

Abstract
Purpose
To investigate the influence of sperm DNA integrity on the zona binding ability of mouse spermatozoa in relation to their sex chromosomal constitution.
Method(s)
In this prospective experimental study, the sperm DNA fragmentation was induced by exposing testicular area of Swiss Albino mice (Mus musculus) to different doses of γ-radiation (0, 2.5, 5.0 and 10.0 Gy). Sperm DNA fragmentation was quantified by single cell gel electrophoresis (comet assay). In vitro sperm zona binding assay was performed and the numbers of zona bound X and Y bearing spermatozoa were determined using fluorescence in situ hybridization (FISH).
Result(s)
The assessment of zona pellucida bound X and Y-bearing spermatozoa using fluorescence in situ hybridization has revealed a unique binding pattern. The number of zona bound Y-spermatozoa declined significantly (P < 0.01 to 0.0001) with increase in the DNA damage. The skewed binding pattern of X and Y-bearing sperm was strongly correlated with the extent of sperm DNA damage.
Conclusion(s)
The zona pellucida may have a role in preventing DNA damaged mouse sperm binding especially towards Y-bearing sperm. However, the exact mechanism behind this observation needs to be elucidated further.
Electronic supplementary material
The online version of this article (doi:10.1007/s10815-013-0123-x) contains supplementary material, which is available to authorized users.
Keywords: Sperm DNA damage, Sperm zona binding, Functional competence
Introduction
Sperm chromatin integrity plays an important role in accurate transmission of genetic material to the offspring [1]. It has been shown that numerous exogenous and endogenous factors play a definitive role on sperm chromatin to carry excessive damage load during spermatogenesis or their stay in the reproductive tract [2]. Earlier studies in murine and bovine species under in vitro conditions have shown that DNA-damaged sperm, regardless of degree of damage has the ability to fertilize the oocyte but further embryonic development and pregnancy was very much related to the extent of DNA damage [3–5].
The zona pellucida has proven to be an important barrier against functionally abnormal spermatozoa. It has been shown that human zona pellucida has the capacity to select against aneuploid spermatozoa by unknown mechanism [6], hence may serve as extremely stringent tool to prevent fertilization with genetically abnormal sperm. However, there are contradicting reports regarding the influence of sperm DNA damage on fertilization [3, 4]. Fertilization with DNA damaged sperm can increase the risk of transgenerational genomic instability in both somatic as well as germ cell compartment of first generation offspring [5]. As the correlation between sperm functional characteristics and DNA integrity is lacking [7], and the ability of the zona pellucida in preventing fertilization by DNA damaged sperm is also possible [8], this study was conducted to understand the zona binding competence of DNA damaged spermatozoa in relation to their sex chromosome. The spermatozoa bound to zona pellucida were identified for sex chromosomes to find out whether there is any sex specific association with DNA integrity and functional competence.
Materials and methods
Animals
The animal care and handling were done according to the Institutional Guidelines for animal experimentation and the proposal was approved by the Institutional Ethical Committee. Six to eight weeks old healthy Swiss Albino mice were used for the experiments and they were maintained under the controlled conditions of temperature (23 ± 2 °C), humidity (55 ± 5 %) and light (12 h light/dark cycle) with standard diet and water ad libitum. Female mice were superovulated with intra peritoneal injection of 5 IU pregnant mare serum gonadotropin (PMSG) followed by the human chronic gonadotropin (hCG) administration (10 IU) after 48 h. After 13 h, oocytes were collected from the oviduct.
Induction of sperm DNA damage
Sperm DNA damage was induced by partial body irradiation to the testicular area of males covering the rest of the body with lead shields. A dose of 0 to 10 Gy was delivered at a dose rate of 1 Gy/min from 60Co (60Cobalt) teletherapy unit.
