

Abstract
Odor signals are transmitted to the olfactory bulb by olfactory nerve (ON) synapses onto mitral/tufted cells (MTCs) and external tufted cells (ETCs); ETCs provide additional feed-forward excitation to MTCs. Both are strongly regulated by intraglomerular inhibition that can last up to 1 s and, when blocked, dramatically increases ON-evoked MC spiking. Intraglomerular inhibition thus limits the magnitude and duration of MC spike responses to sensory input. In vivo, sensory input is repetitive, dictated by sniffing rates from 1 to 8 Hz, potentially summing intraglomerular inhibition. To investigate this, we recorded MTC responses to 1- to 8-Hz ON stimulation in slices. Inhibitory postsynaptic current area (charge) following each ON stimulation was unchanged from 1 to 5 Hz and modestly paired-pulse attenuated at 8 Hz, suggesting there is no summation and only limited decrement at the highest input frequencies. Next, we investigated frequency independence of intraglomerular inhibition on MC spiking. MCs respond to single ON shocks with an initial spike burst followed by reduced spiking decaying to baseline. Upon repetitive ON stimulation peak spiking is identical across input frequencies but the ratio of peak-to-minimum rate before the stimulus (max-min) diminishes from 30:1 at 1 Hz to 15:1 at 8 Hz. When intraglomerular inhibition is selectively blocked, peak spike rate is unchanged but trough spiking increases markedly decreasing max-min firing ratios from 30:1 at 1 Hz to 2:1 at 8 Hz. Together, these results suggest intraglomerular inhibition is relatively frequency independent and can “sharpen” MC responses to input across the range of frequencies. This suggests that glomerular circuits can maintain “contrast” in MC encoding during sniff-sampled inputs.
Keywords: interneurons, microcircuits, inhibition, olfactory, GABA
olfactory sensory input is rhythmic, occurring in patterns dictated by sniffing. Mouse sniffing rates range from 1 to 2 Hz in quiescent respiration to 8 to 10 Hz during investigatory sniffing. As a result, synaptic input from the olfactory nerve (ON) to the olfactory bulb is highly patterned. The transformation of this patterned sensory input into output signals by mitral/tufted cells (MTCs) involves a complex interplay of excitatory and inhibitory circuits.
There are three major inhibitory circuits within the bulb. 1) MCs release glutamate from their lateral dendrites onto granule cells (GCs), which release GABA onto the same or other MCs to generate recurrent and lateral inhibition, respectively. This comprises the MC-GC-MC circuit (Schoppa et al. 1998; Isaacson and Strowbridge 1998; Isaacson and Murphy 2001), which has long been considered the primary inhibitory regulation of MCs (Phillips et al. 1963; Shepherd 1963). 2) Periglomerular (PG) inhibitory neurons in the glomeruli are at least as numerous as GCs (Parrish-Aungst et al. 2007). PGCs are “uniglomerular”; they form intraglomerular inhibitory circuits that provide rapid, strong inhibition to both MTCs and external tufted cells (ETCs; Shao et al. 2009). Intraglomerular inhibition potently regulates the duration of spike output from MCs (Shao et al. 2012). 3) The third major inhibitory circuit is interglomerular, comprised of short axon cells projecting between glomeruli. Activation of the interglomerular circuit inhibits surrounding glomeruli (Aungst et al. 2003; Shirley et al. 2010; Liu et al. 2013). How these three circuits operate to transform temporally pattered input to bulb output is unclear.
In vivo, the magnitude and temporal structure of MTC spike responses to odors are remarkably well maintained across sniffing frequencies (Carey and Wachowiak 2011). This suggests that MTC spiking is rapidly “reset” between sniff-patterned inputs to normalize responses. We investigated the role of intraglomerular inhibition in temporal response patterning by recording MC spiking evoked by 1 to 8 Hz ON stimulation in slices before and after selectively blocking intraglomerular inhibition. Maximum-to-minimum firing ratios of MCs were almost identical across this range of frequencies. Selective block of intraglomerular inhibition significantly reduced MC max-min firing ratios. This suggests that intraglomerular inhibition enhances MC response contrast to odors during sniffing.
