

Abstract
Purpose:
The proposed project is aimed at enhancing the efficiency of epithelial ovarian cancer treatment and reducing adverse side effects of chemotherapy using nanotechnology. Overexpression of the CD44 membrane receptor results in tumor initiation, growth, tumor stem cells specific behavior, development of drug resistance, and metastases. We hypothesize that a developed cancer targeted delivery system which combines CD44 siRNA with paclitaxel would successfully deliver its payload inside cancer cells, effectively induce cell death, and prevent metastases.
Experimental Design:
We synthesized, characterized, and tested a nanoscale-based drug delivery system containing a modified Polypropylenimine (PPI) dendrimer as a carrier; anticancer drug paclitaxel as a cell death inducer; a synthetic analog of luteinizing hormone-releasing hormone (LHRH) peptide as a tumor targeting moiety, and siRNA targeted to CD44 mRNA. The proposed NDDS was tested in vitro and in vivo using metastatic ovarian cancer cells isolated from patients with malignant ascites.
Results:
We found that in contrast to cells isolated from primary tumors, CD44 was highly overexpressed in metastatic cancer cells. Treatment with the proposed tumor-targeted nanoscale-based nucleic acid and drug delivery system led to the suppression of CD44 mRNA and protein, efficient induction of cell death, effective tumor shrinkage, and prevention of adverse side effects on healthy organs.
Conclusion:
We show a high therapeutic potential for combinatorial treatment of ovarian carcinoma with a novel drug delivery system that effectively transports siRNA targeting to CD44 mRNA simultaneously with cytotoxic agents.
Keywords: Nanotechnology, Gene Therapy, Dendrimer, LHRH, Cancer Stem Cells, Paclitaxel
Introduction
Ovarian cancer is the fifth leading cause of cancer-related deaths in women and the most lethal of all gynecologic cancers in the United States. In 2011, an estimated 21,990 new cases of ovarian cancer were registered out of which 15,460 cases were considered deadly (1). In 75% of the cases, the disease is already present as tumor implants throughout the peritoneal cavity (Stage III and higher) at diagnosis (2). Although initial chemotherapy against ovarian cancer is usually effective, the five year survival rates for patients with advanced Stage IIIC disease are extremely low at approximately 30% (2). Consequently, novel approaches to chemotherapy for advanced or recurrent ovarian cancer is very important in order to increase the survival rate.
CD44 is a cell surface glycoprotein involved in cell-cell interactions, cell adhesion, and migration (3). It is considered a major cell surface marker for metastasis and progression in certain types of cancers, including ovarian carcinoma. CD44 is a receptor for hyaluronic acid (4, 5). Hyaluronic acid is the principal glycosaminoglycan found in extracellular matrices. It is a major component of the peritoneum which is considered a major site for ovarian cancer metastases (4, 6). CD44 interacts with hyaluronic acid to activate Nanog-Stat-3 and ankyrin based signaling pathways. Activation of these signaling pathways is considered to be responsible for conferring the tumor stem cell specific behavior like transcriptional activation, tumor cell growth, and multidrug resistance in ovarian and breast cancers (7). CD44 is not expressed in normal cells and certain non-invasive established cell lines (8-11) and interactions of CD44 with tumor stroma and tumor microenvironment are closely related to the metastatic growth of cancer (10, 12). Based on the aforementioned findings, we hypothesized that suppression of CD44 protein could enhance the efficiency of chemotherapy and prevent the development of metastases. Such suppression can potentially be achieved by employing siRNA targeted to CD44 mRNA. However, the poor ability of siRNA to penetrate across the cell membrane into the cytoplasm limits its efficiency as a suppressor of targeted mRNA and protein. Therefore, several types of non-viral delivery vectors were developed in order to enhance the efficiency of siRNA cellular penetration and poor stability in the plasma (13-24). In order to verify the hypothesis, in the present study, we constructed a multifunctional nanocarrier-based drug delivery system (DDS) containing a modified Polypropylenimine (PPI) dendrimer as a carrier; anticancer drug paclitaxel as a cell death inducer; a synthetic analog of luteinizing hormone-releasing hormone (LHRH) peptide as a tumor targeting moiety, and siRNA targeted to CD44 mRNA. The system was capable of simultaneous delivering of an anticancer drug and siRNA specifically to ovarian cancer cells. This DDS was tested in vitro on cells isolated from malignant ascites obtained from patients with advanced ovarian carcinoma and also in vivo on a murine xenograft model of human ovarian carcinoma initiated by subcutaneous injection of tumor cells into nude mice.
Materials and Methods
Materials
Dimethyl-3-3′-dithiobispropionimidate-HCl (DTBP) was obtained from Thermo Fisher Scientific Inc. (Rockford, IL). Polypropylenimine (PPI) tetrahexacontaamine dendrimer was obtained from Symo Chem (Eindhoven, the Netherlands), α-maleimide-ω-N-hydroxysuccinimide ester poly(ethylene glycol) (MAL–PEG–NHS, MW 5000 Da) was purchased from NOF Corporation (White Plains, NY). Synthetic analog of luteinizing hormone-releasing hormone (LHRH) decapeptide (Gln-His-Trp-Ser-Tyr-DLys(D-Cys)-Leu-Arg-Pro) was synthesized according to our design by the American Peptide Company, Inc. (Sunnyvale, CA). Fluorescent RNA duplex, siRNA labeled with Pierce NuLight DY-547 fluorophores (siGLO Red Transfection Indicator, red fluorescence), was also obtained from Applied Biosystems (Ambion, Inc., Foster City, CA). The primary rat CD44 antibody against human was obtained from Developmental studies at hybridoma bank (University of Iowa, Iowa). The secondary anti-rat conjugated with Cy3® goat antibody was obtained from Invitrogen (Eugene, Oregon). CD44 siRNA with a sequence of sense 5'-UAUUCCACGUGGAGAAAAAtt-3' and antisense 5'-UUUUUCUCCACGUGGAAUAca-3' was obtained from Applied Biosystems (Ambion, Inc., Foster City, CA). All other reagents were purchased from Sigma-Aldrich Co. LLC (St. Louis, MO) and used without modifications. Discarded anonymous pathological materials (primary solid gynecologic tumors and malignant ascites) were provided by the Cancer Institute of New Jersey. The samples did not allow for identifying patient information.
Synthesis of Paclitaxel - Succinic Acid Conjugate
Succinic acid as a bis(carboxylic acid moiety) was conjugated with the hydroxyl group in paclitaxel (1 equiv.), leaving another carboxylic group free for further modifications. The flask was charged with paclitaxel (250.0 mg, 0.29 mmol), succinic acid (SA, 34.6 mg, 0.29 mmol) and 4-Dimethylaminopyridine (DMAP, 10.0 mg, 0.08 mmol) in 5.0 mL of anhydrous dimethyl sulfoxide (DMSO) and 20.0 mL of anhydrous CH2Cl2. The reaction mixture was stirred for 30 min at room temperature and finally N-(3-dimethylaminopropyl)-N-ethylcarbodiimide HCl (EDC·HCl, 57.51 mg, 0.29 mmol) was added. The reaction was carried out with continuous stirring for 24 h at room temperature. The resulting reaction mixture turned light yellow due to the formation of dicyclohexylurea (DCU) as a byproduct. Paclitaxel-succinic acid conjugate (paclitaxel-SA) was precipitated using diethyl ether and dried under a vacuum. To remove unreacted paclitaxel, the crude was purified by gel column chromatography.
