

Abstract
Reduced neuronal activation to repetitive stimulation is a common feature of information processing in nervous systems. Such stimulus-specific adaptation (SSA) occurs in many systems, but the underlying neural mechanisms are not well understood. The Neoconocephalus (Orthoptera, Tettigoniidae) TN-1 auditory neuron exhibits an SSA-like process, characterized by reliably detecting deviant pulses after response cessation to common standard pulses. Therefore, TN-1 provides a model system to study the cellular mechanisms underlying SSA with an identified neuron. Here we test the hypothesis that dendritic mechanisms underlie TN-1 response cessation to fast-pulse rate repeated signals. Electrically stimulating TN-1 with either high-rate or continuous-current pulses resulted in a decreased ability in TN-1 to generate action potentials but failed to elicit cessation of spiking activity as observed with acoustic stimulation. BAPTA injection into TN-1 delayed the onset of response cessation to fast-pulse rate acoustic stimuli in TN-1 but did not eliminate it. These results indicate that calcium-mediated processes contribute to the fast cessation of spiking activity in TN-1 but are insufficient to cause spike cessation on its own. Replacing normal saline with low-Na+ saline (replacing sodium chloride with either lithium chloride or choline chloride) eliminated response cessation, and TN-1 no longer responded selectively to the deviant pulses. Sodium-mediated potassium channels are the most likely candidates underlying sodium-mediated response suppression in TN-1, triggered by Na+ influx in dendritic regions activated by acoustic stimuli. On the basis of these results, we present a model for a cellular mechanism for SSA in a single auditory neuron.
Keywords: auditory neuron, hearing, insect
reduced neuronal activation to repetitive stimulation is a common feature of information processing in nervous systems (Grill-Spector et al. 2006). At the level of individual neurons, this is reflected as stimulus-specific adaption (SSA), where a decrease in responses to the repeated stimulus is not generalized to other rare (deviant) stimuli (Gutfreund and Knudsen 2006; Nelken and Ulanovsky 2007; Ulanovsky et al. 2003). It occurs at various levels in the mammalian auditory pathway (Antunes et al. 2010; Malmierca et al. 2009; Ulanovsky et al. 2003). SSA also occurs in other animal groups and sensory modalities (e.g., Gutfreund 2012; Netser et al. 2011; Schul et al. 2012). Despite its universal occurrence and extensive studies, we are only beginning to understand the neural mechanisms underlying SSA, especially the effects of repetitive stimulation on individual neurons (Antunes and Malmierca 2011; Pérez-González et al. 2012).
Recent findings of an SSA-like process in an insect system (Schul et al. 2012; Schul and Sheridan 2006) provide a model system to study the cellular mechanisms underlying SSA. In Neoconocephalus katydids (Orthoptera, Tettigoniidae), the auditory system functions mainly in two contexts: intraspecific communication and predator avoidance (Greenfield 1990). The auditory system of these katydids processes acoustic signals containing amplitude modulation rates several orders of magnitude faster than in mammals or birds. The communication signals typically contain pulse rates between 100 and 200 pulses per second (pps), while relevant predator signals (search-phase echolocation cries of bats) have 5–10 pps pulse rates. Katydid ears (located in the forelegs) contain ∼35 receptor neurons with characteristic frequencies ranging from below 6 kHz to above 60 kHz (Höbel and Schul 2007; Römer 1983). The afferents project into the prothoracic ganglion, where they form excitatory synapses with local and ascending interneurons (Römer et al. 1988). Three primary auditory interneurons ascend on each side, transmitting behaviorally relevant information to the brain (Triblehorn and Schul 2009).
One of these primary interneurons (TN-1) is sensitive to a broad range of carrier frequencies (∼6–60 kHz) with thresholds < 40 dB SPL. TN-1 responds reliably to slow pulse rates (<10 pps) with one to three action potentials (APs) per pulse, while fast pulse rates (>50 pps) do not elicit ongoing responses, as AP activity ceases within a few hundred milliseconds after the onset of the fast pulse rate. When a slow pulse rate (7 pps) was embedded within a fast pulse rate (140 pps), TN-1 reliably encoded the slow pulse rate if the carrier frequencies of the two rates were sufficiently different (Schul et al. 2012). TN-1 responses to the slow pulse rate (i.e., “deviant” pulses) occurred when the deviant pulses had either a lower or a higher carrier frequency than the fast pulse rate. TN-1 continued to respond reliably to deviant pulses with amplitudes as much as 25 dB lower than the fast pulse rate. Schul et al. (2012) argued that this strong adaptation to fast pulse rates while maintaining sensitivity to the deviant pulses of the slow rate exhibited by TN-1 responses parallels SSA described in subcortical stations in the auditory pathway of mammals (e.g., Antunes et al. 2010; Malmierca et al. 2009). Therefore, TN-1 provides an excellent model to study SSA-like processes at the level of a single, identified neuron.
Extensive testing suggested that TN-1 adaptation to fast pulse rates did not result from 1) adaptation of its afferents, 2) synaptic depression of the afferent-TN-1 excitatory synapses, or 3) known inhibitory inputs to TN-1 (Schul et al. 2012). Thus processes intrinsic to TN-1 are likely candidates to generate the SSA-like responses of TN-1 (Schul et al. 2012).
In the present study, we tested the hypothesis that cellular mechanisms intrinsic to TN-1 underlie the cessation of spiking responses to fast pulse rates and repeated signals in this neuron. First, using intracellular current injections, we measured the contribution of axonal, AP-driven processes to response cessation in TN-1. Second, using pharmacological manipulations, we tested the importance of Ca2+- and Na+-activated processes intrinsic to TN-1 for response cessation. We present evidence for at least two processes intrinsic to TN-1, one Na+ activated and another Ca2+ activated. On the basis of these results, we provide a model for how these processes explain the SSA-like responses of TN-1 that could also apply to SSA in vertebrate systems.
METHODS
Animals.
The animals used in these experiments were Neoconocephalus triops (Linnaeus 1758) collected in Puerto Rico and their offspring (F1 and F2) reared in the laboratory under summer conditions (Beckers and Schul 2008). Experiments were conducted on insects at least 2 wk after collection or adult molt. We could not detect differences between the sexes. We used this species instead of N. retusus (the species used by Schul et al. 2012; Schul and Sheridan 2006) because we were able to raise it in culture, while N. retusus is available only for ∼2 mo during its field season. We confirmed in preliminary experiments that the response properties of TN-1 were similar in the stimulus situation used here and by Schul et al. (2012).