METHODS
All experiments were performed on olfactory bulb slices, obtained from male 8- to 10-wk-old C57/BL6 mice (Charles River). Animals were anesthetized with saturated vapor isoflurane and decapitated, and the olfactory bulbs surgically were removed and immediately placed in 4°C oxygenated sucrose-artificial cerebrospinal fluid (sucrose-aCSF) containing the following (in mM): 26 NaHCO3, 1 NaH2PO4, 3 KCl, 5 MgSO4, 0.5 CaCl2, 10 glucose, and 248 sucrose, equilibrated with 95% O2-5% CO2, pH 7.38. Horizontal slices (400-μm thick) were cut with a Leica VT1200 vibratome. Slices were incubated in oxygenated aCSF (in mM; 124 NaCl, 26 NaHCO3, 3 KCl, 1.25 NaH2PO4, 2 MgSO4, 2 CaCl2, and 10 glucose equilibrated with 95% O2-5% CO2, pH 7.38) at 30°C for 20–30 min and then at room temperature (22°C) in aCSF for at least 1 h before use. For recording, individual slices were transferred to a recording chamber and perfused with aCSF (as above) at a rate of 2.5 ml/min maintained at a temperature 30°C (Bipolar Temperature Controller, Norfolk, VA). Mitral cells were observed with a ×40 water-immersion objective using an Olympus BX51W upright microscope equipped for near-infrared differential interference contrast optics (Olympus Optical). All experimental procedures were carried out in accordance with protocols submitted to and approved by the University of Maryland Institutional Animal Care and Use Committee. All drugs used in this study were purchased from Sigma (St. Louis, MO).
Whole cell (current and voltage) patch-clamp recordings were performed as previously described (Shao et al. 2009, 2012). Briefly, recording pipettes were made from thick-wall borosilicate glass with filament (I.D.: 0.75 mm; Sutter Instruments, Novato, CA) pulled on a P-97 Flaming-Brown puller (Sutter). For current clamp, the internal solution contained the following (in mM): 120 K-gluconate, 20 KCl, 10 HEPES, 2 MgCl2, 2 Mg2ATP, 0.2 Na3GTP, 0.1 BAPTA, and 0.02% lucifer yellow, pH 7.3 adjusted with KOH and for voltage clamp contained the following (in mM): 120 CsMeSO4, 10 QX-314, 10 HEPES, 1 MgCl2, 2.5 Mg2ATP, 0.2 Na3GTP, 0.1 BAPTA, 10 phosphocreatin, and 0.02% lucifer yellow, pH 7.3 adjusted with CsOH. Osmolarity for both solutions were in the range 287–295 mOsm. For voltage clamp, MCs were held at −10 mV with a CsMeSO4 intracellular solution containing QX-314 to optimize detection of inhibitory postsynaptic currents (IPSCs) and 0.02% Lucifer yellow (Shao et al. 2012). The final pipette tip resistance was 5.0 to 5.5 MΩ, and seal resistance was routinely >4 GΩ. All data were acquired with pCLAMP 9 software using a MultiClamp 700A amplifier digitized with a DigiData 1322A A/D board (Axon Instruments), low-pass filtered online at 2 (voltage clamp, sampling rate of 5 kHz) and 10 kHz (current clamp, sampling rate of 40 kHz).
MCs were selected for analysis only when they contained an intact apical dendrite with a visible intraglomerular tuft and exhibited membrane bistability in spontaneous activity (Heyward et al. 2001). Membrane bistability, an intrinsic property of MCs, is sensitive to mechanical damage in slice preparations. MCs close to the cut surface of a slice rarely exhibit membrane bistability whereas MCs deeper than 50 μm are typically bistable; MCs can lose bistabilty with time in vitro but never regain this property (Heyward et al. 2001). Thus membrane bistability is taken as an index of MC “health” and experiments were performed only on bistable MCs.
