

Abstract
MC1R is recognized for its role in the regulation of melanin pigmentation. In addition, many investigators believe that it also plays a crucial role in immunomodulation (immunosuppression) and in melanogenesis-independent protective responses against ultraviolet radiation (UVR). Surprisingly Wolnicka-Glubisz et al have shown that loss of function in the MC1R has no effect on inflammatory responses and immunosuppression induced by UVR in C57BL/6 mice as well as on the degree of UVA-induced DNA damage in the epidermis and dermis. These findings, by challenging the existing dogmas on the precise role of MC1R in non-pigmentary responses to the UVR, mandate further research to either validate the presented data or to define to which degree these phenomena are restricted to the C57BL/6 mouse model or are applicable to other species including humans. The alternative target for immunomodulation is represented by MC3R. However, cutaneous expression of MC3R remains to be demonstrated.
Keywords: αMSH, UVR, MC1R, immunosuppression
Commentary
POMC derived peptides including αMSH and ACTH and the melanocortin receptor type 1 (MC1R) are recognized for their role in the regulation of melanin pigmentation (1, 2). In addition, αMSH plays an important role in immunomodulation including induction of anti-inflammatory responses with an assumption that that this pathway is mediated through activation of MC1R (3–5). However, in the recent review paper by Bohm & Grassel (3) it is already envisioned that MC3R is a target for mediation of anti-inflammatory effects by αMSH and related peptides in the osteoarticular system. It is also believed that αMSH activation of MC1R induces protective mechanisms against damaging effects of ultraviolet radiation (UVR) (6, 7).
Most recently, researchers from George Washington and Jagiellonian Universities, using C57BL/6-c, C57BL/6 and C57BL/6-Mc1re/e mouse strains, have made surprising finding that loss of function in the MC1R has neither impacted inflammatory responses to UV nor affected UVR induced immunosuppression (8). These findings are in striking contrast with generally accepted opinion that constitutive or ligand induced MC1R activity plays an important role in modulation of cutaneous immune activity in response to UVR (3–5). Interestingly, the authors also showed that UVR induced similar DNA damage in the epidermis and dermis irrespectively of the genetic background of these mouse strains. These surprising findings challenge the existing dogmas on the precise role of MC1R in non-pigmentary responses to the UVR (3–7) (Fig. 1).
Figure 1.
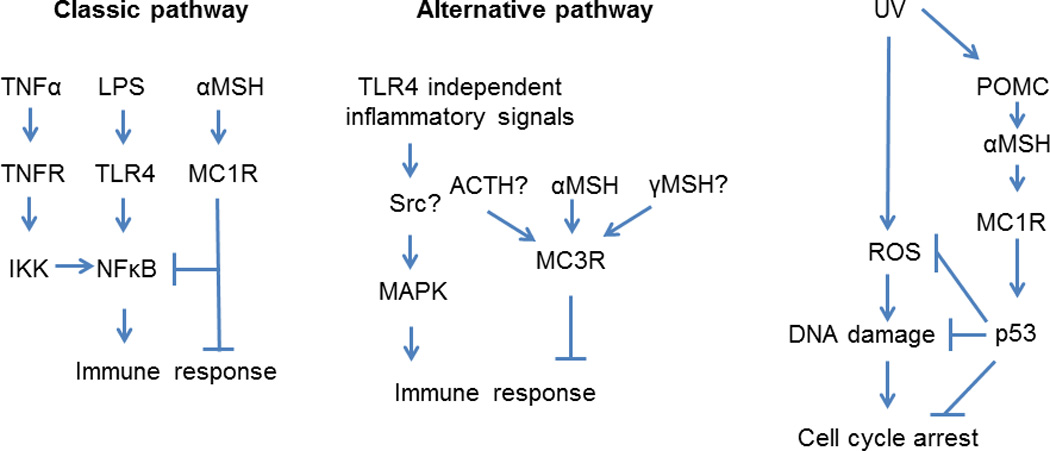
Classic and alternative pathways involved in UVR-induced non-pigmentary responses mediated by melanocortins.