Sperm extraction and quantification of DNA damage by single cell gel electrophoresis (comet assay)
Eighteen hours after irradiation, animals were sacrificed by cervical dislocation and spermatozoa were extracted from the cauda epididymis in two milliliter of pre-warmed Earle’s Balanced Salt Solution (EBSS). The sperm suspension was analyzed for DNA integrity by alkaline comet assay as described earlier [5]. Briefly, the spermatozoa were washed by centrifugation and the resulting sperm pellet was mixed with sterile phosphate buffered saline (pH 7.4). The sperm density was kept constant at 1 × 104 by appropriate dilution in order to maintain the uniform distribution of the spermatozoa during electrophoresis. The sperm suspension was mixed with 0.8 % low melting agarose in equal volume and layered on a slide pre coated with 1 % normal agarose. A third coat of 0.75 % agarose was layered followed by overnight incubation in lysing solution at 4 °C. Sperm head decondensation was done using 10 mM dithiothreitol (DTT) for 2 h at 4 °C. After allowing the sperm DNA to unwind in electrophoresis buffer (pH > 13) for 20 min, electrophoresis was carried out at 25 V for 20 min. The slides were stained with Ethidium bromide (2 μg/ml) and observed under fluorescent microscope (Imager-A1, Zeiss, Germany). The DNA fragmentation was calculated in at least 50 spermatozoa per slide using Kinetic Imaging software (Komet 5.5, UK). A minimum of five animals were used per data point and the experiments were repeated to confirm the reproducibility of the results. The extent of DNA damage in the spermatozoa was determined and presented as olive tail moment (OTM, product of the tail length and the fraction of total DNA in the tail).
Assessment of fertilizing ability of DNA damaged spermatozoa in vivo
Eighteen hours after irradiation, male mice were mated with unirradiated normal female mice (1:1 ratio) in the estrous stage. Mating was allowed for 2 h and successful mating was confirmed by the presence of vaginal plug. The oocytes were collected from oviduct 10 h post-mating and assessed for fertilization by observing the presence of two pronucleus and two polar bodies. A minimum of 50 oocytes were assessed for each group and percentage fertilization rate was calculated.
Sperm zona binding assay
Caudal spermatozoa collected in approximately 2 ml of pre-equilibrated EBSS supplemented with 0.1 % Bovine Serum Albumin (BSA) were incubated at 37 °C and 5 % CO2 for 20 min. Then, sperm suspension was centrifuged at 1,200 rpm for 8 min and this pellet was resuspended in 0.5 mL EBSS supplemented with 0.1 % BSA and the motile sperm concentration was adjusted to 1 million/mL. Oocytes were denuded using 80 IU hyaluronidase and 10 oocytes were inseminated in 100 μl sperm suspension droplet covered with paraffin oil and incubated at 37 °C and 5 % CO2. After 2 h, oocytes were washed three times in 50 μl droplets of EBSS and the zona bound spermatozoa collected using 135 μm denuding pipette (MXL3-135, Origio, USA) in 10 μl EBSS. About 126 to 440 oocytes from >10 females and spermatozoa from >10 males were used for analysis per data point. These zona bound spermatozoa were transferred to poly-L-lysine pre-coated glass slides and subjected to fluorescence in situ hybridization (FISH).
Fluorescence in situ hybridization
The in situ hybridization was performed as described by Sarrate and Anton [9] with minor modifications. The smears were air dried followed by fixation in freshly prepared Carnoy’s fixative (methanol: acetic acid, 3:1). Sperm decondensation was achieved using 25 mM dithiothreitol dissolved in lysis solution for 5 min at room temperature followed by washing with 2X saline-sodium citrate (SSC) buffer. Slides were dried for 10–15 min and then immersed in pre-treatment solution (2 X SSC, pH 7.4) at 73 °C for 2 min followed by washing in water. The cells were treated with protease solution (Pepsin, Cat. No. P7012; Sigma Aldrich Inc. USA) dissolved in 10 mM HCl for 15 min, washed in PBS and then dehydrated by serial graded ethanol solutions. About 12.5 μL of FISH probes (X and Y, ID Labs Inc, UK) were added onto the cells and denatured at 73 °C for 5 min followed by hybridization for 16 h at 37 °C in a hybridization chamber (Thermobrite, Abbott molecular, USA). The slides were placed in 2 X SSC/0.1 % NP-40 at room temperature for 1 min and agitated. The slides were washed in 0.4 X SSC/0.3 % NP-40 at 73 °C for 2 min followed by washing in 2 X SSC/0.1 % NP-40 at room temperature for 1 min and then counter stained with DAPI. The slides were observed using appropriate filter under fluorescent microscope (Imager-A1, Zeiss, Germany) at 100 X magnification (Supplementary figure 1). All the slides were coded to avoid observer’s bias before evaluated for X and Y spermatozoa.