Synthesis of Paclitaxel – PPI Conjugate
Reaction was performed in a similar condition as the synthesis of paclitaxel - succinic acid conjugate. Paclitaxel was conjugated to PPI dendrimer at 1:1 molar ratio as previously described (24, 25). Briefly, the flask was charged with paclitaxel-SA (15.3 mg, 0.016 mmol), PPI (115.1 mg, 0.016 mmol) and DMAP (1.0 mg, 0.001 mmol) in 1.0 mL of anhydrous DMSO and 7.0 mL of anhydrous CH2Cl2. The reaction mixture was stirred for 30 min at room temperature and finally 3.1 mg of EDC·HCl were added. The reaction was carried out with continuous stirring for 24 h at room temperature. The resulting reaction mixture turned light yellow due to the formation of DCU as a byproduct. The reaction mixture (cloudy) was filtered to remove DCU. Paclitaxel-SA-PPI conjugate was precipitated using diethyl ether and dried under a vacuum. To remove unreacted paclitaxel-SA, the crude was purified by a dialysis membrane. The final product was characterized and confirmed by proton NMR and MALDI.
Modification of Paclitaxel – PPI Conjugate with MAL–PEG–NHS and LHRH
Paclitaxel – PPI conjugates dissolved in 5 mM HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid) buffer (pH=7.2) at the concentration 5 μM was mixed with MAL-PEG-NHS with PEG: NH2 ratio equal to 2 rel. units. The NHS groups on the distal end of PEG reacted with amine groups on the periphery of Paclitaxel – PPI conjugate. After the reaction within 1 h at room temperature, LHRH peptide was added to the solution to covalently conjugate the peptide on the distal end of the PEG layer through the maleimide groups on the PEG and the thiol groups in LHRH (LHRH : PEG = 2). After 12 h of the reaction, the modified complexes were then purified by dialysis (MW cut-off 10 kDa) against water for 24 h and used for further studies.
Preparation of PPI – siRNA complexes
PPI-siRNA complexes have been prepared and modified according to our previously developed procedure (24). Briefly, the siRNA complexes were prepared at an amine/phosphate (N/P) ratio of 2.4 in 5 mM HEPES buffer (pH 8.0) by adding stock solution of PPI dendrimer into a pre-prepared siRNA solution. The final concentrations of siRNA and PPI in the solution were 28.6 μM and 45 μM, respectively. The samples were vortexed briefly, and the solutions were then incubated at room temperature for 30 min to ensure complex formation. In order to cross-link individual complexes, DTBP dissolved in 5 mM HEPES buffer was added to the above-formulated PPI –siRNA complexes solution at DTBP:NH2=3.2 cross-linking ratio (ratio between DTBP and total amino groups of PPI available after being complexed with siRNAs). After 3 h of reaction, pH of the solution was adjusted to 7.2, followed by the addition of MAL–PEG–NHS (PEG: NH2= 2). The NHS groups on the distal end of PEG reacted with amine groups on the periphery of PPI –siRNA complex. After the reaction within 1 h at room temperature, LHRH peptide was added to the solution to covalently conjugate the peptide on the distal end of the PEG layer through the maleimide groups on the PEG and the thiol groups in LHRH (LHRH : PEG = 2). After 12 h of the reaction, the modified complexes were then purified by dialysis (MW cut-off 10 kDa) against water for 24 h and used for further studies.
Study of Complex Formation Between siRNA and Dendrimers
In order to evaluate the formation of complexes between siRNA and PPI dendrimers, ethidium bromide (EtBr), and agarose gel retardation assays were used. Various solutions of siRNA with dendrimers were prepared. The N/P ratios in the solutions were 0, 0.25, 0.5, 0.75, 1, 1.25, 1.5, 1.75, 2, 2.25, 2.5, 2.75 and 3. The concentration of siRNA was kept constant at 0.4 μM while the concentration of dendrimer was increased to achieve the aforementioned N/P ratios. EtBr was added to all solutions in the concentration of 126 μM. After 30 min reaction, the fluorescence intensity at excitation wavelength of 526 nm and emission at 592 nm was measured using Hitachi F 7000 Fluorescence Spectrophotometer (Tokyo, Japan). Reduced fluorescence (Ft/F0) was determined for each sample where Ft was the fluorescence of the dendrimer and siRNA complexes and F0 was the fluorescence of the siRNA alone (26). Agarose gel retardation assay of the above samples were also carried out. 15 μl of the complex solution was mixed with 3 μl of gel loading dye (New England Biolabs, Ipswich, MA). These mixtures were then loaded on 4% Nu Sieve Agarose gel (Lonza, Rockland, ME) and allowed to run 1.5 h at 96V.
MALDI TOF Mass Spectroscopy, Atomic Force Microscopy, Particle Size, Zeta Potential, Stability of the Complexes in Serum, Cellular Internalization, Cell Invasion Assay, In Vitro Cytotoxicity
Previously described methods (18-20, 22, 24, 25, 27-29) were used for the analysis of these parameters (please see details in Supplementary Data).
siRNA Serum Stability
Serum stability of both free siRNA and modified siRNA was measured as previously described (24). Briefly, naked siRNA and PPI-siRNA complexes were incubated at 37 °C with equal volume of human serum to give 50% serum concentration. At each predetermined time interval, (0, 5, 15, 30min, 1, 2, 3, 4, 5, 6, 7, 24 and 48 h) 50 μL of the mixture were removed and stored at −20°C until gel electrophoresis was performed. The aliquots from different incubation time periods were subjected to submarine electrophoresis. The gels were stained with EtBr, digitally photographed, and scanned using Gel Documentation System 920 (NucleoTech, San Mateo, CA).
Cancer Cells and Animal Model
The ascitic fluid with cancer cells was obtained from the peritoneum area of the patients with ovarian cancer provided by the Cancer Institute of New Jersey. The samples were centrifuged for 20 min at 2000 g; the supernatant was discarded and cell pellets were consequently re-suspended. The re-suspended cells were cultured in RPMI 1640 media with L-glutamine (Lonza, Walkersvile, MD) supplemented with 15% fetal bovine serum (Invitrogen, Carlsbad, CA), 2.5 μg/ml insulin (Sigma, St. Louis, MO) and 1.2 mL/100 mL penicillin-streptomycin (Gibco, Grand Island, NY). Cells were grown at 37 °C in a humidified atmosphere of 5% CO2 (v/v) in air. All of the experiments were performed on the cells in exponential growth phase. An animal model of human ovarian carcinoma xenografts was created as previously described (30-32). Briefly, human ascitic cells (2x106) were subcutaneously transplanted into the flanks of female athymic nu/nu mice. When the tumors reached a size of about 0.4 cm3 (15-20 days after transplantation), mice were treated intraperitoneally with saline (control) and different formulations: Dendrimer (PPI), LHRH, PPI-Scrambled siRNA, Paclitaxel (TAX), PPI-CD44 siRNA, PPI-TAX, LHRH-CD44 siRNA, LHRH-PPI-TAX, LHRH-PPI-TAX mixed with LHRH-PPI-CD44 siRNA. The dose of TAX (2.5 mg/kg) in all drug-containing formulations corresponded to the maximum tolerated dose of this drug estimated in separate experiments as previously described (31-33). The animals were treated twice per week eight times within four weeks and tumor size was measured by a caliper. The weight of tumors was also measured at the end of the experiment after euthanizing the animals. Animal weight was evaluated every day during the treatment period. All other measurements were performed 48 h after the treatment. Changes in tumor size were used as an overall marker for antitumor activity. Biodistribution of non-targeted and LHRH-targeted delivery systems was estimated in tumor and different organs (liver, kidney, spleen, heart, lung, and brain) as previously described (24, 33-37). Details are provided in the Supplementary Data section. All animal experiments were carried out according to the approved protocol and Institutional guidance.