These experiments also rise several questions. It was recently reported that effects of melanocortins on DNA repair and diminishing of oxidative stress caused by UV irradiation is mediated by MC1R and requires activity of p53 (7, 9). In addition, recent study has shown that MSH bound to the MC1R 1 activates adenylate cyclase activity with following activation of Xeroderma pigmentosum group A (XPA)-binding protein 1 and its nuclear translocation, a critical factor controlling nucleotide excision repair signaling pathways (10). Also, the presence of thymine dimer in UV irradiated skin might not be ideal proof of non-essential role of MC1R receptor in UV response. Therefore, it would be interesting to observe how the murine skin with selected MC1R phenotypes copes with DNA damage and somatic mutations. It must also be noted that polymorphism of MC1R is recognized as one of the skin cancer-risk factors (11).
Although the immunoregulatory role of αMSH is unquestionable (3–5), there are several studies showing that MC1R is not essential for immunomodulatory function of melanocortins. Getting et al (12) showed that the presence of fully functional MC1R receptor is not essential for inactivation of peritoneal macrophages by αMSH, but the effect was abrogated by selective MC3R/MC4R antagonist SHU9119, but not by the selective MC4R antagonist HS024 and, consequently MC3R agonist inhibited peritoneal macrophages. Other study showed that MC3R, but not MC1R is essential for abrogation of urate–crystal-activated inflammation in rat model of arthritis (13). Moreover, Cooper et al (14) showed that immunosuppressive effects of αMSH on streptokinase streptodornase-induced lymphocyte proliferation in human, was not dependent on MC1R allelic variations. They also postulated that presence of MC3R might be required for immunomodulatory activity of αMSH. In contrast Li and Taylor (15) showed that MC1R receptor is essential for effective inhibition of NO generation as well as TNFα production by α-MSH in lipopolysaccharides (LPS)-stimulated RAW264.7 macrophages. Nevertheless, authors also found that RAW264.7 macrophages express MC3R but not MC5R and suggested that MC3R might also be involved in anti-inflammatory response but through non-TLR pathways (15). The same cellular model was used to demonstrate that MC1R is essential for inhibition of LPS induced inflammation as well as 2-chloro-1,3,5-trinitrobenzene (TNCB)-induced atopic dermatitis model and this observation was confirmed by in vivo studies(16). Furthermore, Loser et al have convincingly demonstrated an important role of α-MSH and MC1R in MHC class I-restricted cytotoxicity (17).
Thus, striking observation by Wolnicka-Glubisz et al (8) can be explained by involvement of other members of melanocortin receptor family, e.g., MC3R or perhaps MC5R. MC3R is thought to interact mainly with γMSH with EC50 of 7 nM, but according to the binding efficiency studies αMSH and ACTH could bind to MC3R with comparable affinity with EC50 at 1 and 6 nM, respectively (See (3) for review). MC3R may be involved in activation of steroidogenesis (18) and food intake, but interestingly, Mc3−/− mice show selective upregulation of IL-1β, IL-6, and Nos2 when compared to wt mice (19). Thus, activation of MC3R may represent an alternative pathway for melanocortin induced immunosuppression (Fig. 1). However, cutaneous expression of MC3R remains to be demonstrated.
As discussed by Li and Taylor (15) activation of anti-inflammatory response by melanocortins might depend on the specificity of induction inflammatory factors and MC1R may be essential for attenuation of LPS-induced, TLR4-activated immune responses, while inhibition of urate–crystal-activated inflammation requires MC3R activity and is not affected by MC1R downregulation. It has to be underlined that the observed effects might also reflect cell type or organism specific differences in the regulation of immune response or genetic variations. In fact, the authors analyzed the selected population of CD11b+Ly6G+ cells (8). Also the C57BL/6 background may not be ideal to extrapolate the findings to other rodent species. For example, while the important role of POMC derived peptides in regulation of melanin pigmentation is demonstrated in vast majority of rodent species (1, 20), the POMC−/− C57BL/6 mice produce solely eumelanin pigment because of recessive (a/a) genetic background (21). Finally, there are significant difference in human and rodent (nocturnal specie) skin, which includes differences in local neuropeptide signaling (2, 22). Thus, further research is necessary to validate the presented data or to determine to which degree these phenomena extend beyond the C57BL/6 mouse model.