Statistical analysis
One Way Analysis of Variance (ANOVA) was used to determine the significance of OTM in various groups. The level of significance between the number of Y bearing spermatozoa was determined using Chi-square test (degree of freedom-1), two sided P value is expressed using GraphPad instat 3 (USA).
Result
Successful induction of DNA damage in the spermatozoa
The local testicular irradiation of 0, 2.5, 5.0 and 10.0 Gy was used to induce varying level of DNA damage in mouse spermatozoa. The sperm characteristics at 18 h after irradiation did not show any significant differences with respect to concentration, motility and viability (Table 1). To determine the extent of radiation induced DNA fragmentation in caudal spermatozoa, the data on olive tail moment (OTM, product of the tail length and the fraction of total DNA in the tail) was collected. The spermatozoa exposed to 2.5 Gy radiation had OTM of 6.17 ± 0.32 which was significantly (P < 0.05) higher than the control group (4.51 ± 0.33). Further increase in OTM was observed in 5 Gy (8.84 ± 0.42) and 10 Gy (9.74 ± 0.42) groups which were statistically higher than control group (P < 0.001) (Table 2). The fertilizing ability was performed in vivo and it was significantly different (P < 0.05 to 0.001) between each group (Table 3).
Table 1.
Effect of different doses of γ-radiation on sperm functional characteristics
| Radiation dose (Gy) | Sperm characteristics | ||
|---|---|---|---|
| Concentration (M/ml ± SEM) |
Motility (% ± SEM) |
Viability (% ± SEM) |
|
| Control | 27.4 ± 1.9 | 45.2 ± 1.7 | 66.4 ± 2.6 |
| 2.5 | 29.4 ± 0.9 | 43.4 ± 1.4 | 63.8 ± 3.3 |
| 5.0 | 25.8 ± 1.1 | 43.2 ± 2.6 | 63.4 ± 1.9 |
| 10.0 | 28.2 ± 2.2 | 45.0 ± 1.1 | 58.6 ± 2.7 |
Table 2.
Effect of different levels of sperm DNA fragmentation on the zona pellucida binding competence of X and Y bearing spermatozoa
| Radiation dose (Gy) | OTM (± SEM) |
Oocytes (N) |
Spermatozoa assessed (N) | X bearing spermatozoa (%) | Y bearing spermatozoa (%) |
|---|---|---|---|---|---|
| Control | 4.51 ± 0.33 | 126 | 2537 | 1031(40.4) | 1506 (59.6) |
| 2.5 | 6.17 ± 0.32a | 155 | 2294 | 1018 (44.5) | 1276 (55.5)c |
| 5.0 | 8.84 ± 0.42b | 126 | 1561 | 786 (50.4) | 775(49.6)d |
| 10.0 | 9.74 ± 0.42b | 440 | 4495 | 2399 (53.4) | 2096 (46.6)d |
a = P < 0.05 Vs control; b, c = P < 0.001 Vs corresponding control; d = P < 0.0001 Vs control
Table 3.
