

Abstract
Background
Rate-dependent effects on the Ca2+ sub-system in a rat ventricular myocyte are investigated. Here, we employ a deterministic mathematical model describing various Ca2+ signalling pathways under voltage clamp (VC) conditions, to better understand the important role of calmodulin (CaM) in modulating the key control variables Ca2+/calmodulin-dependent protein kinase-II (CaMKII), calcineurin (CaN), and cyclic adenosine monophosphate (cAMP) as they affect various intracellular targets. In particular, we study the frequency dependence of the peak force generated by the myofilaments, the force-frequency response (FFR).
Methods
Our cell model incorporates frequency-dependent CaM-mediated spatially heterogenous interaction of CaMKII and CaN with their principal targets (dihydropyridine (DHPR) and ryanodine (RyR) receptors and the SERCA pump). It also accounts for the rate-dependent effects of phospholamban (PLB) on the SERCA pump; the rate-dependent role of cAMP in up-regulation of the L-type Ca2+ channel (ICa,L); and the enhancement in SERCA pump activity via phosphorylation of PLB.
Results
Our model reproduces positive peak FFR observed in rat ventricular myocytes during voltage-clamp studies both in the presence/absence of cAMP mediated β-adrenergic stimulation. This study provides quantitative insight into the rate-dependence of Ca2+-induced Ca2+-release (CICR) by investigating the frequency-dependence of the trigger current (ICa,L) and RyR-release. It also highlights the relative role of the sodium-calcium exchanger (NCX) and the SERCA pump at higher frequencies, as well as the rate-dependence of sarcoplasmic reticulum (SR) Ca2+ content. A rigorous Ca2+ balance imposed on our investigation of these Ca2+ signalling pathways clarifies their individual roles. Here, we present a coupled electromechanical study emphasizing the rate-dependence of isometric force developed and also investigate the temperature-dependence of FFR.
Conclusions
Our model provides mechanistic biophysically based explanations for the rate-dependence of CICR, generating useful and testable hypotheses. Although rat ventricular myocytes exhibit a positive peak FFR in the presence/absence of beta-adrenergic stimulation, they show a characteristic increase in the positive slope in FFR due to the presence of Norepinephrine or Isoproterenol. Our study identifies cAMP-mediated stimulation, and rate-dependent CaMKII-mediated up-regulation of ICa,L as the key mechanisms underlying the aforementioned positive FFR.
Background
Cardiac muscle contraction is a result of a transient increase in intracellular Ca2+ concentration[ Ca2+] myo. Sarcolemmal (SL) membrane depolarization triggers Ca2+ influx via the dihydropyridine (DHP)-sensitive L-type Ca2+ channels. Following diffusion across a small sub-membrane dyadic space, this influx activates ryanodine receptors (RyRs) controlling ryanodine-sensitive Ca2+ release channels in the junctional portion of the sarcoplasmic reticulum (jSR). Fabiato and Fabiato [1] named the process calcium-induced calcium release (CICR). Ca2+ subsequently diffuses from the dyadic space into the myoplasm. Ultimately, intracellular Ca2+ concentration [ Ca2+] myo is returned to resting levels by combination of: (a) Ca2+ buffering in the dyadic space and myoplasm; (b) sequestration of Ca2+ by sarcoplasmic/endoplasmic reticulum Ca2+-ATPase (SERCA)-type calcium pumps lining the longitudinal portion of the sarcoplasmic reticulum (LSR); and (c) Ca2+ extrusion from the myoplasm by Na+/ Ca2+ exchangers and Ca2+-ATPase pumps on the sarcolemmal membrane.
Ca2+ is an extremely important and highly versatile second messenger in cardiac cells, which plays a crucial role not only in excitation-contraction (E-C) coupling but also in excitation-transcription coupling [2]. Various inter-connected Ca2+ signalling pathways help preserve the integrity of the cellular Ca2+ system despite any changes in pacing frequency. Specifically, Ca2+ triggers the CaM-mediated rate-dependent effects of CaMKII and CaN on the characteristics of the apposed dihydropyridine (DHP) and ryanodine-sensitive Ca2+ channels in the dyad, whose interaction forms the basis for CICR. The proteins CaMKII and CaN not only influence the release mechanism but also affect the SERCA pump either directly or indirectly via phospholamban (PLB), thus modulating the Ca2+ uptake process [3-6]. It is also known that β-adrenergic stimulation of the cardiac cell, mediated by the second messenger cAMP, modulates the frequency dependence of the peak force generated by the myofilaments, the force-frequency response (FFR). These protein-mediated and second messenger pathways help maintain Ca2+ homeostasis over a wide range of stimulation frequencies.
Cardiac contractile function is closely coupled with heart rate (Bowditch effect [7]). Although positive [8], almost flat [9] and negative [10,11] peak FFR have been reported in the literature, it is clear from in-vitro studies involving stimulation in the physiological range of frequencies [12,13] that rat ventricle exhibits a positive peak FFR. The issue of force-frequency relationship requires broadened investigation into the various underlying cellular and molecular mechanisms. We propose that mathematical modeling would be a useful tool in helping to sort out this complex issue.
Methods
All simulations and analysis were performed on a 2.8GHz Intel Ⓡ Core TM2 Duo CPU-based computer using Microsoft Windows XP operating system. The sarcolemmal membrane charge balance equations, the Ca2+ material balance equations in the myoplasm and SR, and the force balance equations describing the model for myofilament contraction constitute a set of 93 ordinary differential equations (ODEs). A fixed-step Merson-modified Runge-Kutta 4th-order numerical integration scheme [14] was used to solve this set of 1st-order differential equations (ODE) describing the dynamic model. The free Ca2+ concentration in the dyad is governed by the time courses of the Ca2+ fluxes through Ca2+ transport systems, as well as by the time course of Ca2+ binding to Ca2+ buffers present in the junction [15]. Description of the spatio-temporal dynamics of calcium transients in the dyad triggered by Ca2+ stimulus (basis of CICR) requires calculation of the partial differential equations (PDE) of the whole reaction-diffusion system. Formation and dissipation of Ca2+ gradients around an open channel (DHP-sensitive and Ry-sensitive channels in the dyad) is assumed instantaneous as was validated for microsecond timescale and nanoscopic space by Naraghi and Neher [16]. Local Ca2+ concentration in the vicinity of open channels (located on opposing boundaries of the dyadic space) was calculated as the steady state gradient around a point source [17]. The Ca2+ concentration increments from individual channels at each point in space were assumed to be additive [16,18]. The software kernel follows the changes in the state of trigger and release channels together with variables like membrane voltage and spatial Ca2+ concentration to calculate the instantaneous rate constants and estimate the duration of transient events. Crank [19] discusses diffusion problems in a two-phase heterogeneous medium and shows that diffusion through a system of barriers (RyR feet structures in the dyadic cleft space) can be approximated by diffusion in the same region without barriers but with a reduced effective diffusion coefficient. We hence take this approach in modeling the Ca2+ diffusion by solving the 2-D Laplacian equation (Krishna et al. [15], Appendix A3, Eq. 140) in the DCU without explicitly accounting for local potential fields. More specifically, an explicit finite difference scheme was used to solve these Laplacian equations describing Ca2+-diffusion in the dyadic space analogous to the method detailed in Smith et al. [20]. Specifically, a radial symmetry is employed in solving the PDE in the dyadic volume allowing the solution to be computed in a rectangular cross-section discretized into a 20 by 20 cartesian grid. The spatial step size used in the r and z-direction (Figure 1B, Krishna et al. [15]) was 10 nm and 0.76 nm respectively (Table two, Krishna et al. [15]). The 20x20 grid size was used to obtain a stable numerical solution using the explicit finite difference scheme employed to solve the PDE. Obtaining an accurate description for Ca2+-diffusion in the dyadic space is vital to ensure adequate time delays associated with RyR release (z-direction) and Ca2+-diffusion into the cytosol (r-direction) which controls the rate of SR Ca2+-uptake via the SERCA pump. Both of these delays are important in ensuring robust luminal sensor mediated RyR refractory characteristics (described in Krishna et al. [15]). We use the method of lines (discretization in space) to solve the PDE. The full set of ODEs and finite difference equations are solved simultaneously to obtain the complete solution. Execution of a single cycle which translates to 200 ms at 5 Hz took 21 seconds with a time step of 1 μs. A non-linear leastsquares method [21] was used for parameter estimation and data fitting. Results were visualized using Matlab (Mathworks, Natick, MA) and Origin (OriginLab Corp., Northampton, MA).
Figure 1.

CaM-CaMKII-CaN pathways. Reaction Maps. (A) Cooperative binding of 2 Ca2+ to CaM sequentially to the C and then N-terminal hands along with binding of CaM buffers; (B) Probabilistic model of CaMKII subunit switching; (C) Reaction map for reversible binding of CaM, Ca 2CaM and Ca 4CaM to CaN.
Model development
Our objective was to develop a model of the rat ventricular cell under voltage clamp conditions, which includes the description of various Ca2+ signalling pathways in the dyadic space, the myoplasmic medium and the sarcoplasmic reticulum. Therefore, we start with a broad discussion of the key proteins involved in the Ca2+ signalling mechanism and continue with a progressively more detailed description of their influence on respective targets. It is important to note that all Ca2+ concentrations discussed in the model pertain to unbound Ca2+ unless specified. A detailed description of the membrane classification, channel and exchanger distribution as well as the various fluid compartments involved is given in Krishna et al. [15].
Calmodulin mediates the regulation of a variety of Ca2+-dependent signalling pathways in the heart involving CaMKII and CaN [2,22]. These protein-mediated interactions form the basis for a robust mechanism that enables a cell’s response to increased heart rate. CaMKII is reported to be responsive when targeted to Ca2+ release sites such as the dyadic cleft, and CaN is responsive to gradual changes in the lower-amplitude myoplasmic Ca2+ signals [2]. This heterogenous response is a result of the different affinities of CaMKII and CaN for CaM [22] and the non-uniform distribution of these proteins in the cell. Recent study [23] also attributes a key role in frequency-dependent acceleration of relaxation to activated-CaMKII in the cytosol. The model of the CaM-dependent Ca2+ signalling process (Figure 1), which includes a reaction map for cooperative binding of Ca2+ to CaM, the scheme for CaM buffering, probabilistic model of CaMKII subunit switching and the reaction map for reversible binding of CaM, Ca2CaM and Ca4CaM to CaN, is adopted from Saucerman et al. [2]. However, to reproduce relative local CaMKII and CaN activity, modifications were made to the rate constants for CaM buffering in the dyad (Table five, Krishna et al. [15]). Specifically, a limitation of the Saucerman and Bers model is that it is based on little available information regarding the operation of CaM buffers [24] at locations where CaM encounters very high Ca2+ concentrations (compared with the myoplasm).
We incorporate the effects of β-adrenergic stimulation via cAMP-dependent modulation based on a model (Figure 2) derived from Demir et al. [25]. Stimulation of β-adrenoceptors by Isoproterenol (Iso) results in the activation of a G protein (g s) that stimulates Adenylate cyclase (ADC) and enhances the production of cAMP. Subsequently, cAMP may directly or indirectly activate various intracellular targets including ion channels and exchangers. The indirect modulation involves activation of cAMP-dependent Protein kinase A (PKA) before modulation of the channel protein. The reaction kinetics for the cGMP-mediated pathway (Figure 2) involving acetylcholine (ACh), nitric oxide (NO) and soluble guanylate cyclase (sGC) are adopted from Yang et al. [26]. Although it is well known that cGMP modulates its targets via protein kinase G (PKG) or phosphodiesterase (PDE), we have refrained from modeling these protein interactions. Given that NO synthase inhibition and/or NO donor had little or only marginal effects on the FFR in rat ventricular myocardium [27], the cGMP level is kept constant in this study.