LHRH Expression
The expression of genes encoding luteinizing hormone releasing hormone receptors (LHRHR) and β2-microglobulin (β2-m, internal standard) in cell and tissue homogenates was measured by Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR) as previously described (33). The following pairs of primers were used: LHRHR – GAC CTT GTC TGG AAA GAT CC (sense), CAG GCT GAT CAC CAC CAT CA (antisense); β2-m – ACC CCC ACT GAA AAA GAT GA (sense), ATC TTC AAA CCT CCA TGA TG (antisense). Gene expression was calculated as the ratio of analyzed RT-PCR product to the internal standard (β2-m). The expression of LHRHR gene was measured in human ovarian (primary solid tumor and malignant ascites), fallopian and endometrial tumors and different healthy visceral human organs (liver, kidney, spleen, heart, lung, brain, thymus and skeletal muscle). cDNA isolated from human organs was purchased from Clontech Laboratories, Inc (Mountain View, CA).
CD44 and MDR1 Expression
The expression of CD44 and MDR1 mRNA was measured by Quantitative Reverse Transcriptase-Polymerase Chain Reaction (QRT-PCR). RNA was isolated using an RNeasy kit (Qiagen, Valencia, CA) for in vitro and in vivo studies. Beta actin, beta-2-microglobulin and glyceraldehyde-3-phosphate dehydrogenase were used as internal standards. First-strand cDNA was synthesized using High Capacity RNA-to-cDNA kit (Applied biosystems, Carlsbad, CA) using Veriti 96 well thermal cycler (Applied Biosystems, Carlsbad, CA). After synthesis, the reaction mixture was subjected to quantitative PCR, which was carried out by Step One Plus Real time PCR system (Applied Biosystems, Carlsbad, CA). The pairs of primers for CD44, MDR1 and β-actin were obtained from Applied Biosystems (Carlsbad, CA). The PCR regimen was: 94°C/30 s, 55°C/1 min, 72°C/1 min for 30 cycles. A modified immunocytochemistry method (38) was used to analyze the expression of CD44 protein. Briefly, 1·105 cells were seeded to a 4 well chambered glass slide and allowed to incubate overnight with analyzed substances. Control cells received fresh media alone. The cells were then fixed with 4 % formaldehyde solution for 45 min. The fixed cells were washed thrice with PBS and then incubated with 5% bovine serum albumin (BSA) in PBS for 20 min to suppress nonspecific binding of IgG. After another washing with PBS, the cells were incubated with anti-CD44 antibodies (570 ng/mL, Hermes, University of Iowa, Iowa City, IA) in 5% BSA in PBS for 1 h followed by 45 min incubation with Cy3®-conjugated secondary antibody (2.5 μg/mL, Goat, Invitrogen, Eugene, OR) in PBS with 5% BSA. The labeled cells were analyzed by a fluorescence microscope (Olympus America Inc., Melville, NY).
Immunohistochemistry
To visualize the expression of CD44 protein in vitro in tumor tissues after various treatments, immunehistochemical staining was conducted on paraffin-embedded slides. At the end of the experiments, the animals were euthanized, tumors were extracted, immediately fixed in 10% phosphate-buffered formalin. Samples were subsequently dehydrated and embedded in Paraplast®. Slides (5 μm) were deparaffinized in xylene for 5 min followed by progressive rehydration in 100%, 95%, 70%, and 50% ethanol for 3 min during each step. The slides were subjected to 10 mM citrate buffer, pH 6.0 for 5 minutes in a microwave at a power of 300 watt, washed twice with PBS and then incubated with 5% Bovine Serum Albumin (BSA) in PBS for 20 minutes to suppress nonspecific binding of IgG. Tissue slides were washed twice with PBS for 5 minutes. The cells and tissue slides were incubated with anti-CD44 (570 ng/mL, Hermes, University of Iowa, Iowa City, IA) in 5% BSA in PBS for 1 h followed by 45 minute incubation with Cy3® fluorophore- conjugated secondary antibody (2.5 μg/mL, Goat, Invitrogen, Eugene, OR) in PBS with 5% BSA. The nuclei were stained with 600 nM of 4, 6 diamidino-2-phenylindole (DAPI) for 8 minutes. The labeled slides were analyzed by a fluorescence microscope (Olympus America Inc., Melville, NY). Red color represented CD44 protein; blue color represented cell nuclei.
Apoptosis Detection
Apoptosis induction in the tumor and other organs (liver, kidney, spleen, heart, lung and brain) was measured using the Cell Death Plus ELISA and TUNEL kits (F. Hoffmann-La Roche Ltd, Nutley, NJ) as previously described (34, 35, 39). For TUNEL assay, tissue slides were subjected to deparaffinization, rehydration, peroxidase blocking and antigen retrieval as described above. TUNEL reaction mixture (50 μL) was added to the sections. The slides were incubated in the dark for 60 min at 37 °C in a humidified atmosphere, washed with PBS, and mounted with cover slip using fluoromount mounting medium (Sigma, St Louis, MO). The sections were then analyzed using a fluorescence microscope (Olympus America Inc., Melville, NY). Another method of apoptosis detection - the Cell Death ELISA assay is based on the measurement of the enrichment of histone-associated DNA fragments (mononucleosomes and oligonucleosomes). Tumor and other tissues were homogenized and anti-histone and anti-DNA antibodies were used according to manufacturer instructions.
Statistical Analysis
Data were analyzed using descriptive statistics and single-factor ANOVA, and are presented as a mean ± SD from five to ten independent measurements. Ten animals were used in each experimental group. The power analysis (alpha=0.05; power=80%) based on the preliminary results showed that ten mice per group are needed for statistical analysis to determine a 15% difference between the groups. We analyzed data sets for significance with Student’s t test and considered P values of less than 0.05 as statistically significant.
