

Abstract
The vitamin A metabolite retinoic acid (RA) serves as a ligand for nuclear RA receptors that control differentiation of progenitor cells important for vertebrate development. Genetic studies in mouse embryos deficient for RA-generating enzymes have been invaluable for deciphering RA function. RA first begins to act during early organogenesis when RA generated in trunk mesoderm begins to function as a diffusible signal controlling progenitor cell differentiation. In neuroectoderm, RA functions as an instructive signal to stimulate neuronal differentiation of progenitor cells in the hindbrain and spinal cord. RA is not required for early neuronal differentiation of the forebrain, but at later stages RA stimulates neuronal differentiation in forebrain basal ganglia. RA also acts as a permissive signal for differentiation by repressing fibroblast growth factor (FGF) signaling in differentiated cells as they emerge from progenitor populations in the caudal progenitor zone and second heart field. In addition, RA signaling stimulates differentiation of spermatogonial germ cells and induces meiosis in male but not female gonads. A more complete understanding of the normal functions of RA signaling during development will guide efforts to use RA as a differentiation agent for therapeutic purposes.
Keywords: Retinoic acid signaling, FGF signaling, Embryogenesis, Progenitor cells
1. Retinoic Acid Signaling Pathway
Among the various diffusible cell-cell signaling factors that naturally direct developmental processes, retinoic acid (RA) is unique in that it is a small lipophilic molecule (M.W. 300) derived from vitamin A (retinol) (Fig. 1). Genetic studies have demonstrated that RA synthesis in mouse embryos is controlled mostly by retinol dehydrogenase-10 (RDH10) that produces retinaldehyde [1] plus three retinaldehyde dehydrogenases that generate RA (RALDH1, RALDH2, and RALDH3; also known as ALDH1A1, ALDH1A2, and ALDH1A3) [2, 3]. As Rdh10 mutants still maintain a low amount of RA synthesis [1], there must exist at least one additional retinol dehydrogenase required for RA synthesis in embryos, but this has not yet been identified. Alcohol dehydrogenases have been found to participate in adult but not embryonic RA synthesis [4], but one candidate for an additional embryonic retinol dehydrogenase is RDHE2 (SDR16C5) expressed in the neural tube [5]. Raldh1, Raldh2, and Raldh3 single or compound null mutants suggest that no other enzyme is necessary for the second step of RA synthesis prior to stage E10.5 of mouse development, but after this stage it is less clear if these three RALDHs are the only enzymes required [2, 3].
Fig. 1.
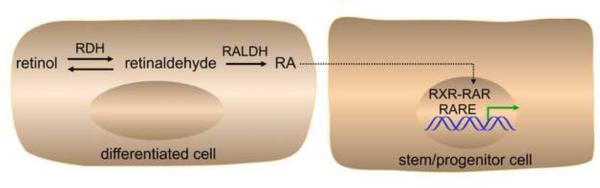
Retinoic acid signaling pathway. Some differentiated cells have the ability to metabolize retinol (vitamin A) to retinoic acid (RA). This is a two-step reaction in which retinol is first metabolized to retinaldehyde by retinol dehydrogenase (RDH) followed by metabolism of retinaldehyde to RA by retinaldehyde dehydrogenase (RALDH). RA can be secreted and taken up by other cells (including progenitor cells) to activate a paracrine signaling response. Inside the nucleus of target cells, RA functions as a ligand for the RA receptor (RAR) that binds as a heterodimer with retinoid-X receptor (RXR) to an RA response element (RARE) near a target gene. In response to binding of RA, RAR undergoes a conformational shift that alters the binding of coregulators that control transcription of RA target genes.
RA directly regulates gene transcription by entering the nucleus of target cells where it functions as a ligand for nuclear RA receptors (RARα, RARβ, RARγ) that bind target genes at a DNA sequence known as a RA response element (RARE) [6-8]. This is in contrast to other common diffusible cell-cell signaling factors such as fibroblast growth factor (FGF) [9, 10], WNT [11, 12], transforming growth factor-beta (TGF β) superfamily [13, 14], and sonic hedgehog (SHH) [15, 16] which bind cell-surface receptors and initiate intracellular signaling pathways that ultimately impact transcription in the nucleus. Also, whereas these other pathways all existed before the evolution of chordate organisms, RA signaling is unique in that it appears to be a chordate invention [17]. All chordates (including amphioxus and tunicates - primitive invertebrate chordates) possess at least one RAR that can bind and respond to the major form of RA (all-trans-RA), as well as at least one retinaldehyde dehydrogenase (RALDH) that can synthesize RA from vitamin A (Table 1). Homology searches have identified genes in non-chordate animals that encode proteins distantly related to chordate RALDHs (aldehyde dehydrogenase family) and RARs (nuclear receptor family), but there has been no demonstration of non-chordate genes that function in the synthesis of all-trans-RA or transduction of an RA signal [18]. Thus, it appears that the ability to synthesize and use RA was invented late in evolution after other cell-cell signaling pathways had evolved. In addition to RAR, PPARdelta has been identified as a non-canonical RA receptor [19] that is required for RA-induced neuronal differentiation in vitro [20].
Table 1.
Retinaldehyde dehydrogenase genes.
| common name | nomenclature | organisms with orthologs |
|---|---|---|
| Raldh1 | Aldh1a1 | mouse, rat, human, chick, frog |
| Raldh2 | Aldh1a2 | mouse, rat, human, chick, frog, zebrafish, tunicate, amphioxus |
| Raldh3 | Aldh1a3 | mouse, rat, human, chick, frog, zebrafish |
Over the years many lessons have been learned about the developmental roles of RA signaling from studies on vitamin A deficiency and RAR null mice [21-23]. Recent studies using model organisms lacking the ability to convert vitamin A to RA (either in the whole embryo or in specific tissues) have provided tremendous insight into the mechanism of RA signaling during development [24, 25]. The functions of RA during development and in the adult are still being deciphered, but it is clear that RA synthesized and secreted by certain differentiated cells plays a role in differentiation of nearby progenitor cells as further examined here (Table 2).
Table 2.
RA-mediated differentiation of progenitor cells in mouse embryos.
| RA-generating cell | Raldh gene | RA-responding progenitor cell |
|---|---|---|
| presomitic mesoderm | Raldh2 | posterior hindbrain neural progenitors |
| presomitic mesoderm or somite | Raldh2 | ventral spinal cord neural progenitors |
| lateral ganglionic eminence | Raldh3 | forebrain basal ganglia progenitors |
| presomitic mesoderm | Raldh2 | axial (neuromesodermal) progenitors |
| presomitic & lateral plate mesoderm | Raldh2 | second heart field progenitors |
| Sertoli cells | Raldh1 & Raldh2 | spermatogonia germ cells |
| premeiotic spermatocytes | Raldh2 | premeiotic spermatocytes (autocrine) |
2. Retinoic acid control of neural progenitor cell differentiation
A role for RA in neural development has been firmly established, reviewed in [26], but there has been some disagreement as to where along the central nervous system RA is required. Treatment of embryonic stem cells with supraphysiological levels of RA (1-10 μM) can induce neuronal differentiation [27, 28], suggesting to some that all neural cells require RA for differentiation. However, this is not the case as shown by several studies. First of all, the concentration of endogenous RA ranges from 1-100 nM in mouse embryonic neural tissues [29, 30], suggesting that treatment of cells or embryos with much higher levels of RA may force neuronal differentiation unnaturally. In addition, RA is not synthesized in mouse embryos until E7.5 after formation of the headfold neuroectoderm comprising the forebrain and midbrain, and although RA activity is initially detected posteriorly, RA is not detected in the head until E8.5; thus, the role of RA in early embryonic neural differentiation may be limited to the hindbrain and spinal cord that develop in posterior neuroectoderm where RA is abundant [31-33]. Loss of RA activity in the forebrain/eye region of Raldh2-/-;Raldh3-/- double mutants resulted in no defect in forebrain development up to E8.75 when these mutants were still healthy enough to compare to wild-type [34]. Also, examination of Rdh10-/- embryos that lack physiological forebrain/eye RA activity (verified using a validated RA-reporter assay), demonstrated no defects in early forebrain development [35]. In contrast, RA activity in the forebrain/eye (which is generated by enzymes in the eye) is required for normal optic cup formation [36, 37]. Thus, RA activity in the early forebrain may simply represent unnecessary spill-over from RA activity needed in the adjacent optic cups that develop as outgrowths from the forebrain neuroepithelium. However, a later role for RA in differentiation of forebrain basal ganglia has been discovered as discussed below.