Lucifer yellow was observed to identify the apical tuft of the MC such that the stimulation electrode could be positioned in the ON lightly anterior to the glomerulus containing the apical tuft (ON bundles course generally anterior to posterior across the bulb) and a microinjection pipette could be positioned at the apical tuft. Constant current electrical stimulation (170-μs duration, 36 ± 5 μA; PG4000A Digital Stimulator and SIU91 stimulus isolator; Cygnus Technology) was applied with dual barrel glass microelectrodes (Borosilicate Theta, 5- to 10-μm tip pulled on a P-97 Flaming-Brown puller). Stimulation with trains of 5 shocks delivered at 1, 3, 5, or 8 Hz with 10-s recovery periods between each train. At least 10 sets of train stimulations were tested in aCSF control and in the presence of gabazine (GBZ). To assess whether rundown across the sets of train stimulations might impact response, we analyzed spikes evoked during the first train of stimulations compared with the spikes evoked during the last set of train stimulations and found no significant differences.
Microinjection of GBZ into a target glomerulus was performed using 30 psi pneumatic pressure applied for 2 ms (via a picospritzer; General Valve). Injection pipettes were made from thick wall borosilicate glass with filament (I.D.: 0.75 mm pulled on a vertical pipette puller, 6-μm tip opening). These parameters were calibrated to deliver an injection volume of ∼20 nl and were previously shown to block inhibition in a single glomerulus (Shao et al. 2012).
IPSCs were detected on the basis of amplitude and area having at least 3:1 signal-to-noise ratio measured in Minianalysis software (Synaptosoft). In current-clamp recordings, action potentials were detected in pClamp and analyzed by Neuroexplorer (NEX Technologies). Comparisons of IPSC/Ps evoked by ON stimulation were performed on amplitude and area under the curve (charge), measured by Minianlysis software. IPSCs were detected for each of multiple sweeps (6–10 sweeps) and the detected IPSCs averaged from across those sweeps to calculate a peristimulus time histograms (PSTH) using a custom Excel spreadsheet for each cell. Population PSTHs were calculated by normalizing each cell response at each time point to the first bin following ON stimulation in aCSF and averaging each time point and treatment condition across multiple cells to generate the population response. Spontaneous IPSCs were analyzed from a minimum of 2 min of recording and >1,000 IPSCs.
Based on our previous study (Shao et al. 2012) we operationally defined an early (0–20 ms) and late (50–800 ms) window for analysis of inhibitory currents. Net charge (pA·ms) was measured from the 0- to 20-ms time window. At the holding potentials utilized near equilibrium potentials the excitatory current is minimized, although the addition of GBZ unmasked a minor excitatory current. Area was measured from 6 to 10 sweeps at each frequency of stimulation under aCSF and then again under GBZ glomerular microinjection. In current-clamp recordings, action potentials were detected in pClamp and analyzed by Neuroexplorer (NEX Technologies).
Spike PSTHs were generated using a custom Excel spreadsheet. Population PSTHs were generated by compiling spike counts in 10-ms time bins for all ON stimulations repeated at a given frequency and by normalizing each cell using the cells total spiking across the whole trace in aCSF and then calculating the population mean (±SE) for each experimental condition. Rundown in spiking can occur during long recording sessions. To assess rundown in our recorded MCs, spiking in sweeps early in the recording session (containing 1, 3, 5, and 8 Hz stimulations) were compared with sweeps late in the recording session. All cells meeting the membrane bistability criteria above also exhibited <10% variance in overall spiking across the duration of the recording session indicating rundown in these cells was not occurring. Decay rate curves were generated with SigmaPlot 12 using a double exponential.
Unless otherwise stated data are represented as means ± SE with n indicating the number of cells examined. Statistical tests were performed on raw nonnormalized data using t-tests, one-way ANOVA, or MANOVA (following the general linear model and post hoc with the Bonferroni test) to test for statistical significance.
RESULTS
Single pulse stimulation of ON input to MCs evokes an initial EPSC (2.12 ± 0.18 ms latency) followed by a fast (6.62 ± 0.09 ms latency), strongly summating barrage of IPSCs lasting ∼40 ms (“early” IPSCs) and then intermittent IPSCs spread over a period of ∼800 ms (“late” IPSCs; Shao et al. 2012). Intraglomerular circuits drive all of the early inhibition and half of late inhibition whereas the MC-GC-MC circuit contributes only to late inhibition (Shao et al. 2012). The magnitude and temporal structure of MTC spike responses to odors are remarkably stable across sniff frequencies in vivo (Carey and Wachowiak 2011). We hypothesized that intraglomerular inhibition might contribute to stabilizing the temporal structure of MC responses. To explore this we recorded synaptic currents evoked in MCs by trains of low intensity (36 ± 5 μA) ON shocks delivered at frequencies that span the range of sniffing (1 to 8 Hz).
