

Abstract
The presynaptic serotonin (5-HT) transporter (SERT) is a key regulator of 5-HT signaling and is a major target for antidepressant medications and psychostimulants. In recent years, studies of natural and engineered genetic variation in SERT have provided new opportunities to understand structural dimensions of drug interactions and regulation of the transporter, to explore 5-HT contributions to antidepressant action, and to assess the impact of SERT-mediated 5-HT contributions to neuropsychiatric disorders. Here we review three examples from our recent studies where genetic changes in SERT, identified or engineered, have led to new models, findings, and theories that cast light on new dimensions of 5-HT action in the CNS and periphery. First, we review our work to identify specific residues through which SERT recognizes antagonists, and the conversion of this knowledge to the creation of mice lacking high-affinity antidepressant and cocaine sensitivity. Second, we discuss our studies of functional coding variation in SERT that exists in commonly used strains of inbred mice, and how this variation is beginning to reveal novel 5-HT-associated phenotypes. Third, we review our identification and functional characterization of multiple, hyperactive SERT coding variants in subjects with autism. Each of these activities has driven the development of new model systems that can be further exploited to understand the contribution of 5-HT signaling to risk for neuropsychiatric disorders and their treatment.
Keywords: serotonin, antidepressant, transporter, cocaine, autism, p38 mitogen-activated protein kinase
Presynaptic, 5-hydroxytryptamine (5-HT, serotonin) transporters (SERTs, SLC6A4) mediate the clearance and re-uptake of 5-HT during synaptic transmission (Ramamoorthy et al., 1993a). SERTs are highly expressed by midbrain raphe nuclei serotonergic neurons, beginning at the earliest stages of 5-HT neuron differentiation (Schroeter and Blakely, 1996; Hansson et al., 1998; Hendricks et al., 2003; Wylie et al., 2010). Although restricted in expression to 5-HT neurons in the adult, SERTs are expressed by nonserotonergic neurons during development (Hansson et al., 1998, 1999; Gaspar et al., 2003). Along with expression by the placenta (Ramamoorthy et al., 1993b), these latter sites are thought to coordinate the availability of 5-HT to act in the modulation of CNS axon guidance and synapse formation (Bonnin et al., 2007). Besides the placenta, SERTs are expressed in platelets (Mercado and Kilic, 2010), lymphocytes (Barkan et al., 2004), pancreatic beta cells (Smith et al., 1999), pulmonary epithelium (Bhat and Block, 1990), bone (Bliziotes et al., 2001), and the gut mucosa (Chen et al., 2001). Whereas the role of SERT in restricting access of targets to 5-HT and in acquiring 5-HT for release has been well-studied, the acquisition of 5-HT for transglutaminase II-catalyzed covalent attachment to small GTP-binding proteins that can modulate the fusion of secretory granules (Walther et al., 2003), and 5-HT generated oxidative stress and stress-activated MAPK/Rho kinases have emerged recently as yet other mechanisms by which SERT influences physiology (Guilluy et al., 2009; Liu et al., 2011). Together, these (and likely other) actions of SERT reveal a broader range of physiological actions of the transporter in the brain and periphery than previously understood and that will require new approaches and models to clarify.
Inhibitors of SERT, including the 5-HT selective re-uptake inhibitors (SSRIs), are well known for their use in the treatment of anxiety disorders, depression, and obsessive-compulsive disorders. Naturally, therefore, polymorphisms in the SERT gene have been extensively studied in humans as potential risk determinants of neuropsychiatric disorders (for detailed reviews, see Lesch et al., 1996; Hahn and Blakely, 2007). Early studies identified two common polymorphisms in the promoter region (5-HTTLPR) and intron 2 (VNTR-2) of SLC6A4 (Lesch et al., 1996; Fiskerstrand et al., 1999). The 5-HTTLPR, a polymorphic, repeat structure 5′ of the sites of initiation for SERT RNA transcription. Initial studies indicated an association of the 5-HTTLPR with anxiety traits in nonclinical subjects and, more recently, in risk for depression and suicide, particularly in the context of early adverse life events (Caspi et al., 2003; Wankerl et al., 2010). These studies, however, have not met with consistent replication, though most recent analyses across larger number of studies where consistency of studies can be maintained, point to significant effects when specific components of mood disorders are assessed (Blakely and Veenstra-Vanderweele, 2011).