Hindbrain
Raldh2 expression in mouse embryos initiates at E7.5 in presomitic mesoderm which secretes RA that can diffuse anteriorly into the developing hindbrain as far as rhombomere 3 (r3) [32]. One of the earliest targets of RA signaling during neural development are the 3’-Hox genes essential for rhombomere formation and identity that are expressed differentially along the anteroposterior axis of the posterior hindbrain beginning at E7.75 in mouse [38, 39]. Hoxb1 is required for facial motor neuron differentiation in r4 [40, 41]. Studies in chick, mouse, and zebrafish have shown that RA generated by Raldh2 is required for induction of Hoxb1 and other Hox genes in the posterior hindbrain [21, 31, 32, 42-44]. During early hindbrain development, Hoxb1 is transiently expressed throughout the entire posterior hindbrain up to the presumptive r3/r4 border, but later Hoxb1 expression is limited to r4 [45]. Transgenic mouse embryos studies have elegantly shown that Hoxb1 is directly regulated by RA through the actions of two RAREs, one located 3’ to the promoter that stimulates early widespread expression in the posterior hindbrain up to the presumptive r3/r4 boundary [46], plus another RARE positioned 5’ to the promoter that is required for repression of Hoxb1 in r3 and r5, thus helping to limit expression to r4 [47]. Although induction of Hoxb1 by RA is consistent with RA stimulating transcription by acting as a RAR ligand to recruit coactivators [48], the mechanism through which RA directly represses Hoxb1 transcription has not been established. Genetic studies in mouse and zebrafish have shown that repression of Hoxb1 posterior to r4 also requires Hnf1b (vHnf1) induced by RA signaling in neuroectoderm posterior to the r4/r5 boundary [32, 49, 50]. Hnf1b has been demonstrated to possess a functional RARE [51].
Spinal Cord
Neuroectoderm fated to become spinal cord emerges from the epiblast during body axis extension and is exposed to RA generated by Raldh2 expressed in the adjacent presomitic mesoderm or somites [33]. In the spinal cord, several studies have concluded that RA produced in mesoderm and SHH produced in notochord and floor plate both act on ventral spinal cord progenitors to stimulate a motor neuron fate [33, 52-56]. Several transcription factors are required for dorsoventral patterning of the spinal cord leading to motor neuron differentiation including Pax6 and Nkx6.1 expressed dorsally and ventrally, respectively, as well as Olig2 expressed in a region overlapping Pax6 and Nkx6.1 that marks the location where motor neurons form [57-59]. Loss of RA synthesis in mouse Raldh2-/- embryos results in loss of Pax6 and Olig2 expression in the spinal cord [33]. During motor neuron specification, RA is needed to induce Pax6 which functions with Nkx6.1 to induce Olig2 that stimulates undifferentiated ventral spinal cord neuroectoderm to acquire a motor neuron fate [55]. As neurogenin-2 (NGN2) is known to induce Pax6 in the spinal cord [60] RA may function by directly upregulating Ngn2 (which has a RARE) that then induces Pax6 [61].
Our present understanding of the spinal cord RA signaling mechanism in vertebrate embryos has been translated into successes in learning how to differentiate embryonic stem (ES) cells in vitro. Both mouse and human ES cells can be stimulated to form motor neurons at high efficiency when RA and SHH are used together as differentiation agents [62-64]. These findings provide hope that knowledge of normal RA function during early organogenesis can be used to generate replacement cells for spinal cord injuries or motor neuron diseases.
Forebrain
RA has not been shown to be required for midbrain development, but a role in the forebrain has been established during late embryonic stages. At E12.5 in the mouse forebrain, Raldh3 expression initiates in the subventricular zone of the lateral ganglionic eminence (LGE) located in ventral forebrain tissue that is fated to become basal ganglia [34, 65]. Raldh3−/− embryos fail to generate RA activity in the ventral forebrain and exhibit a defect in differentiation within the striatum that includes loss of Drd2 expression (encoding dopamine receptor D2) in the nucleus accumbens [34]. A RARE located close to the Drd2 promoter suggests that RA directly induces Drd2 transcription [66]. Consistent with these observations, RARb-/- mice exhibit defects in striatal dopaminergic neurogenesis resulting in motor behavior impairment [67, 68].
RA generated by Raldh3 in the LGE has also been shown to be required for expression of the enzyme that produces the inhibitory neurotransmitter gamma-aminobutyric acid (GABA), i.e. glutamic acid decarboxylase-67 (GAD67). Thus, RA is required for GABAergic differentiation in the striatum which is adjacent to the LGE. Raldh3-/- embryos fail to generate both GABAergic striatal projection neurons and GABAergic interneurons migrating from the striatum to the olfactory bulb and cortex [69]. In addition, LGE-derived neurospheres from wild-type but not Raldh3-/- embryos produce RA needed for GABAergic differentiation in vitro. As no evidence exists for a RARE near the Gad67 gene, it is unclear whether Gad67 is a direct or indirect target of RA signaling in the striatum during GABAergic differentiation. RA treatment protocols have been described for mouse and human ES cells that result in differentiation to GABAergic neurons [69-72], thus providing a potential source of cells for therapeutic purposes.
Raldh genes are not expressed in the embryonic forebrain cortex and RA activity is not observed in this tissue using techniques that easily detect RA activity in the LGE where Raldh3 is expressed [69, 73]. However, Raldh2 is expressed in the meninges by E12.5 [74], a neural crest-derived tissue which provides an outer cover for the brain separating it from the cranial bones. Meningeal RA is likely the source of RA required to control RARs needed for proper cranial bone formation [75]. Rdh10, controlling the metabolism of retinol to retinaldehyde, is also expressed in the meninges by E12.5 [76]. Studies on E14.5 ethylnitrosourea-induced Rdh10 mutants that exhibit a reduction in radial expansion of the forebrain cortex suggested that RA produced in the meninges may be a neurogenic factor inducing the switch from symmetric to asymmetric divisions in the ventricular zone needed for cortical expansion [76]. However, studies on E14.5 Raldh2-/- embryos do not support this conclusion [69]. As Rdh10 mutants exhibit severe neural crest-derived craniofacial defects that distort the cranium [1], whereas Raldh2-/- embryos do not [69], thinning of the cortex in Rdh10 mutants is unlikely to be a specific effect of RA on corticogenesis, but rather a defect in cranial neural crest migration and differentiation which leads to the altered cortical morphology. This conclusion is also supported by studies on E14.5 Rdh10-/- embryos treated early with retinaldehyde (E7.5-E9.5) to rescue the craniofacial defect, but that still lack RA activity later in the meninges at E14.5 and maintain normal cortical morphology [35]. The conclusion that RA is unnecessary for cortical differentiation is also supported by studies showing that mouse ES cells can be differentiated into forebrain cortical neurons without addition of RA [77].
