were for the VP group for the caudate nucleus. Data for the putamen reached P < 0.10.
Dorsal Striatal Volumes Predict Performance Improvements across Training
We first analyzed performance across the entire 20-h training period, collapsed across both training groups, to determine whether striatal volumes were predictive of performance improvements on the Space Fortress game. We used multiple regression analyses between change in performance (session 20 – session 1) and the volume of each region of the striatum (Table 1 and Fig. 3), while including initial performance as covariates in the model. Table 1 summarizes the effect sizes (beta values) and their significance levels for the 4 sub-scores.
Table 1.
Beta values and the significance levels from the multiple regression analyses between each region and the improvement in performance for each sub-score and the Total scores of the Space Fortress game players
| Total | Points | Velocity | Control | Speed | |
| L. Caudate | 0.37; P < 0.03 | 0.39; P < 0.02 | 0.32; P < 0.03 | 0.07; n.s. | 0.30; P < 0.07 |
| R. Caudate | 0.36; P < 0.03 | 0.36; P < 0.04 | 0.27; P < 0.06 | 0.06; n.s. | 0.27; P < 0.10 |
| L. Putamen | 0.29; P < 0.07 | 0.33; P < 0.05 | 0.29; P < 0.05 | 0.03; n.s. | 0.07; n.s. |
| R. Putamen | 0.29; P < 0.08 | 0.23; n.s. | 0.25; P < 0.10 | 0.08; n.s. | 0.05; n.s. |
| L. nucleus accumbens | 0.04; n.s. | −0.02; n.s. | 0.14; n.s. | −0.01; n.s. | −0.16; n.s. |
| R. nucleus accumbens | 0.12; n.s. | 0.09; n.s. | 0.21; n.s | −0.15; n.s. | −0.02; n.s. |
Note: All significance levels of P < 0.10 are given, and the significant relationships (P < 0.05) are in italics. These results suggest a dominant effect of the caudate nucleus volumes in relation to performance skill acquisition for the Total, Points, and Velocity scores. The putamen was at least marginally related to the Total, Points, and Velocity scores. However, the nucleus accumbens volumes were unrelated to improvement for the entire training period. L, left; R, right.
To begin, the overall ANOVAs were significant for the left (F2,33 = 4.00; P < 0.03) and right (F2,33 = 3.94; P < 0.03) caudate nucleus and marginally significant for the left (F2,33 = 3.00; P < 0.06) and right (F2,33 = 2.88; P < 0.07) putamen but failed to reach significance for the left (F2,33 = 1.23; P = 0.30) and right (F2,33 = 1.46; P = 0.24) nucleus accumbens. Given our predictions for the role of the dorsal striatum in learning, we determined the direction of these effects by exploring the beta values resulting from the multiple regression models (described below and in Table 1).
The volumes of the dorsal striatal regions were positively correlated with training-induced performance improvements for the Total scores (accounting for 9% of variance in learning), the Points scores (accounting for 10% of variance in learning), and the Velocity scores (accounting for 11% of variance in learning). By contrast, the volumes for the nucleus accumbens were not correlated with the task acquisition for either the Total score or any of the sub-scores (all accounting for less than 1% of the variance in learning rates). Furthermore, when all dorsal striatal regions were included in the multiple regression model, 23% of the variance in learning amounts across training was accounted for. It should be noted that these values represent statistical values uncorrected for multiple comparisons and therefore may be a liberal estimate of these associations.
To test whether these effects were influenced by gender-related differences in brain volume and learning, we re-ran the regression analyses while including gender as an additional covariate. All results described above remained unchanged after gender was included in the model. These results suggest that larger preexisting volumes of the caudate nucleus and putamen predicted faster overall rates of video game skill acquisition.