At frequencies from 1 to 5 Hz, the integrated area (charge, pA·ms) of the early IPSCs of the first vs. subsequent stimuli in the train was statistically indistinguishable (n = 8 cells; Fig. 1, A and B). At 8 Hz, the IPSC area of the s stimulus was reduced 36.4 ± 8% relative to the first but there were no differences among the second to fifth early IPSCs. Late IPSCs were measured from a 75- to 125-ms poststimulus time window (maximum window size available at 8 Hz). Similar to early IPSCs, late IPSCs did not differ across 1- to 8-Hz stimulation frequencies (n = 7 MCs; Fig. 1C). This shows that the magnitude and duration of both early and late inhibition are independent of ON input frequency over the range of 1 to 5 Hz and early IPSCs are only slightly attenuated at 8 Hz. Thus, while ON evoked late IPSCs might temporally overlap with early IPSCs of the next stimulus, they do not summate.
Fig. 1.
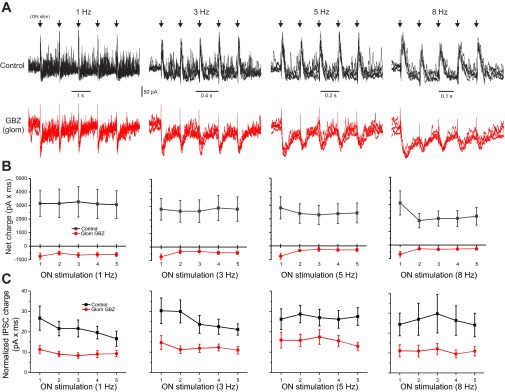
Mitral cell (MC) inhibitory postsynaptic current (IPSC)/excitatory postsynaptic current (EPSC) responses to olfactory nerve (ON) input at frequencies from 1 to 8 Hz are invariant. A: voltage-clamp recording (holding potential = −10 mV) of a MC in artificial cerebral spinal fluid (aCSF; top, black) and following glomerular restricted application of 100 μM gabazine (GBZ; bottom, red). ON stimulation at 1, 3, 5, and 8 Hz (arrows) produced a rapid onset barrage of IPSCs followed by lower frequency intermittent IPSCs. Note, the x-axis scale in each panel is different to maintain stimulation spacing constant. B: population average of MC “early” net charge (pA × ms) at each ON input frequency in aCSF (black, n = 8 cells) and following application of GBZ (red). Addition of glomerular GBZ abolishes early inhibitory currents and unmasks a small excitatory current. C: population average of MC “late” IPSC charge at each ON stimulation frequency. Although, variability between cells in late IPSCs is higher than in early IPSCs, no significant differences with late IPSCs were observed across ON stimulations from 1 to 8 Hz. Addition of GBZ attenuates late IPSCs ∼50%.
Early IPSCs evoked by a single ON shock are due to intraglomerular inhibition as they are completely eliminated when glomerular GABAergic synapses are selectively blocked by discrete intraglomerular microinjection of the GABAA receptor antagonist GBZ (Shao et al. 2012). Early IPSCs are also resistant to the N-methyl-d-aspartate receptor antagonist dl-2-amino-5-phosphonovaleric acid, which selectively attenuates mitral-to-granule transmitter release (Isaacson and Strowbridge 1998; Schoppa et al. 1998; Chen et al. 2000; Shao et al. 2012). In the present experiments, GBZ puffs abolished the early IPSCs at all frequencies of train stimulation (Fig. 1, A and B). Intraglomerular GBZ also reduced the late IPSCs by ∼50% and increased the duration and magnitude of the initial EPSC response, as previously found for single shocks (Shao et al. 2012). In additional experiments (not shown), addition of 50 μM dl-2-amino-5-phosphonovaleric acid did not alter the early IPSC area at any frequency tested. Thus early inhibition and approximately half of the late inhibition are due to intraglomerular GABA release across input frequencies.
The magnitude and temporal structure of MTC spike responses to odors are stable across sniff frequencies. We reasoned that this might be due, in part, to the relative insensitivity of intraglomerular inhibition to changes in input frequency and that intraglomerular inhibition might function, in part, to normalize MC response across sniffs.