3. Retinoic acid control of axial progenitor cells in the caudal progenitor zone
Tissue-specific induction of 3’-Hox genes, Hnf1b, Ngn2, Drd2, Gad67 and other genes by RA signaling constitutes an instructive mechanism of RA action that is certainly an important function for RA during development. However, RA can also control development in a permissive fashion through its ability to repress gene expression as discussed below.
The development of vertebrate embryos occurs in a head-to-tail fashion through the process of body axis extension in which progenitor cells in the caudal progenitor zone generate new tissue [78]. Bipotential axial (neuromesodermal) progenitor cells present in the caudal progenitor zone differentiate into either neuroectoderm to generate the neural tube or presomitic mesoderm to form somites that are precursors of vertebrae and skeletal muscle [79]. Fgf8 expressed in the caudal progenitor zone is essential for maintaining axial progenitor cell function needed to extend the body axis and for the process of somitogenesis [80]. RA generated in the presomitic mesoderm functions as a diffusible signal that represses Fgf8 expression at the anterior end of the caudal progenitor zone, thus allowing cells exiting the progenitor zone to undergo neural differentiation or somitogenesis during body axis extension [54, 81-85]. As RA secreted by presomitic mesoderm was found to act at the neuroectoderm/epiblast junction in order to control somitogenesis [82], the target of RA signaling during body axis extension may be the axial progenitor cells where Fgf8 is most highly expressed. Fgf8 expression initiates in the caudal progenitor zone during early gastrulation with Raldh2 expression following later in the presomitic mesoderm [32], Thus, RA signaling is introduced into the picture during late gastrulation and begins to restrict caudal FGF8 signaling at that time. RA-FGF8 antagonism allows cells at the border of these two signaling pathways to transition away from FGF8 signaling and begin responding to other signals that stimulate differentiation. RA repression of Fgf8 is thus a fundamental permissive function of RA that occurs simultaneous to its instructive function during differentiation of neuroectoderm in the hindbrain and spinal cord.
4. Retinoic acid control of second heart field progenitor cells
The ability of RA signaling to repress gene expression is also important for heart development. The vertebrate heart initially develops as a ventral tube along the anteroposterior axis of the upper trunk from a population of ventral cardiac progenitor cells in the splanchnic lateral plate mesoderm called the first heart field [86]. As the heart tube lengthens, additional cardiac splanchnic mesodermal progenitor cells located dorsally in the second heart field are added to the anterior and posterior poles of the growing heart [86-90]. The LIM homeodomain transcription factor encoded by Isl1 is expressed in cardiac progenitors of the second heart field, and as these cells are recruited to the heart tube Isl1 is down-regulated, although Isl1 expression remains dorsal to the heart and continues to contribute progenitors to the outflow tract at later stages [91]. Genetic studies in mouse embryos indicate that Isl1-/- embryos are missing the outflow tract, right ventricle, and much of the atria which all require a major contribution from second heart field progenitors [91]. In contrast, loss of the T-box transcription factor Tbx5 results in sinoatrial defects and hypoplastic left ventricle, regions associated with a major contribution from the first heart field [92]. Thus, Isl1 is important for maintaining cardiac progenitors along most of the heart anteroposterior axis with the exception of the left ventricle. Recent studies demonstrating the existence of a second heart field expressing an ancestral Isl1 gene in Ciona have revealed an early chordate origin for the second heart field [93].
Raldh2 is first expressed in presomitic mesoderm and lateral plate mesoderm posterior to the heart just after the heart has begun to develop, and secreted RA travels into posterior cardiac mesoderm [94]. Studies in mouse have demonstrated that loss of RA synthesis in Raldh2 mutants results in an enlarged heart tube lacking looping and chamber formation [95]. Zebrafish raldh2 mutants exhibit an expansion in the number of cardiac progenitor cells [96] and vitamin A deficiency studies in avian embryos have shown that RA limits the size of the cardiac field [94, 97]. There are no clear examples of genes induced by RA that control the number of cardiac progenitors, but it appears that RA may function by limiting cardiac FGF signaling. A potential role for FGF signaling in zebrafish heart development was found in studies showing that expression of a constitutive-active FGF receptor resulted in an expanded heart [98]. Genetic studies in mice indicate that Fgf8 expressed in dorsal splanchnic mesoderm is required to drive cardiac progenitors toward an Isl1+ second heart field fate, thus demonstrating that Fgf8 is an important cardiac signaling factor [99, 100]. In E8.5 Raldh2-/- mouse embryos, cardiac expression of Fgf8 and Isl1 was found to expand posteriorly, possibly accounting for the increase in heart size [101, 102]; similar results were observed in E8.5 Rdh10 mutant hearts that lack physiological RA activity [103]. Also, Raldh2-/- and Rdh10 mutant embryos exhibit ectopic FGF signaling posterior to the heart marked by Sprouty2 expression [101, 103]. Thus, RA may limit the expansion of second heart field progenitors by repression of cardiac Fgf8 expression, similar to how RA repression of Fgf8 in the caudal progenitor zone is proposed to restrict expansion of caudal progenitors.
5. Retinoic acid regulation of male germ cell differentiation
Germ cells are specified in mouse embryos at about E6.5, then at E8.5 they begin to migrate through the posterior region of the embryo and the mesonephros until arriving in the developing gonad at E10.5. When they enter the gonad in the genital ridge, germ cells are bipotential but over the next 2-3 days they commit to either the male or female fate [104]. Bipotential germ cells divide mitotically until about E12.5-E13.5 when germ cells in the ovary terminate mitosis and enter prophase of the first meiotic division, a female-specific trait. In contrast, during this time germ cells of the testis fail to enter meiotic prophase, a male-specific trait, and arrest in the Go/G1 phase of the mitotic cycle until about postnatal day 5 when they enter meiosis [105].
RA has long been known to be required postnatally for spermatogenesis [106, 107], but a role for RA in oogenesis was not originally observed. More recent studies suggested that RA controls sex-specific onset of meiosis in germ cells. In those studies, which employed treatment of gonads with high levels of RA or RA receptor antagonists, RA generated by Raldh2 in the embryonic mesonephros was proposed to function as a secreted factor that induces Stra8 (required for meiotic initiation) in the adjacent embryonic ovary but not the embryonic testis due to testis-specific expression of Cyp26b1 encoding a P450 enzyme that stimulates RA degradation [108, 109]. Premature onset of meiosis in the male germ cells was observed in testis organ cultures treated with a P450 inhibitor [108] or in Cyp26b1-/- mice [109]. However, investigation of Raldh2-/- mouse embryos revealed that Stra8 is still expressed at a normal level in ovary despite a complete absence of RA activity in mesonephros and ovary [110]. In addition, treatment of Raldh2-/- testis organ cultures with a P450 inhibitor still allowed Stra8 induction in the absence of RA activity [110]. These findings demonstrate that the critical role Cyp26b1 plays in preventing meiosis in fetal testis does not involve degradation of RA, suggesting an RA-independent function for Cyp26b1 during prevention of premature meiotic initiation in testis. Meanwhile other mechanisms for female-specific initiation of meiosis are suggested by genetic studies on Dmrt1 [111] and Msx1/2 [112].