Preparation.
Animals were anesthetized with CO2 and mounted ventral side up on a freestanding metal holder with a wax-resin mixture or yellow sticky wax (Kerr's). The prothoracic legs (containing the tympanal organs) were fixed on a wire holder perpendicular to the body axis, while the meso- and metathoracic legs were fixed along the body wall. The prothoracic ganglion was exposed by removing a small piece of cuticle and covered with saline (modified after Fielden 1960). A wax well was built around the opening to hold the saline. The ganglion was stabilized with a NiCr spoon. The ascending connectives were cut close to the subesophageal ganglion to minimize descending activity. Removal of descending activity did not alter the TN-1 response properties (Schul and Sheridan 2006).
Electrophysiology.
The soma and dendritic field of TN-1 are located in the prothoracic ganglion. The ascending axon collateral projects into the brain, while a second axon collateral descends toward the abdomen. TN-1 receives direct excitatory input from most auditory receptor cells from the ear ipsilateral to the TN-1 dendrite. Further synaptic input stems from local interneurons. Detailed descriptions of TN-1 morphology and physiology are given elsewhere (Römer et al. 1988; Stumpner 1999; Triblehorn and Schul 2009). The large size of the TN-1 axon and dendrites facilitates intracellular recordings from the dendrite as well as reliable monitoring of spiking activity from the ascending axon.
Intracellular recording and stimulation were made from or close to the TN-1 main dendrite within the prothoracic ganglion with thick-walled borosilicate glass microelectrodes (WPI). Electrodes were filled with 3 M KCl and had impedances between 20 and 50 MΩ. Signals were low-pass filtered (5 kHz) and amplified (Warner Instruments IE-210). Current injections from the intracellular amplifier were gated by a Master-8 pulse stimulator (A.M.P.I.). Prior to data collection in any experiment involving intracellular penetration of TN-1 (i.e., electrical stimulation and calcium chelator injection experiments), two acoustic stimuli were presented to ensure that TN-1 activity was not altered because of electrode penetration: 1) a 1-s pulse train [120 pps containing 3.5 ms of white noise (WN) pulses at 70 dB SPL] to verify that fast-pulse rate stimuli still caused a cessation in TN-1 responses and 2) two standard-deviant stimuli (see below) to verify that TN-1 still responded to deviant pulses (stimulus described in Quantifying detection of deviant pulses by TN-1).
We monitored TN-1 spiking activity extracellularly in the cervical connective, using either a suction electrode (recording from the cut end of the ascending connective) or a tungsten hook electrode (70- to 100-μm diameter). The hook electrode and cervical connective were drawn in a polypropylene pipette tip containing a petroleum jelly-mineral oil mixture that insulated the recording from the surrounding saline. Extracellular recordings were amplified with a custom-built amplifier and band-pass filtered (100–4,000 Hz, Krohn-Hite 3384). Because of its large axonal diameter, TN-1 spiking activity (for the morphology of TN-1 see Stumper 1999; Triblehorn and Schul 2009) is readily identifiable in extracellular recordings from the cervical connective based on its large spike amplitude and its response to acoustic stimulation (Faure and Hoy 2000; McKay 1969; Schul 1997). In addition to monitoring TN-1 spiking activity, the extracellular recording was used to identify intracellular recordings of TN-1 based on exact correspondence between spiking activity in both recordings. Both intracellular and extracellular recordings were digitized (InstruNet 100B, Omega Engineering, New York, NY) and stored with SuperScope II (GW Instruments). SuperScope II was also used for data analysis.
Acoustic stimulation.
Experiments were conducted in an anechoic Faraday cage (1.2 m × 1.2 m × 0.7 m) at 24–26°C. We generated synthetic signals with a 16-bit D/A converter system (National Instruments PCI-6251) at a sampling rate of 250 kHz, using custom-designed software developed in LabVIEW (National Instruments). Stimulus amplitudes were manipulated by a computer-controlled attenuator. Stimuli were delivered via a loudspeaker (Technic 10TH400C) mounted 50 cm from the preparation, perpendicular to the insect's body axis and ipsilateral to the recording side. Signal amplitudes were calibrated at the position of the insect with a 1/4-in. condenser microphone (G.R.A.S. 40BF, Vedbaek, Denmark) and a B&K 2231 sound level meter (Brüel and Kjær, Naerum, Denmark). All sound pressure levels are given as dB peak sound pressure level (SPL; re 20 μPa). At the recording site slight echoic influences were unavoidable, but these influences did not alter the envelope of the signals by more than ±1 dB.
Quantifying TN-1 responses to pulse rates.
We presented pulse trains (3-s duration) consisting of 3.5-ms pulses (0.5 ms rise/fall time) repeated at a slow (7 pps) and a fast (120 or 140 pps) pulse rate. Carrier signals of the pulse trains were 15 kHz, 40 kHz, or WN. All pulse rates for one carrier signal were presented before testing another, and pulse rates were presented in random order. We kept a minimum of 30 s between stimulus presentations.
To quantify the TN-1 responses to the stimuli, we 1) counted the total number of TN-1 spikes occurring during the 3-s stimulus presentation and 2) created peristimulus time histograms (PSTHs), using 50-ms time bins. For pharmacological treatments, we subtracted the bins of the baseline PSTH from the treatment PSTH to generate “difference PSTHs.” Positive values of the difference PSTH indicate an increase and negative values a decrease in responses during the treatment.
Quantifying detection of deviant pulses by TN-1.
To quantify TN-1's ability to respond to deviant pulses, we used the standard/deviant stimulus used by Schul and Sheridan (2006) and Schul et al. (2012) (Fig. 1, A and B). Both standard and deviant pulses had 3.5-ms duration, including 0.5-ms rise and fall time, and carrier frequencies of either 15 or 40 kHz. Standard pulses were presented with a 3.5-ms interpulse interval, equivalent to a pulse rate of ∼140 pps; deviant pulses were inserted in the silent interval every 20 standard pulses (i.e., the pulse rate of the deviants was 7 pps). The standard was presented for a total of 8 s: the first 3 s alone (i.e., without deviant pulses inserted) and the following 5 s with deviant pulses inserted into the standard signal (35 deviant pulses total).