Dorsal Striatal Volumes Predict Learning Performance with Variable Priority Training but Not Fixed Priority Training Protocols
The dorsal striatal volumes were predictive of performance only for subjects trained on the variable priority version of the Space Fortress game, despite the fact that large performance gains were achieved by both groups of subjects (Figs 3 and 4). There were significant positive training group × volume interactions for the Points, Velocity, and Speed sub-scores. For the Points sub-score, the training group × volume interactions were significant for the left caudate nucleus (β = 0.39, P < 0.02), the right caudate nucleus (β = 0.40, P < 0.02), and the left putamen (β = 0.41, P < 0.01). For the Velocity sub-score, there were also significant training group × volume interactions for both sides of the caudate nucleus (left: β = 0.29, P < 0.05; right: β = 0.30, P < 0.04) and for the left putamen (β = 0.32, P < 0.03). As for the Points sub-scores, there was only a trend for a relation between performance and right putamenal volumes. For the Speed sub-score, there was a significant training group × volume interaction for the right caudate nucleus (β = 0.32, P < 0.04). There was a trend toward significance for the left caudate nucleus (β = 0.31, P < 0.06), but interactions for the putamen were not significant (left: β = 0.13, P > 0.05; right: β = 0.06, P > 0.05).
Figure 4.
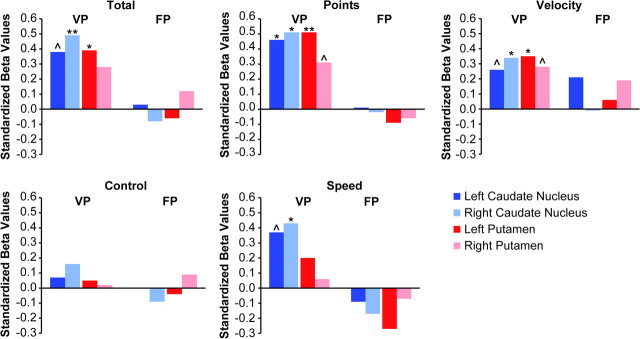
Relationship between each measure of performance on the Space Fortress game and dorsal striatum volumes, grouped by training method (VP, variable priority training; FP, fixed priority training). This data analysis demonstrates that the relationship between volume and performance improvement was driven by the VP method and was nonexistent for the individuals trained by the fixed priority method. (^ = P < 0.10; * = P < 0.05; ** = P < 0.001).
All of these interactions resulted from positive associations between preexisting striatal volumes and performance improvements for those subjects trained with variable priority methods. We found no consistent associations between striatal volume and performance improvements for the fixed priority training group (Fig. 3) and no significant interactions with gender (all P > 0.10). None of the regressions (main effects and interactions) between performance improvements and ventral striatal volumes were significant. Thus, the predictive value of the striatal volumes on learning was selective both for striatal region and for training strategy.
Volumes of the Nucleus Accumbens Predict Performance Improvement during Early Phases of Learning
Despite the lack of significant correlations between the volumes of the nucleus accumbens region and overall performance on the video game, the ventral striatal volumes were positively correlated with improvements in performance early during task acquisition. When the training periods were broken up into 4 separate phases, consisting of 5 sessions in each phase, we found that improvements in performance within the first 5 sessions (phase 1) for the Total score were related to both left (β = 0.37, P < 0.03) and right (β = 0.33, P < 0.05) nucleus accumbens volume (Fig. 5). This relationship with Total score appeared to be driven by a relationship with the Velocity sub-score. Improvements in Velocity scores for the first phase of learning were related to the volumes of the left (β = 0.41, P < 0.01) and the right (β = 0.46, P < 0.004) nucleus accumbens. There was no relationship between nucleus accumbens volumes and performance improvements in the later training sessions (phases 2–4) or for other measures of Space Fortress performance (all P > 0.10). Furthermore, all interactions between training group and volume also failed to reach significance levels of P < 0.05, suggesting that this correlation between the ventral striatal volumes and the initial learning phase held for both the fixed priority and variable priority training methods. Similarly, when gender was added to the model, the results remained unchanged. The volumes of the ventral striatum thus were selectively predictive of early acquisition, independent of gender and independent of the training method experienced by the subjects.
Figure 5.