To investigate this, we recorded MC spike responses to 1- to 8-Hz ON train stimuli. At 1 Hz, MCs respond with initial burst of spikes followed by reduced spiking that decays back to baseline firing before the next pulse in the train (Fig. 2, A and B). To analyze spike responses to higher frequency trains we analyzed the steady-state spiking responses from the third to fifth stimuli in the train. In response to the first stimulus in a train (equivalent to a single shock), MCs respond with a burst of spikes peaking at ∼60 Hz with spike rate decaying back to baseline after ∼600 ms (time constant 93.7 ms; present experiments and Shao et al. 2012). At higher frequencies (3–8 Hz), spiking does not return to baseline before the next stimulus in the train. Despite this overlap the maximum MC firing rate to subsequent stimuli is unchanged (Fig. 2E). However, response contrast (maximum-to-minimum firing rate) is reduced because the response begins from a higher baseline firing rate. To quantify this we took the ratio of the firing rate in a 20-ms window after stimulation to the minimum rate 20 ms preceding the stimulus to give a max-min ratio. Max-min ratio ranged from 30:1 at 1 Hz to 10:1 at 8 Hz (Fig. 2D). Thus, even at 8 Hz, the highest train frequency tested, there is still ∼10-fold difference between the maximum and minimum response, providing significant spike response contrast to each stimulus in the train.
Fig. 2.

Intraglomerular inhibition reduces MC spike responses to ON stimulation at input frequencies >3 Hz. A: current-clamp recording of ON evoked spikes in a MC in aCSF (black) and following restricted glomerular GBZ application (red). Note, the x-axis scale in each panel is different to maintain stimulation spacing constant. B1 and B2: population peristimulus time histograms (PSTH) of MC spike count following ON stimulation at 1, 3, 5, and 8 Hz (arrows, n = 8 cells) in aCSF (black) superimposed on the spike PSTH following glomerular restricted application of 100 μM GBZ (red). Glomerular injection of 100 μM GBZ dramatically increases spike output. Note, the x-axis time scale in B1 vary to maintain constant stimulation spacing, B2 shows the same PSTH with a fixed time scale. C: curve fitting of the population MC spiking decay following stimulation 1 (top trace) and stimulation 5 (bottom trace) at 1, 3, 5, and 8 Hz. D: curve fitting of the population MC spiking decay in aCSF and glomerular injection of GBZ showing average for stimulations 1–5 showing the major slowing of decay rate with GBZ. E: curve fitting of the population MC spiking decay in aCSF and glomerular injection of GBZ averaged for frequencies 1, 3, 5, and 8 Hz. In aCSF or GBZ there is a slight slowing of decay rate at higher frequencies. F: ratio of maximum/minimum spike counts at each frequency. aCSF (black, n = 16 cells); Glom GBZ: restricted glomerular gabazine application (red, n = 8 cells); Glom GBZ (hyperpolarization): restricted glomerular gabazine application on hyperpolarized MTCs (blue, n = 8 cells) and computation model of tonic inhibition (green, n = 8 cells). G: population mean of the ratio between maximum and minimum spiking at each stimulation frequency in aCSF (left) and following GBZ (right) for steady-state responses from the 3rd to 5th stimulations in the ON stimulation train. Blocking interglomerular inhibition with GBZ decreases the max/min ratio as input frequency increases. H: population mean of the maximum spike frequency in each stimulation frequency in aCSF (left) and following GBZ (right) shows no significant differences in maximum spiking across frequencies.
The max-min ratio shows a response contrast as input frequency changed but does not provide detail on the “shape” of MC responses as they decay back to baseline. To analyze spiking decay back to baseline, we curve fitted aCSF responses from stimulation 1 through to stimulation 5 at each of the frequencies utilizing a double exponential (best fit). Stimulation 1 showed a highly similar decay at all frequencies (Fig. 2C); however, by stimulation 5 for the 8-Hz input there was a modest slowing in the MC spiking decay rate (Fig. 2C). Averaging all frequency responses for stimulations 1–5 indicate decay rates relatively similar to each stimulation in aCSF (Fig. 2D). When the decay rates are sorted by frequency, there is a slowing in MC spiking decay rate at 8 Hz (Fig. 2E).