In contrast, recent genetic studies on the role of RA in spermatogenesis have provided conclusive evidence for specific RA-regulated functions. Studies on compound Raldh conditional knockout mice have shown that RA generated at postnatal day 5 in Sertoli cells by Raldh1 and Raldh2 functions as a paracrine signal for initiation of spermatogonial germ cell differentiation to generate pre-meiotic spermatocytes [113]. Importantly, those studies also demonstrated that Raldh2 subsequently expressed in pre-meiotic spermatocytes synthesizes RA needed to locally drive expression of Stra8 (which has a nearby RARE) and initiate meiosis [113]. Thus, genetic studies provide strong evidence that RA is required for male but not female germ cell differentiation.
6. Conclusions and Future Directions
Studies focused on how RA controls nuclear receptor function during development have provided deep insight into how RA controls differentiation of progenitor cells. Instrumental in this investigation has been the discovery of key enzymes encoded by Rdh10, Raldh1, Raldh2, and Raldh3 that generate RA in specific differentiated cell types designed to control differentiation of nearby progenitor cells in a paracrine fashion, or perhaps in some cases change the differentiation status of the RA-generating cell in an autocrine fashion. The defects observed in mouse embryos carrying Raldh mutations have identified differentiation events controlled by RA signaling during embryogenesis, a process which is very similar in mice and humans. From such studies, we know that RA is required to induce specific genes for differentiation of neural progenitor cells in the posterior hindbrain, spinal cord, and forebrain basal ganglia. These studies have also demonstrated that RA antagonizes FGF signaling emanating from axial progenitor cells in the embryonic caudal progenitor zone, thus controlling differentiation as the body axis extends in a head-to-tail fashion. RA signaling also antagonizes FGF signaling in progenitor cells of the second heart field, suggesting that RA antagonism of FGF signaling may be a common mode of RA action. Regulation of male germ cell differentiation in the testis provides a glimpse into how RA continues to control progenitor cell differentiation postnatally.
Several open questions remain that can direct future studies. In addition to RDH10, what other enzyme(s) are required for metabolism of retinol to retinaldehyde in the first step of RA synthesis? Are RALDH1, RALDH2, and RALDH3 sufficient for the second step of RA synthesis or do additional enzymes play a role in the embryo or postnatally? During motor neuron differentiation, is the Ngn2 RARE required for RA regulation, and are other genes direct targets of RA action? What is the mechanism of Gad67 induction in the forebrain basal ganglia by RA? During body axis extension and second heart field development, does RA repress Fgf8 expression directly or does repression occur indirectly through RA induction of a repressor? If RA does not function as a diffusible signal controlling female meiosis, what is the nature of this signal?
Discovery of how RA normally controls progenitor cells during organogenesis will provide information essential for the field of regenerative medicine. This knowledge will help establish regenerative treatments for damaged or aged human tissues based upon therapies designed to stimulate endogenous progenitor cells to produce differentiated cells in vivo or therapies that rely on introduction of differentiated cells produced in vitro from embryonic stem cells or induced pluripotent stem cells.
Highlights.
Retinoic acid stimulates neuronal differentiation of progenitor cells in the posterior CNS and forebrain basal ganglia.
Differentiation of progenitor cells in the caudal progenitor zone is controlled by retinoic acid antagonism of FGF signaling.
FGF signaling in progenitor cells of the second heart field is limited by retinoic acid signaling.
Retinoic acid is required for spermatogonial differentiation plus meiotic initiation in male but not female germ cells.
Acknowledgements
Thanks go to past and present members of my laboratory for stimulating discussions on the role of retinoic acid signaling during development. Funding was provided by National Institutes of Health grant GM062848.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errorsmaybe discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Sandell LL, Sanderson BW, Moiseyev G, Johnson T, Mushegian A, Young K, Rey JP, Ma JX, Staehling-Hampton K, Trainor PA. RDH10 is essential for synthesis of embryonic retinoic acid and is required for limb, craniofacial, and organ development. Genes Dev. 2007;21:1113–1124. doi: 10.1101/gad.1533407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Niederreither K, Subbarayan V, Dollé P, Chambon P. Embryonic retinoic acid synthesis is essential for early mouse post-implantation development. Nature Genet. 1999;21:444–448. doi: 10.1038/7788. [DOI] [PubMed] [Google Scholar]
- 3.Mic FA, Haselbeck RJ, Cuenca AE, Duester G. Novel retinoic acid generating activities in the neural tube and heart identified by conditional rescue of Raldh2 null mutant mice. Development. 2002;129:2271–2282. doi: 10.1242/dev.129.9.2271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kumar S, Sandell LL, Trainor PA, Koentgen F, Duester G. Alcohol and aldehyde dehydrogenases: Retinoid metabolic effects in mouse knockout models. Biochimica et Biophysica Acta. 2012;1821:198–205. doi: 10.1016/j.bbalip.2011.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Belyaeva OV, Lee SA, Adams MK, Chang C, Kedishvili NY. Short chain dehydrogenase/reductase rdhe2 is a novel retinol dehydrogenase essential for frog embryonic development. J Biol Chem. 2012;287:9061–9071. doi: 10.1074/jbc.M111.336727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chambon P. A decade of molecular biology of retinoic acid receptors. FASEB J. 1996;10:940–954. [PubMed] [Google Scholar]
- 7.Mangelsdorf DJ, Evans RM. The RXR heterodimers and orphan receptors. Cell. 1995;83:841–850. doi: 10.1016/0092-8674(95)90200-7. [DOI] [PubMed] [Google Scholar]
- 8.Kumar S, Duester G. SnapShot: Retinoic Acid Signaling. Cell. 2011;147:1422. doi: 10.1016/j.cell.2011.11.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Thisse B, Thisse C. Functions and regulations of fibroblast growth factor signaling during embryonic development. Dev Biol. 2005;287:390–402. doi: 10.1016/j.ydbio.2005.09.011. [DOI] [PubMed] [Google Scholar]