The ability of γ-irradiated spermatozoa to fertilize normal oocytes in vivo
| Group | Number of egg assessed (N) | Fertilized/Unfertilized egg | Fertilization rate (%) (Mean ± SEM) |
|---|---|---|---|
| Control | 59 | 56/3 | 95.0 ± 3.33 |
| 2.5 Gy | 58 | 51/7 | 88.03 ± 6.30 |
| 5.0 Gy | 58 | 39/19 | 67.43 ± 3.91a,b |
| 10 Gy | 55 | 26/29 | 47.63 ± 6.18c,d |
a P < 0.001 Vs Control; b P < 0.05 Vs 2.5 Gy; c P < 0.001 Vs Control and 2.5 Gy; d P < 0.05 Vs 5.0 Gy
Number of spermatozoa bound to zona pellucida in relation to sex chromosome
The distribution of X and Y bearing sperm in control group prior to insemination revealed about 46.4 % X and 53.7 % Y bearing sperm. The average number of sperm bound to an oocyte in control group was approximately 20 and the number was reduced to 14.8, 12.3 and 10.2 in 2.5, 5.0 and 10.0 Gy groups respectively. In addition, there was a moderate difference in the number of X and Y bearing spermatozoa bound to zona pellucida in control group (40.4 % Vs 59.3 %). However, in 2.5 Gy group, the number of Y bearing sperm bound to zona pellucida reduced significantly from control (P < 0.001) whereas the number of X bearing spermatozoa bound to the zona pellucida was significantly higher than the control (P < 0. 001). In addition, at 5.0 and 10.0 Gy group, a further difference in the binding pattern was observed which was significantly different from the control group (P < 0.0001). In contrast, the distribution of X and Y bearing spermatozoa was inversely related with increase in DNA damage (OTM) (Table 2). Small fractions of the spermatozoa also showed signals for both X and Y bearing chromosomes. However, the incidence was very low and specific to 5 and 10 Gy group (0.19 and 0.7 % in 5 and 10 Gy group respectively).
Discussion
Fertilization begins when spermatozoa bind to the extracellular matrix coating the oocyte, zona pellucida. Here, we investigated the zona binding competence of DNA damaged spermatozoa in relation to their sex chromosome status. Interestingly, the data presented in this study has demonstrated a novel, negative influence of sperm DNA fragmentation preferentially on Y chromosome bearing spermatozoa resulting in the reduced zona binding with respect to the extent of sperm DNA damage.
Sperm DNA integrity is an important factor in establishing successful pregnancy and reproductive outcome [10]. Gathering facts from our earlier data [11] and other studies [12, 13], there is apparently evidence to show that fertilization with DNA damaged sperm induces variety of biological response in embryos and even increases the risk of transgenerational genomic instability in both somatic as well as germ cell compartment of first generation offspring [5].
During natural postovulatory mating, there is a preliminary selection process against spermatozoa with low motility and fragmented DNA during the transport through the female reproductive tract. However, the ability of the zona pellucida in recognizing the DNA damaged spermatozoa is achieved during sperm zona penetration and it depends on the source and type of damage [8]. Hence, it is possible that selectivity of DNA intact spermatozoa by zona pellucida might be a significant mechanism at the gamete level as fertilization with zona free hamster oocytes did not reduce the incidence of chromosomal abnormalities in pronuclei [14]. In addition, it has been shown that zona pellucida can also act as selective barrier to aneuploid spermatozoa [15]. However, till date there is no report available on the differential zona binding competence of spermatozoa with respect to their sex chromosome and the extent of DNA damage carried by these spermatozoa.
The Y chromosome is particularly vulnerable to DNA damage, partly because of its genetic structure and partly because it cannot correct double-stranded DNA fragments by homologous recombination [16]. However, in somatic cells, high dose of radiation can immediately induce breakage of Y chromosome [17]. As the mature sperm has unique chromatin architecture and poor DNA repair capacity, the radiation doses used in the present study and a short post-irradiation interval of 18 h is not expected to target germ cell populations other than mature epididymal spermatozoa and eventually may not have any specific impact on X or Y bearing sperm. The reason for 18 h post-irradiation interval used in our study is to induce DNA fragmentation specifically in epididymal sperm (possibly ROS mediated) and not other types of abnormalities as prolonged post irradiation interval also increases a substantial risk of sperm aneuploidy due to meiotic errors and it is difficult to rule out the influence of other form of sperm abnormalities.