Results
Synthesis and Characterization of Cytotoxic Tumor Targeted siRNA Nanocomplexes
In order to synthesize a cytotoxic tumor-targeted nanocarrier, an anticancer drug (cytotoxic agent), paclitaxel (TAX), was conjugated to the dendrimer via a biodegradable succinic acid spacer while a cancer targeting moiety, and the LHRH peptide, was bound to the distal end of non-biodegradable PEG polymer that surrounds the DTBP-caged dendrimer (Fig. 1A). Since endosomes and lysosomes show acidic pH (5.6–6.5), the incorporation of a pH sensitive ester bond between drug and carrier enables the specific release of TAX from the carrier in cancer cells. The MALDI TOF mass spectrometry confirmed that TAX was indeed conjugated to the dendrimer with 7162 of mass/charge (m/z) ratio resulting in the strong peak of the subsequent complex with Na at 8122 m/z (Fig. 1B). The complex formation between the synthesized PPI carriers and siRNA were studied by ethidium bromide (EtBr) and agarose gel retardation assays (Fig.1C). It was found that when the N/P ratio exceeded 0.5 related units, all used siRNA was completely conjugated to the carrier(s) resulting in disappearing of a small band of naked siRNA on gel electrophoresis. At the same time, binding of siRNA to the dendrimer carrier at N/P>0.5 led to the quenching of EtBr fluorescence (Fig. 1C). The nanoparticles with N/P ratio equal to 2.4 were used in all studies. AFM data (Fig. 1D) showed that the synthesized carrier condensated siRNA into stable compact nanoparticles with average diameter of 100 - 200 nm. The size of nanocarriers was also confirmed by light scattering method. The size of nanoparticles did not change significantly after incubation in human serum. The surface charge of initially synthesized dendrimers was +20.8 ± 4.01 mV. After making a complex of the PPI dendrimer with siRNA, caging with DTBP and PEGylation, the nanoparticles became practically neutral with an average surface charge of +1.10 ± 1.54 mV. Incubation of naked siRNA and PPI-siRNA complexes in 50% human serum at 37 °C demonstrated that the condensation of siRNA and dendrimers into nanoparticles, caging and PEGylation significantly increased the stability of siRNA. We found that the band of naked siRNA on the agarose gel almost completely disappeared after 6-7 h of incubation reflecting the degradation of siRNA. In contrast, the siRNA-PPI band did not decrease its intensity even after 48 h of incubation (Fig. 1D). These data indicate an increase in the stability of siRNA after conjugation of PPI dendrimer carriers.
Fig. 1.

Complex cancer targeted drug delivery system. A, Schema of the delivery system containing poly(propylenimine) (PPI) dendrimer as a carrier; paclitaxel (TAX) as an anticancer drug, conjugated to the dendrimer via succinic acid (SA) spacer; luteinizing-hormone-releasing hormone (LHRH) as a cancer cell targeting moiety, conjugated to the dendrimer via poly(ethylene glycol) (PEG) spacer; siRNA targeted to CD44 mRNA and caged by Dimethyl 3,3′-dithio-bis(propionimidate) dihydrochloride (DTBP). B, Representative image of matrix-assisted laser desorption/ionisation-time of flight (MALDI-TOF) mass spectrometry analysis of PPI-TAX conjugate. The most abundant peak [M+Na] was observed at mass M=8,122 indicating that paclitaxel was conjugated with the PPI dendrimer (M=7162). C, Representative gel electrophoresis image of siRNA complexated with PPI dendrimer at different nitrogen to phosphate (N/P). The complexes were synthesized with 0.4 μM siRNA and 126 μM ethidium bromide. The concentration of PPI dendrimer was increased in order to obtain N/P ratios from 0 (no PPI dendrimer) to 3. The nanoparticles with N/P ratio equal to 2.4 were used in all studies. D, Atomic force microscope (AFM) images of nanoparticles resulted from the complex formation between the carrier and siRNA and quantitative analysis of the stability of naked siRNA and siRNA-PPI complexes in plasma.
Expression of CD44, MDR1 and LHRH in Cancer Cells
In order to effectively deliver the drugs to the cell, the tumor must express the LHRH receptor to be identified by the target peptide. Fig. 2 shows that CD44 mRNA and protein were overexpressed in primary and metastatic tumor tissues isolated from patients with different types of gynecological cancers and in cancer cells isolated from malignant ascites (Fig. 2A and B). The expression of CD44 mRNA in normal ovarian tissues was significantly less pronounced when compared with tissues from primary and metastatic tumors of the same patient (Fig. 2A). Moreover, the expression was significantly higher in metastatic tissues when compared with primary tumors (Fig. 2A, P < 0.05). The expression of CD44 mRNA correlates with the expression of MDR1 mRNA (Fig. 2B). Fig. 2C shows that endometrial, fallopian, ovarian (primary and metastatic) tumors overexpressed LHRH receptors; in contrast, in the healthy liver, kidney, spleen, heart and lung tissues, detectable levels of LHRHR were not observed. Staining with anti-CD44 antibodies followed by a fluorescence microscopy analysis showed that the overexpressed CD44 proteins were localized predominately in the plasma membrane and partially in the cytoplasm of cancer cells (Fig. 2D). Ovarian cancer metastatic cells isolated from malignant ascites obtained from patients with advanced ovarian carcinoma were treated with non-targeted and LHRH receptor-targeted PPI-siRNA complexes. It was found that naked siRNA as well as PPI complexes with siRNA with scrambled sequence did not change the expression of CD44 mRNA (Fig. 2B). In contrast, incubation of cells with PPI-siRNA complexes significantly (P < 0.05) decreased the expression of CD44. Furthermore, LHRH tumor-targeted PPI-siRNA complexes reduced the expression much more efficiently (P < 0.05) when compared with a non-targeted system (PPI-CD44 siRNA). In fact, the expression of CD44 mRNA in ascitic cells treated with PPI-siRNA decreased down to 53.5% when compared with control cells (Fig. 2B). Targeting of the complex specifically to ovarian cancer cells by LHRH peptide significantly decreased the overexpression (down to 18.6% of control). In other words, the overexpression of CD44 in cells treated with cancer-targeted complexes was almost three fold lower when compared with cells treated with non-targeted complexes. A high efficiency of LHRH-PPI-siRNA in suppression of the targeted protein was confirmed by direct measurement of the expression of CD44 protein (Fig. 2D). Moreover, the suppression of CD44 protein by siRNA targeted to CD44 mRNA simultaneously inhibited the expression of the MDR1 gene (Fig. 2B).
Fig. 2.

Expression of CD44, MDR1 and LHRHR mRNA in cancer cells and tumor tissues. A, Expression of CD44 mRNA in normal tissues, primary tumor and metastases isolated from tissues of the same patients with different types of gynecological cancers. The expression of CD44 mRNA in corresponding primary tumor was set to 1 unit. Means ± SD are shown. *P < 0.05 when compared with primary tumor. B, Expression of CD44 and MDR1 mRNA in cancer cells isolated from malignant ascites obtained from patients with advanced ovarian carcinoma. Cells were incubated with substances indicated. Means ± SD are shown. *P < 0.05 when compared with control (cells incubated with fresh media); †P < 0.05 when compared with non-targeted complexes. C, Expression of LHRH receptors in gynecological tumors and healthy tissues. D, Expression of CD44 protein (immunocytochemistry) in human cancer cells isolated from malignant ascites from patient with advanced ovarian carcinoma. Cells were treated with PPI-siRNA-LHRH nanocomplex. Untreated cells received media alone. CD44 protein was stained using primary anti-CD44 antibody and secondary antibody conjugated with Cy3® fluorescence dye. Nuclei were stained with DAPI nuclear dye. Red color represents CD44 protein; blue color represents cell nuclei. Representative light and fluorescence microscope images are shown.