Figure 2.

cAMP and cGMP pathways. Reaction pathways triggered by sympathetic and parasympathetic neural stimuli invoking cAMP [25] and cGMP-mediated [26] modulation of various sub-cellular targets respectively. ACh-mediated effects of junctional receptor on Na+ current and background Na+ current and extrajunctional muscarinic M 2/K ACh receptor on I K,ACh (direct muscarinic pathway) and cAMP-mediated effects of β-adrenoreceptor (adrenergic pathway) and cholinergic M 2/ADC receptor (indirect muscarinic pathway) on L-type Ca2+ (I Ca,L), K+ (I K), hyperpolarization-activated (I f), and Na+/ K+ (I NaK)currents are shown ‡. Although cAMP-dependent modulation of I Ca,L and Iup are known to be PKA-mediated, we make use of a lumped cAMP term to model this influence. A similar lumped cAMP term is employed to modulate phosphorylation of PLB which in turn affects the SERCA pump. ACh triggered NO-mediated synthesis of cGMP by sGC is modeled employing a 3-state model for sGC transition based on Yang et al. [26]. cGMP is involved in suppressing cAMP activity via PDE. cGMP also enhances IPMCA, Ca2+ activated K+ channel (IKCa) and suppresses ICa,L, Icyt,serca via PKG (not explicitly modeled, but lumped into a cGMP term). ‡The rat ventricular cell model used in this study is however limited to Ca2+ related channel, exchanger and pumps (ICa,L, INaCa, IPMCA and Icyt,serca), while lacking exclusive Na+ or K+ related channels and transporters (as shown in Figure 2, Krishna et al. [15]). The part of the model describing cAMP-mediated pathway used in our study is highlighted (blue).
DHP-sensitive Ca2+ channel
Upon cell depolarization, the L-Type Ca2+ channel (ICa,L) brings trigger Ca2+ into the dyad that facilitates Ca2+ release from the apposed Ry-sensitive Ca2+ channel associated with the membrane of the junctional sarcoplasmic reticulum (jSR). Feedback controlled interaction between these apposed channels is critical in CICR as well as in the maintenance of the overall cellular Ca2+ balance. Besides membrane voltage, gating of the ICa,L channel is also influenced by two prominent Ca2+-mediated effects, namely Ca2+-dependent inactivation (CDI) and Ca2+-dependent facilitation (CDF).
In CDI, Ca2+ that enters the dyad either via the ICa,L channel or through RyR release from the jSR binds to the protein Calmodulin (CaM) which is tethered to the C-terminus of the ICa,L channel [28], modulating the interaction of CaM with the Ca2+ channel. Leucine-Alanine (LA) and Isoleucine-Glutamine (IQ) are 2 adjacent motifs in the Ca2+ sensing domain of the C-terminus of the ICa,L channel. A Ca2+-dependent switch of CaM from LA to IQ motif removes CaM from the inner mouth of the channel pore, thus causing an enhancement in inactivation by facilitating the constriction of the pore [29,30]. CDI is a critical negative feedback mechanism which causes decreased Ca2+ entry via ICa,L when the SR load is high with an accompanying large myoplasmic Ca2+ transient, and it results in increased Ca2+ entry via ICa,L when [ Ca2+] myo is small due to a low SR load. We utilize a 2-state model, as shown in Figure 3A of our previous study Krishna et al. [15], to simulate CDI.
Figure 3.
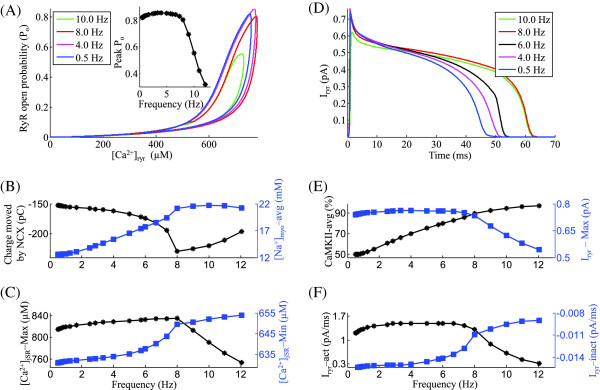
Iryr. Frequency-dependent characteristics of the SR Ca2+ release channel Iryr. Voltage clamp protocol used is a 50 ms step pulse to 10 mV from a holding potential of -40 mV. (A) RyR open probability vs [ Ca2+] ryr for increasing frequency of stimulation (0.5 Hz to 10.0 Hz). The inset shows the frequency-dependent modulation of peak RyR open probability. (B) Frequency dependence of average charge transported by NCX and average [ Na+] myo in the myoplasm. (C) Rate dependence of pre-release maximum and post-release minimum SR Ca2+ level. (D) Traces for RyR release for the corresponding frequencies of stimulation in panel A. (E) Dependence of average level of activated-CaMKII and peak Iryr current on frequency of stimulation. (F) Frequency-dependent changes in average Iryr activation and inactivation rates. The legend for traces in panel A is shared by panel D.
In contrast, Ca2+-dependent facilitation (CDF) is a Ca2+/CaM-mediated enhancement in ICa,L via activation of CaMKII [31], which has been shown to tether to the ICa,L channel [32], functioning as a Ca2+ signalling sensor for facilitation. Activated Calcineurin (CaN) is also observed to facilitate Ca2+ entry via the ICa,L channel [33]. CDF is implicated in causing a gradual increase in amplitude and an accompanying decrease in inactivation over consecutive pulses after a resting interval [34]. Enhancement of ICa,L caused by activated-CaMKII and CaN plays a key role in negating the effects of incomplete ICa,L channel recovery at faster heart rates, thus helping to improve cardiac performance during exercise. Although CDF and CDI of ICa,L coexist, CDI responds much faster (within the same beat) than CDF (over several beats). Our model incorporates CDF by allowing the rate constants in the 6-state Markovian model of the ICa,L channel (, and in Figure 3A, Krishna et al. [15]) to be a function of the available active CaMKII and CaN.
Rate-dependent increases in ICa,L can also be caused by frequency-dependent increase in β-adrenergic stimulation increasing the level of available cAMP (Table 1), which in turn causes an enhancement in ICa,L channel current via protein kinase-A (PKA). Although PKA is involved in the indirect regulation of the ICa,L channel, its effect is considered lumped into the conductance term in the ionic current description (Appendix A3, Equations 1-2). The effect of β-adrenergic stimulation via cAMP, particularly its dose-dependent influence on L-Type Ca2+ channels both in terms of modifying the single channel behavior such as Ca2+ ion permeability as well as overall channel recruitment characteristics, is not clearly understood. While cAMP has been shown to increase channel open probability [35,36], increased levels of cAMP also result in increased phosphorylation of L-type Ca2+ channels, causing an increased permeability to Ca2+ ions [36-38].
Table 1.
Frequency dependence of intracellular cAMP concentration
| Stimulation frequency (Hz) | Intracellular cAMP concentration (μM) |
|---|---|
| 0.50 - 1.67 |
2.67 δ |
| 2.50 |
3.00 |
| 2.86 |
3.19 |
| 3.33 |
3.45 |
| 4.00 |
3.85 |
| 5.00 |
4.67 |
| 5.71 |
5.58 |
| 6.67 |
18.06 |
| 8.00 |
24.58 |
| 10.00 |
27.03 |
| 12.00 | 29.17 |
δ- Value used under basal conditions. Intracellular cAMP concentration corresponding to maximal β-adrenergic stimulation at various stimulation frequencies used in the model (based on Equation 2, Demir et al. [25]).
Ry-sensitive Ca2+ release channel
Ry-sensitive Ca2+ channels on the jSR membrane respond to the trigger Ca2+ entering the dyadic space via the ICa,L channel on the plasma membrane. A larger Ca2+ release from the jSR follows, forming the basis for Ca2+ induced Ca2+ release (CICR) and subsequent contraction of the ventricular cell. CaM that is tethered to the Ry-sensitive Ca2+ channel [39] facilitates frequency-dependent CaMKII and CaN-assisted modulation of that channel. Although CaMKII is known to bind to the RyR [40-42], the effect of this association has not yet been resolved. In lipid bilayer studies, CaMKII has been shown to increase [40,41,43] or decrease [44] RyR open probability. In studies on rat ventricular myocytes, it has been shown that endogenous CaMKII has an activating effect on the RyR Ca2+ release channel [45-48]. However, a contrasting study shows that constitutively active CaMKII depresses RyR release [49]. Thus, the functional consequence of phosphorylation of RyR by CaMKII remains controversial. Since the bulk of the published literature on this topic points toward an activating effect of CaMKII on RyR, this concept is adopted in our model by making the rate constants in our 4-state Markovian model of the RyR channel (, , and in Figure 3B, Krishna et al. [15]) functions of the available active CaMKII. Although CaN is reported to regulate ryanodine receptor Ca2+ release channels in rat heart [50], we have refrained from modeling its influence on the RyR channel because CaN is known to be constitutively active in the dyad [2] exhibiting only minor frequency-dependent modulation in its level, hence making its rate-dependent regulatory role insignificant.
SERCA pump
In rat ventricular myocytes, 85-90% [51,52] of the systolic increase in Ca2+ in the myoplasm is recovered back into the SR stores via the sarcoplasmic reticulum Ca2+-ATPase (SERCA) pump. Frequency-dependent CaMKII activity is known to cause an acceleration of relaxation [3,23]. CaMKII affects the SERCA pump via direct phosphorylation, assisting in enhancement of SR Ca2+ transport by increasing the pumping rate [4]. This feature is incorporated in our model by letting the rate constants for Ca2+ binding to/release from the SERCA pump depend on the available active CaMKII. The SERCA pump is also indirectly affected by CaMKII via phosphorylation of unphosphorylated phospholamban (PLB), relieving the inhibition caused by PLB on the SERCA pump and thereby increasing the sensitivity of the pump for Ca2+ uptake. This indirect effect is modeled by having the rate constant for phosphorylation of PLB be a function of active CaMKII in the myoplasm. These two effects cause enhancement in SR Ca2+ uptake in an activity-dependent fashion. Expression of CaN, a protein that is a phosphatase, paradoxically has been reported to cause an increased level of PLB phosphorylation via an unknown indirect mechanism in transgenic mice. In CaN knock-out mice, decreasing phosphorylation of PLB allowed an increased level of inhibition of the SERCA pump, which resulted in poor muscle contraction and relaxation [5,6,53]. However, it is unclear if this behavior is a result of other compensatory mechanisms such as decreased CaMKII expression or enhanced PLB to SERCA ratio. On the contrary, CaN has been reported to inhibit SERCA activity in isolated non-failing human myocardium [54] in-vitro. Hence, we model the role of CaN in rate-dependent inhibition of the SERCA pump via PLB dephosphorylation by allowing the rate constant for phosphorylation of PLB to be dependent on available active CaN in the myoplasm. With increasing heart rates, β-adrenergic stimulation results in increasing levels of cAMP in vivo (Table 1 shows values for maximal β-adrenergic stimulation), which in turn phosphorylates PLB via PKA, causing enhanced uptake by the SERCA pump (Appendix A3, Equations 5-6). Together, activity-dependent recruitment of these CaMKII and cAMP-mediated effects at high frequencies counter the effect of CaN as well as decreasing cardiac cycle duration on SR refilling.