Although, the number of X and Y-chromosome sperm binding was significantly perturbed in relation to the extent of DNA damage, at this juncture it is not possible to explain the mechanism by how zona pellucida regulates the differential binding of spermatozoa. An earlier study has demonstrated the existence of differential functional survival of X and Y bearing spermatozoa where Y chromosome bearing spermatozoa had longer functional survival under in vitro conditions [15]. In contrary, our results have suggested the compromised zona binding ability of Y bearing spermatozoa possibly due to their higher susceptibility to undergo DNA fragmentation than X-bearing spermatozoa as suggested earlier [16].
Though the amount of DNA damage in the irradiated spermatozoa increased in a dose-dependent manner, the sperm functional characteristics were not significantly compromised (Table 1). In addition, the sex ratio was unskewed in the offspring derived from the DNA damaged sperm at birth [18] even though, an earlier study has demonstrated mid gestational demise of the fetus derived from the X-irradiated sperm [12]. Nonetheless, our early study also showed reduction in litter size in relation to sperm DNA damage level [18]. It has been shown that radiation induced genomic instability through the germ cells can be transmitted to the offspring [11, 13, 19] which can affect both somatic and germ cell compartments [5]. The period in which survival reduction occurred in our earlier study [18] was only until 10 weeks of postnatal development which roughly correspond to the rapidly growing phase in the life of mice. We speculate that the escape of X bearing spermatozoa at zona level subsequently resulted in the aberrant damage response pathways or increased genomic instability in fetus which eventually caused early postnatal death of the offspring. Radiation induced genomic instability describes the effects seen at delayed times several years later and is manifested for many cell divisions after the initial radiation insult [20].
The novel observation made in this study hypothesize that zona pellucida may have the ability to selectively prevent DNA damaged Y bearing but not X bearing sperm. Sex determination of the early embryos derived from the irradiated sperm may prove the role of zona pellucida in preventing fertilization by the DNA damaged Y bearing sperm. One of the limitations of this study was lack of simultaneous detection of DNA integrity and sex chromosome status of zona bound and unbound spermatozoa. Due to technical difficulties, several attempts to determine this relationship were not successful. More importantly, the exact mechanisms behind this unique and clinically important observation need to be elucidated further.
Electronic supplementary material
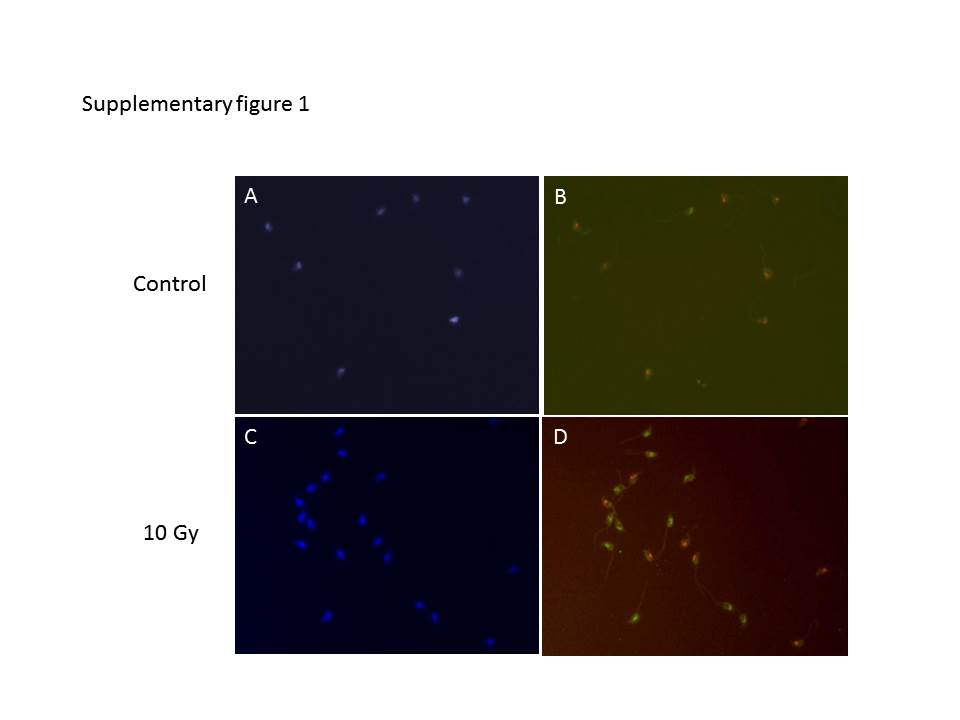{kind=link}