Cytotoxicity and Cell Invasiveness
The effect of the targeted LHRH-PPI-siRNA conjugate in cell invasion and cytotoxicity were tested in vitro. It was found that the drug conjugate significantly suppressed the invasiveness of cancer cells isolated from malignant ascites (Fig. 3A). It should be stressed that a similar effect was registered in the cells incubated both with fresh media alone and with media supplemented with a chemoattractant. In both cases, the percentage of cell invasion was inhibited by 6-7–fold. The incubation of ascitic cells with naked siRNA targeted to CD44 mRNA, naked scrambled siRNA, PPI dendrimer alone and siRNA with a scrambled sequence delivered by PPI dendrimer did not demonstrate toxicity in maximum available concentrations (Fig. 3B). Free non-bound TAX at a concentration of 55 μM led to the death of around 50-60% of cells. Therefore, this concentration of TAX was close to its IC50 dose in ascitic cells. Conjugation of TAX to a non-targeted PPI dendrimer enhanced its cytotoxic effect leading to the 1.6-fold decrease in cellular viability under the same drug concentration. PPI-siRNA conjugates induced a cell death comparable with that originated by PPI-TAX. Combination of siRNA and TAX in one dendrimer-based delivery system substantially increased the cytotoxicity of TAX leading to the further decrease in cellular viability down to 2.5-fold when compared with free non-bound TAX in the same concentration. Finally, the use of LHRH peptide to enhance cellular internalization of the LHRH-PPI-siRNA and LHRH-PPI-TAX complexes specifically by cancer cells enhanced the cytotoxicity of the conjugate. The viability of ascitic cells in this case was decreased almost 10-fold when compared with control cells, more than 5-fold when compared with free TAX and more than 2-fold when compared with non-targeted PPI-TAX-siRNA complex (Fig. 3B).
Fig. 3.

Invasion and viability of cancer cells isolated from malignant ascites obtained from patients with advanced ovarian carcinoma. A, Cell invasion was measured in the cells growing with media alone or media containing 15% fetal bovine serum (FBS, chemoattractant) and treated with LHRH-PPI-CD44 siRNA. B, Influence of different substances on cellular viability. Cells were incubated with substances indicated. Means ± SD are shown. *P < 0.05 when compared with control (cells incubated with fresh media); †P < 0.05 when compared with non-targeted complexes; ‡P < 0.05 when compared with free non-bound TAX.
Cellular Internalization of siRNA
We hypothesized that an increase in the cytotoxicity and efficiency of the suppression of targeted mRNA/protein by CD44 siRNA delivered by the PPI dendrimer is attributed to its higher internalization by cancer cells. Analysis of cellular internalization and distribution of naked and conjugated siRNA (Fig. 4) showed that naked siRNA poorly penetrates cancer cells leading to high red fluorescence of labeled siRNA in the medium and very low fluorescence inside the cell (Fig. 4A). In contrast, high level of both green (PPI dendrimer labeled by FITC) and red fluorescence (siRNA) was registered in cellular cytoplasm when siRNA was conjugated with the dendrimer. Moreover, targeting of dendrimer to cancer cells by LHRH peptide substantially enhanced cellular internalization of the dendrimer (Fig. 4B) and siRNA (Fig. 4B, C, D). It should be stressed that fluorescence of siRNA conjugated to the dendrimer is quenched. Consequently, the appearance of red fluorescence indicates that siRNA is separated from the dendrimer. These data show that (1) PPI dendrimer substantially enhanced cellular internalization of siRNA and (2) siRNA delivered by dendrimer was separated (released) from the carrier and accumulated predominately in the cytoplasm. Serial confocal microscope images (z-series) showed that distribution of siRNA was uniform from the top to the bottom of the cell and more uniform for siRNA delivered by targeted dendrimer (Fig. 4C, D). These data clearly show the high efficiency of cellular penetration of siRNA delivered by the PPI dendrimer and also the successful release of siRNA from the complex inside the cells.
Fig. 4.

Cellular internalization of naked siRNA and siRNA delivered by non-targeted and cancer cell targeted delivery systems. A, Representative images of naked fluorophore-labeled siRNA (siGLO Red, red fluorescence) incubated with ovarian cancer cells (z-series, from the top of the cell to the bottom). B, Representative images of ovarian cancer cells incubated with fluorescently labeled siRNA (siGLO Red, red fluorescence) complexated with non-targeted and LHRH-targeted PPI dendrimers labeled with FITC (green fluorescence). Superimposition of red (siRNA) and green (dendrimer) fluorescence images gives yellow color. C, Representative confocal microscopy images of ovarian cancer cells incubated with PPI-siRNA (z-series, from the top of the cell to the bottom). Red color represents siRNA. D, Representative confocal microscopy images of ovarian cancer cells incubated with LHRH-PPI-siRNA (z-series, from the top of the cell to the bottom). Red color represents siRNA. Scale bar – 15 μm.
In Vivo Antitumor Activity
In order to support the biologic significance of the in vitro findings and analyze the antitumor efficacy of the synthesized complexes, we performed in vivo experiments with nude mouse xenografts using human ovarian cancer cells isolated from malignant ascites from patients with advanced ovarian carcinoma. Mice were treated eight times, twice per week, within four weeks starting from day 0 when tumor volume reached 0.4 cm3. Expression of CD44 protein and apoptosis induction in tumor tissues isolated at the end of the experiments as well as tumor volume was measured. The animals were treated with the proposed formulations. Corresponding controls were used. The concentration of TAX in all drug-containing formulations was equal to 2.5 mg/kg. The experiments were terminated if the tumor size reached the maximum allowed by the Institutional Animal Care and Use Committee approved animal protocol (~2 cm3 that corresponded to approximately 10% of body mass). The data showed that either LHRH-PPI dendrimer (without drug and siRNA), free TAX, PPI-TAX or LHRH-PPI-TAX did not change significantly the expression of CD44 protein (Fig. 5). On the other hand, complexes of CD44 siRNA with tumor-targeted dendrimers containing TAX (LHRH-PPI-CD44 siRNA mixed with LHRH-PPI-TAX) almost completely inhibited the expression of targeted protein (Fig. 5, lower panel). The measurement of body distribution of non-targeted dendrimer showed a substantial accumulation of the dendrimer in the tumor, however, a significant amount of PPI dendrimer was also registered in the liver, kidney, lungs, spleen and heart (Fig. 6A, left panel). Targeting of the dendrimer to the tumor cells by LHRH peptide dramatically enhanced the accumulation of PPI dendrimer in the tumor and radically limited its accumulation in healthy organs (Fig. 6A, right panel). Analysis of cell death induction by two independent methods showed that in agreement with the in vitro data, conjugation of TAX to PPI dendrimer significantly enhanced apoptosis and, consequently, drug antitumor activity (Fig. 6A and B). Further enhancement was achieved by employing LHRH peptide as a tumor-targeting moiety. Finally, the suppression of CD44 protein by siRNA delivered with the PPI dendrimer targeted by LHRH peptide dramatically enhanced apoptosis in tumor by the proposed complexes (Fig. 6B, C). It should be stressed that targeting of drug composition by LHRH peptide significantly limited apoptosis induction in healthy organs (Fig. 6C). Consequently, such enhancements led to the more effective suppression of tumor growth by the tested formulations (Fig. 6D). The data obtained show that tumors in control animals (treated with saline) as well as those treated with PPI dendrimer alone, LHRH, PPI-siRNA with a scrambled sequence reached the maximum allowed by the institutional policy size by 10 days after beginning of the treatment (Fig. 6D, curves 1-4). As expected, free unbound TAX limited tumor growth and the maximal tumor size was reached at day 17 (Fig. 6D, curve 5). PPI-siRNA complexes and PPI-TAX conjugates showed more pronounced antitumor effect when compared with free TAX (Fig. 6D, curves 6 and 7) and slowed down tumor growth (maximum tumor size was reached on day 24). Consistently with in vitro data, targeting of PPI complexes to ovarian tumor by LHRH peptide significantly enhanced antitumor activity of TAX and CD44 siRNA (Fig. 6D, curves 8 and 9, respectively). Finally, the combination of the anticancer drug (TAX), siRNA targeted to CD44 mRNA, and tumor-specific targeting agent (LHRH peptide) delivered by the nanocarrier (PPI dendrimer) led to the almost complete shrinkage of the tumor within the 28-day study period (Fig. 6D, curve 10).