As common, genetic variation such as the 5-HTTLPR is often found in noncoding or complex regulatory regions of genes, it can be difficult to establish mechanistic contributions to disease risk. On the other hand, SLC6A4 coding variation is rare, with the most common variation (Gly56Ala in the SERT N-terminus) exhibiting an allele frequency generally under 1% (Glatt et al., 2001; Sutcliffe et al., 2005). Though the low frequency of these structural changes in SERT protein might appear seemingly irrelevant to population disease risk, they can be quite functionally penetrant (Prasad et al., 2009), and are easier to study in vitro since SERT activity and regulation can be reconstituted following heterologous expression. Such efforts can permit the identification of broader mechanisms whose compromised control of SERT expression, activity, and regulation could phenocopy SERT-influenced disorders and even identify new targets within which more common gene variation can contribute more broadly to disease risk (Veenstra-Vanderweele and Blakely, in press). Indeed, in several cases, familial segregation of rare SERT variants has provided strong support for a contribution of disrupted 5-HT signaling neuropsychiatric disorders, including OCD (Ozaki et al., 2003) and autism (Sutcliffe et al., 2005; Prasad et al., 2009). These studies draw attention to a network of SERT-regulatory signaling pathways that may bear additional disease-associated gene variation, or be modified by epigenetic influences, such as early-life adverse events. Although these alterations can institute limits to the behavioral flexibility needed to deal with everyday life, their origins in perturbed 5-HT signaling suggest opportunities for pharmacological treatments that, properly targeted, can provide relief from many disabling symptoms.
The translocation of 5-HT across the plasma membrane by SERT is a multi-step process that involves the binding of neurotransmitter along with Na+/Cl−, followed by protein conformational changes, the intracellular release of substrates, and the binding of K+ to drive return of the unloaded carrier for subsequent rounds of transport. Together, these steps energize the 5-HT transport mechanism and provide multiple steps for regulation of SERT activity. But exactly where critical events, such as 5-HT binding, internal occlusion, and membrane transit, occur in SERT have only recently begun to emerge. Since the cloning of SERT cDNAs two decades ago (Ramamoorthy et al., 1993a), many mutation studies have been conducted with the hope of pinpointing key features of substrate and antagonist binding, as well as determinants of transporter regulation. The high-resolution crystal structure of a bacterial SLC6 transporter, LeuTAa (Yamashita et al., 2005; Singh et al., 2008), provided a critical framework on which the effects of such could be appreciated in molecular terms. Unfortunately LeuTAa lacks key cytoplasmic sequences (most of the N- and C-termini and cytoplasmic loops are either unordered or absent) that have been implicated in transporter regulation. As we will discuss later in the text, the LeuTAa crystal structure has validated our hypotheses concerning the location of 5-HT and high-affinity antagonist-binding sites in SERT and encouraged further exploration of their manipulation in vivo.
Structural determinants of SERT regulation remain to be fully elucidated, though a combination of engineered and natural gene variation has provided remarkable opportunities to elucidate opportunities for drug development as well as to elucidate normal regulatory networks where disruption can support neuropsychiatric diseases associated with compromised 5-HT signaling. Neuropsychiatric diseases are polygenic in nature and feature many opportunities for environmental triggers or modulation. Thus, though the connections between SERT dysfunction and disorders such as depression and autism continue to be explored, deciphering the exact roles of SERT requires a more nuanced understanding of (1) the transporter’s interactions with other gene products, (2) a careful dissection of where among the many sites of SERT expression such interactions are relevant, and (3) whether these interactions are most critical during early life or in the adult (or both). In this context, the discussions of our efforts to examine the impact of natural and engineered genetic variation in this review should be seen not as a comprehensive review of the literature, but more as an outline of possible new entry points and tools that can assist us in understanding the complex roles of SERT and 5-HT in human behavior and drug action.
IDENTIFICATION OF KEY SITES FOR SSRI INTERACTIONS AT SERT: Tyr95 AND Ile172
Before the structural insights afforded by the LeuTAa structure, we developed a new approach to probe SERT structure, species-scanning mutagenesis (Barker et al., 1998; Adkins et al., 2001; Henry et al., 2006b), whereby amino acid sequences are sequentially interconverted between SERT species variants, to search for sites of high-affinity interaction of 5-HT and SSRIs. Key to this effort is the fact that many SSRIs, including citalopram and fluoxetine, as well as 5-HT analogs (but not 5-HT itself) display substantial differences in potency for inhibition of 5-HT uptake at human and Drosophila melanogaster SERT. By switching amino acid residues in human SERT to their fly counterparts (and vice versa) in regions of the transporter implicated by chimera studies in 5-HT or SSRI recognition, we identified Tyr95 and Ile172 in human SERT as two major structural determinants for binding of many antidepressant medications, as well as cocaine (Barker et al., 1998; Henry et al., 2006b). With the elucidation of the LeuTAa structure, the location of Tyr95 and Ile172 in transmembrane domain (TM) 1 and 3, respectively, was revealed to correspond to sites on opposite sides of the leucine-binding site (Yamashita et al., 2005). By inference from homology models, these residues could then be predicted to lie on opposite sides of the binding site for 5-HT in SERT (Henry et al., 2006a) (Fig. 1).