Changes in correlation coefficients as a function of training phase for the Total performance scores on the variable priority version of the Space Fortress game in relation to the volumes of the 3 striatal regions analyzed. Volumes are represented as the mean of values for each region. The pretraining volumes of the caudate nucleus and putamen were significantly correlated with performance improvement across all phases of training for the variable priority group, whereas the nucleus accumbens volumes were significantly correlated with performance only during the early part of task training (phase 1).
We performed a similar analysis for the dorsal striatal structures to determine whether volumes were more related to the learning slopes of the early or later phases of task acquisition. In a multiple regression analysis, we found that dorsal striatal volume was predictive of performance improvements during both early and later learning phases for subjects trained with the variable priority strategy (Fig. 5). For example, the volumes of the left and right caudate nucleus were predictive of learning rates in phase 1 (left: β = 0.42, P < 0.01; right: β = 0.31, P < 0.05) as well as in phase 4 (left: β = 0.46, P < 0.01; right: β = 0.36, P < 0.03). The volumes for the putamen showed a similar trend as those of the caudate nuclei for phases 1–4 for the left and right hemispheres (all P < 0.05).
Discussion
Our findings demonstrate that the preexisting volumes of the striatum, but not the preexisting hippocampal volumes, predicted the improvement in performance of healthy young subjects learning a complex video game. The predictive power of the striatal measurements was dependent both on the striatal region and on the training strategy. The correlations between learning and dorsal striatal volume were specific to variable priority training methods; they were not significant for fixed priority training methods. For the variable priority game version, larger dorsal striatal volumes were associated with higher overall performance scores and some performance sub-scores following training.
There were no significant correlations between these performance scores and ventral striatal volumes. The ventral striatal volumes, however, were predictive of learning during the initial stages. When we analyzed performance improvement for the first and later phases of training, ventral striatal volumes were predictive of learning during initial phases of learning but not during later phases, whereas dorsal striatal volumes were predictive of both early and late learning. These findings point to the striatum as a key element of neural circuits underpinning video game learning and suggest that both the nature of the demands of the procedural learning and the dynamics of the learning trajectories are reflected in the dominant striatal circuits involved.
The selectivity of the ventral striatum for predicting performance during the early stages of learning highlights the importance of limbic circuits related to this striatal region to initial learning and the reinforcement-related processes involved in early task acquisition and suggests that these do not have as dominant an effect later in the learning process. This result is consistent with studies in rodents and humans demonstrating a critical role of the nucleus accumbens in early stages of learning and the dorsal striatum in both earlier and later stages of learning (Hernandez et al. 2002; Doyon et al. 2003; Barnes et al. 2005; Atallah et al. 2007; Graybiel 2008; Balleine et al. 2009; Belin et al. 2009). The striatum is interconnected with the neocortex by trans-thalamic circuits that can be broadly divided into sensorimotor, associative, and limbic zones, with the dorsal striatum contributing to motor and associative processing including cognitive flexibility, and the ventral striatum contributing especially to motivational and affective processing (Packard and Knowlton 2002; Postuma and Dagher 2006; Belin and Everitt 2008; Di Martino et al. 2008; Draganski et al. 2008; Graybiel 2008; Takahashi et al. 2008). The early periods of acquisition of the Space Fortress game may have invoked heightened activity of limbic inputs related to reward and motivation, processes regulated by the ventral striatum. By contrast, during later learning phases, the limbic inputs may have been diminished while cognitive switching and procedural processes, regulated by the dorsal striatum, were heightened. The fact that we could detect correlations between dorsal striatal volumes and performance for all phases of learning suggests that the dorsal striatum operates with the continued input throughout acquisition from circuits associated with cognitive flexibility and procedural learning.