Fig. 1.

A and B: schematic view of the standard (std)/deviant (dev) stimulus at millisecond (A) and second (B) timescales. The standard stimulus consisted of pulses (3.5-ms duration) repeated after a silent interval of 3.5 ms. After 3 s, a deviant pulse (3.5-ms duration) was inserted after every 20th standard pulse into the silent interval. Note that standard and deviant pulses were played through the same loudspeaker. C: example of TN-1 response in Neoconocephalus triops to the standard/deviant stimulus demonstrating stimulus-specific adaptation (SSA). TN-1 responds to the onset of the standard stimulus (15-kHz carrier frequency) with a short burst of action potentials (APs) and subsequently only responds to deviant pulses (40-kHz carrier frequency) presented together with the standard pulses. The TN-1 response to the standard/deviant stimulus in N. triops is similar to that observed in N. retusus (Schul et al. 2012; Schul and Sheridan 2006).
Schul et al. (2012) demonstrated that TN-1 exhibits SSA-like responses to a variety of standard and deviant carrier frequency combinations when the carrier frequencies are sufficiently different, regardless of whether the deviant carrier frequency is higher or lower than the standard (Schul et al. 2012). In this study, we chose to test two combinations (all pulse amplitudes = 70 dB SPL): 1) standard pulses = 15 kHz, deviant pulses = 40 kHz and 2) the inverse situation with standard pulses = 40 kHz, deviant pulses = 15 kHz.
We quantified the spiking responses to the latter 5 s of each stimulus (i.e., the part with deviant pulses inserted; Fig. 1). Spikes occurring within 35 ms of the onset of deviant pulses (i.e., within the first third of the deviant pulse period) were classified as responses to the deviant (= deviant time window). Spikes occurring during the 105 ms between the deviant time windows were considered responses to the standard stimulus. We present the proportion of the 35 deviants and standards that elicited at least one AP as the TN-1 response probability for deviant and standard stimuli, respectively. We chose response probability rather than spikes per pulse as a measure because we wanted to test the reliability of the encoding of the deviant pulses. This response measure is consistent with previous studies of change detection in TN-1 (Schul et al. 2012; Schul and Sheridan 2006).
Electrical stimulation.
We used electrical intracellular stimulation to test the contribution of axonal, AP-driven processes to SSA in TN-1. We stimulated TN-1 with 10-ms depolarizing pulses presented at 7, 15, 30, 50, and 75 pps. This pulse duration evoked reliable responses of 2 APs/pulse. A shorter pulse duration (i.e., 3.5 ms as in acoustic experiments) was not used since the pulses did not reliably elicit spiking responses at the slow pulse rate (7 pps). For each individual, the pulse stimulator was set to the minimum voltage required to reliably elicit two spikes per pulse for all seven pulses presented at 7 pps for 1 s (3–4 V). We tested two different series during electrical stimulation: 1) Equal Duration (all pulse rates were presented for 1 s, i.e., the number of pulses differed among stimuli) and 2) Equal Pulse Number (all pulse rates were presented for 100 pulses, i.e., the duration of the pulse train differed).
A stimulus with 7 pps was tested at the beginning and end of each series as well as between each of the other pulse rates (15, 30, 50, and 75 pps). The latter pulse rates were presented in random order. We kept a 30-s interval between the stimuli. The stimulus voltage was kept constant for all pulse rates tested within each series. We included data only from experiments in which a complete series was tested. If both stimulus series were tested in an individual TN-1, the threshold voltage was rechecked before beginning the second stimulus series. Data were discarded if 1) >1 of the pulses presented at 7 pps for 1 s failed to elicit two spikes per pulse (for Equal Duration) or 2) >5 of the 100 pulses presented at 7 pps failed to elicit two spikes per pulse (for Equal Pulse Number).
In some preparations, TN-1 responses to a 500-ms current pulse and a 500-ms acoustic pulse (WN at 70 dB SPL) were tested.
Pharmacological manipulations.
To test the importance of dendritic Ca2+- and Na+-activated processes on TN-1 response cessation, we conducted pharmacological manipulations of TN-1 and its environment by either removing free intracellular Ca2+ through the injection of a calcium chelator or replacing Na+ in the extracellular medium.
The role of intrinsic Ca2+-activated events was tested by injecting the Ca2+ chelator 1,2-bis(2-aminophenoxy)ethane-N,N,N′,N′-tetraacetic acid (BAPTA; Sigma) into the main dendrite. Fifty millimolar BAPTA was iontophoretically injected for 25–30 min with a 3-nA hyperpolarizing current. Responses were measured with extracellular recordings before (baseline) and after BAPTA injection.
We tested the importance of Na+-activated processes (such as Na+-activated K+ currents) with low-Na+ saline by replacing NaCl with either LiCl or choline chloride. This method has been commonly used (e.g., Hess et al. 2007; Le Guennec and Noble 1994; Wallén et al. 2007) to eliminate Na+-activated cellular processes, as there are no established Na+ chelators. Na+ could not be completely eliminated since the saline also contains sodium bicarbonate (Fielden 1960), which cannot be replaced (Sanchez-Vives et al. 2000). The chemical replacing NaCl was added in an amount to maintain the same total osmolarity of the normal saline. Replacing NaCl with either choline chloride or LiCl removes sodium-activated events, in particular the opening of Na+-activated K+ channels, while supporting APs since Li+ and choline pass through the voltage-gated Na+ channels (Hess et al. 2007; Sanchez-Vives et al. 2000; Wallén et al. 2007).
TN-1 responses to acoustic pulse trains were tested before (baseline), during (low Na+), and after (wash) application of the pharmacological manipulation. We replaced the hemolymph/saline surrounding the prothoracic ganglion with the low-Na+ saline until we registered a change in TN-1 responses to fast pulse rates. After the changes stabilized, we tested the physiology during low-Na+ conditions. We then reapplied the normal saline to wash out the low-Na+ saline until TN-1 responses to fast pulse rates returned to baseline activity. We included only data from experiments in which experimental effects washed out.
TN-1 spiking activity was only monitored extracellularly (described above) during experiments involving low Na+ and after BAPTA injection.