Electro-mechanics
Our model for cardiac contractile mechanics is based on the approximate model of cooperative activation and crossbridge cycling reported by Rice et al. [55] with the following modifications: (a) the first-order rate constants for the transformation of the troponin/tropomyosin regulatory complex (outside the single overlap region between the thick and thin filaments) from a crossbridge non-permitting state to a crossbridge permitting state and vice-versa are chosen as 500 s-1 and 50 s-1 respectively in order to reproduce results reported by Rice et al. [55]; (b) the β-adrenergic agonist isoproterenol (ISO) is known to cause a decrease in myofilament Ca2+ sensitivity as a result of PKA mediated phosphorylation of troponin I [51,56] at Ser23/Ser24. Specifically, a two-state Markovian model is added to allow cAMP-dependent PKA-mediated interaction between troponin I (TnI) and the Ca2+-binding regulatory site on troponin. As reported by Messer et al. [57], the unphosphorylated form of TnI (TnI u) modulates the Ca2+ affinity of the regulatory site on troponin. We model the effects of cAMP by allowing the cumulative activation rate constant for Ca2+-binding to the troponin regulatory site to be a function of unphosphorylated TnI u, the availability of which is in turn dependent on the amount of [cAMP] present (Appendix, Equations 10,12); (c) the large Q10 values used by Rice et al. (Qf app, Qh f, Qh b and Qg xb, Table 1, [55]) are decreased from 6.25 to 2.25 in order to reproduce temperature dependence of peak force developed in intact thin rat ventricular trabeculae [58]. The rate constants (Appendix, Equations 13,17) governing cross-bridge kinetics are modeled as functions of [cAMP] to reproduce stimulation frequency dependent increase in contraction and relaxation rates. Although a calmodulin (CaM) mediated pathway has been reported [59] to be responsible for modulation of myofibrillar Ca2+-sensitivity (implying a possible CaM mediated role for Ca-dependent kinases or phosphatases in regulating myofilament contractility, particularly in frequency dependent acceleration of relaxation), we refrain from modeling this effect as the molecular mechanisms involved remain unresolved.
Results
From our modeling standpoint, the dyadic coupling unit (DCU) as defined by Krishna et al. [15] is a fundamental element involved in the mechanism of CICR. These previous studies have described the control features of this unit, as well as its interaction with the SERCA pump and free sarcolemmal pumps and exchangers to achieve a homeostatic regulation of myoplasmic Ca2+ concentration. We now extend our voltage clamp studies to address the subject of frequency-dependent characteristics of CICR and begin with the study of frequency dependence of the DCU, one of the most important components of the model. All the frequency-dependent behavior discussed here is at steady state (150 cycles) unless otherwise specified. The first task is to examine the frequency dependence of the ICa,L trigger current, followed by the RyR Ca2+ channel, highlighting the rate-dependent CaM-mediated signalling involved. This analysis is followed by examining the SR Ca2+ content and the various factors controlling it in a rate-dependent manner. A quantitative study of the overall cellular Ca2+ balance is performed to highlight its rate-dependent feature. Emphasis is placed on relative roles played by the longitudinal sarcoplasmic reticulum (LSR) membrane SERCA pump and the plasma membrane Na+/ Ca2+ exchanger, as two principal Ca2+ transport routes in the maintenance of Ca2+ homeostasis. We finally examine the myoplasmic Ca2+ transient as a function of frequency with a particular interest in the rate dependence of the force-frequency response (FFR) generated by the coupled electromechanical model. This is subsequently followed by an investigation of the rate-dependent influence of cAMP-mediated β-adrenergic stimulation on the cardiac contractile response.
L-type Ca2+ current (ICa,L)
An increase in stimulation frequency from 0.5 Hz to 8 Hz, results in a frequency-dependent monotonic increase in the peak trigger current (inset in Figure 4A and Figure 4B) while slowing down the rate of decline after it reaches its maximum (note the traces corresponding to 0.5 Hz to 8 Hz in Figure 4A). The most critical mechanism involved in frequency encoding of the ICa,L channel activity is the rate-dependent change in the average level of activated-CaMKII (Figure 4C), which is known to assist CaM-mediated Ca2+-dependent facilitation (CDF). As stimulation frequency is increased from 0.5 Hz to 8 Hz, the increase in peak ICa,L(Figure 4B) closely tracks the increase in the average level of activated-CaMKII. However, beyond 8 Hz a decrease in peak is observed (Figure 4B) despite a further increase in CaMKII (Figure 4C). This occurs as a result of incomplete channel recovery at high (> 8Hz) stimulation rates, which results in a decline in peak channel open probability (Figure 4D). At low (0.5 Hz ≤ f ≤ 4.0 Hz) stimulation rates, an increase in activated CaN (85% to 97%) is also known to enhance ICa,L channel activity, whereas at higher (> 4 Hz) rates, the lack of a substantial rate-dependent increase in its average level (Figure 4C) minimizes its role in Ca2+-dependent facilitation. The maximum value attained by the open probability of the DHP-sensitive Ca2+ channel (Figure 4D) reflects the trend shown by the peak value of the trigger current over the entire range of stimulation frequencies (0.5 to 12.0 Hz) investigated. At frequencies less than 8 Hz, the upstroke velocity of ICa,L current can be seen to increase with an increase in frequency, but above 8 Hz it begins to decline (Figure 4E) due to insufficient time for full channel recovery. It is important to note that the model predicts a frequency-dependent modulation in peak ICa,L current of less than 20% over the entire frequency range (0.5 Hz to 12.0 Hz). This small modulation of peak current (Figure 4B) is far less than the percentile changes in CaMKII activation (50%, Figure 4C), due to the insufficient time for channel recovery at high (> 4 Hz) stimulation rates.
Figure 4.
ICa,L. Frequency-dependent characteristics of the DHP-sensitive Ca2+ channel ICa,L, which forms the trigger current. Voltage clamp protocol used is a 50 ms step pulse to 10 mV from a holding potential of -40 mV. (A)ICa,L at different frequencies of stimulation ranging from 0.5 Hz to 8.0 Hz. Inset gives an expanded view of the rate-dependent modulation of ICa,L over the range of frequencies investigated (B) Dependence of peak ICa,L current on frequency of stimulation (C) Frequency-dependent increase in average levels of activated auxiliary proteins CaMKII and CaN in the dyad (D) Frequency-dependent modulation of peak open probability of the DHP-sensitive Ca2+ channel. (E) Rate-dependent changes in ICa,L upstroke velocity.
RyR Ca2+ release
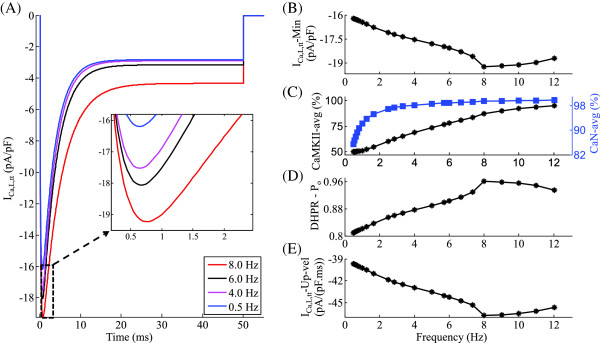
Figure 3A shows a phase plot of RyR open probability (Po) versus [ Ca2+] RyR constructed from model-generated data corresponding to different stimulation rates with the inset showing the peak open probability attained by each of the phase loops. As the stimulation frequency is increased from 0.5 Hz to 4 Hz, a marginal increase (< 1%) in peak RyR open probability occurs (inset in Figure 3A) as a result of the factors: (a) a frequency-dependent CaMKII mediated (Figure 3B) ICa,L facilitation (Figure 4A); (b) a moderate increase in SR Ca2+ content (despite a relatively constant maximal SERCA uptake rate), due to increased trigger current and the subsequent increase in [ Ca2+] myo combined with an increase in [ Na+] myo (30% in Figure 3B) which impedes Ca2+ extrusion via the NCX; and (c) a direct CaMKII-mediated enhancement in RyR release. As the stimulation frequency is increased from 4 Hz to 8 Hz, despite a moderate increase in both the trigger current and SR Ca2+ content, a small decrease in RyR open probability occurs as a result of an increase in post-release [ Ca2+] jSR (Figure 3C), which forces an incomplete luminal sensor-based RyR recovery (as described in Krishna et al. [15]). Beyond 8.0 Hz, the peak RyR open probability decreases due to two mechanisms: (a) a small parallel decrease in the trigger current (Figure 4B) indicating a close coupling enforced by a stable CICR, and (b) insufficient time for full channel recovery, accompanied by a falling pre-release diastolic jSR Ca2+ level (Figure 3C). Both the declining trigger current and the declining [ Ca2+] jSR result in a strong decline in [ Ca2+] RyR. The decrease in [ Ca2+] RyR at high (> 8 Hz) frequencies can be seen in Figure 3A, where the open probability loop for 10 Hz is enclosed within that of 8 Hz. The frequency dependence of the area enclosed by the loops (which indicates the amount of SR Ca2+ released into the dyad) mirrors that of the peak RyR open probability. The SR Ca2+ release from the Ry-sensitive receptor exhibits strong frequency-dependent behavior as shown in Figure 3D. Instantaneous RyR flux is obtained by multiplying the open probability by the concentration gradient across the channel. The rate dependence of the peak instantaneous RyR flux (Figure 3D, E) is a result of two factors: (a) the frequency dependence of peak RyR open probability (Fig 3A), which indicates the degree of recruitment of RyR release channels; and (b) the pre-release SR Ca2+ content (Figure 3C), which establishes the initial concentration gradient across the release channel. Figure 3D also shows the gradation in the time required for RyR recovery.
The frequency dependence of RyR release activation (Figure 3F), shares its characteristics with the peak RyR open probability (Figure 3A). Besides increasing trigger current, RyR release is also facilitated by increasing levels of activated-CaMKII (Figure 3E) and increasing SR Ca2+ content(Figure 3C). The frequency-dependent modulation of RyR inactivation rate (Figure 3F) mimics the rate dependence of minimum SR Ca2+ levels reached after release, mediated via the luminal sensor. With increasing stimulation frequency (0.5 Hz ≤ f ≤ 8.0 Hz), despite decreasing time available for uptake, increasing pre-release SR Ca2+ content is achieved only by increased rate of SR filling. This increased rate of SR filling translates into a faster decline in inhibition via the luminal sensor (a RyR inactivation mechanism described in Krishna et al. [15]). Hence, the rate of inactivation decreases as the frequency is increased from 0.5 Hz to 8.0 Hz (Figure 3F) owing to increasing SR Ca2+ content in this range of frequencies. With increase in frequency, the rate-dependent increase in the level of activated-CaMKII in the dyadic space also causes an increased CaMKII-mediated upregulation of the ryanodine receptor [40,41,43,45-48]. This CaMKII mediated up-regulation delays the onset of declining SR Ca2+ content-driven (8.0 Hz ≤ f ≤ 12.0 Hz) increase in the rate of RyR channel inactivation. The RyR channel experiences a frequency-dependent modulation by both the trigger current and the amount of activated-CaMKII on its dyadic side. On its luminal side, it experiences modulation by the luminal sensor (as described in Krishna et al. [15]) controlling refractoriness, and the SR Ca2+ content providing the drive for Ca2+ through the channel.