- 10.Itoh N, Ornitz DM. Functional evolutionary history of the mouse Fgf gene family. Dev Dyn. 2008;237:18–27. doi: 10.1002/dvdy.21388. [DOI] [PubMed] [Google Scholar]
- 11.Reya T, Clevers H. Wnt signaling in stem cells and cancer. Nature. 2005;434:843–850. doi: 10.1038/nature03319. [DOI] [PubMed] [Google Scholar]
- 12.Nelson WJ, Nusse R. Convergence of Wnt, β-catenin, and cadherin pathways. Science. 2004;303:1483–1487. doi: 10.1126/science.1094291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Derynck R, Zhang YE. Smad-dependent and Smad-independent pathways in TGF-β family signaling. Nature. 2003;425:577–584. doi: 10.1038/nature02006. [DOI] [PubMed] [Google Scholar]
- 14.Shen MM. Nodal signaling: developmental roles and regulation. Development. 2007;134:1023–1034. doi: 10.1242/dev.000166. [DOI] [PubMed] [Google Scholar]
- 15.Lum L, Beachy PA. The hedgehog response network: sensors, switches, and routers. Science. 2004;304:1755–1759. doi: 10.1126/science.1098020. [DOI] [PubMed] [Google Scholar]
- 16.Ingham PW, Placzek M. Orchestrating ontogenesis: variations on a theme by sonic hedgehog. Nature Rev Genet. 2006;7:841–850. doi: 10.1038/nrg1969. [DOI] [PubMed] [Google Scholar]
- 17.Marlétaz F, Holland LZ, Laudet V, Schubert M. Retinoic acid signaling and the evolution of chordates. Int J Biol Sci. 2006;2:38–47. doi: 10.7150/ijbs.2.38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sobreira TJ, Marletaz F, Simoes-Costa M, Schechtman D, Pereira AC, Brunet F, Sweeney S, Pani A, Aronowicz J, Lowe CJ, Davidson B, Laudet V, Bronner M, de Oliveira PS, Schubert M, Xavier-Neto J. Structural shifts of aldehyde dehydrogenase enzymes were instrumental for the early evolution of retinoid-dependent axial patterning in metazoans. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:226–231. doi: 10.1073/pnas.1011223108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Schug TT, Berry DC, Shaw NS, Travis SN, Noy N. Opposing effects of retinoic Acid on cell growth result from alternate activation of two different nuclear receptors. Cell. 2007;129:723–733. doi: 10.1016/j.cell.2007.02.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Yu S, Levi L, Siegel R, Noy N. Retinoic Acid Induces Neurogenesis by Activating Both Retinoic Acid Receptors (RARs) and Peroxisome Proliferator-activated Receptor beta/delta (PPARbeta/delta) Journal of Biological Chemistry. 2012;287:42195–42205. doi: 10.1074/jbc.M112.410381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Maden M, Gale E, Kostetskii I, Zile MH. Vitamin A-deficient quail embryos have half a hindbrain and other neural defects. Curr Biol. 1996;6:417–426. doi: 10.1016/s0960-9822(02)00509-2. [DOI] [PubMed] [Google Scholar]
- 22.Clagett-Dame M, DeLuca HF. The role of vitamin A in mammalian reproduction and embryonic development. Annu Rev Nutr. 2002;22:347–381. doi: 10.1146/annurev.nutr.22.010402.102745E. [DOI] [PubMed] [Google Scholar]
- 23.Kastner P, Mark M, Chambon P. Nonsteroid nuclear receptors: What are genetic studies telling us about their role in real life? Cell. 1995;83:859–869. doi: 10.1016/0092-8674(95)90202-3. [DOI] [PubMed] [Google Scholar]
- 24.Duester G. Retinoic acid synthesis and signaling during early organogenesis. Cell. 2008;134:921–931. doi: 10.1016/j.cell.2008.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rhinn M, Dolle P. Retinoic acid signalling during development. Development. 2012;139:843–858. doi: 10.1242/dev.065938. [DOI] [PubMed] [Google Scholar]
- 26.Maden M. Retinoic acid in the development, regeneration and maintenance of the nervous system. Nat Rev Neurosci. 2007;8:755–765. doi: 10.1038/nrn2212. [DOI] [PubMed] [Google Scholar]
- 27.Cai C, Grabel L. Directing the differentiation of embryonic stem cells to neural stem cells. Dev Dyn. 2007;236:3255–3266. doi: 10.1002/dvdy.21306. [DOI] [PubMed] [Google Scholar]
- 28.Gudas LJ, Wagner JA. Retinoids regulate stem cell differentiation. J Cell Physiol. 2011;226:322–330. doi: 10.1002/jcp.22417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Horton C, Maden M. Endogenous distribution of retinoids during normal development and teratogenesis in the mouse embryo. Dev Dyn. 1995;202:312–323. doi: 10.1002/aja.1002020310. [DOI] [PubMed] [Google Scholar]
- 30.Mic FA, Molotkov A, Benbrook DM, Duester G. Retinoid activation of retinoic acid receptor but not retinoid X receptor is sufficient to rescue lethal defect in retinoic acid synthesis. Proc Natl Acad Sci USA. 2003;100:7135–7140. doi: 10.1073/pnas.1231422100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Niederreither K, Vermot J, Schuhbaur B, Chambon P, Dollé P. Retinoic acid synthesis and hindbrain patterning in the mouse embryo. Development. 2000;127:75–85. doi: 10.1242/dev.127.1.75. [DOI] [PubMed] [Google Scholar]
- 32.Sirbu IO, Gresh L, Barra J, Duester G. Shifting boundaries of retinoic acid activity control hindbrain segmental gene expression. Development. 2005;132:2611–2622. doi: 10.1242/dev.01845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Molotkova N, Molotkov A, Sirbu IO, Duester G. Requirement of mesodermal retinoic acid generated by Raldh2 for posterior neural transformation. Mech Dev. 2005;122:145–155. doi: 10.1016/j.mod.2004.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Molotkova N, Molotkov A, Duester G. Role of retinoic acid during forebrain development begins late when Raldh3 generates retinoic acid in the ventral subventricular zone. Dev Biol. 2007;303:601–610. doi: 10.1016/j.ydbio.2006.11.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chatzi C, Cunningham TJ, Duester G. Investigation of retinoic acid function during embryonic brain development using retinaldehyde-rescued Rdh10 knockout mice. Dev Dyn. 2013 doi: 10.1002/dvdy.23999. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Matt N, Dupé V, Garnier J- M, Dennefeld C, Chambon P, Mark M, Ghyselinck NB. Retinoic acid-dependent eye morphogenesis is orchestrated by neural crest cells. Development. 2005;132:4789–4800. doi: 10.1242/dev.02031. [DOI] [PubMed] [Google Scholar]
- 37.Molotkov A, Molotkova N, Duester G. Retinoic acid guides eye morphogenetic movements via paracrine signaling but is unnecessary for retinal dorsoventral patterning. Development. 2006;133:1901–1910. doi: 10.1242/dev.02328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Simeone A, Acampora D, Arcioni L, Andrews PW, Boncinelli E, Mavilio F. Sequential activation of HOX2 homeobox genes by retinoic acid in human embryonal carcinoma cells. Nature. 1990;346:763–766. doi: 10.1038/346763a0. [DOI] [PubMed] [Google Scholar]