RESULTS
In response to the standard-deviant stimulus situations, TN-1 responded with a burst of spikes at the onset of the standard pulses, but spiking responses ceased within 100–200 ms (Fig. 1C). Typically, 80–90% of deviant pulses elicited spiking responses (1–3 APs), while spikes occurred in <10% of the interspersed time windows with standard pulses. These response properties of the N. triops TN-1 were similar to those of TN-1 in N. retusus (Schul et al. 2012; Schul and Sheridan 2006). The following experiments elucidate the neuronal mechanisms underlying the cessation of spiking responses to fast pulse rates (i.e., the standard in Fig. 1).
Electrical stimulation.
Intracellular current injection elicits APs without activation of postsynaptic channels by afferent input, which allows us to separate the effects of processes triggered by the opening of postsynaptic channels from those triggered by AP activity.
During electrical stimulation with low pulse rates (7, 15, and 30 pps), >95% of the current pulses elicited two spikes per pulse (Fig. 2A). This percentage progressively decreased as pulse rate increased (∼75% for 50 pps and ∼30% for 75 pps). Both stimulus series (see methods) produced similar results. The results of the current injection followed the same trend as seen with acoustic stimulation (from Schul and Sheridan 2006). However, the reduction in TN-1 spiking responses as pulse rate increased was much greater for acoustic stimulation. For acoustic stimulation, <10% of the pulses elicited any spikes at 50 and 75 pps (Fig. 2A) as opposed to electrical stimulation, where the majority of pulses presented at these rates elicited at least one spike (Fig. 2B).
Fig. 2.

TN-1 responses during electrical stimulation. A: TN-1 response to 10-ms electrical or 3.5-ms acoustic stimulus pulses presented at different rates. Electrical stimulation involved 2 different series of stimuli (see methods), and TN-1 responses were quantified as % of pulses eliciting 2 spikes per pulse (Equal Duration n = 9; Equal Pulse Number n = 6; means ± SE). Acoustic stimulation involved 15- and 40-kHz pulses, and TN-1 responses were measured as the average spike per pulse (data from Schul and Sheridan 2006). B: histograms displaying the average number of TN-1 spikes elicited by each pulse during the Equal Pulse Number series for all 5 pulse rates (7, 15, 30, 50, and 75 pps). The dashed line indicates an average of 2 spikes per pulse; values below the line represent a decrease in TN-1 responses, while values above the line indicate enhancement. C: extracellular recordings of TN-1 responses to a 500-ms white noise (WN) burst (top) or a 500-ms electrical stimulation (bottom). D: instantaneous frequency of TN-1 responses during the 500-ms constant stimulation result shown in C beginning with the first spike.
During the Equal Pulse Number stimulation, responses progressively declined over the course of the stimulus train for 50 and 75 pps (Fig. 2B). Current pulses at the beginning of the stimulation reliably elicited two spikes, but the number of spikes decreased as the pulse train continued. The decline was faster and greater for the 75 pps than the 50 pps stimulation and accounts for the overall decrease in response percentages for 50 and 75 pps. This result was consistent across the animals tested.
During stimulation with a 500-ms continuous current injection (Fig. 2C), the instantaneous firing rate dropped from >300 Hz to 100 Hz within 100–150 ms and remained at this level for the remainder of the stimulus (Fig. 2D). Acoustically stimulating TN-1 with a 500-ms WN burst initially produced a high firing rate, but spiking responses ceased within 50–100 ms.
Electrical stimulation failed to elicit the TN-1 spiking cessation observed during acoustic stimulation with fast pulse rates or continuous sound bursts. However, electrical stimulation activated processes that reduced spiking activity during fast pulse rates and rapidly limited sustained spike rates to ∼100 Hz. The pharmacological experiments (below) tested the importance of Ca2+- and Na+-activated processes in both the dendrite and the axon.
Effects of pharmacological manipulations on TN-1 response cessation to fast pulse rates.
To determine the role of intracellular Ca2+ in the cessation of spiking activity to fast pulse rates, we compared TN-1 responses to acoustic stimulation before and after the injection of the Ca2+ chelator BAPTA, which buffers intracellular calcium. Prior to BAPTA injection, responses to the three carrier signals (WN, 15 kHz, 40 kHz) were similar. TN-1 responded to each pulse during the entire 7 pps train, while spiking responses ceased during the 120 pps train (Fig. 3A, left). BAPTA injection had little, if any, systematic effect on responses to the slow pulse rate for all carrier signals. For 120 pps, the effect of BAPTA injection was similar for WN and 40-kHz stimuli: TN-1 spikes persisted for a longer duration, but spiking ceased before the stimulus ended (Fig. 3, A and B). Responses increased by up to 2 spikes/bin at the beginning of the stimulus, while responses increased by <0.5 spikes/bin toward the end of the stimulus (Fig. 3B). Thus Ca2+-related processes appeared to contribute to the suppression of TN-1 spikes but were not solely responsible for sustained spike suppression, since TN-1 spiking activity still ceased after BAPTA injection. For 15-kHz pulses, responses to 120 pps were relatively unaffected by BAPTA injection, i.e., they tended to cease with a similar time course as prior to BAPTA injection (Fig. 3B).
Fig. 3.

TN-1 responses before and after BAPTA injection. A: extracellular recordings of TN-1 responses to 3.5-ms WN acoustic pulses presented at 7 and 120 pps before (Baseline, left) and after (BAPTA, right) BAPTA injection (3-s stimulus duration). B: histograms of the change of TN-1 spiking after BAPTA injection (mean, n = 6): acoustic stimulation with WN pulses (3.5-ms duration; top), 15 kHz (middle), and 40 kHz (bottom) repeated at 7 (left) and 120 (right) pps. Bins were 50 ms wide. Positive values indicate an increase in TN-1 responses after BAPTA injection.
We used low-Na+ saline to test the importance of Na+-activated processes for the TN-1 spike cessation. TN-1 responses to fast pulse rates were dramatically altered by replacement of NaCl with LiCl (Fig. 4) or choline chloride (Fig. 5). Under low-Na+ conditions, TN-1 responses continued throughout the duration of the fast-pulse rate stimulus and the spike cessation observed before the treatment disappeared (note that both Na+ replacements, Li+ and choline, pass through Na+ channels and support AP generation). Responses increased by 2–4 APs/bin (50-ms bin width) for WN and 40-kHz stimuli and by 1 or 2 APs/bin for 15-kHz stimuli (Fig. 4B, Fig. 5B). Under low-Na+ conditions, TN-1 produced more spikes at the beginning of the stimulus and stabilized after 1–1.5 s.