Fig. 1.
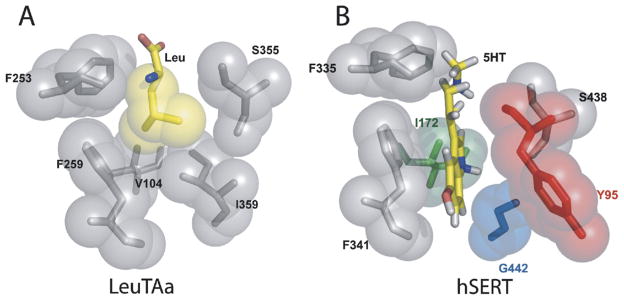
Model of the 5-HT binding pocket in human SERT. (A) Hydrophobic binding pocket of LeuTAa-transporter side chains (gray) and L-leucine side chain (yellow) are represented by stick models surrounded by semi-transparent Van der Waals surface spheres (Figure from Yamashita et al., 2005). (B) Modeling of 5-HT within the substrate-binding site of hSERT. hSERT residues were mapped onto LeuTAa coordinates and probable side-chain conformations determined by SCWRL3 rotomer library analysis while maintaining the LeuTAa backbone. Amino acid changes in the binding pocket are represented as colored residues: Y95 (red), I172 (green), and G442 (blue). Figure reproduced from Henry et al. (2006a), by permission from Elsevier.
Nearly identical losses in antagonist potency were seen by substitution of Ile172 with the corresponding fly residue (Met) at mouse SERT (Henry et al., 2006b). The reverse mutation, where a human residue is introduced into the Drosophila SERT at the corresponding position to Ile172 produced an increase in inhibitor sensitivity (Henry et al., 2006b). Interestingly, potency of the SSRI paroxetine was unaffected by the Ile172Met mutation, as was the apparent affinity of 5-HT itself (5-HT analogs do differentially respond to the mutation; Adkins et al., 2001). We speculate that either paroxetine binds to a site outside of the substrate-binding site or that it occludes the substrate-binding site in an orientation that does not utilize the hydrophobic interactions afforded by the Ile172 side chain. The insensitivity of 5-HT to the Ile172Met mutation is consistent with a requirement for more bulky 5-HT analogs that can mimic the larger size of an SSRI to interact with Tyr95.
Since the initial study, a series of structures with bound SSRIs and TCAs have been reported (Singh et al., 2007; Zhou et al., 2007, 2009). These studies revealed that SERT antagonists bind above, and stabilize, an extracellular gate in LeuTAa, thus effectively prohibiting substrate access and transport in a noncompetitive mechanism. However, the interactions of most SERT antagonists are competitive in nature and, as inhibitory potency and binding affinity are profoundly reduced by Tyr95 and Ile172 mutations that lie much deeper in the membrane, it is likely that studies with the bacterial transporters point more to potential, lower affinity contacts for SSRIs and cocaine, or are only relevant for the binding of these agents to bacterial SLC6 proteins (Henry et al., 2007). Recently, Javitch and coworkers provided evidence of a second substrate-binding site in LeuTAa and mammalian dopamine (DA) transporters lying in the general area of the external SSRI site (Shan et al., 2011; Zhao et al., 2011). These findings have not been without controversy (Piscitelli et al., 2010), but merits future consideration for its potential importance in allosteric drug development. Of course, neither the sites around the substrate-binding pocket, nor the outer gate sites, might be relevant in vivo, but as we discuss later in the text, this does not appear to be the case.
CREATION AND CHARACTERIZATION OF THE SERT Ile172Met MOUSE: A NEW TOOL TO DISSECT THE CONTRIBUTIONS OF SEROTONIN SIGNALING FOR SSRI AND COCAINE ACTION
The discrepancy between the delay (3– 6 weeks) in the clinical efficacy of antidepressants, as compared with their actions (within minutes) in blocking brain transporters, has been clear to psychopharmacologists and clinicians for decades. This distinction suggests that other mechanisms besides, or beyond, the blockade of monoamine reuptake are required for the treatment of depression (Duman and Monteggia, 2006; Baudry et al., 2010). Indeed, SSRIs have been found to exert their actions on many other targets besides SERT (Lenkey et al., 2006; Levkovitz et al., 2007; Keiser et al., 2009; Rainey et al., 2010), and there are many CNS targets that we do not even yet know how to assay. Certainly, long-term plasticities arise in gene (Sillaber et al., 2008) and protein (Abumaria et al., 2007) expression following chronic antidepressant treatments that are not evident with short-term administration.