Taken together, recently published studies including Wolnicka-Glubisz et al paper (8), brought additional level of complexity to the model of αMSH regulatory functions on immune responses indicating that MC1R might not be its only target in the skin and implying MC3R as an additional target triggering alternative pathways leading to immunosuppression (Fig. 1). Furthermore, Wolnicka-Glubisz et al findings (8) challenge the existing dogma that MC1R is crucial and/or very important in non-pigmentary responses to the UVR, which should stimulate further studies in this area.
Acknowledgement
Writing of this commentary was supported in part by grants from National Science Foundation (# IOS-0918934), National Institutes of Health (#1R01AR056666-01A2) to AS, and a grant from Polish Ministry of Science and Higher Education, project no. N405 623238 to M.A.Z. and A.S.. Both authors contributed equally by writing the paper.
Footnotes
Conflicts of Interest
There are no conflicts of interest to declare for all the authors
References
- 1.Slominski A, Tobin DJ, Shibahara S, Wortsman J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol Rev. 2004;84:1155–1228. doi: 10.1152/physrev.00044.2003. [DOI] [PubMed] [Google Scholar]
- 2.Slominski AT, Zmijewski MA, Skobowiat C, Zbytek B, Slominski RM, Steketee JD. Sensing the environment: regulation of local and global homeostasis by the skin's neuroendocrine system. Advances in anatomy, embryology, and cell biology. 2012;212:v–vii. 1–115. doi: 10.1007/978-3-642-19683-6_1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Böhm M, Grässel S. Role of proopiomelanocortin-derived peptides and their receptors in the osteoarticular system: from basic to translational research. Endocr Rev. 2012;33:623–651. doi: 10.1210/er.2011-1016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Brzoska T, Luger TA, Maaser C, Abels C, Bohm M. Alpha-melanocyte-stimulating hormone and related tripeptides: biochemistry, antiinflammatory and protective effects in vitro and in vivo, and future perspectives for the treatment of immune-mediated inflammatory diseases. Endocr Rev. 2008;29:581–602. doi: 10.1210/er.2007-0027. [DOI] [PubMed] [Google Scholar]
- 5.Luger TA, Scholzen T, Brzoska T, Becher E, Slominski A, Paus R. Cutaneous immunomodulation and coordination of skin stress responses by alpha-melanocyte-stimulating hormone. Annals of the New York Academy of Sciences. 1998;840:381–394. doi: 10.1111/j.1749-6632.1998.tb09577.x. [DOI] [PubMed] [Google Scholar]
- 6.Abdel-Malek ZA, Knittel J, Kadekaro AL, Swope VB, Starner R. The melanocortin 1 receptor and the UV response of human melanocytes--a shift in paradigm. Photochemistry and photobiology. 2008;84:501–508. doi: 10.1111/j.1751-1097.2008.00294.x. [DOI] [PubMed] [Google Scholar]
- 7.Kadekaro AL, Chen J, Yang J, et al. Alpha-melanocyte-stimulating hormone suppresses oxidative stress through a p53-mediated signaling pathway in human melanocytes. Mol Cancer Res. 2012;10:778–786. doi: 10.1158/1541-7786.MCR-11-0436. [DOI] [PubMed] [Google Scholar]
- 8.Wolnicka-Glubisz A, De Fabo E, Noonan F. Functional melanocortin 1 receptor Mc1r is not necessary for an inflammatory response to UV radiation in adult mouse skin. Experimental Dermatology. 2013 doi: 10.1111/exd.12100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wong SS, Ainger SA, Leonard JH, Sturm RA. MC1R variant allele effects on UVR-induced phosphorylation of p38, p53, and DDB2 repair protein responses in melanocytic cells in culture. J Invest Dermatol. 2012;132:1452–1461. doi: 10.1038/jid.2011.473. [DOI] [PubMed] [Google Scholar]