Fig. 4.

TN-1 responses before and after application of low-Na+ saline replacing NaCl with LiCl. A: extracellular recordings of TN-1 responses to 3.5-ms WN acoustic pulses presented at 7 and 120 pps before (Baseline, left) and after (Lithium, right) application of low-Na+ saline (3-s stimulus duration). B: histograms of the change of TN-1 spiking after introduction of low-Na+ saline (mean, n = 3): acoustic stimulation with WN pulses (3.5-ms duration) presented at 7 and 120 pps (top), 15 kHz at 140 pps (bottom left), and 40 kHz at 140 pps (bottom right). Bins were 50 ms wide. Positive values indicate an increase in TN-1 responses after introduction of low-Na+ saline.
Fig. 5.

TN-1 responses before and after application of low-Na+ saline replacing NaCl with choline chloride. A: extracellular recordings of TN-1 responses to 3.5-ms WN acoustic pulses presented at 7 and 120 pps (3-s stimulus duration) before (Baseline, left) and after (Choline, right) application of low-Na+ saline. B: histograms of the change of TN-1 spiking after introduction of low-Na+ saline (mean, n = 3): acoustic stimulation with WN pulses (3.5-ms duration) presented at 7 and 120 pps (top), 15 kHz at 140 pps (bottom left), and 40 kHz at 140 pps (bottom right). Bins were 50 ms wide. Positive values indicate an increase in TN-1 responses after introduction of low-Na+ saline.
Replacement of Na+ also affected responses to the slow pulse rate (7 pps). Responses to each pulse increased by 1 or 2 APs, as evidenced by the regularly spaced positive bars in Fig. 4B and Fig. 5B. This pulse rate was tested with WN pulses only.
Figure 6 compares the average increase in APs during the 3-s stimulation among the three treatments (BAPTA and two low-Na+ conditions), pulse rates (7, 120/140 pps) and pulse types (WN, 15 kHz, 40 kHz). All three treatments had only weak effects on responses to the slow pulse rate. BAPTA moderately increased TN-1 responses to fast pulse rates with 40-kHz and WN carrier signals and had little, if any, effect on responses with the 15-kHz carrier signal. However, during low-Na+ conditions, responses to fast pulse rates increased substantially. For WN and 40-kHz pulses, TN-1 activity increased by >100 spikes during the 3-s stimulus. This increase was weaker for the 15-kHz pulses (∼40 spikes).
Fig. 6.

Comparison of the effects of pharmacological manipulations on TN-1 responses. Left: BAPTA injection. Center: LiCl replacement of NaCl. Right: choline chloride replacement of NaCl. Bar heights indicate the mean (±SE) increase in number of APs elicited during the 3-s stimulation.
Effects of low-Na+ condition on detection of deviant pulses by TN-1.
Since the low-Na+ treatments greatly increased the number of TN-1 spikes to fast pulse rates, we tested how they would affect TN-1's ability to detect deviant stimuli. We used two versions of the 8-s standard/deviant stimulus (Fig. 1): 1) standard pulses = 15 kHz, deviant pulses = 40-kHz carrier frequencies and 2) carrier frequencies reversed: standard = 40 kHz, deviant = 15 kHz.
During baseline conditions (i.e., with normal Na+ concentration), TN-1 responses to standard pulses ceased shortly after stimulus onset for both stimuli (Fig. 7) and few responses to standard pulses occurred during the last 5 s of the stimulus that included both standard and deviant pulses (see methods). Deviant pulses consistently elicited TN-1 responses throughout the stimulation (Fig. 7). This basic response pattern was similar for both versions of the stimulus.
Fig. 7.
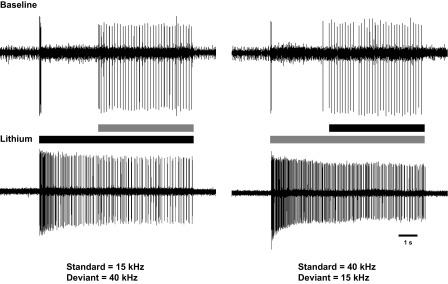
Extracellular recordings of TN-1 responses during the 8 s of the standard/deviant stimulus before (Baseline) and after replacement of normal saline with low-Na+ saline (LiCl replaced NaCl in example) with the carrier frequencies of the standard/deviant pulses being 15/40 kHz (left) and 40/15 kHz (right).
After application of the low-Na+ saline, the TN-1 response pattern changed substantially. Standard pulses continued to evoke TN-1 responses throughout the stimulus duration, including between the deviant pulses (Fig. 7, bottom). TN-1 still reliably responded to the deviant pulses, and responses even increased by 1–3 APs/pulse in some preparations. The responses to the deviant pulses were now, however, masked by the continued spiking to the standard pulses.
Because replacing NaCl with either LiCl or choline chloride had similar effects (Figs. 4–6), we pooled the data of both treatments for quantitative analysis. During baseline conditions, response probabilities to the standard pulses were <0.1 for both stimulus versions while the deviant pulses were much more likely to elicit spikes (Fig. 8). During low-Na+ conditions, response probabilities to the standard pulses increased to 0.57 (15-kHz standard/40-kHz deviant) or 0.65 (40/15 kHz). Response probabilities to the deviant pulses remained high during the low-Na+ conditions and even increased relative to the baseline condition.
Fig. 8.

Response probability (mean ± SE, n = 6) of TN-1 to the standard and deviant pulses during baseline (BL) and low-Na+ conditions. The probability (see methods) of eliciting at least 1 AP during the 35 presentations is given for standard and deviant pulses. TN-1 responses were tested with the carrier frequencies of the standard/deviant pulses being 15/40 kHz (left) and 40/15 kHz (right). Data from both low-Na+ treatments (LiCl or choline chloride replacing NaCl) were pooled.
DISCUSSION
The neural detection of rare or deviant signals requires that the responses to common or repeated signals are suppressed while the neuron or network remains sensitive to novel signals that differ sufficiently from the background. This stimulus-specific adaptation (SSA) to the repeated signal can be interpreted as sensory memory about previous stimulation (Ulanovsky et al. 2003). There is currently very little known about the neural mechanisms underlying SSA in vertebrates (recently reviewed in Ayala and Malmierca 2013). Current hypotheses focus primarily on network mechanisms (i.e., inhibitory GABAergic inputs) or the concept of “memory” neurons (Jääskeläinen et al. 2007) but not on potential cellular mechanisms.