Fig. 5.
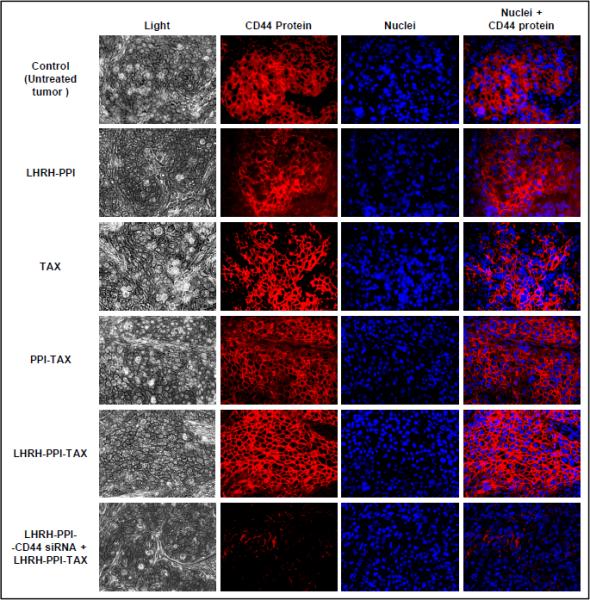
Expression of CD44 protein (immunohistochemistry) in tumor tissues from mice bearing xenografts of human cancer cells isolated from malignant ascites from patient with advanced ovarian carcinoma. Mice were treated eight times twice per week within four weeks starting from the day 0 with the indicated formulations. CD44 protein was stained with primary anty-CD44 and secondary antibodies conjugated with Cy3® fluorescence dye (red fluorescence). Nuclei were stained with DAPI dye (blue fluorescence). Representative fluorescence images are shown.
Fig. 6.

Organ distribution of non-targeted and tumor-targeted dendrimers, apoptosis and tumor volume in mice bearing xenografts of human cancer cells isolated from malignant ascites from patient with advanced ovarian carcinoma. After tumors reached a size of about 0.4 cm3, mice were treated eight times twice per week within four weeks starting from the day 0 with the indicated formulations. A, Organ distribution of non-targeted (PPI) and tumor-targeted (LHRH-PPI) dendrimers. B and C, Apoptosis induction in tumor and other organs at the end of the experiments (day 28 of the treatment). B, Representative fluorescence microscopy images of tumor tissue slides labeled by TUNEL. C, The enrichment of histone-associated DNA fragments (mono- and oligonucleosomes) per gram tissue in the tumor and different organs. Values in control animals were set to unit 1, and the degree of apoptosis was expressed in relative units. Means ± SD are shown. *P < 0.05 when compared with control. D, Tumor volume. Means ± SD are shown.
Discussion
In the present investigation, we successfully delivered siRNA targeting a commonly found surface receptor in ovarian cancer cells. We show that CD44 mRNA and protein are both overexpressed in the plasma membrane of cancer cells isolated from patients with gynecologic malignancies. Moreover, this overexpression is more pronounced in metastatic cells when compared with the cells from primary tumor. One can hypothesize that CD44 is involved in the development of resistance and invasiveness of cancer cells. Several facts support these suggestions. Previously, we found that the expression of CD44 correlates with the expression of the main drug efflux pump – P-glycoprotein, resistance to chemotherapy and progression of metastases (9, 10, 38). Moreover, current in vitro and in vivo data showed that the suppression of CD44 protein enhances the activity of the anticancer drug – paclitaxel, initiates and promotes cell death by apoptosis and leads to tumor shrinkage. This finding also indicates that CD44 might be involved in the development of metastases and its expression correlates with cell invasiveness. Present data obtained using cell invasion assays support this suggestion and show that the suppression of CD44 protein dramatically decreases the invasiveness of malignant cells. Consequently, it is understandable that CD44 may represent an attractive target for chemotherapy both as a primary target for cell death induction and as an inducer of drug resistance. In order to validate the hypothesis, we constructed a novel dendrimer-based delivery system that is capable of simultaneously inducing cell death and suppressing cellular resistance specifically in ovarian tumor cells.
The proposed multifunctional and multicomponent delivery system includes a PPI dendrimer as a carrier, paclitaxel as an anticancer drug/cell death inducer, LHRH peptide as a cancer-specific targeting moiety, and siRNA targeted to CD44 mRNA as a specific CD44+ ovarian cancer cell death enhancer. In order to increase the stability of the proposed drug-siRNA combination, complexes of siRNA with dendrimers were protected from the harsh extracellular environment by caging with DTBP. We previously used such caging of PPI dendrimer-siRNA complexes and showed its high efficiency in increasing the stability of complexed siRNA in plasma (24). Detailed characterization of resulting complexes showed that indeed siRNA and the carrier were condensed into spherical nanoparticles with an increased stability.
Previously, we revealed that a similar dendrimer-based system effectively delivered and released inside the cells different anticancer drugs including paclitaxel (25, 40). Present data shows that siRNA is also effectively delivered into the cytoplasm of cancer cells by the proposed nanosystem and separated (released) from the carrier in the cytoplasm. This separation is possible due in part to lower pH in the cytoplasm and cellular organelles that contains and transports the system after its internalization into cancer cells. Under these conditions, negative phosphate groups of siRNA become protonated and lose their charge, so the siRNA molecule becomes less negative which helps to release electrostatically bound siRNA. The so called proton sponge effect (41) might also be responsible for the effective release of siRNA into the cytoplasm. Moreover, the distribution of siRNA in the cytoplasm is homogenous in all directions including so-called z-section from the top to the bottom of the cells. Our present data show that the delivered and released siRNA has specific activity and effectively suppresses the targeted CD44 mRNA and consequently CD44 protein expression. It should be stressed that the suppression of CD44 protein in turn led to the inhibition of MDR1 mRNA. Therefore, the use of siRNA targeted to CD44 inhibited both genes primarily responsible for multidrug resistance in ovarian cancer cells. Consequently, such suppression significantly enhanced the ability of TAX to induce cell death by apoptosis and as a result the proposed complex delivery system has high antitumor efficiency in vitro and in vivo.