The relative lack of specificity of some SERT antagonists, such as cocaine (Torres et al., 2003), presents further challenges in attributing specific actions to one or another monoamine system. Although much evidence supports the key role of altered DA signaling in the reinforcing and addictive actions of cocaine, genetic loss of DA transporters (DATs) in mice does not eliminate cocaine-conditioned place preference (Rocha et al., 1998), unlike a DAT/SERT double KO (Sora et al., 2001). These data are difficult to link to physiological aspects of DA and 5-HT signaling given the significant compensatory changes that arise with the constitutive DAT and SERT KOs. Nonetheless, these findings remind us of functional interactions between 5-HT and DA systems (Carlsson et al., 2007; Navailles and De Deurwaerdere, 2010; Larsen et al., 2011), particularly given the higher affinity that SERT exhibits for cocaine as compared with DAT (Ritz et al., 1987; Kozikowski et al., 1998).
The uncertainties noted previously regarding the actions of antidepressants and psychostimulants encouraged us to develop an animal model that can compromise drug action at SERT in the absence of compensatory changes in 5-HT signaling (Murphy et al., 2008). Our discovery of sites that dictate high-affinity recognition at SERT provided the opportunity to achieve this goal. As neither 5-HT recognition nor 5-HT transport is compromised by the Ile172Met mutation in either human or mouse SERT, we hypothesized that a substitution of genomic sequence that establishes the Met172 substitution in “knock-in” mice could result in an animal with normal 5-HT signaling, yet compromised SERT-mediated drug sensitivities. Initial ex vivo and in vivo pharmacological assessments showed complete agreement with our kinetic and pharmacological observations using transfected cell models (Fig. 2).
Fig. 2.

Impact of SERT Met172 substitution on synaptosomal 5-HT transport inhibitor potency. (A) 5-HT, (B) paroxetine, (C) fluoxetine, (D) cocaine, (E) ctalopram, and (F) escitalopram were assessed for their ability to compete with [3H] 5-HT uptake. Fluoxetine, cocaine, citalopram, and escitalopram demonstrated significantly reduced potencies in SERT M172 relative to SERT I172 synaptosomes. Reprinted from Thompson et al. (2011) with permission.
Synaptosomes prepared from the Met172 mice display significant reductions in potency for the SSRIs fluoxetine and citalopram (but not for paroxetine as expected), as well as a loss of sensitivity to cocaine and cocaine analogs, such as RTI-55. In contrast, SERT protein levels, SERT 5-HT transport kinetics, and the affinity of 5-HT at SERT appear to be identical to that of the wild-type SERT. Additionally, brain 5-HT levels are equivalent, comparing mutant and wild-type animals. Finally, in vitro brain slice studies and in vivo amperometry experiments demonstrate a loss of antidepressant action in producing 5HT1A autoreceptor inhibition of raphe firing and/or reductions in SERT-dependent 5-HT clearance, respectively.
The behavioral impact of acute treatment with antidepressants can be assessed in the tail suspension and forced-swim tests (TST and FST, respectively). The well-documented strain differences in responses to SSRIs (Jacobson and Cryan, 2007) provide challenges to studies of the initial line of SERT Met172 mice as the 129S6/S4 background used to generate these mice displays an opposite response to SSRIs than most mouse strains. Although SERT proteins exhibit a two amino acid difference in their structure, comparing 129S6 lines with some other commonly used strains (such as C57BL/6) as noted below, the difference in behavioral response to SSRIs may also derive from other elements of the 129S6/S4 background. Regardless, in the FST, SERT Met172 mice display a significant loss in citalopram and fluoxetine sensitivity, whereas paroxetine sensitivity appears normal. Together, these findings encourage further use of the SERT Met172 model to elucidate 5-HT dependent and independent contributions to the actions of antidepressants and cocaine, and may be of benefit in developing novel treatments for mood disorders that do not gain efficacy through SERT blockade. We are currently assessing gene expression changes in the Met172 mice that arise following acute and chronic antagonist administration as a probe for SERT dependent and independent actions of SSRIs and cocaine. In this regard, Bonnin and coworkers have recently used our mice to reveal a novel action of the SSRI during development that is SERT independent, but rather mediated by sigma receptors (Bonnin et al., unpublished observation).
STRAIN-BASED GENETIC VARIATION IN MOUSE SERT AND ITS USE IN IDENTIFYING NOVEL PHENOTYPES LINKED TO 5-HT SIGNALING
The inbred and clonal nature of mouse strains in common use by neuroscientists provides an important opportunity to limit variation in experiments, and to thereby reduce the numbers of animals that must be tested to provide confidence in results. These lines, however, also present challenges, as noted previously, due to differences in physiology and behavior across strains that often require extension of studies to other strains, an effort that incurs significant time and expense. Interestingly, the genetic and functional diversity of mouse strains can also be useful in identifying the molecular underpinnings of complex traits, as well as in defining a broader molecular and physiological impact of medications. Essential to this strategy is the presence of genetic or functional variation between strains that can be exploited to identify heritable influences (Holmes et al., 2002; Lu et al., 2008). We identified coding variation in SERT between C57BL/6J (Gly39 and Lys152=GK haplotype) and DBA/2J (Glu39 and Arg152=ER haplotype) and, using heterologous expression studies, demonstrated that this sequence variation imparted significant differences in 5-HT transport capacity (Carneiro et al., 2009) (Fig. 3).