SR Ca2+ content
The frequency dependence of pre-release Ca2+ level in the SR is a result of several factors namely: (a) available [ Ca2+] myo for sequestration; (b) the rate-dependent behavior of the SERCA pump; (c) the frequency dependence of the release characteristics of Ry-sensitive Ca2+ channel; and (d) the cumulative transmembrane Ca2+ transport via ICa,L and NCX. The combined effect of these factors results in the biphasic relationship (inset in Figure 5A) between free Ca2+ content in the SR and the frequency of stimulation. As the frequency is increased from 0.5 Hz to 8.0 Hz, the SR Ca2+ content increases due to the following frequency-dependent, active CaMKII-mediated effects: (a) enhancement of maximal uptake rate of the SERCA pump due to increase in PLB phosphorylation (Figure 5B) assisted by increasing levels of activated-CaMKII (Figure 5C); (b) decrease in the half-activation constant for the forward (myoplasm to SR) operation of the SERCA pump (Figure 5D) translating into increased Ca2+ sensitivity of the pump for uptake; and (c) increase in half-activation constant for the backward (SR to myoplasm) operation of the SERCA pump (Figure 5D), reducing tendency for back-flow via the SERCA pump. The increase in SR Ca2+ content is also facilitated by a CaMKII mediated enhancement in ICa,L as well as inhibition of Ca2+ extrusion via NCX due to increasing [ Na+] myo. As the stimulation frequency is gradually increased further from 8.0 Hz to 12.0 Hz, a steep decrease in SR Ca2+ content is observed (inset in Figure 5A). This frequency-dependent decrease in pre-release SR Ca2+ content at very high (> 8.0 Hz) stimulation frequencies is a result of the following: (a) a decrease in maximal uptake rate of the SERCA pump (Figure 5B) along with a significant decrease in time available for resequestration of cytosolic Ca2+; (b) a decrease in Ca2+ entry into the cell via the trigger current ICa,L (Figure 4B); and (c) relatively constant intracellular [ Na+] myo (Figure 3B). Between stimulation frequencies 0.5 Hz and 8.0 Hz, the characteristics of the post-release SR Ca2+ level tracks the pre-release peak SR Ca2+ content. However, beyond 8.0 Hz, a declining SR Ca2+ content causes increasing inhibition on RyR release via the luminal sensor [15], resulting in a gradual increase in post-release SR Ca2+ level.
Figure 5.

[Ca2+] jSR.Mechanisms underlying the frequency-dependent modulation in SR Ca2+ content. Voltage clamp protocol used is a 50 ms step pulse to 10 mV from a holding potential of -40 mV. (A) [ Ca2+] jSR traces indicating rate-dependent changes in SR filling. The inset shows the frequency dependence of the maximum and minimum SR Ca2+ concentration. (B) Frequency dependence of the maximal pumping rate of SERCA and the average level of phosphorylated PLB. (C) Frequency-dependent increase in average levels of activated auxiliary proteins CaMKII and CaN in the myoplasm. (D) Frequency dependence of the half-activation constant for the forward and backward flow via the SERCA pump. (E) Rate dependence of the relative role of the SERCA pump (when compared to the Na+/ Ca2+ exchanger (INaCa)and the plasma membrane Ca2+ ATPase pump (IPMCA).
The frequency encoding involved in the uptake mechanism via the SERCA pump is strongly regulated by two key proteins, PLB and activated-CaMKII. The rate-dependent decrease in the level of unphosphorylated phospholamban (PLB) (Figure 5B) capable of inhibiting the SERCA pump is a result of increasing levels of activated-CaMKII (Figure 5C). It is important to note that phosphorylation of PLB is sensitive to small changes in activated-CaMKII [23]. Our results show that activated-CaMKII (Figure 5C) has a prominent role in rate-dependent modulation of the SERCA pump despite its low level (three orders of magnitude compared to activated CaN (note the 10 −5 on left ordinate in Figure 5C)) in the myoplasm [2]. Our results show that activated CaN (Figure 5C) has minimal role in rate-dependent inhibition of the SERCA pump. However, over-expression of CaN in failing heart is known to significantly compromise SERCA activity [54]. At a given frequency of stimulation, an increase in trigger current forces a gradual increase in SR Ca2+ uptake which in turn results in a corresponding increase in RyR release, which translates into an elevated cytosolic Ca2+ level and subsequently an increased activated-CaMKII level. This causes an increase in SR Ca2+ uptake rate via phosphorylation of PLB, thus completing a positive feedback loop. Hence, the peak ICa,L trigger current sets the peak SR Ca2+ content if the SERCA pump is not operating in saturation. Thus the frequency dependence of the maximal rate of uptake by the SERCA pump (Figure 5B) shares its characteristics with peak ICa,L current (Figure 4B) linked together by activated-CaMKII (Figure 5C). It is important to note that rate-dependent enhancement in SERCA activty by CaMKII significantly overrides the inhibitory influence of CaN in a non-failing heart [54]. Figure 5B also shows the underlying rate-dependent decline in unphosphorylated PLB responsible for rate-dependent increase in SR uptake.
Although marginally (< 1% change over 0.5 Hz ≤ f ≤ 12.0 Hz) frequency-dependent, the SERCA pump has a primary role in resequestration of myoplasmic Ca2+ when compared with the combined influence of Na+/ Ca2+ exchange (INaCa) as well as Ca2+ transport via the plasma membrane Ca2+ ATPase pump,(IPMCA). As the frequency of stimulation is increased from 0.5 Hz to 4.0 Hz, increasing activated-CaMKII-dependent enhancement in SERCA activity assisted by a decrease in Ca2+ extrusion via NCX due to increasing [ Na+] myo (Figure 3B) result in a marginal increase in the relative role of SERCA pump as shown in Figure 5E. However, as frequency is further increased, declining SERCA-mediated uptake activity due to a decrease in CaMKII-mediated SERCA enhancement, as well as reduced time available for uptake results in a small negative slope to the frequency dependence of the relative role of SERCA pump (Figure 5E).
Myoplasmic Ca2+ transient [Ca2+]myo
Figure 6A shows the traces of [ Ca2+] myo at increasing frequencies showing an increase in the peak myoplasmic Ca2+ transient during systole with increasing stimulation frequency in the range 0.5 Hz to 8.0 Hz. A ’primary phase’ negative peak FFR (a decrease in peak contractile force with increase in stimulation frequency observed at low (< 1.0 Hz) stimulation rates) reported in studies on rat ventricular myocytes [8,9,60,61] could be a result of low unphysiological temperatures employed in those studies. The inset in Figure 6A shows that the peak rate of rise (R rise) and decay (R decay) of the [ Ca2+] myo transient increases with the frequency of stimulation from 0.5 Hz to 8.0 Hz. The increase in rate of rise is a result of an increase in peak trigger current ICa,L (Figure 4B) and SR Ca2+ content (inset in Figure 5A) while the increase in rate of decay is a result of CaMKII-mediated acceleration of relaxation (Figure 5C) due to a rate-dependent enhancement in uptake by the SERCA pump. An opposite effect is observed in the rate of rise and decay of the [ Ca2+] myo transient beyond a stimulation rate of 8.0 Hz due to declining peak trigger current and reduced CaMKII activity. As shown in Figure 6B, increasing stimulation rate from 0.5 Hz to 8.0 Hz results in a steep increase in peak [ Ca2+] myo (maximum), which parallels a strong increase in pre-release diastolic [ Ca2+] jSR (Figure 5A). This increase in [ Ca2+] jSR is mirrored by a corresponding increase in pre-release minimum value attained by [ Ca2+] myo during diastole (Figure 6B). As the stimulation frequency is increased beyond 8.0 Hz, although the SERCA activity is saturated [12], the declining SR Ca2+ content (inset in Figure 5A) aids in the decrease of peak [ Ca2+] myo by compromising SR release. However, a lack of sufficient time for uptake continues to manifest in increasing pre-release diastolic [ Ca2+] myo (Figure 6B) at these rates. Our study suggests that a decrease in pH i causing a decrease in myofilament Ca2+ sensitivity [62] is not required to cause the ’secondary phase’ negative peak FFR (a decrease in peak contractile force with increase in stimulation frequency observed at high (> 8.0 Hz) stimulation rates). We attribute this phase which involves a decrease in force response, to the declining peak [ Ca2+] myo as seen in Figure 6B. The rate-dependence of peak [ Ca2+] myo for increasing stimulation frequencies (0.5 Hz ≤ f ≤ 12.0 Hz) is comparable to that reported by Kentish et al. (Figure 1, [63]) in rat ventricular trabeculae. Figure 6C shows the frequency-dependent changes in gain calculated as the ratio of peak Ca2+ transient in the presence of CICR to the peak calcium transient in its absence, contributed by the trigger calcium alone [64]. The EC coupling gain increases with the frequency of stimulation from 0.5 Hz to 8.0 Hz, due to rate dependent CaMKII-mediated increase in uptake rate of the SERCA pump (Figure 5B), which decreases the peak [ Ca2+] myo in the absence of CICR and increases peak [ Ca2+] myo in its presence (due to increase in SR content (inset in Figure 5A)). Beyond a stimulation frequency of 8.0 Hz, the rapid decrease in SR content causes a steep decline in EC coupling gain.
Figure 6.
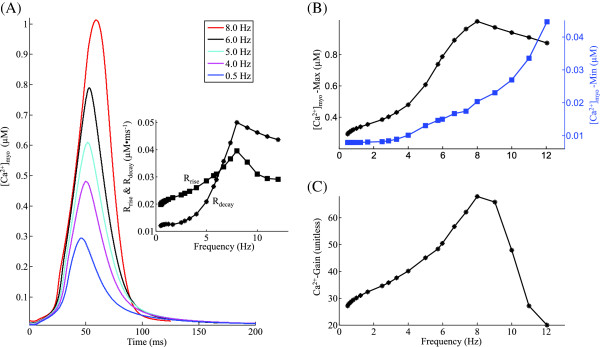
[Ca2+] myo. Frequency-dependent modulation of myoplasmic Ca2+ transient. Voltage clamp protocol used is a 50 ms step pulse to 10 mV from a holding potential of -40 mV. (A) [ Ca2+] myo traces showing rate-dependent effects on systolic Ca2+ concentration in the myoplasm for increasing stimulation frequencies from 0.5 Hz to 8.0 Hz. The inset shows the change in the maximum rate of rise and decay of the [ Ca2+] myo transient with stimulation frequency. (B) Frequency dependence of the maximum and minimum myoplasmic Ca2+ concentration. (C) Bi-phasic rate dependence of EC coupling gain.
Positive force-frequency relationship
Changing the stimulation rate has a very prominent effect on cardiac muscle contraction by way of affecting the amplitude and time course of the intracellular Ca2+ transient (Figures 6 and 7A). Here, we simulate a positive peak force-frequency response elicited using a voltage clamp stimulation protocol. Figure 7B shows the traces for the isometric contractile force generated for increasing stimulation frequencies. As stimulation frequency increases from 0.5 Hz to 8.0 Hz, peak contractile force increases (Figures 7B and C) mirroring an increase in peak [ Ca2+] myo. As seen in Figures 4B and 5B, this is mainly a result of CaMKII-dependent upregulation of ICa,L and SERCA activity which translates into a frequency-dependent increase in SR Ca2+ content, and thereby SR Ca2+ release. As reported in the literature [55], the non-linear sigmoidal steady state Force- Ca2+ relationship (overlayed in Figure 7D) causes a non-linear response in peak force developed for a corresponding change in peak [ Ca2+] myo. As shown in Figure 7D the contraction-relaxation coupling point (CRCP) moves to larger values of force and Ca2+ concentration with increase in rate of stimulation. The peak contractile force is attained at a physiological rate of 8.0 Hz, beyond which the gradual decline in peak ICa,L current (Figure 4B), which forces a corresponding decrease in SR Ca2+ content (Figure 5A), and hence RyR release (Figure 3E), causes a marginal decline in peak contractile force generated. The frequency at which peak contractile force is generated is temperature-dependent as reported by Layland et al. [12] and shifts to the right on the frequency axis as the temperature increases. The peak force frequency response obtained mimics the experimentally observed frequency dependence (Figure 1, [63]). The minimum value of contractile force per cycle (end-diastolic) increases with increasing rate of stimulation as a result of the combination of factors: (a) increasing myoplasmic Ca2+ levels due to rate-dependent increase in SR release (Fig 6); and (b) decreasing time available for resequestering released Ca2+ from the myoplasm. The frequency dependence of the force response translates into a reciprocal rate dependent characteristic of the sarcomere length as shown in Figures 7E and F.