The Space Fortress task has substantial motor learning demands related to controlling the ship's position and direction in a frictionless environment. In addition, variable priority training promotes cognitive flexibility by shifting the emphasis of training from one task component to another during the training sessions (Gopher et al. 1994; Kramer et al. 1995, 1999). Our results argue that, in the human, preexisting variations in striatal volume can affect the rate of learning in a complex task that involves the coordination and integration of many cognitive, motor, and perceptual parameters and rules, at least when conditions of learning capitalize on flexible learning strategies. The fact that the striatal volumes were predictive of learning in the variable priority version of the task but not in the fixed priority version is consistent with this interpretation. In contrast to the variable priority training group, the fixed priority training group, due to the generalized nature of their training approach, was not encouraged to prioritize different aspects of the task flexibly, but rather, was trained to use a “flat” priority approach. This strategy reduced the need for flexible task prioritization and may have led to basal ganglia-based circuitry being less relevant for this group during learning. The basal ganglia may have a central function in flexible priority allocation of task goals.
The Space Fortress task requires a number of cognitive and motor processes for successful performance. It is, therefore, likely that a network of brain regions comprising visual, parietal, and frontal areas contribute to successful task performance and learning. This makes our results all the more compelling—that is, the measurement of the basal ganglia accounted for 23% of the variance in learning, a value that is quite high given that many other brain regions are probably contributing to performance and learning. However, the relationship between basal ganglia volume and learning was only significant for those trained with a variable priority strategy. Given that the fixed priority group also learned the task, brain regions other than the basal ganglia must be involved in the acquisition of the task for this group. This result complements the argument that the association between regional brain volume and learning is dependent on the degree to which the brain region supports the learning strategies employed in the task.
Our results argue that the volume of basal ganglia is predictive of learning on the Space Fortress Task when subjects are exposed to a variable priority learning strategy. Unfortunately, the cellular and molecular factors that contribute to such volumetric assessments are unknown but could include trophic factor influences, greater dendritic or axonal arborizations, more synaptic connections, greater synaptic plasticity, more neurons or more active neurons, or greater vascularization (Schubert et al. 2009). Nearly three-quarters (72%) of the participants in our sample were females with relatively little experience with video game playing. In our regression analyses, we included gender as a covariate to remove variance in learning or brain volume associated with gender, and gender did not moderate any of the effects reported in this study. Prior studies have found that women experience faster learning rates in action video games compared with those of men, resulting in a reduction of gender differences in spatial attention (Feng et al. 2007). Given that we failed to find any moderating effects of gender in this study, we can be reasonably certain that our results do not reflect differential learning rates and brain volume between the genders. Nonetheless, the skewed gender distribution with only 36 participants may limit the generalization of our findings and may have also affected the power to detect gender differences if they exist. Such questions would be better addressed with larger sample sizes and an equal proportion of males and females.
Our finding that the association between striatal volume and learning occurred only for participants receiving variable priority training is also important practically, given that this training method emphasizes cognitive flexibility and leads to superior learning more generally (Kramer et al. 1995, 1999; Bherer et al. 2005, 2006, 2008). If variable priority training is more effective than fixed training for learning and capitalizes more on basal ganglia-based circuits as a consequence, then this type of training could prove more useful for enhancing cognitive function in a number of applied settings. This interpretation of our findings requires further testing to determine whether the association between learning and basal ganglia volumes holds for other complex tasks and whether such training will generalize to the performance of other experimental and real-world cognitive tasks.
Funding
Office of Naval Research (N00014-07-1-0903 to A.F.K. and A.M.G.).
Acknowledgments
We thank Nancy Dodge, Holly Tracy, Tracey Wszalek, Emily Wee, Harish Sharma, Brad Sutton, Zuha Warraich, Dawn Epstein, Sarah Dalton, Matt Windsor, Maulik Sheth, and Yasuo Kubota for their help in this project. Conflict of Interest: None declared.