Fig. 3.
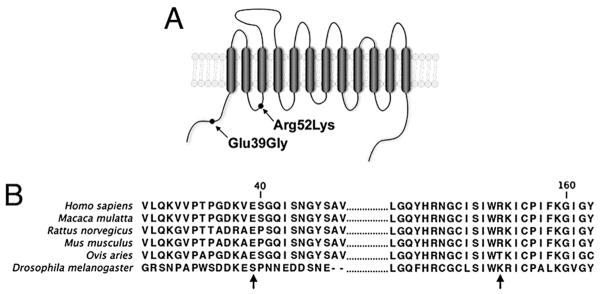
Sites of coding differences between DBA/2J and C57BL/6 SERT proteins. (A) Schematic illustration of the location of the amino acid changes in Slc6a4 between DBA/2J (Glu39/Arg172) and C57BL/6 (Gly39/Lys172). (B) Alignment of SERT protein sequences from human, macaque, rat, mouse, sheep, and fruit fly to illustrate relative conservation at positions analogous to amino acids 39 and 172 of mouse SERT. Reprinted from Carneiro et al. (2009) with permission.
These findings were particularly useful since a large panel of recombinant inbred mice, formed by breeding C57BL/6J and DBA2J parents (termed BXD lines), exists that differentially segregate millions of sequence variants as well as many biochemical and behavioral phenotypes (Taylor et al., 1977, 1999; Peirce et al., 2004). The dense genotyping of the BXD lines, along with the full sequencing of the parental genomes, provides the researcher with a powerful paradigm to map quantitative trait loci (QTLs) supporting functional variation (Lu et al., 2008; Gaglani et al., 2009). Additionally, correlations can also be identified between biochemical or functional traits due to their co-segregation across BXD lines, and then their common genetic underpinnings can be pursued (Koutnikova et al., 2009; Tapocik et al., 2009).
The major difference between the 5-HT transport kinetics of the ER and GK SERTs was found to be in their maximal transport capacity (Vmax-ER has higher activity than GK) rather than the transporter’s apparent affinity for 5-HT (Km) (Carneiro et al., 2009). In vitro mutagenesis studies revealed that the contributor to this activity difference is the E/G substitution that interestingly lies near a site in the N-terminus impacting SERT regulation (see later in the text). Although BXD strains that harbor the GK haplotype have reduced 5-HT uptake than those harboring ER haplotype, brain 5-HT levels are comparable between strains. This may be expected as alterations in synthesis could readily offset changes in 5-HT reuptake as long as the alterations in 5-HT transport are not severe, as with the SERT knockout mouse (Bengel et al., 1998). Regardless, we reasoned that differences in the ability of different strains to clear 5-HT after release could lead to changes in extracellular 5-HT levels for the same amount of nerve terminal excitation and produce altered activation of pre-and postsynaptic 5-HT receptors in vivo. To explore this hypothesis, we conducted multiple behavioral assays on lines differentially segregating the SERT ER and GK coding variation (Carneiro et al., 2009). In the TST, GK expressing lines display reduced basal immobility time relative to ER lines, though both lines exhibited equivalent responses to the SSRI citalopram. We also found that GK mice spend less time in the chamber center of open field test (OFT), suggesting a greater level of anxiety in these animals that may arise from constitutively altered 5-HT clearance capacity.
The community of scientists studying the phenotypes of BXD lines has created a phenotype database (http://www.genenetwork.org) where the quantitative levels of multiple traits for each line have been archived. By querying this database to examine traits whose levels are influenced by the SERT GK/ER haplotypes, we identified multiple traits correlating with SERT variation that were previously associated with 5-HT signaling pathways, such as body weight (Garattini et al., 1992) and ethanol consumption (Kranzler et al., 2002). Consistent with the interactions of DA and 5-HT pathways noted earlier in this review, multiple DA-related phenotypes were also found to segregate with SERT haplotype (Carneiro et al., 2009). Finally, and perhaps the most important use of the BXD trait database, we identified multiple, unexpected SERT-associated traits. For example, the SERT ER/GK haplotype distinguishes levels of handling-induced convulsions, cocaine-induced seizure activity, retinal area, and T cell proliferation. Additionally, we found that brain iron levels were influenced. To provide a direct test of whether SERT influences one of these traits, we examined iron levels in SERT KO mice. Indeed, we found midbrain iron levels to be significantly decreased in the KO animals vs. wild-type animals.