Figure 7.

Contractile force and sarcomere length. Rate-dependent changes in myoplasmic Ca2+ concentration, the corresponding contractile force generated and the resulting sarcomere length. Voltage clamp protocol used is a 50 ms step pulse to 10 mV from a holding potential of -40 mV. (A) Frequency dependence of the maximum and minimum myoplasmic Ca2+ concentration (B) Traces for normalized isometric force developed at stimulations rates of 4, 5, 6, and 8 Hz. (C) Rate dependence of the maximum and minimum normalized force generated. (D) Phase loops for normalized isometric force vs [ Ca2+] myo for increasing frequency of stimulation (4.0 Hz to 8.0 Hz) corresponding to panel B. (E) Traces for sarcomere contraction corresponding to isometric force in panel B. (F) Frequency dependence of minimum sarcomere length achieved at peak contraction. The legend for traces in panel B is shared by panel D and E.
Effect of maximal β-adrenergic stimulation
β-adrenergic stimulation bears a very crucial physiological relevance in normal functioning of the heart. We have investigated the effect of β-adrenergic stimulation on [ Ca2+] myo under voltage clamp conditions. The degree of CaMKII mediated ICa,L facilitation is kept identical to the control case discussed earlier. Besides a moderate CaMKII-mediated [23,65] enhancement in ICa,L, we observe that increase in stimulation frequency causes a strong β-adrenergic stimulation-dependent facilitation in ICa,L, resulting in an enhanced positive slope in the FFR as the stimulation frequency is increased. These results tend to agree with studies that show that increased cAMP levels reverse the abnormalities in the FFR that are found in end-stage heart failure [66], and that moderate stimulation by isoproterenol partly reverses the negative FFR found in NYHA (New York Heart Association) class IV (severe) type heart failure and preserves the positive FFR in nonfailing myocardium [67]. It is important to note that variations in experimental protocol could result in undesirable artifacts owing to the time-sensitive nature of the effects of cAMP [68]. Here, we show that the presence of maximal β-adrenergic stimulation results in a stronger positive FFR (frequency dependence of peak contractile force generated) at physiological stimulation frequencies.
At low (< 2.5 Hz) stimulation frequencies β-adrenergic stimulation plays no role in altering intracellular Ca2+ dynamics. However, in the presence of maximal β-adrenergic stimulation (elevated levels of cAMP), an increase in stimulation rate (> 2.5 Hz) causes increased up-regulation of ICa,L[69-71]. The model predicts (Figure 8A) that as the frequency of stimulation is increased in steps, peak ICa,L magnitude increases at a regular rate, before declining at high stimulation frequencies (> 8.0 Hz) as a result of incomplete channel recovery. As shown in Figure 8B, as the frequency is increased from 2.5 Hz to 8.0 Hz, the peak SR Ca2+ content increases due to two mechanisms, namely cAMP-dependent enhancement of both trigger current and a rate-dependent increase in uptake by the SERCA pump as a result of a stronger decline in the average level of unphosphorylated PLB, which stimulates the uptake. It is important to note that although the increase in peak [ Ca2+] myo (traces in Figure 8C) causes a strong CaM-mediated CaMKII-dependent up-regulation of the SERCA pump due to an increase in PLB phosphorylation, cAMP-mediated phosphorylation of PLB causes further enhancement of the uptake mechanism. An increase in stimulation rate beyond 8.0 Hz, causes a decrease in SR Ca2+ content due to insufficient time for SR recovery along with a saturated operation of the SERCA pump. The presence of β-adrenergic stimulation significantly alters the relationship between the peak force response and the frequency of stimulation. As shown in the traces for force response in Figure 9A, besides an increase in peak force generated (a result of increased peak [ Ca2+] myo (Figures 8C and 9B)), β-adrenergic stimulation causes an increase in contraction and relaxation rate. This cAMP-mediated increase in the rate of sarcomere contraction and relaxation is a combined result of increasing rate of rise and decline of the Ca2+-transient and a cAMP-mediated enhancement in rate kinetics governing cross-bridge formation. The inset in Figure 9A is a plot of the maximum rate of relaxation (R relax) versus maximum rate of contraction (R act) for increasing stimulation frequency with and without β-adrenergic stimulation (shows significant overlap). This linear relationship highlights contraction-relaxation coupling, and represents a key intrinsic property of the contractile myofilaments [72]. As shown in Figure 9B, although β-adrenergic stimulation further assists (due to the upregulation of ICa,L and SERCA pump) a CaMKII-mediated rate-dependent increase in peak [ Ca2+] myo, CaMKII is the key factor responsible for this behavior (a 2-fold increase in peak [ Ca2+] myo from 4 Hz to 8 Hz both in the presence/absence of cAMP-mediated effects). In the presence of β-adrenergic stimulation, Figure 9C shows an enhancement in peak force generated and a decrease in minimum force as a result of a parallel trend in the peak systolic and end-diastolic levels of [ Ca2+] myo (Figure 9B) supported by faster cross-bridge kinetics. The delta increment in force due to β-adrenergic stimulation (compared to the control case) is maximum at the stimulation rate of 5 Hz (Figure 9C) and decreases both above and below this frequency. The cAMP-mediated effect on the dependence of maximum sarcomere contraction (Figure 9D) on stimulation frequency mirrors that of the peak force response (Figure 9C) showing an opposite trend.
Figure 8.

Effect of maximalβ-adrenergic stimulation onCa2+ dynamics. A comparison between cAMP-mediated maximal β-adrenergic stimulation and basal condition. Voltage clamp protocol used is a 50 ms step pulse to 10 mV from a holding potential of -40 mV. (A) Rate dependence of peak ICa,L current. (B) Frequency dependence of the maximum pre-release SR Ca2+ concentration. (C) [ Ca2+] myo traces showing rate-dependent effects on systolic Ca2+ concentration in the myoplasm for increasing stimulation frequencies from 4.0 Hz to 8.0 Hz.
Figure 9.

Effect of maximal β-adrenergic stimulation on force response. A comparison between cAMP-mediated maximal β-adrenergic stimulation and basal condition. Voltage clamp protocol used is a 50 ms step pulse to 10 mV from a holding potential of -40 mV. (A) Traces for normalized isometric force developed for increasing stimulation frequencies from 4.0 Hz to 8.0 Hz. The inset shows a plot of the maximum rate of relaxation (R relax) versus maximum rate of contraction (R act) for increasing stimulation frequency. (B) The frequency dependence of the maximum and minimum myoplasmic Ca2+ concentration. (C) Rate dependence of the maximum and minimum normalized force generated. (D) Frequency dependence of minimum sarcomere length achieved at peak contraction.
Discussion
In this study, we have examined the role of two key mechanisms: (a) CaMKII-mediated upregulation; and (b) frequency-dependent cAMP-mediated β-adrenergic stimulation on Ca2+ cycling and their effect on the force-frequency response. In particular, we have modeled their two crucial signalling mechanisms: the rate-dependent upregulation of the DHP-sensitive ICa,L channel [25,69,73,74] and SERCA pump activity [75,76], which our study suggests are the key factors responsible for the characteristics of FFR in rat ventricular myocytes. These two signalling pathways mediate ICa,L enhancement bringing an extra supply of trigger current into the dyad to both enhance CICR and assist in a rate-dependent build up in pre-release diastolic SR Ca2+ concentration. This frequency-dependent increase in SR Ca2+ content is strongly assisted by a β-adrenergic stimulation-dependent cAMP-mediated increase in PLB phosphorylation, which relieves the inhibition exerted on the SERCA pump. This is especially important in the light of decreasing time for Ca2+ uptake with increasing stimulation rate.
We have systematically analyzed the important role of Ca2+-dependent CaM-mediated function of CaMKII and CaN as rate sensors. CaMKII is known to regulate EC coupling [41,45] and has substrates in both the dyadic domain (ICa,L and Iryr) [32,41] as well as the myoplasm (phospholamban) [77], and immunoprecipitates with the ICa,L channel [32] and RyR [78]. Our study confirms that activated-CaMKII causes rate-dependent acceleration of relaxation [3] both by direct phosphorylation of the SERCA pump as well as phosphorylation of PLB resulting in a decreased inhibition of the SERCA pump. Although the time course of the indirect effect of CaMKII via PLB phosphorylation is reported to be delayed with regard to frequency-dependent acceleration of relaxation [79], the direct phosphorylation of the SERCA pump by activated-CaMKII could cause an immediate increase in maximal pumping rate [4,23]. Despite poor sensitivity of CaMKII to myoplasmic Ca2+ signals due to its relatively low affinity for CaM and an inadequate supply of myoplasmic Ca2+-CaM complex (compared to the dyad), activated-CaMKII in the cytosol plays an important role in the enhancement of uptake via the SERCA pump except at high (> 8 Hz, inset in Figure 5A) stimulation frequencies. Here, the rapid decrease in time available for uptake results in gradually declining pre-release SR Ca2+ content. CaMKII has a more critical role in the dyadic cleft where we show [2] substantial CaMKII activity owing to large local Ca2+ concentration and CaM enrichment. It is reported that frequency-dependent CaMKII activity contributes to ICa,L channel facilitation [31] and assists a positive force-frequency relationship [80]. Our study shows that CaMKII causes a significant enhancement of ICa,L even under basal conditions (absence of β-adrenergic stimulation) as the stimulation frequency is increased to 8.0 Hz (Figure 4B), beyond which incomplete recovery begins to cause a decline in peak ICa,L. A rate-dependent increase in SR Ca2+ content can be achieved only when the CaMKII-mediated (Figure 4C) increase in trigger current (Figure 4B) is supported by an increase in uptake via the SERCA pump by activated-CaMKII in the myoplasm (Figure 5C) which enhances PLB phosphorylation. CaMKII fails to sustain a positive FFR at very high (> 8.0 Hz) stimulation frequencies due to incomplete channel recovery causing a decline in trigger current and subsequently SR Ca2+ content. Data from atrial myocytes [81] show that myoplasmic CaN is also found to be responsive to increased pacing frequency.
Although CaN is also involved in ICa,L enhancement in the dyad at low (< 2.0 Hz) stimulation frequencies, at high stimulation rates (> 2.0 Hz), the large Ca2+ signals along with very slow dissociation of the Ca4CaM-CaN complex force it to be constitutively active, limiting its modulatory role. The presence of β-adrenergic stimulation, which also supports a sustained frequency-dependent increase in ICa,L (Figure 8A) and SERCA activity (Figure 8B) results in enhanced contractile response (Figure 9A and C).
Here, we have integrated an electro-mechanical model with our electrophysiological model of a rat ventricular myocyte [15] to describe isometric contractile force generated by myofibrils and investigate its rate-dependent characteristics. Our model for the cardiac contractile mechanism is derived from the description of cooperative activation and cross-bridge cycling given by Rice et al. [55]. However, we have incorporated a rate-dependent cAMP mediated enhancement in cross-bridge kinetics which is shown to be responsible for sustaining linear contraction-relaxation coupling (inset in Figure 9A) at increasing frequencies. It is important to note that the presence of CaMKII-mediated ICa,L enhancement or β-adrenergic stimulation results in physiologically relevant rate-dependent increase in peak cardiac contractile force (positive FFR).
Model limitations
1. This model of a rat ventricular myocyte is limited to Ca2+ related channel, exchanger and pumps (ICa,L, INaCa, IPMCA, Iryr and SERCA pump), while lacking exclusive Na+ or K+ related channels and transporters. Hence, it is aimed at mimicking voltage clamp conditions where channels other than calcium are blocked, and it cannot be used to study any action potential-induced Ca2+ transient-frequency relationship. However, its focus on the Ca2+ dynamics alone allows one to comprehend more clearly the important role of various Ca2+-dependent regulatory proteins such as CaMKII, CaN, PLB and cAMP in affecting multiple targets and thus generating a cell’s response to a change in the frequency of stimulation.