Fluorescent in situ hybridization (FISH) of zona pellucida bound X-bearing (green) and Y-bearing (red) spermatozoa. (JPEG 22 kb)
Acknowledgments
Part of this work was supported by Indian Council of Medical Research in the form of senior research fellowship to DU (IRISID No. 2006–01640). Authors are thankful to Radiation Oncology department for providing radiation facility.
Footnotes
Capsule
Differences in zona pellucida binding of mouse X and Y bearing spermatozoa are related to their DNA integrity.
References
- 1.Donnelly ET, O’Connell M, McClure N, Lewis SE. Differences in nuclear DNA fragmentation and mitochondrial integrity of semen and prepared human spermatozoa. Hum Reprod. 2000;15:1552–1561. doi: 10.1093/humrep/15.7.1552. [DOI] [PubMed] [Google Scholar]
- 2.Aitken RJ, De Iuliis GN. Origins and consequences of DNA damage in male germ cells. RBM Online. 2007;14:727–733. doi: 10.1016/s1472-6483(10)60676-1. [DOI] [PubMed] [Google Scholar]
- 3.Ahmadi A, Ng SC. Developmental capacity of damaged spermatozoa. Hum Reprod. 1999;14:2279–2285. doi: 10.1093/humrep/14.9.2279. [DOI] [PubMed] [Google Scholar]
- 4.Fatehi AN, Bevers MM, Schoevers E, Roelen BA, Colenbrander B, Gadella BM. DNA damage in bovine sperm does not block fertilization and early embryonic development but induces apoptosis after the first cleavages. J Androl. 2006;27:176–188. doi: 10.2164/jandrol.04152. [DOI] [PubMed] [Google Scholar]
- 5.Adiga SK, Upadhya D, Kalthur G, Bola Sadashiva SR, Kumar P. Transgenerational changes in somatic and germ line genetic integrity of first-generation offspring derived from the DNA damaged sperm. Fertil Steril. 2010;93:2486–2490. doi: 10.1016/j.fertnstert.2009.06.015. [DOI] [PubMed] [Google Scholar]
- 6.Van Dyk Q, Lanzendorf S, Kolm P, Hodgen GD, Mahony MC. Incidence of aneuploid spermatozoa from subfertile men: selected with motility versus hemizona-bound. Hum Reprod. 2000;15:1529–1536. doi: 10.1093/humrep/15.7.1529. [DOI] [PubMed] [Google Scholar]
- 7.Estop A, Catala V, Santalo J. Chromosome constitution of highly motile mouse sperm. Mol Reprod Dev. 1990;27:168–172. doi: 10.1002/mrd.1080270213. [DOI] [PubMed] [Google Scholar]
- 8.Hourcade JD, Pérez-Crespo M, Fernández-González R, Pintado B, Gutiérrez-Adán A. Selection against spermatozoa with fragmented DNA after postovulatory mating depends on the type of damage. Reprod Biol Endocrinol. 2010;8:9. doi: 10.1186/1477-7827-8-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sarrate Z, Anton E. Fluorescence in situ hybridization (FISH) protocol in human sperm. J Vis Exp. 2009;31:1405. doi: 10.3791/1405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Aitken RJ, De Iuliis GN. On the possible origins of DNA damage in human spermatozoa. Mol Hum Reprod. 2010;16:3–13. doi: 10.1093/molehr/gap059. [DOI] [PubMed] [Google Scholar]