It is worth noting that other genes participating in 5-HT signaling harbor coding variation in the BXD parental strains. These findings indicate that the BXD recombinant inbred lines represent a rich opportunity to dissect multiple genetic interactions that influence 5-HT signaling and behavior. For example, the tryptophan hydroxylase 2 (Tph2) Arg447 allele has a twofold lower enzyme activity than Pro447 allele (Zhang et al., 2004; Kulikov et al., 2005), and coding variation across strains has also been found in Slc18a2 gene (Vesicular Monoamine Transporter 2, VMAT2), though the direct, functional impact of this variation remains an area of investigation (Crowley et al., 2006). Interestingly, mice with different Tph2 or Slc18a2 genotypes exhibit varied responses to SSRIs (Cervo et al., 2005; Crowley et al., 2006). C57BL/6J mice express the high-functioning Pro447 Tph2 allele and the P117/P505 haplotype in Slc18a2 in the background of the low-capacity GK SERT haplotype, whereas DBA/2J expresses the low-functioning Arg447 Tph2, L117/S505 haplotype in Slc18a2 in the context of the high-capacity ER SERT haplotype. In our study, we found epistatic effects involving SERT and Tph2 in retinal ganglion cell number, cerebellar weight, cholesterol levels after high-fat diet, and adult dentate gyrus cell proliferation (Carneiro et al., 2009). The genetic variation present in these and other molecular determinants of 5-HT signaling is of great importance to our current efforts to decipher risk in neuropsychiatric disorders in man that derive from the intersection of gene and environmental variation. Further studies with the BXD lines, particularly employing 5-HT signaling manipulations (Canal et al., 2010), may provide important clues as to how altered genomic architecture can drive risk for 5-HT linked brain disorders and under what conditions.
HUMAN SERT CODING VARIATION
As noted earlier, SERT coding variation is rare in humans. Despite this, we found that publically accessible sequence databases (e.g. dbSNP) contained multiple changes in SERT exon sequences that encoded rare SERT coding variants. We suspected that the general ignorance of these variants represented less a lack of functional impact on but rather the fact that population geneticists could not assess their impact in population studies. Therefore, we generated and expressed the 10 known SERT coding variants established and found that multiple variants exhibited a gain of 5-HT transport function. Even more remarkably, we found that multiple variants displayed a loss of sensitivity to PKG and p38 MAPK-linked signaling pathways (Prasad et al., 2005). In a completely unrelated effort, we were establishing the impact of rare coding variation in the SERT gene in subjects with autism, identifying five coding variants that associated with high levels of rigid-compulsive traits as well as sensory aversion (Anderson, 2002; Sutcliffe et al., 2005). We have since learned that all five of these variants (Fig. 4), including two that were present in our parallel study of deposited SERT variants, exhibit elevated 5-HT transport activity.
Fig. 4.
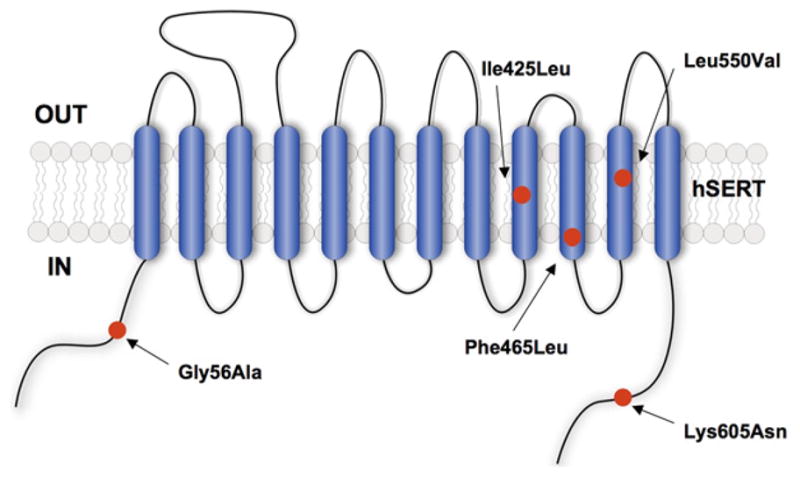
Location of autism-associated SERT coding variants. Five coding variants are overlaid on an hSERT topology model with 12 transmembrane (TM) domains. Reprinted from Prasad et al. (2009) with permission.