References
- Atallah HE, Lopez-Paniagua D, Rudy JW, O'Reilly RC. Separate neural substrates for skill learning and performance in the ventral and dorsal striatum. Nat Neurosci. 2007;10:126–131. doi: 10.1038/nn1817. [DOI] [PubMed] [Google Scholar]
- Balleine BW, Liljeholm M, Ostlund SB. The integrative function of the basal ganglia in instrumental conditioning. Behav Brain Res. 2009;199:43–52. doi: 10.1016/j.bbr.2008.10.034. [DOI] [PubMed] [Google Scholar]
- Barnes TD, Kubota Y, Hu D, Jin DZ, Graybiel AM. Activity of striatal neurons reflects dynamic encoding and recoding of procedural memories. Nature. 2005;437:1158–1161. doi: 10.1038/nature04053. [DOI] [PubMed] [Google Scholar]
- Basak C, Boot WR, Voss MW, Kramer AF. Can training in a real-time strategy videogame attenuate cognitive decline in older adults? Psychol Aging. 2008;23:765–777. doi: 10.1037/a0013494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belin D, Everitt BJ. Cocaine seeking habits depend upon dopamine-dependent serial connectivity linking the ventral with the dorsal striatum. Neuron. 2008;57:432–441. doi: 10.1016/j.neuron.2007.12.019. [DOI] [PubMed] [Google Scholar]
- Belin D, Jonkman S, Dickinson A, Robbins TW, Everitt BJ. Parallel and interactive learning processes within the basal ganglia: relevance for the understanding of addiction. Behav Brain Res. 2009;199:89–102. doi: 10.1016/j.bbr.2008.09.027. [DOI] [PubMed] [Google Scholar]
- Bherer L, Kramer AF, Peterson M, Colcombe SJ, Erickson KI, Becic E. Transfer effects in task-set cost and dual-task cost after dual-task training in older and younger adults: further evidence for cognitive plasticity in attentional control in late adulthood. Exp Aging Res. 2008;334:188–219. doi: 10.1080/03610730802070068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bherer L, Kramer AF, Peterson MS, Colcombe S, Erickson K, Becic E. Training effects on dual-task performance: are there age-related differences in plasticity of attentional control? Psychol Aging. 2005;20:695–709. doi: 10.1037/0882-7974.20.4.695. [DOI] [PubMed] [Google Scholar]
- Bherer L, Kramer AF, Peterson MS, Colcombe S, Erickson K, Becic E. Testing the limits of cognitive plasticity in older adults: application to attentional control. Acta Psychol (Amst) 2006;123:261–278. doi: 10.1016/j.actpsy.2006.01.005. [DOI] [PubMed] [Google Scholar]
- Boot WR, Kramer AF, Simons DJ, Fabiani M, Gratton G. The effects of video game playing on attention, memory, and executive control. Acta Psychol (Amst) 2008;129:387–398. doi: 10.1016/j.actpsy.2008.09.005. [DOI] [PubMed] [Google Scholar]
- Cools R, Clark L, Robbins TW. Differential responses in human striatum and prefrontal cortex to changes in object and rule relevance. J Neurosci. 2004;24:1129–1135. doi: 10.1523/JNEUROSCI.4312-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dahlin E, Neely AS, Larsson A, Backman L, Nyberg L. Transfer of learning after updating training mediated by the striatum. Science. 2008;320:1510–1512. doi: 10.1126/science.1155466. [DOI] [PubMed] [Google Scholar]
- De Martino B, Kumaran D, Holt B, Dolan RJ. The neurobiology of reference-dependent value computation. J Neurosci. 2009;29:3833–3842. doi: 10.1523/JNEUROSCI.4832-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Martino A, Scheres A, Margulies DS, Kelly AM, Uddin LQ, Shehzad Z, Biswal B, Walters JR, Castellanos FX, Milham MP. Functional connectivity of human striatum: a resting state FMRI study. Cereb Cortex. 2008;18:2735–2747. doi: 10.1093/cercor/bhn041. [DOI] [PubMed] [Google Scholar]