How might intraglomerular inhibition influence response contrast and shape? When GBZ was microinjected into the glomerulus containing the recorded MC dendrite, maximum spiking in the first 20 ms did not significantly change (65 Hz in aCSF to 70 Hz in GBZ, measured from the steady-state responses at stimulations 3–5 at each train frequency; Fig. 2H). This suggests that the maximum sustainable MC spiking rate is approximately 60–80 Hz. However, as with single stimulations (Shao et al. 2012), the duration of ON-evoked MC spiking dramatically increased when intraglomerular inhibition was blocked (Fig. 2B). Although GBZ increased the duration of spiking response, it still decayed back to baseline before the next pulse for 1-Hz trains; thus the max-min ratio was ∼30:1 in both aCSF and GBZ (Fig. 2F, no significant difference). However, at 3–8 Hz trains block of intraglomerular inhibition caused a significant, frequency-dependent decrease in the max-min ratio from 30:1 in aCSF to 10:1 in GBZ at 3 Hz, 20:1 in to 5:1 at 5 Hz, and 15:1 to 2:1 at 8 Hz (Fig. 2G). The drop in response contrast with block of intraglomerular inhibition is mirrored by a dramatic increase in the time taken for the response to decay to baseline (Fig. 2, D and E). While stimulation frequency does have an impact, with slower decay at higher frequency, the most dramatic effect is following GBZ block of intraglomerular inhibition.
Blocking intraglomerular GABAA receptors by focal intraglomerular application of GBZ increases MC spontaneous spiking due to loss of postsynaptic inhibition (Shao et al. 2012). How might this influence response contrast during repetitive stimulation? To assess this, we arithmetically added the baseline firing in the presence of GBZ to the spiking in aCSF (imposing a ceiling of 80 Hz to replicate maximum firing rates) and recalculated response contrast for each rate of stimulation. At 8 Hz the estimated increase in spiking attributable to GBZ-induced spontaneous firing could account for ∼40% of response contrast decrease. However, this computational estimate assumes a degree of response linearity that may not be present in live cells. Therefore, we mimicked the tonic inhibition in GBZ blocked MCs by injecting current into the cell sufficient to restore spontaneous spike rate to that observed in aCSF. Cells were first held in aCSF, and the spontaneous spiking rate was measured for each cell (mean 2.16 ± 0.73 Hz; n = 8 cells). Intraglomerular inhibition was blocked by microinjection of GBZ, and the cell hyperpolarized (−8.8 ± 2 pA; n = 8 cells) sufficient to restore each cell to its pre-GBZ aCSF spontaneous spike rate (mean 2.02 ± 0.62 Hz in GBZ with hyperpolarization; n = 8 cells). In this condition, ON stimulation still produced marked elevation of spiking across the response envelope compared with aCSF (Fig. 2F) with restoration of ∼50% of the response contrast. Thus our computational estimate and experimental results are in agreement and suggest that intraglomerular inhibition has both a tonic drive (mimicked by our hyperpolarization) and a stimulus-evoked phasic that act in concert to improve MC response contrast approximately fivefold for repetitive inputs ≥3 Hz by rapidly “resetting” MC spiking following stimulation. Taken together, these results suggest that intraglomerular inhibition could play a key role in shaping MC response contrast to odors during repetitive sniffing.
DISCUSSION
The olfactory system receives rhythmic sensory input dictated by respiration rate and active sniffing. Animals dynamically alter sniffing rates and patterns when investigating the environment. Thus the neural circuits that transform peripheral sensory signals into outputs to higher brain regions must maintain information fidelity across varying input frequencies up to ∼8 Hz. Intraglomerular circuits are well suited to regulate temporal patterns of MC output to downstream olfactory networks.