2. It is known that CaMKII alters the function of numerous ion channels and Ca2+ regulatory targets in a rate-dependent fashion. However, disparate findings exist on its modulation of targets such as the SERCA [82,83] and RyR channel [40,44,45]. Although our model aligns with rate-dependent CaN-mediated inhibition of the SERCA pump, its role in modulating SERCA activity is controversial due to conflicting findings [53,54]. Similarly, the effect of β-adrenergic stimulation via cAMP particularly, its dose-dependent influence on L-Type Ca2+ channels both in terms of modifying the single channel behavior such as Ca2+ ion permeability (channel conductance) as well as overall channel recruitment characteristics (open probability) is not clearly understood. While cAMP has been shown to increase channel open probability [35,36], increased levels of cAMP also result in increased phosphorylation of L-type Ca2+ channels, causing an increased permeability to Ca2+ ions [36-38]. Although we have modeled enhancement in ICa,L due to beta-adrenergic stimulation solely by a rate-dependent cAMP-mediated increase in channel conductance, the relative contribution of these factors to cAMP-dependent ICa,L enhancement has to be examined further. A detailed investigation is required to clarify the nature of these interactions. We hope that this study would help motivate more pointed experimental investigation of frequency-dependent CaMKII, CaN and cAMP effects on FFR in rat ventricular myocytes.
3. The effect of β-adrenergic stimulation on cardiac Na+/ Ca2+ exchange has been controversial. Perchenet et al. [84] report an enhancement in Na+/ Ca2+ exchange by the β-adrenergic/PKA-mediated phosphorylation of the exchanger protein. However, a cAMP-mediated enhancement in NCX activity would impede rate dependent increase in SR Ca2+ content. We base our model for the Na+/ Ca2+ exchanger on a more recent study by Lin et al. [85] which supports the view that β-adrenergic stimulation does not upregulate Na+/ Ca2+ exchange current.
4. The cooperative activation of the thin filament and the strain-dependent transitions of the cross-bridge cycle have been approximately modeled as non-spatial, state-variables. However, this simplification is valid as these transitions are inherently local phenomena and the model reproduces a wide range of steady state and dynamic responses in cardiac muscle [55].
Conclusion
Using a mathematical model of an isolated rat ventricular myocyte in a voltage clamp setting, we have systematically examined the issue of rate-dependence in the proper functioning of the dyadic coupling unit, the regulation of SERCA function to provide adequate SR Ca2+ content, the peak amplitude of the myoplasmic Ca2+ transient and the complex interaction of all these factors. Given the complexity of these interacting systems, computer modeling gives an insight into the relative roles of different Ca2+ transport mechanisms. Our simulations explain the Ca2+-dependent, CaM-mediated, rate sensitive effects of CaMKII and CaN on various intracellular targets. We also investigate a significant, frequency-dependent, cAMP-mediated effect of β-adrenergic stimulation and its modulatory influence on the ICa,L channel as well as the SERCA pump. Rate-dependent CaMKII mediated ICa,L facilitation as well as cAMP-dependent upregulation of intracellular targets could play a vital role in reversing the negative FFR found in failing hearts. However, further studies are required to develop a clear understanding of the relative role of CaMKII and cAMP in the rate-dependent up-regulation of various intra-cellular targets especially the DHP-sensitive ICa,L channel. This would help in assigning rate-dependent weights to these signalling pathways. One could use KN-93 and autocamtide-2 related inhibitory peptide [86] for a study to delineate these effects. Our coupled electro-physiological and electro-mechanical model also sheds light on the rate dependence of the cardiac contractile mechanism. In particular, our model accounts for cAMP-dependent modulation of the rate kinetics governing cross-bridge formation. In agreement with Janssen [87], we also demonstrate a key linear relationship between the rate of contraction and relaxation, which is shown here to be intrinsically coupled over the full range of physiological frequencies both in the absence/presence of β-adrenergic stimulation. This study provides mechanistic, biophysically based explanations for the rate-dependent Ca2+ signalling underlying the force-frequency response in rat ventricular myocytes, generating useful and testable hypotheses.
Appendix
Below is the set of equations modified in the model.
Equations for currents modified (from Krishna et al. [15]) in the model
L-Type Ca2+ current
Ca2+ current through the DHP-sensitive ICa,L channel
| (1) |
| (2) |
| (3) |
| (4) |
§- ICa,L described in Krishna et al. [15]. ‡- ξ described in Krishna et al. [15] causes rate-dependent modulation of the ICa,L channel.
Uptake of Ca2+ from the cytosol into the LSR
Differential equation for phospholamban phosphorylation
| (5) |
| (6) |
| (7) |
=6800 s −1 ; =1000 s −1; (Krishna et al. [15]).
Equations governing electro-mechanics modified (from Rice et al. [55]) in the model
Regulatory Ca 2+-binding to troponin
| (8) |
| (9) |
k onT = 22.22 μM−1s−1 ; k offHT = 17.36 s −1; k offLT = 173.61 s −1; (Rice et al. [55]).
| (10) |
k onTI = 698.69 s −1 ; k offTI = 80.0 s −1; (estimated from Roof et al. [88]).
| (11) |
| (12) |
| (13) |
| (14) |
| (15) |
| (16) |
| (17) |
‡- Rate constants governing cross-bridge kinetics described in Rice et al. [55].
Abbreviations
[Ca2+]: calcium ion concentration; [Ca2+]jSR: luminal Ca2+ concentration in the jSR; [Ca2+]myo: myoplasmic Ca2+ concentration; [Ca2+]o: extracellular Ca2+ concentration; [Ca2+]ryr: Ca2+ concentration at the “mouth” of the RyR channel on the dyadic side; CaM: calmodulin; CaMKII: Ca2+/calmodulin-dependent protein kinase II; CaMKIIact: activated Ca2+/calmodulin-dependent protein kinase II; cAMP: cyclic adenosine monophosphate; CaN: calcineurin; CaNact: activated calcineurin; CDF: calcium-dependent facilitation; CDI: calcium-dependent inactivation; CICR: calcium-induced calcium-release; CRCP: contraction-relaxation coupling point; DCU: dyadic coupling unit; DHP: dihydropyridine; DHPR: dihydropyridine receptor; E-C: excitation contraction; EC50bwd: affinity of backward Ca2+ flux from LSR to myoplasm; : ; ICa,L: L-type Ca2+ current; Icyt,serca: Ca2+ uptake current directed from the myoplasm to the SERCA; INa,b: background sodium current; INaCa: sodium calcium exchanger current; INaCs: sodium cesium pump current; IPMCA: plasma membrane Ca2+ ATPase pump current; Iryr: Ca2+ current due to CICR from an individual jSR; Iserca,sr: Ca2+ uptake current directed from the SERCA to the LSR; jSR: junctional portion of the sarcoplasmic reticulum; LCC: L-type DHP-sensitive Ca2+ channel; L-type: long lasting type; LSR: longitudinal portion of the sarcoplasmic reticulum; mM: milli molar; mV: milli volt; [Na+]myo: myoplasmic Na+ concentration; NCX: Na+/Ca2+ exchanger; nM: nano molar; NO: Nitric oxide; pC: pico coulomb; Po: Open probability; PKA: protein kinase A; PLB: phospholamban; : ; PLBp: phosphorylated phospholamban; PSR: phospholamban to SERCA ratio; Ry: ryanodine; RyR: ryanodine receptor; SERCA: sarcoplasmic reticulum Ca2+ ATPase; SL: sarcolemma; SR: sarcoplasmic reticulum; VC: voltage clamp; VDI: voltage-dependent inactivation.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
AK developed the coupled electromechanical model of the rat ventricular myocyte, carried out the voltage clamp modeling studies, and drafted the manuscript. MV made substantial intellectual contributions to the study and in drafting of the manuscript. PTP made intellectual contributions to the manuscript as well as significant contributions to the drafting of the manuscript. JWC made key contributions to the conception of the study, design, analysis and interpretation of results, and drafting of the manuscript. All authors read and approved the final manuscript.
Contributor Information
Abhilash Krishna, Email: abhilash@rice.edu.
Miguel Valderrábano, Email: MValderrabano@tmhs.org.
Philip T Palade, Email: PPalade@uams.edu.
John W Clark, Jr, Email: jwc@rice.edu.
Acknowledgements
This work was supported by a research grant from Methodist Hospital Research Institute. The authors would like to thank Liang Sun for his early contributions to this work.