- 39.Krumlauf R. Hox genes and pattern formation in the branchial region of the vertebrate head. Trends Genet. 1993;9:106–112. doi: 10.1016/0168-9525(93)90203-t. [DOI] [PubMed] [Google Scholar]
- 40.Goddard JM, Rossel M, Manley NR, Capecchi MR. Mice with targeted disruption of Hoxb-1 fail to form the motor nucleus of the VIIth nerve. Development. 1996;122:3217–3228. doi: 10.1242/dev.122.10.3217. [DOI] [PubMed] [Google Scholar]
- 41.Studer M, Lumsden A, Ariza-McNaughton L, Bradley A, Krumlauf R. Altered segmental identity and abnormal migration of motor neurons in mice lacking Hoxb-1. Nature. 1996;384:630–634. doi: 10.1038/384630a0. [DOI] [PubMed] [Google Scholar]
- 42.Dupé V, Lumsden A. Hindbrain patterning involves graded responses to retinoic acid signalling. Development. 2001;128:2199–2208. doi: 10.1242/dev.128.12.2199. [DOI] [PubMed] [Google Scholar]
- 43.Begemann G, Schilling TF, Rauch GJ, Geisler R, Ingham PW. The zebrafish neckless mutation reveals a requirement for raldh2 in mesodermal signals that pattern the hindbrain. Development. 2001;128:3081–3094. doi: 10.1242/dev.128.16.3081. [DOI] [PubMed] [Google Scholar]
- 44.Maves L, Kimmel CB. Dynamic and sequential patterning of the zebrafish posterior hindbrain by retinoic acid. Dev Biol. 2005;285:593–605. doi: 10.1016/j.ydbio.2005.07.015. [DOI] [PubMed] [Google Scholar]
- 45.Wilkinson DG, Bhatt S, Cook M, Boncinelli E, Krumlauf R. Segmental expression of Hox-2 homeobox-containing genes in the developing mouse hindbrain. Nature. 1989;341:405–409. doi: 10.1038/341405a0. [DOI] [PubMed] [Google Scholar]
- 46.Marshall H, Studer M, Pöpperl H, Aparicio S, Kuroiwa A, Brenner S, Krumlauf R. A conserved retinoic acid response element required for early expression of the homeobox gene Hoxb-1. Nature. 1994;370:567–571. doi: 10.1038/370567a0. [DOI] [PubMed] [Google Scholar]
- 47.Studer M, Pöpperl H, Marshall H, Kuroiwa A, Krumlauf R. Role of a conserved retinoic acid response element in rhombomere restriction of Hoxb-1. Science. 1994;265:1728–1732. doi: 10.1126/science.7916164. [DOI] [PubMed] [Google Scholar]
- 48.Kashyap V, Gudas LJ, Brenet F, Funk P, Viale A, Scandura JM. Epigenomic reorganization of the clustered hox genes in embryonic stem cells induced by retinoic Acid. Journal of Biological Chemistry. 2011;286:3250–3260. doi: 10.1074/jbc.M110.157545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wiellette EL, Sive H. vhnf1 and Fgf signals synergize to specify rhombomere identity in the zebrafish hindbrain. Development. 2003;130:3821–3829. doi: 10.1242/dev.00572. [DOI] [PubMed] [Google Scholar]
- 50.Hernandez RE, Rikhof HA, Bachmann R, Moens CB. vhnf1 integrates global RA patterning and local FGF signals to direct posterior hindbrain development in zebrafish. Development. 2004;131:4511–4520. doi: 10.1242/dev.01297. [DOI] [PubMed] [Google Scholar]
- 51.Pouilhe M, Gilardi-Hebenstreit P, Desmarquet-Trin Dinh C, Charnay P. Direct regulation of vHnf1 by retinoic acid signaling and MAF-related factors in the neural tube. Dev Biol. 2007;309:344–357. doi: 10.1016/j.ydbio.2007.07.003. [DOI] [PubMed] [Google Scholar]
- 52.Roelink H, Porter JA, Chiang C, Tanabe Y, Chang DT, Beachy PA, Jessell TM. Floor plate and motor neuron induction by different concentrations of the amino-terminal cleavage product of Sonic hedgehog autoproteolysis. Cell. 1995;81:445–455. doi: 10.1016/0092-8674(95)90397-6. [DOI] [PubMed] [Google Scholar]
- 53.Sockanathan S, Jessell TM. Motor neuron-derived retinoid signaling specifies the subtype identity of spinal motor neurons. Cell. 1998;94:503–514. doi: 10.1016/s0092-8674(00)81591-3. [DOI] [PubMed] [Google Scholar]
- 54.Del Corral RD, Olivera-Martinez I, Goriely A, Gale E, Maden M, Storey K. Opposing FGF and retinoid pathways control ventral neural pattern, neuronal differentiation, and segmentation during body axis extension. Neuron. 2003;40:65–79. doi: 10.1016/s0896-6273(03)00565-8. [DOI] [PubMed] [Google Scholar]
- 55.Novitch BG, Wichterle H, Jessell TM, Sockanathan S. A requirement for retinoic acid-mediated transcriptional activation in ventral neural patterning and motor neuron specification. Neuron. 2003;40:81–95. doi: 10.1016/j.neuron.2003.08.006. [DOI] [PubMed] [Google Scholar]
- 56.Wilson L, Gale E, Chambers D, Maden M. Retinoic acid and the control of dorsoventral patterning in the avian spinal cord. Dev Biol. 2004;269:433–446. doi: 10.1016/j.ydbio.2004.01.034. [DOI] [PubMed] [Google Scholar]
- 57.Takebayashi H, Nabeshima Y, Yoshida S, Chisaka O, Ikenaka K. The basic helix-loop-helix factor Olig2 is essential for the development of motoneuron and oligodendrocyte lineages. Curr Biol. 2002;12:1157–1163. doi: 10.1016/s0960-9822(02)00926-0. [DOI] [PubMed] [Google Scholar]
- 58.Lu QR, Sun T, Zhu Z, Ma N, Garcia M, Stiles CD, Rowitch DH. Common developmental requirement for Olig function indicates a motor neuron/oligodendrocyte connection. Cell. 2002;109:75–86. doi: 10.1016/s0092-8674(02)00678-5. [DOI] [PubMed] [Google Scholar]
- 59.Marquardt T, Pfaff SL. Cracking the transcriptional code for cell specification in the neural tube. Cell. 2001;106:651–654. doi: 10.1016/s0092-8674(01)00499-8. [DOI] [PubMed] [Google Scholar]
- 60.Scardigli R, Schuurmans C, Gradwohl G, Guillemot F. Crossregulation between Neurogenin2 and pathways specifying neuronal identity in the spinal cord. Neuron. 2001;31:203–217. doi: 10.1016/s0896-6273(01)00358-0. [DOI] [PubMed] [Google Scholar]
- 61.Ribes V, Stutzmann F, Bianchetti L, Guillemot F, Dolle P, Le Roux I. Combinatorial signalling controls Neurogenin2 expression at the onset of spinal neurogenesis. Dev Biol. 2008;321:470–481. doi: 10.1016/j.ydbio.2008.06.003. [DOI] [PubMed] [Google Scholar]
- 62.Wichterle H, Lieberam I, Porter JA, Jessell TM. Directed differentiation of embryonic stem cells into motor neurons. Cell. 2002;110:385–397. doi: 10.1016/s0092-8674(02)00835-8. [DOI] [PubMed] [Google Scholar]
- 63.Li XJ, Du ZW, Zarnowska ED, Pankratz M, Hansen LO, Pearce RA, Zhang SC. Specification of motoneurons from human embryonic stem cells. Nat Biotechnol. 2005;23:215–221. doi: 10.1038/nbt1063. [DOI] [PubMed] [Google Scholar]