- Donchin E, Fabiani M, Sanders A. editors. 1989. The learning strategies program: an examination of the strategies in skill acquisition [Special Issue] Acta Psychol (Amst).71:1–311. [PubMed] [Google Scholar]
- Doyon J, Penhune V, Ungerleider LG. Distinct contribution of the cortico-striatal and cortico-cerebellar systems to motor skill learning. Neuropsychologia. 2003;41:252–262. doi: 10.1016/s0028-3932(02)00158-6. [DOI] [PubMed] [Google Scholar]
- Draganski B, Kherif F, Kloppel S, Cook PA, Alexander DC, Parker GJ, Deichmann R, Ashburner J, Frackowiak RS. Evidence for segregated and integrative connectivity patterns in the human basal ganglia. J Neurosci. 2008;28:7143–7152. doi: 10.1523/JNEUROSCI.1486-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erickson KI, Prakash RS, Voss MW, Chaddock L, Hu L, Morris KS, White SM, Wojcicki TR, McAuley E, Kramer AF. Aerobic fitness is associated with hippocampal volume in elderly humans. Hippocampus. 2009;19:1030–1039. doi: 10.1002/hipo.20547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fabiani M, Buckley J, Gratton G, Coles MH, Donchin E, Logie R. The training of complex task performance. Acta Psychol (Amst) 1989;71:259–299. [Google Scholar]
- Feng J, Spence I, Pratt J. Playing an action video game reduces gender differences in spatial cognition. Psychol Sci. 2007;18:850–855. doi: 10.1111/j.1467-9280.2007.01990.x. [DOI] [PubMed] [Google Scholar]
- Frederickson JR, White BY. An approach to training based on principled task decomposition. Acta Psychol (Amst) 1989;71:89–146. [Google Scholar]
- Gopher D, Weil M, Bareket T. Transfer of skill from a computer game trainer to flight. Hum Factors. 1994;36:387–405. [Google Scholar]
- Graybiel AM. The basal ganglia: learning new tricks and loving it. Curr Opin Neurobiol. 2005;15:638–644. doi: 10.1016/j.conb.2005.10.006. [DOI] [PubMed] [Google Scholar]
- Graybiel AM. Habits, rituals and the evaluative brain. Annu Rev Neurosci. 2008;31:359–387. doi: 10.1146/annurev.neuro.29.051605.112851. [DOI] [PubMed] [Google Scholar]
- Green CS, Bavelier D. Action video game modifies visual selective attention. Nature. 2003;423:534–537. doi: 10.1038/nature01647. [DOI] [PubMed] [Google Scholar]
- Green CS, Bavelier D. Effect of action video games on the spatial distribution of visuospatial attention. J Exp Psychol Hum Percept Perform. 2006;32:1465–1478. doi: 10.1037/0096-1523.32.6.1465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green CS, Bavelier D. Action-video-game experience alters the spatial resolution of vision. Psychol Sci. 2007;18:88–94. doi: 10.1111/j.1467-9280.2007.01853.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernandez PJ, Sadeghian K, Kelley AE. Early consolidation of instrumental learning requires protein synthesis in the nucleus accumbens. Nat Neurosci. 2002;5:1327–1331. doi: 10.1038/nn973. [DOI] [PubMed] [Google Scholar]
- Kennedy KM, Erickson KI, Rodrigue KM, Voss MW, Colcombe SJ, Kramer AF, Acker JD, Raz N. Age-related differences in regional brain volumes: a comparison of optimized voxel-based morphometry to manual volumetry. Neurobiol Aging. 2009;30:1657–1676. doi: 10.1016/j.neurobiolaging.2007.12.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimchi EY, Laubach M. Dynamic encoding of action selection by the medial striatum. J Neurosci. 2009;29:3148–3159. doi: 10.1523/JNEUROSCI.5206-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koepp MJ, Gunn RN, Lawrence AD, Cunningham VJ, Dagher A, Jones T, Brooks DJ, Bench CJ, Grasby PM. Evidence for striatal dopamine release during a video game. Nature. 1998;393:266–268. doi: 10.1038/30498. [DOI] [PubMed] [Google Scholar]