References
- Fabiato A, Fabiato F. Contractions induced by a calcium-triggered release of calcium from the sarcoplasmic reticulum of single skinned cardiac cells. J Physiol. 1975;249:469–495. doi: 10.1113/jphysiol.1975.sp011026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saucerman JJ, Bers DM. Calmodulin mediates differential sensitivity of CaMKII and Calcineurin to local Ca2+ in Cardiac Myocytes. Biophys J. 2008;95:4597–4612. doi: 10.1529/biophysj.108.128728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassani RA, Mattiazzi A, Bers DM. CaMKII is responsible for activity-dependent acceleration of relaxation in rat ventricular myocytes. Am J Physiol. 1995;268:703–712. doi: 10.1152/ajpheart.1995.268.2.H703. [DOI] [PubMed] [Google Scholar]
- Xu A, Hawkins C, Narayanan N. Phosphorylation and activation of the Ca2+-pumping ATPase of cardiac sarcoplasmic reticulum by Ca2+/calmodulin-dependent protein kinase. J Biol Chem. 1993;268(12):8394–8397. [PubMed] [Google Scholar]
- Wolska BM. Calcineurin and cardiac function: is more or less better for the heart? Am J Physiol Heart Circ Physiol. 2009;297:H1576–H1577. doi: 10.1152/ajpheart.00833.2009. [DOI] [PubMed] [Google Scholar]
- Schaeffer PJ, Desantiago J, Yang J, Flagg TP, Kovacs A, Weinheimer CJ, Courtois M, Leone TC, Nichols CG, Bers DM, Kelly DP. Impaired contractile function and calcium handling in hearts of cardiac-specific calcineurin b1-deficient mice. Am J Physiol Heart Circ Physiol. 2009;297:1263–1273. doi: 10.1152/ajpheart.00152.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowditch HP. Ueber die Eigenthümlichkeiten der Reizbarkeit, welche die Muskelfasern des Herzens zeigen. Arb Physiol Anst Leipzig. 1871;6:139–176. [Google Scholar]
- Schouten VJ, ter Keurs H E. Role of Ica and Na+/Ca2+ exchange in the force-frequency relationship of rat heart muscle. J Mol Cell Cardiol. 1991;23(9):1039–1050. doi: 10.1016/0022-2828(91)91639-9. [DOI] [PubMed] [Google Scholar]
- Vornanen M. Force-frequency relationship, contraction duration and recirculating fraction of calcium in postnatally developing rat heart ventricles: correlation with heart rate. Acta Physiol Scand. 1992;145(4):311–321. doi: 10.1111/j.1748-1716.1992.tb09371.x. [DOI] [PubMed] [Google Scholar]
- Jouannot P, Hatt PY. Rat myocardial mechanics during pressure-induced hypertrophy development and reversal. Am J Physiol. 1975;229(2):355–364. doi: 10.1152/ajplegacy.1975.229.2.355. [DOI] [PubMed] [Google Scholar]
- Maier LS, Bers DM, Pieske B. Differences in Ca2+-handling and sarcoplasmic reticulum Ca2+-content in isolated rat and rabbit myocardium. J Mol Cell Cardiol. 2000;32(12):2249–2258. doi: 10.1006/jmcc.2000.1252. [DOI] [PubMed] [Google Scholar]
- Layland J, Kentish JC. Positive force- and [Ca2+]i-frequency relationships in rat ventricular trabeculae at physiological frequencies. Am J Physiol. 1999;276:H9–H18. doi: 10.1152/ajpheart.1999.276.1.H9. [DOI] [PubMed] [Google Scholar]
- Monasky MM, Janssen PM. The positive force-frequency relationship is maintained in absence of sarcoplasmic reticulum function in rabbit, but not in rat myocardium. J Comp Physiol B. 2009;179(4):469–479. doi: 10.1007/s00360-008-0331-3. [DOI] [PubMed] [Google Scholar]
- Butcher JC. The Numerical Analysis of Ordinary Differential Equations: Runge-Kutta and General Linear Methods. Chichester: John Wiley and Sons; 1987. [Google Scholar]
- Krishna A, Sun L, Valderrábano M, Palade PT, Clark JWJ. Modeling CICR in rat ventricular myocytes: voltage clamp studies. Theor Biol Med Model. 2010;6:199–222. doi: 10.1186/1742-4682-7-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naraghi M, Neher E. Linearized buffered Ca2+ diffusion in microdomains and its implications for calculation of [Ca2+] at the mouth of a calcium channel. J Neurosci. 1997;17:6961–6973. doi: 10.1523/JNEUROSCI.17-18-06961.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simon SM, Llinás RR. Compartmentalization of the submembrane calcium activity during calcium influx and its significance in transmitter release. Biophys J. 1985;48:485–498. doi: 10.1016/S0006-3495(85)83804-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shirokova N, Garcia J, Pizarro G, Rios E. Ca2+ release from the sarcoplasmic reticulum compared in amphibian and mammalian skeletal muscle. J Gen Physiol. 1996;107:1–18. doi: 10.1085/jgp.107.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crank J. The Mathematics of Diffusion. Oxford, London: Clarendon Press; 1975. [Google Scholar]
- Smith GD, Keizer JE, Stern MD, Lederer WJ, Cheng H. A simple numerical model of calcium spark formation and detection in cardiac myocytes. Biophys J. 1998;75:15–32. doi: 10.1016/S0006-3495(98)77491-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marquardt DW. An algorithm for least-squares estimation of nonlinear parameters. J Soc Ind Appl Math. 1963;11:431–441. doi: 10.1137/0111030. [DOI] [Google Scholar]
- Persechini A, Stemmer PM. Calmodulin is a limiting factor in the cell. Trends Cardiovasc Med. 2002;12:32–37. doi: 10.1016/S1050-1738(01)00144-X. [DOI] [PubMed] [Google Scholar]
- Picht E, DeSantiago J, Huke S, Kaetzel MA, Dedman JR, Bers DM. CaMKII inhibition targeted to the sarcoplasmic reticulum inhibits frequency-dependent acceleration of relaxation and Ca2+ current facilitation. J Mol Cell Cardiol. 2007;42:196–205. doi: 10.1016/j.yjmcc.2006.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu X, Bers DM. Free and bound intracellular calmodulin measurements in cardiac myocytes. Cell Calcium. 2007;41:353–364. doi: 10.1016/j.ceca.2006.07.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Demir SS, Clark JW, Giles WR. Parasympathetic modulation of sinoatrial node pacemaker activity in rabbit heart: a unifying model. Am J Physiol. 1999;276:2221–2244. doi: 10.1152/ajpheart.1999.276.6.H2221. [DOI] [PubMed] [Google Scholar]
- Yang J, Clark JW, Bryan RM, Robertson CS. Mathematical modeling of the nitric oxide/cGMP pathway in the vascular smooth muscle cell. Am J Physiol Heart Circ Physiol. 2005;289(2):886–897. doi: 10.1152/ajpheart.00216.2004. [DOI] [PubMed] [Google Scholar]
- Prabhu SD, Azimi A, Frosto T. Nitric oxide effects on myocardial function and force-interval relations: regulation of twitch duration. J Mol Cell Cardiol. 1999;31(12):2077–2085. doi: 10.1006/jmcc.1999.1038. [DOI] [PubMed] [Google Scholar]
- Mori MX, Erickson MG, Yue DT. Functional stoichiometry and local enrichment of calmodulin interacting with Ca2+ channels. Science. 2004;304:394–395. doi: 10.1126/science.1097507. [DOI] [PubMed] [Google Scholar]
- Pitt GS, Zühlke RD, Hudmon A, Schulman H, Reuter H, Tsien RW. Molecular basis of calmodulin tethering and Ca2+-dependent inactivation of L-type Ca2+ channels. J Biol Chem. 2001;276(33):30794–30802. doi: 10.1074/jbc.M104959200. [DOI] [PubMed] [Google Scholar]
- Soldatov NM. Ca2+ channel moving tail: link between Ca2+-induced inactivation and Ca2+ signal transduction. TRENDS Pharmacol Sci. 2003;24(4):167–171. doi: 10.1016/S0165-6147(03)00065-8. [DOI] [PubMed] [Google Scholar]
- Yuan W, Bers DM. Ca-dependent facilitation of cardiac Ca current is due to Ca-calmodulin-dpenedent protein kinase. Am J Physiol. 1994;267:H082–H993. doi: 10.1152/ajpheart.1994.267.3.H982. [DOI] [PubMed] [Google Scholar]
- Hudmon A, Schulman H, Kim J, Maltez M, Tsien RW, Pitt GS. CaMKII tethers to L-type Ca2+ channels, establishing a local and dedicated integrator of Ca2+ signals for facilitation. J Cell Biol. 2005;171:537–547. doi: 10.1083/jcb.200505155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tandan S, Wang Y, Wang TT, Jiang N, Hall DD, Hell JW, Luo X, Rothermel BA, Hill JA. Physical and functional interaction between Calcineurin and the cardiac L-type Ca2+ channel. Circ Res. 2009;105:51–60. doi: 10.1161/CIRCRESAHA.109.199828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hryshko LV, Bers DM. Ca current facilitation during postrest recovery depends on Ca entry. Am J Physiol. 1990;259:951–961. doi: 10.1152/ajpheart.1990.259.3.H951. [DOI] [PubMed] [Google Scholar]
- Yue DT, Herzig S, Marban E. β-Adrenergic stimulation of calcium channels occurs by potentiation of high-activity gating modes. Proc Natl Acad Sci. 1990;87:753–757. doi: 10.1073/pnas.87.2.753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klein G, Schröder F, Vogler D, Schaefer A, Haverich A, Schieffer B, Korte T, Drexler H. Increased open probability of single cardiac L-type calcium channels in patients with chronic atrial fibrillation: Role of phosphatase 2A. Cardiovascular Res. 2003;59:37–45. doi: 10.1016/S0008-6363(03)00357-2. [DOI] [PubMed] [Google Scholar]
- Salhanick SD, Shannon MW. Management of calcium channel antagonist overdose. Drug Saf. 2003;26:65–79. doi: 10.2165/00002018-200326020-00001. [DOI] [PubMed] [Google Scholar]
- Klabunde RE. Cardiovascular Physiology Concepts, 1st edition. 351 West Camden St., Baltimore, MD 21201: Lippincott Williams & Wilkins; 2004. Cellular structure and function; pp. 41–58. [Google Scholar]
- Balshaw DM, Xu L, Yamaguchi N, Pasek DA, Meissner G. Calmodulin binding and inhibition of cardiac muscle calcium release channel (ryanodine receptor) J Biol Chem. 2001;276:20144–20153. doi: 10.1074/jbc.M010771200. [DOI] [PubMed] [Google Scholar]
- Witcher DR, Kovacs RJ, Schulman H, Cefali DC, Jones LR. Unique phosphorylation site on the cardiac ryanodine receptor regulates calcium channel activity. J Biol Chem. 1991;266(17):11144–11152. [PubMed] [Google Scholar]
- Wehrens XH, Lehnart SE, Reiken SR, Marks AR. Ca2+/calmodulin-dependent protein kinase II phosphorylation regulates the cardiac ryanodine receptor. Circ Res. 2004;94(6):61–70. doi: 10.1161/01.RES.0000125626.33738.E2. [DOI] [PubMed] [Google Scholar]
- Rodriguez P, Bhogal MS, Colyer J. Stoichiometric phosphorylation of cardiac ryanodine receptor on serine 2809 by calmodulin-dependent kinase II and protein kinase A. J Biol Chem. 2003;278:38593–38600. doi: 10.1074/jbc.C301180200. [DOI] [PubMed] [Google Scholar]
- Hain J, Onoue H, Mayrleitner M, Fleischer S, Schindler H. Phosphorylation modulates the function of the calcium release channel of sarcoplasmic reticulum from cardiac muscle. J Biol Chem. 1995;270(5):2074–2081. doi: 10.1074/jbc.270.5.2074. [DOI] [PubMed] [Google Scholar]
- Lokuta AJ, Rogers TB, Lederer WJ, Valdivia HH. Modulation of cardiac ryanodine receptors of swine and rabbit by a phosphorylation-dephosphorylation mechanism. J Physiol. 1995;487:609–622. doi: 10.1113/jphysiol.1995.sp020904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Satoh H, Ginsburg KS, Bers DM. The effect of Ca2+-calmodulin-dependent protein kinase II on cardiac excitation-contraction coupling in ferret ventricular myocytes. J Physiol. 1997;501:17–32. doi: 10.1111/j.1469-7793.1997.017bo.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maier LS, Zhang T, Chen L, DeSantiago J, Brown JH, Bers DM. Transgenic CaMKIIδC overexpression uniquely alters cardiac myocyte Ca2+ handling: reduced SR Ca2+ load and activated SR Ca2+ release. Circ Res. 2003;92:904–911. doi: 10.1161/01.RES.0000069685.20258.F1. [DOI] [PubMed] [Google Scholar]