Studies performed with both transfected cells and cultured lymphoblast lines from variant-carrying probands also demonstrated that multiple variants are refractory to PKG- and p38 MAPK-dependent regulation (Prasad et al., 2009). Detailed kinetic studies indicate that I425L, F465L, and L550V variants all possess higher affinity for 5-HT as well as higher transport capacity. In comparison, the G56A and K605N variants bear a comparable Vmax to wild-type alleles along with an increased affinity for 5-HT. Despite this, all SERT variants display similar levels of SERT protein expression, supporting the idea that SERT coding variants bear alterations in post-translational regulation. For the variants with both higher Km and Vmax values, enhanced 5-HT uptake is accompanied by increased levels of surface SERT expression as measured by surface protein biotinylation (Prasad et al., 2009). However, the elevated SERT activity for G56A and K605N variants is not accounted for by elevated surface protein expression. Interestingly, the G56A variant is also hyperphosphorylated in transfected cells relative to wild-type SERT, and phosphorylation cannot be further enhanced by PKG activation (Prasad et al., 2005). These findings led us to hypothesize that mechanisms sustaining reduced synaptic 5-HT availability during development could increase risk of autism traits. Moreover, the nature of the functional perturbations we identified point to regulatory networks involving the control of PKG-dependent surface trafficking and p38 MAPK-dependent catalytic function as points of focus for future studies (Steiner et al., 2009). We suspect that altered regulation of SERT imposed by SERT gene variation altered regulation can modify the dynamic range achievable by SERT to track changes in 5-HT signaling.
An impact on SERT coding variation on brain development would not be limited to SERT-expressing neurons since 5-HT serves as a major axonal guidance molecule in the developing brain (Haydon et al., 1984; Bonnin et al., 2007). Recently, Bonnin and coworkers identified the placenta as an unrecognized source for 5-HT during early brain development (Bonnin et al., 2011). SERTs are also highly expressed in the placenta, and thus we must also consider whether the functional impact of SERT variants on brain development originates with malfunction in the periphery, not the brain. We recently engineered the most common of the autism-associated, gain of function SERT variants (G56A) variant into mice (Veenstra-Vanderweele et al., 2009). Studies with this model are ongoing but we are optimistic that this model will prove suitable for a mechanistic dissection of the contribution of human SERT gene variation to physiology and behavior, as well as risk for brain disorders associated with compromised 5-HT signaling. As noted earlier, inbred strains of mice also display different SERT coding sequences, with one variant (G39E) lying close to the autism-associated variant G56A. Perhaps the different behaviors of inbred strains of mice speak more clearly as to risk for neuropsychiatric disorders than we have previously recognized, and at least some of that risk derives from differences in their 5-HT systems.
CONCLUSIONS
The study of 5-HT and its control mechanisms, to some, is understandably a well-trodden field, where surely all stones have been turned. However, to those working in the area, many issues remain unclear, and the need for new models and tools is paramount. Previously, we have described three new models that have emerged from our studies of SERT variation. The first, knock-in mice bearing an SSRI and cocaine sensitivity-disrupting mutation, has no direct parallel in human coding variation. Thus, the utility of this model lies not in an understanding of the differential responses of individuals to antidepressants or drugs of abuse, but in their use to tease apart the complex actions of these drugs, elucidating what truly is 5-HT dependent and what is not. Data from this model, now freely available to others in the community, may lead us to better ways to treat subjects with 5-HT associated disorders and/or eliminate side effects. We often assume that because some agents have been tested for decades that we fully understand their actions, not recognizing that assays do not exist for many gene products at all, and that, with chronic dosing, even highly specific reagents can build up to levels that render nonspecific targets more important, if not essential to their action.
Our other two examples of novel models for the study of 5-HT biology derive not from SERT variants engineered to track down a specific facet of transporter function, but rather from natural genetic variation whose revealed functional impact has revealed new facets of 5-HT action that were largely unknown. One is a product of the generation of mouse strains by “mouse fanciers,” who sought to generate animals, as with dog breeding, with unique physical and behavioral traits. The second derives from humans with unique behavioral traits, in this case producing changes that can limit social and intellectual development and impact the lives of children and families for decades. Remarkably, both of the latter models further stimulate our interests in SERT regulation as a critical determinant of 5-HT biology. Studies in heterologous expression systems and ex vivo preparations, such as synaptosomes, have established such regulatory capacities. However, the contribution of this regulation to physiology and behavior remains largely unexplored. Thus, whether perturbed SERT regulation underlies disease risk has been limited to speculation. We believe that further study of recombinant inbred lines bearing SERT, TPH2, and VMAT2 regulation can elucidate molecular and physiological relationships heretofore unsuspected. Finally, we feel that the study of disease-associated natural variation in human SERT using engineered mouse models will identify actions of 5-HT that we would be unable to detect in human studies. In so doing, we suspect that we will learn how limited our understanding of 5-HT actions has been.