ON excitatory inputs to MCs are followed by inhibition with two distinct temporal phases: an early, summating barrage of IPSCs and later intermittent IPSCs. Intraglomerular block of GABAA receptors selectively and completely abolishes the early inhibition whereas selective suppression of GABA release from GCs attenuates the late but not the early IPSCs (Shao et al. 2012) indicating that MCs are differentially regulated by intraglomerular and MC-GC-MC circuits. Intraglomerular inhibition potently regulates ON evoked MC spike responses: ∼10-fold more spikes occur when intraglomerular inhibition is selectively blocked. MCs receive tonic intraglomerular postsynaptic inhibition (Shao et al. 2012). Blocking this tonic inhibition increases MC spontaneous firing rates. However, linear addition of the increase in firing does not fully replicate the increased time constants of MC spiking in the presence of GBZ (Shao et al. 2012). This indicates that intraglomerular inhibition regulates MC spiking in response to repetitive input, partially through ongoing tonic inhibition as well as by phasic inhibition evoked by input. We tested this idea and showed that intraglomerular inhibition greatly improves MC spike response contrast to successive stimuli across the range of sniffing frequencies. When intraglomerular inhibition is blocked, response contrast falls approximately fivefold across 3- to 8-Hz input frequencies.
In behaving rats, the initial MC response is invariant to changes in respiration (Cury and Uchida 2010). However, increased sniff frequency is associated with faster rise and decay times in the MC spike response envelope (Carey and Wachowiak 2011). We showed that intraglomerular inhibition shortened decay times of responses to train stimuli; when intraglomerular inhibition is blocked, decay times are prolonged and spiking persists across successive stimuli. This suggests that glomerular inhibitory circuits regulate the temporal structure of MC responses to maintain coding contrast of sniff-sampled inputs. However, in vivo at higher frequencies (>8 Hz) decay kinetics of MC spiking deviate from a linear prediction at lower frequencies, which suggested that additional inhibitory mechanisms engage at elevated frequencies to bring MC spiking back towards baseline more rapidly. From our observations, inhibition arising from glomeruli is relatively constant across all input frequencies tested. The additional inhibition observed at high frequencies in vivo may be due in part to cortically mediated feedback onto granule (Gao and Strowbridge 2009; Strowbridge 2010; Boyd et al. 2012; Markopoulos et al. 2012) cells, a circuit truncated within slices, or as a consequence of odorant receptor desensitization dynamics at the level of the ORN.
The relative strengths of intraglomerular excitation and inhibition, and the timing of these excitatory-inhibitory sequences, determine how MC spiking is gated. Intraglomerular inhibition operates as a fast-feed forward circuit: ON input activate ETCs, which in turn activate GABAergic PGCs to generate postsynaptic inhibition in MCs. Rapid feed forward inhibition by this ON→ETC→PGC→MC circuit is important in gating MC spike output. Other inhibitory circuits, interglomerular inhibition, the MC→GC→MC circuit, and cortical feedback to GCs (Balu et al. 2007; Strowbridge 2009; Boyd et al. 2012; Markopoulos et al. 2012), act in concert with intraglomerular feed forward inhibition to regulate the olfactory bulb input-output transform. The relative contributions of each of these inhibitory circuits during odor input in awake behaving animals remain to be determined. Recent work suggests that difficult odor discriminations involve longer time courses in vivo. These perceptual processes may be more heavily impacted by late IPSCs from the MC-GC-MC circuit (Abraham et al. 2010) and/or cortical feedback (Balu et al. 2007; Strowbridge 2009; Doucette et al. 2011; Boyd et al. 2012; Markopoulos et al. 2012) across multiple sniffing cycles.
GRANTS
This work was supported by National Institutes of Health Grants DCD-005676 and DCD-19015.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
Author contributions: Z.S., A.C.P., and M.T.S. conception and design of research; Z.S. performed experiments; Z.S. and A.C.P. analyzed data; Z.S., A.C.P., and M.T.S. interpreted results of experiments; Z.S. and A.C.P. prepared figures; Z.S., A.C.P., and M.T.S. drafted manuscript; Z.S., A.C.P., and M.T.S. edited and revised manuscript; M.T.S. approved final version of manuscript.