- 11.Adiga SK, Toyoshima M, Shiraishi K, Shimura T, Takeda J, Taga M, et al. p21 provides stage specific DNA damage control to preimplantation embryos. Oncogene. 2007;26:6141–6149. doi: 10.1038/sj.onc.1210444. [DOI] [PubMed] [Google Scholar]
- 12.Shimura T, Toyoshima M, Taga M, Shiraishi K, Uematsu N, Inoue M, et al. The novel surveillance mechanism of the Trp53-dependent s-phase checkpoint ensures chromosome damage repair and preimplantation-stage development of mouse embryos fertilized with x-irradiated sperm. Radiat Res. 2002;158:735–742. doi: 10.1667/0033-7587(2002)158[0735:TNSMOT]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 13.Shimura T, Inoue M, Taga M, Shiraishi K, Uematsu N, Takei N, et al. Niwa O p53-dependent S-phase damage checkpoint and pronuclear cross talk in mouse zygotes with X-irradiated sperm. Mol Cell Biol. 2002;22:2220–2228. doi: 10.1128/MCB.22.7.2220-2228.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Martínez-Pasarell O, Vidal F, Colls P, Nogués C, Egozcue J, Templado C. Sex chromosome aneuploidy in sperm-derived pronuclei, motile sperm and unselected sperm, scored by three-color FISH. Cytogenet Cell Genet. 1997;78:27–30. doi: 10.1159/000134620. [DOI] [PubMed] [Google Scholar]
- 15.Van Dyk Q, Mahony MC, Hodgen GD. Differential binding of X- and Y-chromosome-bearing human spermatozoa to zona pellucida in vitro. Andrologia. 2001;33:199–205. doi: 10.1046/j.1439-0272.2001.00427.x. [DOI] [PubMed] [Google Scholar]
- 16.Aitken RJ, Krausz C. Oxidative stress, DNA damage and the Y chromosome. Reproduction. 2001;122:497–506. doi: 10.1530/rep.0.1220497. [DOI] [PubMed] [Google Scholar]
- 17.Mühlmann-Díaz MC, Bedford JS. Breakage of human chromosomes 4, 19 and Y in G0 cells immediately after exposure to gamma-rays. Int J Radiat Biol. 1994;65:165–173. doi: 10.1080/09553009414550201. [DOI] [PubMed] [Google Scholar]
- 18.Kumar D, Upadhya D, Salian SR, Rao SB, Kalthur G, Kumar P, et al. The extent of paternal sperm DNA damage influences early post-natal survival of first generation mouse offspring. Eur J Obstet Gynecol Reprod Biol. 2013;166:164–167. doi: 10.1016/j.ejogrb.2012.09.021. [DOI] [PubMed] [Google Scholar]
- 19.Niwa O. Induced genomic instability in irradiated germ cells and in the offspring; reconciling discrepancies among the human and animal studies. Oncogene. 2003;22:7078–7086. doi: 10.1038/sj.onc.1207037. [DOI] [PubMed] [Google Scholar]
- 20.Morgan WF. Non-targeted and delayed effects of exposure to ionizing radiation: II. Radiation-induced genomic instability and bystander effects in vivo, clastogenic factors and transgenerational effects. Radiat Res. 2003;159:581–596. doi: 10.1667/0033-7587(2003)159[0581:NADEOE]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fluorescent in situ hybridization (FISH) of zona pellucida bound X-bearing (green) and Y-bearing (red) spermatozoa. (JPEG 22 kb)