- Kramer AF, Larish JF, Strayer DL. Training for attentional control in dual-task settings: a comparison of young and old adults. J Exp Psychol Appl. 1995;1:50–76. [Google Scholar]
- Kramer AF, Larish JF, Strayer DL. Training for executive control: task coordination strategies and aging. In: Gopher D, Koriat A, editors. Attention and performance XVII. Cambridge (MA): MIT Press; 1999. [Google Scholar]
- Meiran N, Friedman G, Yehene E. Parkinson's disease is associated with goal setting deficits during task switching. Brain Cogn. 2004;54:260–262. doi: 10.1016/j.bandc.2004.02.043. [DOI] [PubMed] [Google Scholar]
- O'Doherty J, Dayan P, Schultz J, Deichmann R, Friston K, Dolan RJ. Dissociable roles of ventral and dorsal striatum in instrumental conditioning. Science. 2004;304:452–454. doi: 10.1126/science.1094285. [DOI] [PubMed] [Google Scholar]
- Packard MG, Knowlton BJ. Learning and memory functions of the basal ganglia. Annu Rev Neurosci. 2002;25:563–593. doi: 10.1146/annurev.neuro.25.112701.142937. [DOI] [PubMed] [Google Scholar]
- Patenaude B, Smith SM, Kennedy D, Jenkinson M. Bayesian shape and appearance models. Oxford: FMRIB Centre, University of Oxford; 2007. Technical Report TR07BP1. [Google Scholar]
- Patenaude B, Smith SM, Kennedy D, Jenkinson M. FIRST-FMRIB's integrated registration and segmentation tool. 2007. Human Brain Mapping Conference. Organization for Human Brain Mapping Conference Abstracts. Chicago, IL. [Google Scholar]
- Postuma RB, Dagher A. Basal ganglia functional connectivity based on a meta-analysis of 126 positron emission tomography and functional magnetic resonance imaging publications. Cereb Cortex. 2006;16:1508–1521. doi: 10.1093/cercor/bhj088. [DOI] [PubMed] [Google Scholar]
- Ragozzino ME, Jih J, Tzavos A. Involvement of the dorsomedial striatum in behavioral flexibility: role of muscarinic cholinergic receptors. Brain Res. 2002;953:205–214. doi: 10.1016/s0006-8993(02)03287-0. [DOI] [PubMed] [Google Scholar]
- Raz N, Lindenberger U, Rodrigue KM, Kennedy KM, Head D, Williamson A, Dahle C, Gerstorf D, Acker JD. Regional brain changes in aging healthy adults: general trends, individual differences and modifiers. Cereb Cortex. 2005;15:1676–1689. doi: 10.1093/cercor/bhi044. [DOI] [PubMed] [Google Scholar]
- Schubert MI, Porkess MV, Dashdorj N, Fone KC, Auer DP. Effects of social isolation rearing on the limbic brain: a combined behavioral and magnetic resonance imaging volumetry study in rats. Neuroscience. 2009;159:21–30. doi: 10.1016/j.neuroscience.2008.12.019. [DOI] [PubMed] [Google Scholar]
- Smith SM, Jenkinson M, Woolrich MW, Beckmann CF, Behrens TE, Johansen-Berg H, Bannister PR, De Luca M, Drobnjak I, Flitney DE, et al. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage. 2004;23(Suppl 1):S208–S219. doi: 10.1016/j.neuroimage.2004.07.051. [DOI] [PubMed] [Google Scholar]
- Squire LR, Stark CE, Clark RE. The medial temporal lobe. Annu Rev Neurosci. 2004;27:279–306. doi: 10.1146/annurev.neuro.27.070203.144130. [DOI] [PubMed] [Google Scholar]
- Takahashi Y, Schoenbaum G, Niv Y. Silencing the critics: understanding the effects of cocaine sensitization on dorsolateral and ventral striatum in the context of an actor/critic model. Front Neurosci. 2008;2:86–99. doi: 10.3389/neuro.01.014.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin HH, Knowlton BJ. The role of the basal ganglia in habit formation. Nat Rev Neurosci. 2006;7:464–476. doi: 10.1038/nrn1919. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Brady M, Smith S. Segmentation of brain MR images through a hidden Markov random field model and the expectation-maximization algorithm. IEEE Trans Med Imaging. 2001;20:45–57. doi: 10.1109/42.906424. [DOI] [PubMed] [Google Scholar]