- 64.Hu BY, Zhang SC. Differentiation of spinal motor neurons from pluripotent human stem cells. Nature Protocols. 2009;4:1295–1304. doi: 10.1038/nprot.2009.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Li H, Wagner E, McCaffery P, Smith D, Andreadis A, Dräger UC. A retinoic acid synthesizing enzyme in ventral retina and telencephalon of the embryonic mouse. Mech Dev. 2000;95:283–289. doi: 10.1016/s0925-4773(00)00352-x. [DOI] [PubMed] [Google Scholar]
- 66.Samad TA, Krezel W, Chambon P, Borrelli E. Regulation of dopaminergic pathways by retinoids: Activation of the D2 receptor promoter by members of the retinoic acid receptor retinoid X receptor family. Proc Natl Acad Sci USA. 1997;94:14349–14354. doi: 10.1073/pnas.94.26.14349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Krezel W, Ghyselinck N, Samad TA, Dupé V, Kastner P, Borrelli E, Chambon P. Impaired locomotion and dopamine signaling in retinoid receptor mutant mice. Science. 1998;279:863–867. doi: 10.1126/science.279.5352.863. [DOI] [PubMed] [Google Scholar]
- 68.Liao WL, Tsai HC, Wang HF, Chang J, Lu KM, Wu HL, Lee YC, Tsai TF, Takahashi H, Wagner M, Ghyselinck NB, Chambon P, Liu FC. Modular patterning of structure and function of the striatum by retinoid receptor signaling. Proc Natl Acad Sci USA. 2008;105:6765–6770. doi: 10.1073/pnas.0802109105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Chatzi C, Brade T, Duester G. Retinoic acid functions as a key GABAergic differentiation signal in the basal ganglia. PLoS Biol. 2011;9:e1000609. doi: 10.1371/journal.pbio.1000609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Chatzi C, Scott RH, Pu J, Lang B, Nakamoto C, McCaig CD, Shen S. Derivation of homogeneous GABAergic neurons from mouse embryonic stem cells. Experimental Neurology. 2009;217:407–416. doi: 10.1016/j.expneurol.2009.03.032. [DOI] [PubMed] [Google Scholar]
- 71.Addae C, Yi X, Gernapudi R, Cheng H, Musto A, Martinez-Ceballos E. All-trans-retinoid acid induces the differentiation of encapsulated mouse embryonic stem cells into GABAergic neurons. Differentiation. 2012;83:233–241. doi: 10.1016/j.diff.2012.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Shin E, Palmer MJ, Li M, Fricker RA. GABAergic Neurons from Mouse Embryonic Stem Cells Possess Functional Properties of Striatal Neurons In Vitro, and Develop into Striatal Neurons In Vivo in a Mouse Model of Huntington's Disease. Stem Cell Reviews. 2012;8:513–531. doi: 10.1007/s12015-011-9290-2. [DOI] [PubMed] [Google Scholar]
- 73.Luo TL, Wagner E, Grün F, Dräger UC. Retinoic acid signaling in the brain marks formation of optic projections, maturation of the dorsal telencephalon, and function of limbic sites. J Comp Neurol. 2004;470:297–316. doi: 10.1002/cne.20013. [DOI] [PubMed] [Google Scholar]
- 74.Smith D, Wagner E, Koul O, McCaffery P, Dräger UC. Retinoic acid synthesis for the developing telencephalon. Cereb Cortex. 2001;11:894–905. doi: 10.1093/cercor/11.10.894. [DOI] [PubMed] [Google Scholar]
- 75.Dupe V, Pellerin I. Retinoic acid receptors exhibit cell-autonomous functions in cranial neural crest cells. Dev Dyn. 2009;238:2701–2711. doi: 10.1002/dvdy.22087. [DOI] [PubMed] [Google Scholar]
- 76.Siegenthaler JA, Ashique AM, Zarbalis K, Patterson KP, Hecht JH, Kane MA, Folias AE, Choe Y, May SR, Kume T, Napoli JL, Peterson AS, Pleasure SJ. Retinoic acid from the meninges regulates cortical neuron generation. Cell. 2009;139:597–609. doi: 10.1016/j.cell.2009.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Gaspard N, Bouschet T, Hourez R, Dimidschstein J, Naeije G, van den Ameele J, Espuny-Camacho I, Herpoel A, Passante L, Schiffmann SN, Gaillard A, Vanderhaeghen P. An intrinsic mechanism of corticogenesis from embryonic stem cells. Nature. 2008;455:351–357. doi: 10.1038/nature07287. [DOI] [PubMed] [Google Scholar]
- 78.Wilson V, Olivera-Martinez I, Storey KG. Stem cells, signals and vertebrate body axis extension. Development. 2009;136:1591–1604. doi: 10.1242/dev.021246. [DOI] [PubMed] [Google Scholar]
- 79.Tzouanacou E, Wegener A, Wymeersch FJ, Wilson V, Nicolas JF. Redefining the progression of lineage segregations during mammalian embryogenesis by clonal analysis. Dev Cell. 2009;17:365–376. doi: 10.1016/j.devcel.2009.08.002. [DOI] [PubMed] [Google Scholar]
- 80.Naiche LA, Holder N, Lewandoski M. FGF4 and FGF8 comprise the wavefront activity that controls somitogenesis. Proc Natl Acad Sci USA. 2011;108:4018–4023. doi: 10.1073/pnas.1007417108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Vermot J, Llamas JG, Fraulob V, Niederreither K, Chambon P, Dollé P. Retinoic acid controls the bilateral symmetry of somite formation in the mouse embryo. Science. 2005;308:563–566. doi: 10.1126/science.1108363. [DOI] [PubMed] [Google Scholar]
- 82.Sirbu IO, Duester G. Retinoic acid signaling in node ectoderm and posterior neural plate directs left-right patterning of somitic mesoderm. Nature Cell Biol. 2006;8:271–277. doi: 10.1038/ncb1374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Ribes V, Le Roux I, Rhinn M, Schuhbaur B, Dolle P. Early mouse caudal development relies on crosstalk between retinoic acid, Shh and Fgf signalling pathways. Development. 2009;136:665–676. doi: 10.1242/dev.016204. [DOI] [PubMed] [Google Scholar]
- 84.Zhao X, Sirbu IO, Mic FA, Molotkova N, Molotkov A, Kumar S, Duester G. Retinoic acid promotes limb induction through effects on body axis extension but is unnecessary for limb patterning. Curr Biol. 2009;19:1050–1057. doi: 10.1016/j.cub.2009.04.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Patel NS, Rhinn M, Semprich CI, Halley PA, Dolle P, Bickmore WA, Storey KG. FGF Signalling Regulates Chromatin Organisation during Neural Differentiation via Mechanisms that Can Be Uncoupled from Transcription. PLoS Genetics. 2013;9:e1003614. doi: 10.1371/journal.pgen.1003614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Buckingham M, Meilhac S, Zaffran S. Building the mammalian heart from two sources of myocardial cells. Nature Rev Genet. 2005;6:826–835. doi: 10.1038/nrg1710. [DOI] [PubMed] [Google Scholar]
- 87.Mjaatvedt CH, Nakaoka T, Moreno-Rodriguez R, Norris RA, Kern MJ, Eisenberg CA, Turner D, Markwald RR. The outflow tract of the heart is recruited from a novel heart-forming field. Dev Biol. 2001;238:97–109. doi: 10.1006/dbio.2001.0409. [DOI] [PubMed] [Google Scholar]
- 88.Waldo KL, Kumiski DH, Wallis KT, Stadt HA, Hutson MR, Platt DH, Kirby ML. Conotruncal myocardium arises from a secondary heart field. Development. 2001:128, 3179–3188. doi: 10.1242/dev.128.16.3179. [DOI] [PubMed] [Google Scholar]