- Currie S, Loughrey CM, Craig MA, Smith GL. Calcium/calmodulin-dependent protein kinase IIdelta associates with the ryanodine receptor complex and regulates channel function in rabbit heart. Biochem J. 2004;377:357–366. doi: 10.1042/BJ20031043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo T, Zhang T, Mestril R, Bers DM. Ca2+/Calmodulin-dependent protein Kinase II phosphorylation of Ryanodine receptor does affect calcium sparks in mouse ventricular Myocytes. Circ Res. 2006;99:398–406. doi: 10.1161/01.RES.0000236756.06252.13. [DOI] [PubMed] [Google Scholar]
- Yang D, Zhu WZ, Xiao B, Brochet DX, Chen SR, Lakatta EG, Xiao RP, Cheng H. Ca2+/calmodulin kinase II-dependent phosphorylation of ryanodine receptors suppresses Ca2+ sparks and Ca2+ waves in cardiac myocytes. Circ Res. 2007;100(3):399–407. doi: 10.1161/01.RES.0000258022.13090.55. [DOI] [PubMed] [Google Scholar]
- Bandyopadhyay A, Shin DW, Ahn JO, Kim DH. Calcineurin regulates ryanodine receptor/Ca2+-release channels in rat heart. Biochem J. 2000;352:61–70. doi: 10.1042/0264-6021:3520061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bers DM. Excitation-Contraction Coupling and Cardiac Contractile Force. Dordrecht: Kluwer Academic; 2001. [Google Scholar]
- Negretti N, O’Neill SC, Eisner DA. The effects of inhibitors of sarcoplasmic reticulum function on the systolic Ca2+ transient in rat ventricular myocytes. J Physiol. 1993;468:35–52. doi: 10.1113/jphysiol.1993.sp019758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu G, Carr AN, Young KB, Lester JW, Yatani A, Sanbe A, Colbert MC, Schwartz SM, Frank KF, Lampe PD, Robbins J, Molkentin JD, Kranias EG. Enhanced myocyte contractility and Ca2+ handling in a calcineurin transgenic model of heart failure. Cardiovasc Res. 2002;54:105–116. doi: 10.1016/S0008-6363(02)00230-4. [DOI] [PubMed] [Google Scholar]
- Münch G, Bölck B, Karczewski P, Schwinger RH. Evidence for calcineurin-mediated regulation of SERCA 2a activity in human myocardium. J Mol Cell Cardiol. 2002;34(3):321–334. doi: 10.1006/jmcc.2001.1515. [DOI] [PubMed] [Google Scholar]
- Rice JJ, Wang F, Bers DM, de Tombe PP. Approximate model of cooperative activation and crossbridge cycling in cardiac muscle using ordinary differential equations. Biophys J. 2008;95(5):2368–2390. doi: 10.1529/biophysj.107.119487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varian KD, Raman S, Janssen PM. Measurement of myofilament calcium sensitivity at physiological temperature in intact cardiac trabeculae. Am J Physiol Heart Circ Physiol. 2006;290(5):H2092—H2097. doi: 10.1152/ajpheart.01241.2005. [DOI] [PubMed] [Google Scholar]
- Messer AE, Jacques AM, Marston SB. Troponin phosphorylation and regulatory function in human heart muscle: dephosphorylation of Ser23/24 on troponin I could account for the contractile defect in end-stage heart failure. J Mol Cell Cardiol. 2007;42:247–259. doi: 10.1016/j.yjmcc.2006.08.017. [DOI] [PubMed] [Google Scholar]
- Janssen PM, Stull LB, Marbán E. Myofilament properties comprise the rate-limiting step for cardiac relaxation at body temperature in the rat. Am J Physiol Heart Circ Physiol. 2002;282:H499–H507. doi: 10.1152/ajpheart.00595.2001. [DOI] [PubMed] [Google Scholar]
- Suematsu N, Satoh S, Ueda Y, Makino N. Effects of calmodulin and okadaic acid on myofibrillar Ca2+ sensitivity in cardiac myocytes. Basic Res Cardiol. 2002;97(2):137–144. doi: 10.1007/s003950200004. [DOI] [PubMed] [Google Scholar]
- Mubagwa K, Lin W, Sipido K, Bosteels S, Flameng W. Monensin-induced reversal of positive force-frequency relationship in cardiac muscle: role of intracellular sodium in rest-dependent potentiation of contraction. J Mol Cell Cardiol. 1997;29(3):977–989. doi: 10.1006/jmcc.1996.0342. [DOI] [PubMed] [Google Scholar]
- Orchard CH, Lakatta EG. Intracellular calcium transients and developed tension in rat heart muscle: a mechanism for the negative interval strength relationship. J Gen Physiol. 1985;86:637–651. doi: 10.1085/jgp.86.5.637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morii I, Kihara Y, Konishi T, Inubushi T, Sasayama S. Mechanism of the negative force-frequency relationship in physiologically intact rat ventricular myocardium–studies by intracellular Ca2+ monitor with indo-1 and by 31P-nuclear magnetic resonance spectroscopy. Jpn Circ J. 1996;60(8):593–603. doi: 10.1253/jcj.60.593. [DOI] [PubMed] [Google Scholar]
- Layland J, Kentish JC. Positive force- and [Ca2+]i-frequency relationships in rat ventricular trabeculae at physiological frequencies. Am J Physiol. 1999;276:H9–H18. doi: 10.1152/ajpheart.1999.276.1.H9. [DOI] [PubMed] [Google Scholar]
- Stern MD. Theory of excitation-contraction coupling in cardiac muscle. Biophys J. 1992;63:497–517. doi: 10.1016/S0006-3495(92)81615-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo J, Duff HJ. Calmodulin kinase II accelerates L-type Ca2+ current recovery from inactivation and compensates for the direct inhibitory effect of [Ca2+]i in rat ventricular myocytes. J Physiol. 2006;574:509–518. doi: 10.1113/jphysiol.2006.109199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phillips PJ, Gwathmey JK, Feldman MD, Schoen FJ, Grossman W, Morgan JP. Post-extrasystolic potentiation and the force-frequency relationship: differential augmentation of myocardial contractility in working myocardium from patients with end-stage heart failure. J Mol Cell Cardiol. 1990;22:99–110. doi: 10.1016/0022-2828(90)90975-8. [DOI] [PubMed] [Google Scholar]
- Schwinger RH, Böhm M, Müller-Ehmsen J, Uhlmann R, Schmidt U, Stäblein A, Überfuhr P, Kreuzer E, Reichart B, Eissner HJ. et al. Effect of inotropic stimulation on the negative force-frequency relationship in the failing human heart. J Mol Cell Cardiol. 1993;23(9):1039–1050. doi: 10.1161/01.cir.88.5.2267. [DOI] [PubMed] [Google Scholar]
- Giesen J, Sondermann M, Juengling E, Kammermeier H. Time dependent partial loss of the effects of isoproterenol on function and energy metabolism of isolated rat hearts. Basic Res Cardiol. 1980;75:515–525. doi: 10.1007/BF01907833. [DOI] [PubMed] [Google Scholar]
- Lemaire S, Piot C, Leclercq F, Leuranguer V, Nargeot J, Richard S. Heart rate as a determinant of L-type Ca2+ channel activity: mechanisms and implication in force-frequency relation. Basic Res Cardiol. 1998;93:51–59. doi: 10.1007/pl00007389. [DOI] [PubMed] [Google Scholar]
- Liu SJ, Zhou W, Kennedy RH. Suppression of beta-adrenergic responsiveness of L-type Ca2+ current by IL-1 beta in rat ventricular myocytes. Am J Physiol-Heart Circ Physiol. 1996;276:H141–H148. doi: 10.1152/ajpheart.1999.276.1.H141. [DOI] [PubMed] [Google Scholar]
- Tiaho F, Piot C, Nargeot J, Richard S. Regulation of the frequency-dependent facilitation of L-type Ca2+ currents in rat ventricular myocytes. J Physiol. 1994;477:237–251. doi: 10.1113/jphysiol.1994.sp020187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janssen PM. Kinetics of cardiac muscle contraction and relaxation are linked and determined by properties of the cardiac sarcomere. Am J Physiol Heart Circ Physiol. 2010;299:H1092–H1099. doi: 10.1152/ajpheart.00417.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delpech N, Soustre H, Potreau D. Antagonism of beta-adrenergic stimulation of L-type Ca2+ current by endothelin in guinea-pig atrial cells. Eur J Pharmacol. 1995;285:217–220. doi: 10.1016/0014-2999(95)00517-O. [DOI] [PubMed] [Google Scholar]
- Nagykáldi Z, Kem D, Lazzara R. Szabó B. The coupling of canine ventricular myocyte beta2-adrenoceptors to L-type calcium current. Acta Pharm Hung. 1999;69:247–257. [PubMed] [Google Scholar]
- Schwinger RH, Münch G, Bölck B, Karczewski P, Krause EG, Erdmann E. Reduced Ca2+-sensitivity of SERCA 2a in failing human myocardium due to reduced serin-16 phospholamban phosphorylation. J Mol Cell Cardiol. 1999;31(3):479–491. doi: 10.1006/jmcc.1998.0897. [DOI] [PubMed] [Google Scholar]
- Ait-Mamar B, Cailleret M, Rucker-Martin C, Bouabdallah A, Candiani G, Adamy C, Duvaldestin P, Pecker F, Defer N, Pavoine C. The cytosolic phospholipase A2 pathway, a safeguard of beta2-adrenergic cardiac effects in rat. J Biol Chem. 2005;280:18881–18890. doi: 10.1074/jbc.M410305200. [DOI] [PubMed] [Google Scholar]
- Maier LS, Bers DM. Role of Ca2+/calmodulin-dependent protein kinase (CaMK) in excitation-contraction coupling in the heart. Cardiovasc Res. 2007;73:631–640. doi: 10.1016/j.cardiores.2006.11.005. [DOI] [PubMed] [Google Scholar]
- Zhang T, Maier LS, Dalton ND, Miyamoto S, Ross JJ, Bers DM, Brown JH. The δC isoform of CaMKII is activated in cardiac hypertrophy and induces dilated cardiomyopathy and heart failure. Circ Res. 2003;92:912–919. doi: 10.1161/01.RES.0000069686.31472.C5. [DOI] [PubMed] [Google Scholar]
- Huke S, Bers DM. Temporal dissociation of frequency-dependent acceleration of relaxation and protein phosphorylation by CaMKII. J Mol Cell Cardiol. 2007;42(3):590–599. doi: 10.1016/j.yjmcc.2006.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hund TJ, Rudy Y. Rate dependence and regulation of action potential and calcium transient in a canine cardiac ventricular cell model. Circulation. 2004;110:3168–3174. doi: 10.1161/01.CIR.0000147231.69595.D3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tavi P, Pikkarainen S, Ronkainen J, Ilves M, Weckström M, Vuolteenaho O, Bruton J, Westerblad H, Ruskoaho H. Niemelä P. Pacing-induced calcineurin activation controls cardiac Ca2+ signalling and gene expression. J Physiol. 2004;554:309–320. doi: 10.1113/jphysiol.2003.053579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toyofuku T, Curotto Kurzydlowski K, Narayanan N, MacLennan DH. Identification of Ser38 as the site in cardiac sarcoplasmic reticulum Ca2+-ATPase that is phosphorylated by Ca2+/calmodulin-dependent protein kinase. J Biol Chem. 1994;269(42):26492–29496. [PubMed] [Google Scholar]
- Odermatt A, Kurzydlowski K, MacLennan DH. The vmax of the Ca2+-ATPase of cardiac sarcoplasmic reticulum (SERCA2a) is not altered by Ca2+/calmodulin-dependent phosphorylation or by interaction with phospholamban. J Biol Chem. 1996;271(24):14206–14213. doi: 10.1074/jbc.271.24.14206. [DOI] [PubMed] [Google Scholar]
- Perchenet L, Hinde AK, Patel KC, Hancox JC, Levi AJ. Stimulation of Na/Ca exchange by the beta-adrenergic/protein kinase A pathway in guinea-pig ventricular myocytes at 37 degrees C. Pflugers Arch. 2000;439:822–828. doi: 10.1007/s004240051010. [DOI] [PubMed] [Google Scholar]
- Lin X, Jo H, Sakakibara Y, Tambara K, Kim B, Komeda M, Matsuoka S. Beta-adrenergic stimulation does not activate Na+/Ca2+ exchange current in guinea pig, mouse, and rat ventricular myocytes. Am J Physiol Cell Physiol. 2006;290:C601–C608. doi: 10.1152/ajpcell.00452.2005. [DOI] [PubMed] [Google Scholar]
- DeSantiago J, Maier LS, Bers DM. Frequency-dependent acceleration of relaxation in the heart depends on CaMKII, but not phospholamban. J Mol Cell Cardiol. 2002;34(8):975–984. doi: 10.1006/jmcc.2002.2034. [DOI] [PubMed] [Google Scholar]
- Janssen PM. Myocardial contraction-relaxation coupling. Am J Physiol Heart Circ Physiol. 2010;299:H1741–H1749. doi: 10.1152/ajpheart.00759.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roof SR, Shannon TR, Janssen PM, Ziolo MT. Effects of increased systolic Ca2+ and phospholamban phosphorylation during β-adrenergic stimulation on Ca2+ transient kinetics in cardiac myocytes. Am J Physiol Heart Circ Physiol. 2011;301:H1570–H1578. doi: 10.1152/ajpheart.00402.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]