Here we studied the cellular mechanisms underlying the response cessation to fast repeated pulses in TN-1 and specifically tested the role of processes intrinsic to TN-1. Our main findings were that 1) electrical stimulation resulted in a reduced ability to generate APs (for pulse trains) and a decrease in firing rate but not in the complete response cessation observed during acoustic stimulation; 2) BAPTA injection delayed, but did not eliminate, response cessation; and 3) low-Na+ conditions eliminated response cessation and TN-1 no longer responded selectively to the deviant pulses.
Calcium-activated processes underlie spike rate adaptation.
Electrically stimulating TN-1 intracellularly should trigger spike-dependent currents (e.g., voltage-gated Ca2+ currents) without activation of postsynaptic ion channels that depend on afferent transmitter release. At fast pulse rates (50 and 75 pps), the percentage of stimulus pulses that elicited 2 APs/pulse declined. Thus the electrical stimulation resulted in spike rate adaptation (Fig. 2B).
BAPTA is a Ca2+ chelator that reduces intracellular Ca2+ elevation when injected into neurons (e.g., Ogawa et al. 2001) and therefore should largely eliminate Ca2+-activated currents (such as Ca2+-activated K+ channels). Our BAPTA experiments support the idea that Ca2+-triggered processes underlie spike rate adaptation in TN-1, since the duration of the initial spike burst to acoustic fast pulse rates increased after BAPTA treatment (Fig. 3). Furthermore, responses to the acoustic slow pulse rate (7 pps) increased by ∼1 AP/pulse. Both effects can be explained by reduced spike rate adaptation in the absence or reduction of Ca2+-activated currents due to Ca2+ buffering by BAPTA.
The spike rate adaptation observed in the electrical stimulation and BAPTA experiments probably contributes to the fast onset of the cessation of spiking responses during acoustic stimulation with fast pulse rates but appears insufficient to cause spike cessation on its own. The latter statement is further supported by the results from the low-Na+ conditions (Figs. 4 and 5), where spike cessation did not occur during the fast pulse rate even though Ca2+-activated processes remained unaffected. Whether additional Ca2+-mediated processes exist in TN-1, e.g., through voltage-gated Ca2+ channels activated by synaptic activity, needs to be addressed in further studies (e.g., calcium imaging experiments or the use of channel blockers).
Sodium-activated processes underlie spike cessation.
While Ca2+-activated mechanisms contribute to the fast onset of response cessation, the failure to completely eliminate spike cessation after BAPTA injection suggests the involvement of non-Ca2+ mechanisms. A likely mediator is Na+, which might contribute to spike cessation through Na+-activated K+ channels (Bhattacharjee and Kaczmarek 2005; Sah 1996; Sah and Faber 2002). Application of low-Na+ saline should largely eliminate Na+-activated events.
Under low-Na+ conditions, response cessation to fast pulse rates was largely eliminated and spikes occurred throughout the stimulus duration (Figs. 4, 5). The decrease of TN-1's firing frequency over the duration of the stimulus was likely caused by the Ca2+-mediated spike frequency adaptation (see above). The elimination of response cessation during low-Na+ conditions resulted in standard pulses eliciting additional TN-1 responses while responses to the deviant persisted or increased (Figs. 7, 8). The increased response to standard pulses under low-Na+ conditions effectively masked the deviant pulses. In terms of auditory function, this masking would strongly interfere with detecting changes in the auditory scene.
The response properties of TN-1 to slow pulse rates remained largely unchanged during low-Na+ conditions, indicating that neither replacement method substantially interfered with afferent APs or synaptic transmission to TN-1. The choline treatment could potentially increase excitatory synaptic transmission from afferents to TN-1 since acetylcholine is the neurotransmitter used by most insect sensory neurons (Leitch and Pitman 2009; Pitman 1985) and presynaptic uptake of the added choline could lead to higher transmitter synthesis and release. However, the results from the low-Na+ saline using Li+ replacement paralleled that involving choline replacement, which indicates that the absence of Na+, not the replacement ions themselves, caused the treatment effects.
Na+-activated K+ channels (Bhattacharjee and Kaczmarek 2005; Dryer 1994) would be the most likely candidates underlying response cessation in TN-1, triggered by Na+ influx through ligand-gated Na+ or mixed-cation channels opened during synaptic transmission. We propose that opening of Na+-activated K+ channels within the TN-1 dendrites would act as a “shunt,” attenuating the excitatory postsynaptic potentials (EPSPs) reaching the spike initiating zone of TN-1. Furthermore, only fast pulse rates would allow enough Na+ to open sufficient numbers of the K+ channels to provide this “current shunt.” Intracellular recordings from the main TN-1 dendrite (which register PSPs and APs) revealed that EPSPs occur for the duration of stimulation with fast pulse rates (Triblehorn and Schul 2009). The cessation of TN-1 spiking activity coincides with a decrease in EPSP amplitude. This agrees with the proposed dendritic mechanisms (EPSP amplitude decreasing because of current shunting), as there is no evidence for decreased activity of afferents or the synaptic transmission to TN-1 (Schul et al. 2012). The continued spiking of TN-1 during both low-Na+ conditions highlights the ability of the afferents to transmit fast pulse rates to TN-1. Although synaptic depression cannot be completely ruled out, it is unlikely that low-Na+ conditions would eliminate synaptic depression. In contrast, low-Na+ conditions created by replacing NaCl with either LiCl or choline chloride can prevent the activation of Na+-activated K+ channels in the dendritic mechanism that we propose (e.g., Hess et al. 2007; Le Guennec and Noble 1994; Wallén et al. 2007).
A cellular model for SSA-like response properties generated by a single neuron.