- 89.Brade T, Gessert S, Kuhl M, Pandur P. The amphibian second heart field: Xenopus islet-1 is required for cardiovascular development. Dev Biol. 2007;311:297–310. doi: 10.1016/j.ydbio.2007.08.004. [DOI] [PubMed] [Google Scholar]
- 90.Abu-Issa R, Kirby ML. Heart field: from mesoderm to heart tube. Annu Rev Cell Dev Biol. 2007;23:45–68. doi: 10.1146/annurev.cellbio.23.090506.123331. [DOI] [PubMed] [Google Scholar]
- 91.Cai CL, Liang X, Shi Y, Chu PH, Pfaff SL, Chen J, Evans S. Isl1 identifies a cardiac progenitor population that proliferates prior to differentiation and contributes a majority of cells to the heart. Dev Cell. 2003;5:877–889. doi: 10.1016/s1534-5807(03)00363-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Bruneau BG, Nemer G, Schmitt JP, Charron F, Robitaille L, Caron S, Conner DA, Gessler M, Nemer M, Seidman CE, Seidman JG. A murine model of Holt-Oram syndrome defines roles of the T-box transcription factor Tbx5 in cardiogenesis and disease. Cell. 2001;106:709–721. doi: 10.1016/s0092-8674(01)00493-7. [DOI] [PubMed] [Google Scholar]
- 93.Stolfi A, Gainous TB, Young JJ, Mori A, Levine M, Christiaen L. Early chordate origins of the vertebrate second heart field. Science. 2010;329:565–568. doi: 10.1126/science.1190181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Hochgreb T, Linhares VL, Menezes DC, Sampaio AC, Yan CYI, Cardoso WV, Rosenthal N, Xavier-Neto J. A caudorostral wave of RALDH2 conveys anteroposterior information to the cardiac field. Development. 2003;130:5363–5374. doi: 10.1242/dev.00750. [DOI] [PubMed] [Google Scholar]
- 95.Niederreither K, Vermot J, Messaddeq N, Schuhbaur B, Chambon P, Dollé P. Embryonic retinoic acid synthesis is essential for heart morphogenesis in the mouse. Development. 2001;128:1019–1031. doi: 10.1242/dev.128.7.1019. [DOI] [PubMed] [Google Scholar]
- 96.Keegan BR, Feldman JL, Begemann G, Ingham PW, Yelon D. Retinoic acid signaling restricts the cardiac progenitor pool. Science. 2005;307:247–249. doi: 10.1126/science.1101573. [DOI] [PubMed] [Google Scholar]
- 97.Dersch H, Zile MH. Induction of normal cardiovascular development in the vitamin A-deprived quail embryo by natural retinoids. Dev Biol. 1993;160:424–433. doi: 10.1006/dbio.1993.1318. [DOI] [PubMed] [Google Scholar]
- 98.Marques SR, Lee Y, Poss KD, Yelon D. Reiterative roles for FGF signaling in the establishment of size and proportion of the zebrafish heart. Dev Biol. 2008;321:397–406. doi: 10.1016/j.ydbio.2008.06.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Ilagan R, Abu-Issa R, Brown D, Yang YP, Jiao K, Schwartz RJ, Klingensmith J, Meyers EN. Fgf8 is required for anterior heart field development. Development. 2006;133:2435–2445. doi: 10.1242/dev.02408. [DOI] [PubMed] [Google Scholar]
- 100.Park EJ, Ogden LA, Talbot A, Evans S, Cai CL, Black BL, Frank DU, Moon AM. Required, tissue-specific roles for Fgf8 in outflow tract formation and remodeling. Development. 2006;133:2419–2433. doi: 10.1242/dev.02367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Sirbu IO, Zhao X, Duester G. Retinoic acid controls heart anteroposterior patterning by down-regulating Isl1 through the Fgf8 pathway. Dev Dyn. 2008;237:1627–1635. doi: 10.1002/dvdy.21570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Ryckebusch L, Wang Z, Bertrand N, Lin S-C, Chi X, Schwartz R, Zaffran S, Niederreither K. Retinoic acid deficiency alters second heart field formation. Proc Natl Acad Sci USA. 2008;105:2913–2918. doi: 10.1073/pnas.0712344105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Cunningham TJ, Zhao X, Sandell LL, Evans SM, Trainor PA, Duester G. Antagonism between retinoic acid and fibroblast growth factor signaling during limb development. Cell Reports. 2013;3:1503–1511. doi: 10.1016/j.celrep.2013.03.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.McLaren A. Primordial germ cells in the mouse. Dev Biol. 2003;262:1–15. doi: 10.1016/s0012-1606(03)00214-8. [DOI] [PubMed] [Google Scholar]
- 105.McLaren A, Southee D. Entry of mouse embryonic germ cells into meiosis. Dev Biol. 1997;187:107–113. doi: 10.1006/dbio.1997.8584. [DOI] [PubMed] [Google Scholar]
- 106.Van Pelt AMM, De Rooij DG. Retinoic acid is able to reinitiate spermatogenesis in vitamin A-deficient rats and high replicate doses support the full development of spermatogenic cells. Endocrinology. 1991;128:697–704. doi: 10.1210/endo-128-2-697. [DOI] [PubMed] [Google Scholar]
- 107.Lufkin T, Lohnes D, Mark M, Dierich A, Gorry P, Gaub M- P, LeMeur M, Chambon P. High postnatal lethality and testis degeneration in retinoic acid receptor a mutant mice. Proc Natl Acad Sci USA. 1993;90:7225–7229. doi: 10.1073/pnas.90.15.7225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Koubova J, Menke DB, Zhou Q, Capel B, Griswold MD, Page DC. Retinoic acid regulates sex-specific timing of meiotic initiation in mice. Proc Natl Acad Sci USA. 2006;103:2474–2479. doi: 10.1073/pnas.0510813103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Bowles J, Knight D, Smith C, Wilhelm D, Richman J, Mamiya S, Yashiro K, Chawengsaksophak K, Wilson MJ, Rossant J, Hamada H, Koopman P. Retinoid signaling determines germ cell fate in mice. Science. 2006;312:596–600. doi: 10.1126/science.1125691. [DOI] [PubMed] [Google Scholar]
- 110.Kumar S, Chatzi C, Brade T, Cunningham TJ, Zhao X, Duester G. Sex-specific timing of meiotic initiation is regulated by Cyp26b1 independent of retinoic acid signaling. Nature Commun. 2011;2:151. doi: 10.1038/ncomms1136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Krentz AD, Murphy MW, Sarver AL, Griswold MD, Bardwell VJ, Zarkower D. DMRT1 promotes oogenesis by transcriptional activation of Stra8 in the mammalian fetal ovary. Dev Biol. 2011;356:63–70. doi: 10.1016/j.ydbio.2011.05.658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Le Bouffant R, Souquet B, Duval N, Duquenne C, Herve R, Frydman N, Robert B, Habert R, Livera G. Msx1 and Msx2 promote meiosis initiation. Development. 2011;138:5393–5402. doi: 10.1242/dev.068452. [DOI] [PubMed] [Google Scholar]
- 113.Raverdeau M, Gely-Pernot A, Feret B, Dennefeld C, Benoit G, Davidson I, Chambon P, Mark M, Ghyselinck NB. Retinoic acid induces Sertoli cell paracrine signals for spermatogonia differentiation but cell autonomously drives spermatocyte meiosis. Proc Natl Acad Sci USA. 2012;109:16582–16587. doi: 10.1073/pnas.1214936109. [DOI] [PMC free article] [PubMed] [Google Scholar]