- 10.Dong L, Wen J, Pier E, et al. Melanocyte-stimulating hormone directly enhances UV-Induced DNA repair in keratinocytes by a xeroderma pigmentosum group A-dependent mechanism. Cancer research. 2010;70:3547–3556. doi: 10.1158/0008-5472.CAN-09-4596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Law MH, Macgregor S, Hayward NK. Melanoma genetics: recent findings take us beyond well-traveled pathways. J Invest Dermatol. 2012;132:1763–1774. doi: 10.1038/jid.2012.75. [DOI] [PubMed] [Google Scholar]
- 12.Getting SJ, Christian HC, Lam CW, et al. Redundancy of a functional melanocortin 1 receptor in the anti-inflammatory actions of melanocortin peptides: studies in the recessive yellow (e/e) mouse suggest an important role for melanocortin 3 receptor. J Immunol. 2003;170:3323–3330. doi: 10.4049/jimmunol.170.6.3323. [DOI] [PubMed] [Google Scholar]
- 13.Getting SJ, Lam CW, Chen AS, Grieco P, Perretti M. Melanocortin 3 receptors control crystal-induced inflammation. FASEB J. 2006;20:2234–2241. doi: 10.1096/fj.06-6339com. [DOI] [PubMed] [Google Scholar]
- 14.Cooper A, Robinson SJ, Pickard C, Jackson CL, Friedmann PS, Healy E. Alpha-melanocyte-stimulating hormone suppresses antigen-induced lymphocyte proliferation in humans independently of melanocortin 1 receptor gene status. J Immunol. 2005;175:4806–4813. doi: 10.4049/jimmunol.175.7.4806. [DOI] [PubMed] [Google Scholar]
- 15.Li D, Taylor AW. Diminishment of alpha-MSH anti-inflammatory activity in MC1r siRNA-transfected RAW264.7 macrophages. J Leukoc Biol. 2008;84:191–198. doi: 10.1189/jlb.0707463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chen W, Li J, Qu H, et al. The melanocortin 1 receptor (MC1R) inhibits the inflammatory response in Raw 264.7 cells and atopic dermatitis (AD) mouse model. Mol Biol Rep. 2013;40:1987–1996. doi: 10.1007/s11033-012-2256-x. [DOI] [PubMed] [Google Scholar]
- 17.Loser K, Brzoska T, Oji V, et al. The neuropeptide alpha-melanocyte-stimulating hormone is critically involved in the development of cytotoxic CD8+ T cells in mice and humans. PloS one. 2010;5:e8958. doi: 10.1371/journal.pone.0008958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Harmer SC, Bicknell AB. Role of gamma-MSH peptides in the regulation of adrenal steroidogenesis. Peptides. 2005;26:1944–1951. doi: 10.1016/j.peptides.2004.11.028. [DOI] [PubMed] [Google Scholar]
- 19.Patel HB, Bombardieri M, Sampaio AL, et al. Anti-inflammatory and antiosteoclastogenesis properties of endogenous melanocortin receptor type 3 in experimental arthritis. FASEB J. 2010;24:4835–4843. doi: 10.1096/fj.10-167759. [DOI] [PubMed] [Google Scholar]
- 20.Slominski A, Wortsman J, Luger T, Paus R, Solomon S. Corticotropin releasing hormone and proopiomelanocortin involvement in the cutaneous response to stress. Physiol Rev. 2000;80:979–1020. doi: 10.1152/physrev.2000.80.3.979. [DOI] [PubMed] [Google Scholar]
- 21.Slominski A, Plonka PM, Pisarchik A, et al. Preservation of eumelanin hair pigmentation in proopiomelanocortin-deficient mice on a nonagouti (a/a) genetic background. Endocrinology. 2005;146:1245–1253. doi: 10.1210/en.2004-0733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Slominski A, Wortsman J, Pisarchik A, et al. Cutaneous expression of corticotropin-releasing hormone (CRH), urocortin, and CRH receptors. FASEB J. 2001;15:1678–1693. doi: 10.1096/fj.00-0850rev. [DOI] [PubMed] [Google Scholar]