In summary, we were able to verify the hypothesis and show that suppression of CD44 enhances the efficiency of chemotherapy. The proposed cancer targeted complex chemotherapy approach and the multifunctional nanocarrier-based delivery system especially designed to effectively suppress invasiveness and kill ovarian cancer cells substantially enhances efficiency of therapy of ovarian cancer and possibly other gynecologic malignancies, limiting at the same time adverse side effect of chemotherapy on healthy tissues. In the new era of personalized targeted therapies, having a drug delivery system that can efficiently deliver siRNA to a precise genomic target is pivotal. By targeting CD44 in ovarian cancer cells we show proof of principle that targeted nanomedicine is a possible approach to precision therapies.
Supplementary Material
Statement of Translational Relevance.
The majority of patients with ovarian carcinoma are diagnosed in the metastatic stage of the disease. Chemotherapy and surgery are the initial treatment options; however, the development of drug resistance is almost universal. Consequently, the development of novel, effective, and safe approaches of treatment is vitally important. However, low therapeutic efficiency and significant toxicity represent two critical barriers limiting progress in the therapy. The presented research addresses these important challenges by substantially enhancing the efficiency of therapy and reducing its adverse side effects. This paper represents proof-of-concept of a novel treatment approach that combines chemo and gene therapy by tumor specific targeted delivery of siRNA and anticancer drugs. The present preclinical research forms a basis for future clinical trials using this novel nanoscale-based multifunctional delivery system that among its many assets is the successful delivery of siRNA and a very specific targeted delivery to ovarian cancer cells sparing normal organs.
Acknowledgments
Financial support: T.M. – NIH/NCI R01 CA100098, NIH/NCI R01 CA138533; NSF CBET 0933966; L.R. – NIH/NCI R01 CA120429.
Grant Support
This research was supported in part by grants from NIH/NCI (R01 CA100098 and R01 CA138533 to T.M.; R01 CA120429 to L.R.) and NSF (CBET 0933966 to T.M.).
Footnotes
Conflict of Interest Disclosure Statement: There are no conflicts to disclose.
References
- 1.Siegel R, Ward E, Brawley O, Jemal A. Cancer statistics, 2011: the impact of eliminating socioeconomic and racial disparities on premature cancer deaths. CA: a cancer journal for clinicians. 2011;61:212–36. doi: 10.3322/caac.20121. [DOI] [PubMed] [Google Scholar]
- 2.Cannistra SA. Cancer of the ovary. N Engl J Med. 2004;351:2519–29. doi: 10.1056/NEJMra041842. [DOI] [PubMed] [Google Scholar]
- 3.Naor D, Sionov RV, Ish-Shalom D. CD44: structure, function, and association with the malignant process. Adv Cancer Res. 1997;71:241–319. doi: 10.1016/s0065-230x(08)60101-3. [DOI] [PubMed] [Google Scholar]
- 4.Bajorath J, Greenfield B, Munro SB, Day AJ, Aruffo A. Identification of CD44 residues important for hyaluronan binding and delineation of the binding site. J Biol Chem. 1998;273:338–43. doi: 10.1074/jbc.273.1.338. [DOI] [PubMed] [Google Scholar]
- 5.Screaton GR, Bell MV, Jackson DG, Cornelis FB, Gerth U, Bell JI. Genomic structure of DNA encoding the lymphocyte homing receptor CD44 reveals at least 12 alternatively spliced exons. Proc Natl Acad Sci U S A. 1992;89:12160–4. doi: 10.1073/pnas.89.24.12160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Aruffo A, Stamenkovic I, Melnick M, Underhill CB, Seed B. CD44 is the principal cell surface receptor for hyaluronate. Cell. 1990;61:1303–13. doi: 10.1016/0092-8674(90)90694-a. [DOI] [PubMed] [Google Scholar]
- 7.Bourguignon LY, Peyrollier K, Xia W, Gilad E. Hyaluronan-CD44 interaction activates stem cell marker Nanog, Stat-3-mediated MDR1 gene expression, and ankyrin-regulated multidrug efflux in breast and ovarian tumor cells. J Biol Chem. 2008;283:17635–51. doi: 10.1074/jbc.M800109200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mackay CR, Terpe HJ, Stauder R, Marston WL, Stark H, Gunthert U. Expression and modulation of CD44 variant isoforms in humans. J Cell Biol. 1994;124:71–82. doi: 10.1083/jcb.124.1.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rodriguez-Rodriguez L, Sancho-Torres I, Leakey P, Gibbon DG, Comerci JT, Ludlow JW, et al. CD44 splice variant expression in clear cell carcinoma of the ovary. Gynecol Oncol. 1998;71:223–9. doi: 10.1006/gyno.1998.5108. [DOI] [PubMed] [Google Scholar]
- 10.Rodriguez-Rodriguez L, Sancho-Torres I, Mesonero C, Gibbon DG, Shih WJ, Zotalis G. The CD44 receptor is a molecular predictor of survival in ovarian cancer. Med Oncol. 2003;20:255–63. doi: 10.1385/MO:20:3:255. [DOI] [PubMed] [Google Scholar]
- 11.Speiser P, Wanner C, Breitenecker G, Kohlberger P, Kainz C. CD-44 is not involved in the metastatic spread of ovarian cancer in vivo. Anticancer Res. 1995;15:2767–9. [PubMed] [Google Scholar]
- 12.Stadlmann S, Raffeiner R, Amberger A, Margreiter R, Zeimet AG, Abendstein B, et al. Disruption of the integrity of human peritoneal mesothelium by interleukin-1beta and tumor necrosis factor-alpha. Virchows Arch. 2003;443:678–85. doi: 10.1007/s00428-003-0867-2. [DOI] [PubMed] [Google Scholar]
- 13.Chiu YL, Ali A, Chu CY, Cao H, Rana TM. Visualizing a correlation between siRNA localization, cellular uptake, and RNAi in living cells. Chem Biol. 2004;11:1165–75. doi: 10.1016/j.chembiol.2004.06.006. [DOI] [PubMed] [Google Scholar]
- 14.de Fougerolles A, Vornlocher H-P, Maraganore J, Lieberman J. Interfering with Disease: A Progress Report on siRNA-based Therapeutics. Nat Rev Drug Discovery. 2007;6:443–53. doi: 10.1038/nrd2310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Song E, Zhu PC, Lee S-K, Chowdhury D, Kussman S, Dykxhoorn DM, et al. Antibody Mediated in vivo Delivery of Small Interfering RNAs via Cell-Surface Receptors. Nature Biotech. 2005;23:709–17. doi: 10.1038/nbt1101. [DOI] [PubMed] [Google Scholar]
- 16.Uprichard SL. The Therapeutic Potential of RNA Interference. FEBS Lett. 2005;579:5996–6007. doi: 10.1016/j.febslet.2005.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Betigeri S, Pakunlu RI, Wang Y, Khandare JJ, Minko T. JNK1 as a molecular target to limit cellular mortality under hypoxia. Mol Pharm. 2006;3:424–30. doi: 10.1021/mp060014x. [DOI] [PubMed] [Google Scholar]