REFERENCES
- Abraham NM, Egger V, Shimshek DR, Renden R, Fukunaga I, Sprengel R, Seeburg PH, Klugmann M, Margrie TW, Schaefer AT, Kuner T. Synaptic inhibition in the olfactory bulb accelerates odor discrimination in mice. Neuron 65: 399–411, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aungst JL, Heyward PM, Puche AC, Karnup SV, Hayar A, Szabo G, Shipley MT. Centre-surround inhibition among olfactory bulb glomeruli. Nature 426: 623–629, 2003 [DOI] [PubMed] [Google Scholar]
- Balu R, Pressler RT, Strowbridge BW. Multiple modes of synaptic excitation of olfactory bulb granule cells. J Neurosci 27: 5621–5632, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boyd AM, Sturgill JF, Poo C, Isaacson JS. Cortical feedback control of olfactory bulb circuits. Neuron 76: 1161–1174, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carey RM, Wachowiak M. Effect of sniffing on the temporal structure of mitral/tufted cell output from the olfactory bulb. J Neurosci 31: 10615–10626, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen WR, Xiong W, Shepherd GM. Analysis of relations between NMDA receptors and GABA release at olfactory bulb reciprocal synapses. Neuron 25: 625–633, 2000 [DOI] [PubMed] [Google Scholar]
- Cury KM, Uchida N. Robust odor coding via inhalation-coupled transient activity in the mammalian olfactory bulb. Neuron 68: 570–585, 2010 [DOI] [PubMed] [Google Scholar]
- Doucette W, Gire DH, Whitesell J, Carmean V, Lucero MT, Restrepo D. Associative cortex features in the first olfactory brain relay station. Neuron 69: 1176–1187, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao Y, Strowbridge BW. Long-term plasticity of excitatory inputs to granule cells in the rat olfactory bulb. Nat Neurosci 12: 731–733, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heyward P, Ennis M, Keller A, Shipley MT. Membrane bistability in olfactory bulb mitral cells. J Neurosci 21: 5311–5320, 2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Isaacson JS, Murphy GJ. Glutamate-mediated extrasynaptic inhibition: direct coupling of NMDA receptors to Ca2+-activated K+ channels. Neuron 31: 1027–1034, 2001 [DOI] [PubMed] [Google Scholar]
- Isaacson JS, Strowbridge BW. Olfactory reciprocal synapses: dendritic signaling in the CNS. Neuron 20: 749–761, 1998 [DOI] [PubMed] [Google Scholar]
- Liu S, Plachez C, Shao Z, Puche A, Shipley MT. Olfactory bulb short axon cell release of GABA and dopamine produces a temporally biphasic inhibition-excitation response in external tufted cells. J Neurosci 33: 2916–2926, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Markopoulos F, Rokni D, Gire DH, Murthy VN. Functional properties of cortical feedback projections to the olfactory bulb. Neuron 76: 1175–1188, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parrish-Aungst S, Shipley MT, Erdelyi F, Szabo G, Puche AC. Quantitative analysis of neuronal diversity in the mouse olfactory bulb. J Comp Neurol 501: 825–836, 2007 [DOI] [PubMed] [Google Scholar]
- Phillips CG, Powel TPS, Shepherd GM. Responses of mitral cells to stimulation of the lateral olfactory tract in the rabbit. J Physiol 168: 65–88, 1963 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schoppa NE, Kinzie JM, Sahara Y, Segerson TP, Westbrook GL. Dendrodendritic inhibition in the olfactory bulb is driven by NMDA receptors. J Neurosci 18: 6790–6802, 1998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shao Z, Puche AC, Kiyokage E, Szabo G, Shipley MT. Two GABAergic intraglomerular circuits differentially regulate tonic and phasic presynaptic inhibition of olfactory nerve terminals. J Neurophysiol 101: 1988–2001, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shao Z, Puche AC, Liu S, Shipley MT. Intraglomerular inhibition shapes the strength and temporal structure of glomerular output. J Neurophysiol 108: 782–793, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shepherd GM. Neuronal systems controlling mitral cell excitability. J Physiol 168: 101–117, 1963 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shirley CH, Coddington EJ, Heyward PM. All-or-none population bursts temporally constrain surround inhibition between mouse olfactory glomeruli. Brain Res Bull 81: 406–415, 2010 [DOI] [PubMed] [Google Scholar]
- Strowbridge BW. Role of cortical feedback in regulating inhibitory microcircuits. Ann NY Acad Sci. 1170: 270–274, 2009 [DOI] [PubMed] [Google Scholar]
- Strowbridge BW. Linking local circuit inhibition to olfactory behavior: a critical role for granule cells in olfactory discrimination. Neuron 65: 295–297, 2010 [DOI] [PubMed] [Google Scholar]