Our results provide evidence for a dendritic origin of the response cessation to fast pulse rates in TN-1. However, the main characteristic of SSA is that the sensitivity to deviant signals is maintained while the responses to the standard are suppressed. Schul et al. (2012) demonstrated that TN-1 responses to deviants occurred when standard and deviant pulses had sufficiently different carrier frequencies, when the carrier frequency of the deviant was either lower or higher than the standard (e.g., Fig. 8). We propose here a model based on the morphological organization of the prothoracic auditory neuropile and TN-1 in tettigoniids. Projections of auditory receptor cells are tonotopically organized according to their spectral selectivity (Ebendt et al. 1994; Römer et al. 1988), strikingly resembling the tonotopic organization of vertebrate hearing systems (Feng and Ratnam 2000; Montealegre-Z et al. 2012). Projections of individual receptor cells overlap with only a limited portion of the TN-1 dendrite (Römer et al. 1988), and therefore signals with different carrier frequencies should stimulate different regions of the TN-1 dendritic field. Our model proposes that the Na+-based dendritic mechanism is activated only in the stimulated regions of the TN-1 dendrite and therefore so will the attenuation of the EPSPs to the spike initiating zone via the current shunt mediated by Na+-activated K+ channels. Stimulation of other dendritic branches (from afferents with different spectral sensitivity) would remain unaffected and could still elicit APs in TN-1. This mechanism would work for different carrier frequencies of standard and deviant, as long as they are sufficiently different, stimulating separate regions of the TN-1 dendritic field. With the tonotopic organization of the auditory neuropile, this dendritic, Na+-activated mechanism of spike suppression in TN-1 would thus provide a mechanism for generating SSA-like response properties in a single neuron.
Support for this model comes from Schul et al. (2012), where TN-1 responses to deviant pulses occurred for large spectral differences between standard and deviant, i.e., when they were transmitted by different groups of receptor cells and stimulated different regions of the TN-1 dendritic field. In contrast, when deviant and standard pulses were spectrally similar and thus stimulated the same TN-1 dendritic region, TN-1 did not respond to the deviant (Schul et al. 2012).
Whether Na+ (and possibly Ca2+) accumulation is spatially limited in the stimulated regions of the TN-1 dendritic field needs to be tested with ion imaging experiments. However, localized Ca2+ accumulation does occur in parts of the dendrite of another large insect auditory neuron depending on the carrier frequency of the stimulation (Baden and Hedwig 2006).
SSA in Neoconocephalus occurred at standard pulse rates (140 pps) that were an order of magnitude faster than SSA in vertebrates, while the deviant pulse rates (7 pps) were closer to the standard pulse rates typically used in vertebrate studies (Ulanovsky et al. 2003). Nevertheless, SSA in TN-1 strikingly resembles that occurring at lower levels of the mammal auditory system, such as the medial geniculate body (e.g., cf. our Fig. 1 and Antunes and Malmierca 2011, Fig. 2), the thalamus (Antunes et al. 2010), and the inferior colliculus (Malmierca et al. 2009). In mammalian auditory systems, SSA depends primarily on mechanisms acting on the neuron's inputs (Ulanovsky et al. 2003, 2004) and cortical control does not contribute significantly to SSA (Antunes and Malmierca 2011). The strong frequency dependence of SSA in mammals suggests that the tonotopic organization of the hearing system might play a role in SSA. Our findings indicate that intracellular processes are responsible for SSA in TN-1 and that the spatial change of the input pattern on the TN-1 dendritic field underlies the detection of change in the auditory scene. Similar mechanisms may be responsible for SSA at lower levels of the mammalian hearing system.
GRANTS
This work was supported by a grant from the National Science Foundation to J. Schul (IOS 0722109). J. D. Triblehorn was supported by a Ruth L. Kirschstein National Research Service Award (NRSA) of the National Institute on Deafness and other Communication Disorders (1 F32 DC-008925) and by grants from the National Center for Research Resources (5 P20 RR-016461) and the National Institute of General Medical Sciences (8 P20 GM-103499). The experiments followed current US laws.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
Author contributions: J.D.T. and J.S. conception and design of research; J.D.T. performed experiments; J.D.T. and J.S. analyzed data; J.D.T. and J.S. interpreted results of experiments; J.D.T. and J.S. prepared figures; J.D.T. and J.S. drafted manuscript; J.D.T. and J.S. edited and revised manuscript; J.D.T. and J.S. approved final version of manuscript.
ACKNOWLEDGMENTS
We thank the members of the Schul lab for support and helpful discussions. J. Prešern and S. L. Bush provided input to the manuscript. We also thank the anonymous reviewers for comments that improved the manuscript.
REFERENCES
- Antunes FM, Malmierca MS. Effect of auditory cortex deactivation on stimulus-specific adaptation in the medial geniculate body. J Neurosci 31: 17306–17316, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Antunes FM, Nelken I, Covey E, Malmierca MS. Stimulus-specific adaptation in the auditory thalamus of the anesthetized rat. PLoS One 5: e14071, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayala YA, Malmierca MS. Stimulus-specific adaptation and deviance detection in the inferior colliculus. Front Neural Circuits 6: 1–16, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baden T, Hedwig B. Neurite-specific Ca2+ dynamics underlying sound processing in an auditory interneurone. Dev Neurobiol 67: 68–80, 2006 [DOI] [PubMed] [Google Scholar]
- Beckers OM, Schul J. Developmental plasticity of mating calls enables acoustic communication in diverse environments. Proc Biol Sci 275: 1243–1248, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhattacharjee A, Kaczmarek LK. For K+ channels, Na+ is the new Ca2+. Trends Neurosci 28: 422–428, 2005 [DOI] [PubMed] [Google Scholar]
- Dryer SE. Na+-activated K+ channels: a new family of large-conductance ion channels. Trends Neurosci 17: 155–160, 1994 [DOI] [PubMed] [Google Scholar]