- 18.Patil ML, Zhang M, Betigeri S, Taratula O, He H, Minko T. Surface-modified and internally cationic polyamidoamine dendrimers for efficient siRNA delivery. Bioconjug Chem. 2008;19:1396–403. doi: 10.1021/bc8000722. [DOI] [PubMed] [Google Scholar]
- 19.Patil ML, Zhang M, Minko T. Multifunctional triblock Nanocarrier (PAMAM-PEG-PLL) for the efficient intracellular siRNA delivery and gene silencing. ACS Nano. 2011;5:1877–87. doi: 10.1021/nn102711d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Patil ML, Zhang M, Taratula O, Garbuzenko OB, He H, Minko T. Internally cationic polyamidoamine PAMAM-OH dendrimers for siRNA delivery: effect of the degree of quaternization and cancer targeting. Biomacromolecules. 2009;10:258–66. doi: 10.1021/bm8009973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Saad M, Garbuzenko OB, Minko T. Co-delivery of siRNA and an anticancer drug for treatment of multidrug-resistant cancer. Nanomedicine (Lond) 2008;3:761–76. doi: 10.2217/17435889.3.6.761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Taratula O, Garbuzenko O, Savla R, Wang YA, He H, Minko T. Multifunctional nanomedicine platform for cancer specific delivery of siRNA by superparamagnetic iron oxide nanoparticles-dendrimer complexes. Curr Drug Deliv. 2011;8:59–69. doi: 10.2174/156720111793663642. [DOI] [PubMed] [Google Scholar]
- 23.Taratula O, Garbuzenko OB, Chen AM, Minko T. Innovative strategy for treatment of lung cancer: targeted nanotechnology-based inhalation co-delivery of anticancer drugs and siRNA. J Drug Target. 2011;19:900–14. doi: 10.3109/1061186X.2011.622404. [DOI] [PubMed] [Google Scholar]
- 24.Taratula O, Garbuzenko OB, Kirkpatrick P, Pandya I, Savla R, Pozharov VP, et al. Surface-engineered targeted PPI dendrimer for efficient intracellular and intratumoral siRNA delivery. J Control Release. 2009;140:284–93. doi: 10.1016/j.jconrel.2009.06.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Taratula O, Savla R, He H, Minko T. Poly(propyleneimine) dendrimers as potential siRNA delivery nanocarrier: from structure to function. Int J Nanotechnol. 2011;8:36–52. [Google Scholar]
- 26.Zinselmeyer BH, Mackay SP, Schatzlein AG, Uchegbu IF. The lower-generation polypropylenimine dendrimers are effective gene-transfer agents. Pharm Res. 2002;19:960–7. doi: 10.1023/a:1016458104359. [DOI] [PubMed] [Google Scholar]
- 27.Jayant S, Khandare JJ, Wang Y, Singh AP, Vorsa N, Minko T. Targeted sialic acid-doxorubicin prodrugs for intracellular delivery and cancer treatment. Pharm Res. 2007;24:2120–30. doi: 10.1007/s11095-007-9406-1. [DOI] [PubMed] [Google Scholar]
- 28.Pakunlu RI, Wang Y, Tsao W, Pozharov V, Cook TJ, Minko T. Enhancement of the efficacy of chemotherapy for lung cancer by simultaneous suppression of multidrug resistance and antiapoptotic cellular defense: novel multicomponent delivery system. Cancer Res. 2004;64:6214–24. doi: 10.1158/0008-5472.CAN-04-0001. [DOI] [PubMed] [Google Scholar]
- 29.Shah V, Taratula O, Garbuzenko OB, Patil ML, Savla R, Zhang M, et al. Genotoxicity of Different Nanocarriers: Possible Modifications for the Delivery of Nucleic Acids. Curr Drug Discov Technol. 2013;10:8–15. [PMC free article] [PubMed] [Google Scholar]
- 30.Chandna P, Khandare JJ, Ber E, Rodriguez-Rodriguez L, Minko T. Multifunctional tumor-targeted polymer-peptide-drug delivery system for treatment of primary and metastatic cancers. Pharm Res. 2010;27:2296–306. doi: 10.1007/s11095-010-0235-2. [DOI] [PubMed] [Google Scholar]
- 31.Chandna P, Saad M, Wang Y, Ber E, Khandare J, Vetcher AA, et al. Targeted proapoptotic anticancer drug delivery system. Mol Pharm. 2007;4:668–78. doi: 10.1021/mp070053o. [DOI] [PubMed] [Google Scholar]
- 32.Zhang M, Garbuzenko OB, Reuhl KR, Rodriguez-Rodriguez L, Minko T. Two-in-one: combined targeted chemo and gene therapy for tumor suppression and prevention of metastases. Nanomedicine (Lond) 2012;7:185–97. doi: 10.2217/nnm.11.131. [DOI] [PubMed] [Google Scholar]
- 33.Dharap SS, Wang Y, Chandna P, Khandare JJ, Qiu B, Gunaseelan S, et al. Tumor-specific targeting of an anticancer drug delivery system by LHRH peptide. Proc Natl Acad Sci U S A. 2005;102:12962–7. doi: 10.1073/pnas.0504274102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Betigeri S, Zhang M, Garbuzenko O, Minko T. Non-viral systemic delivery of siRNA or antisense oligonucleotides targeted to Jun N-terminal kinase 1 prevents cellular hypoxic damage. Drug Deliv Transl Res. 2011;1:13–24. doi: 10.1007/s13346-010-0003-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Garbuzenko OB, Saad M, Pozharov VP, Reuhl KR, Mainelis G, Minko T. Inhibition of lung tumor growth by complex pulmonary delivery of drugs with oligonucleotides as suppressors of cellular resistance. Proc Natl Acad Sci U S A. 2010;107:10737–42. doi: 10.1073/pnas.1004604107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Garbuzenko OB, Saad M, Betigeri S, Zhang M, Vetcher AA, Soldatenkov VA, et al. Intratracheal versus intravenous liposomal delivery of siRNA, antisense oligonucleotides and anticancer drug. Pharm Res. 2009;26:382–94. doi: 10.1007/s11095-008-9755-4. [DOI] [PubMed] [Google Scholar]
- 37.Saad M, Garbuzenko OB, Ber E, Chandna P, Khandare JJ, Pozharov VP, et al. Receptor targeted polymers, dendrimers, liposomes: which nanocarrier is the most efficient for tumor-specific treatment and imaging? J Control Release. 2008;130:107–14. doi: 10.1016/j.jconrel.2008.05.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Miletti-Gonzalez KE, Chen S, Muthukumaran N, Saglimbeni GN, Wu X, Yang J, et al. The CD44 receptor interacts with P-glycoprotein to promote cell migration and invasion in cancer. Cancer Res. 2005;65:6660–7. doi: 10.1158/0008-5472.CAN-04-3478. [DOI] [PubMed] [Google Scholar]
- 39.Dharap SS, Minko T. Targeted proapoptotic LHRH-BH3 peptide. Pharm Res. 2003;20:889–96. doi: 10.1023/a:1023839319950. [DOI] [PubMed] [Google Scholar]
- 40.Khandare JJ, Jayant S, Singh A, Chandna P, Wang Y, Vorsa N, et al. Dendrimer versus linear conjugate: Influence of polymeric architecture on the delivery and anticancer effect of paclitaxel. Bioconjug Chem. 2006;17:1464–72. doi: 10.1021/bc060240p. [DOI] [PubMed] [Google Scholar]
- 41.Benjaminsen RV, Mattebjerg MA, Henriksen JR, Moghimi SM, Andresen TL. The Possible "Proton Sponge " Effect of Polyethylenimine (PEI) Does Not Include Change in Lysosomal pH. Mol Ther. 2013;21:149–57. doi: 10.1038/mt.2012.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.