- Ebendt R, Friedel J, Kalmring K. Central projection of auditory receptors in the prothoracic ganglion of the bushcricket Psorodonotus illyricus (Tettigoniidae): computer-aided analysis of the end branch pattern. J Neurobiol 25: 35–49, 1994 [DOI] [PubMed] [Google Scholar]
- Faure PA, Hoy RR. Neuroethology of the T-cell. I. Tuning and responses to pure tones. J Exp Biol 203: 3225–3242, 2000 [DOI] [PubMed] [Google Scholar]
- Feng AS, Ratnam R. Neural basis of hearing in real-world situations. Annu Rev Psychol 51: 699–725, 2000 [DOI] [PubMed] [Google Scholar]
- Fielden A. Transmission through the last abdominal ganglion of the dragonfly nymph, Anax imperator. J Exp Biol 37: 832–844, 1960 [Google Scholar]
- Greenfield MD. Evolution of acoustic communication in the genus Neoconocephalus: discontinuous songs, synchrony, and interspecific interactions. In: The Tettigoniidae: Biology, Systematics, and Evolution, edited by Bailey WJ, Rentz DC. Heidelberg, Germany: Springer, 1990 [Google Scholar]
- Grill-Spector K, Henson R, Martin A. Repetition and the brain: neural models of stimulus-specific effects. Trends Cogn Sci 10: 14–23, 2006 [DOI] [PubMed] [Google Scholar]
- Gutfreund Y. Stimulus-specific adaptation, habituation and change detection in the gaze control system. Biol Cybern 106: 657–668, 2012 [DOI] [PubMed] [Google Scholar]
- Gutfreund Y, Knudsen EI. Adaptation in the auditory space map of the barn owl. J Neurophysiol 96: 813–825, 2006 [DOI] [PubMed] [Google Scholar]
- Hess D, Nanou E, El Manira A. Characterization of Na+-activated K+ currents in larval lamprey spinal cord neurons. J Neurophysiol 97: 3484–3493, 2007 [DOI] [PubMed] [Google Scholar]
- Höbel G, Schul J. Listening for males and bats: spectral processing in the hearing organ of Neoconocephalus bivocatus (Orthoptera: Tettigoniidae). J Comp Physiol A Neuroethol Sens Neural Behav Physiol 193: 917–925, 2007 [DOI] [PubMed] [Google Scholar]
- Jääskeläinen IP, Ahveninen J, Belliveau JW, Raij T, Sams M. Short-term plasticity in auditory cognition. Trends Neurosci 30: 653–661, 2007 [DOI] [PubMed] [Google Scholar]
- Le Guennec J, Noble D. Effects of rapid changes of external Na+ concentration at different moments during the action potential in guinea-pig myocytes. J Physiol 478: 493–504, 1994 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leitch B, Pitman RM. Neuromodulation of mechanosensory input to the insect CNS. In: Mechanosensitivity of the Nervous System Part II. edited by Kamkin A, Kiseleva I. Amsterdam: Springer, 2009, p. 145–170 [Google Scholar]
- McKay JM. The auditory system of Homorocoryphus (Tettigoniidae, Orthoptera). J Exp Biol 51: 787–802, 1969 [Google Scholar]
- Malmierca MS, Cristaudo S, Perez-Gonzalez D, Covey E. Stimulus-specific adaptation in the inferior colliculus of the anesthetized rat. J Neurosci 29: 5483–5493, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montealegre ZF, Jonsson T, Robson-Brown KA, Postles M, Robert D. Convergent evolution between insect and mammalian audition. Science 338: 968–971, 2012 [DOI] [PubMed] [Google Scholar]
- Nelken I, Ulanovsky N. Mismatch negativity and stimulus-specific adaptation in animal models. J Psychophysiol 21: 214–223, 2007 [Google Scholar]
- Netser S, Zahar Y, Gutfreund Y. Stimulus-specific adaptation: can it be a neural correlate of behavioral habituation? J Neurosci 31: 17811–17820, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ogawa H, Baba Y, Oka K. Dendritic calcium accumulation regulates wind sensitivity via short-term depression at cercal sensory-to-giant interneuron synapses in the cricket. J Neurobiol 46: 301–313, 2001 [PubMed] [Google Scholar]
- Pérez-González D, Hernández O, Covey E, Malmierca MS. GABAA-mediated inhibition modulates stimulus-specific adaptation in the inferior colliculus. PLoS One 7: e34297, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pitman RM. Nervous system. In: Comprehensive Insect Physiology, Biochemistry and Pharmacology, edited by Kerkut GA, Gilbert LI. Oxford, UK: Pergamon, 1985, vol. 11, p. 5–54 [Google Scholar]
- Römer H. Tonotopic organization of the auditory neuropile in the bushcricket Tettigonia viridissima. Science 306: 60–62, 1983 [Google Scholar]
- Römer H, Marquardt V, Hardt M. Organization of a sensory neuropile in the auditory pathway of two groups of Orthoptera. J Comp Neurol 275: 201–215, 1988 [DOI] [PubMed] [Google Scholar]
- Sah P. Ca2+-activated K+ currents in neurons: types, physiological roles and modulation. Trends Neurosci 19: 150–154, 1996 [DOI] [PubMed] [Google Scholar]
- Sah P, Faber ES. Channels underlying neuronal calcium-activated potassium currents. Prog Neurobiol 66: 345–353, 2002 [DOI] [PubMed] [Google Scholar]
- Sanchez-Vives MV, Nowak LG, McCormick DA. Cellular mechanisms of long-lasting adaptation in visual cortical neurons in vitro. J Neurosci 20: 4286–4299, 2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schul J. Neuronal basis of phonotactic behaviour in Tettigonia viridissima: processing of behaviourally relevant signals by auditory afferents and thoracic interneurons. J Comp Physiol A 180: 573–583, 1997 [Google Scholar]
- Schul J, Mayo AM, Triblehorn JD. Auditory change detection by a single neuron in an insect. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 198: 695–704, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schul J, Sheridan RA. Auditory stream segregation in an insect. Neuroscience 138: 1–4, 2006 [DOI] [PubMed] [Google Scholar]
- Stumpner A. Comparison of morphology and physiology of two plurisegmental sound-activated interneurones in a bushcricket. J Comp Physiol A 185: 199–205, 1999 [Google Scholar]
- Triblehorn JD, Schul J. Sensory-encoding differences contribute to species-specific call recognition mechanisms. J Neurophysiol 102: 1348–1357, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulanovsky N, Las L, Farkas D, Nelken I. Multiple time scales of adaptation in auditory cortex neurons. J Neurosci 24: 10440–10453, 2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulanovsky N, Las L, Nelken I. Processing of low-probability sounds by cortical neurons. Nat Neurosci 6: 391–398, 2003 [DOI] [PubMed] [Google Scholar]
- Wallén P, Robertson B, Cangiano L, Low P, Bhattacharjee A, Kaczmarek LK, Grillner S. Sodium-dependent potassium channels of a Slack-like subtype contribute to the slow afterhyperpolarization in lamprey spinal neurons. J Physiol 585: 75–90, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]