Acknowledgments
We gratefully acknowledge support of the Brain and Behavior Research Foundation and NIH awards P50MH078028, MH094527, and HD065278 to R.D.B.
Abbreviations
- SERT
serotonin transporter
- SSRI
5-HT selective re-uptake inhibitors
References
- Abumaria N, Rygula R, Hiemke C, Fuchs E, Havemann-Reinecke U, Rüther E, Flügge G. Effect of chronic citalopram on serotonin-related and stress-regulated genes in the dorsal raphe nucleus of the rat. Eur Neuropsychopharmacol. 2007;17:417–429. doi: 10.1016/j.euroneuro.2006.08.009. [DOI] [PubMed] [Google Scholar]
- Adkins EM, Barker EL, Blakely RD. Interactions of tryptamine derivatives with serotonin transporter species variants implicate transmembrane domain I in substrate recognition. Mol Pharmacol. 2001;59:514–523. doi: 10.1124/mol.59.3.514. [DOI] [PubMed] [Google Scholar]
- Anderson GM. Genetics of childhood disorders: XLV. Autism, part 4: serotonin in autism. J Am Acad Child Adolesc Psychiatry. 2002;41:1513–1516. doi: 10.1097/00004583-200212000-00025. [DOI] [PubMed] [Google Scholar]
- Barkan T, Gurwitz D, Levy G, Weizman A, Rehavi M. Biochemical and pharmacological characterization of the serotonin transporter in human peripheral blood lymphocytes. Eur Neuropsychopharmacol. 2004;14:237–243. doi: 10.1016/S0924-977X(03)00107-X. [DOI] [PubMed] [Google Scholar]
- Barker EL, Perlman MA, Adkins EM, Houlihan WJ, Pristupa ZB, Niznik HB, Blakely RD. High affinity recognition of serotonin transporter antagonists defined by species-scanning mutagenesis. An aromatic residue in transmembrane domain I dictates species-selective recognition of citalopram and mazindol. J Biol Chem. 1998;273:19459–19468. doi: 10.1074/jbc.273.31.19459. [DOI] [PubMed] [Google Scholar]
- Baudry A, Mouillet-Richard S, Schneider B, Launay JM, Kellermann O. miR-16 targets the serotonin transporter: a new facet for adaptive responses to antidepressants. Science. 2010;329:1537–1541. doi: 10.1126/science.1193692. [DOI] [PubMed] [Google Scholar]
- Bengel D, Murphy DL, Andrews AM, Wichems CH, Feltner D, Heils A, Mössner R, Westphal H, Lesch KP. Altered brain serotonin homeostasis and locomotor insensitivity to 3, 4-methylenedioxy-methamphetamine (“Ecstasy”) in serotonin transporter-deficient mice. Mol Pharmacol. 1998;53:649–655. doi: 10.1124/mol.53.4.649. [DOI] [PubMed] [Google Scholar]
- Bhat GB, Block ER. Hypoxia directly increases serotonin transport by porcine pulmonary artery endothelial cell plasma membrane vesicles. Am J Respir Cell Mol Biol. 1990;3:363–367. doi: 10.1165/ajrcmb/3.4.363. [DOI] [PubMed] [Google Scholar]
- Blakely RD, Veenstra-Vanderweele J. Genetic indeterminism, the 5-HTTLPR, and the paths forward in neuropsychiatric genetics. Arch Gen Psychiatry. 2011;68:457–458. doi: 10.1001/archgenpsychiatry.2011.34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bliziotes MM, Eshleman AJ, Zhang XW, Wiren KM. Neurotransmitter action in osteoblasts: expression of a functional system for serotonin receptor activation and reuptake. Bone. 2001;29:477–486. doi: 10.1016/s8756-3282(01)00593-2. [DOI] [PubMed] [Google Scholar]
- Bonnin A, Goeden N, Chen K, Wilson ML, King J, Shih JC, Blakely RD, Deneris ES, Levitt P. A transient placental source of serotonin for the fetal forebrain. Nature. 2011;472:347–350. doi: 10.1038/nature09972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonnin A, Torii M, Wang L, Rakic P, Levitt P. Serotonin modulates the response of embryonic thalamocortical axons to netrin-1. Nat Neurosci. 2007;10:588–597. doi: 10.1038/nn1896. [DOI] [PubMed] [Google Scholar]
- Canal CE, Olaghere da Silva UB, Gresch PJ, Watt EE, Sanders-Bush E, Airey DC. The serotonin 2C receptor potently modulates the head-twitch response in mice induced by a phenethylamine hallucinogen. Psychopharmacology (Berl) 2010;209:163–174. doi: 10.1007/s00213-010-1784-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlsson T, Carta M, Winkler C, Björklund A, Kirik D. Serotonin neuron transplants exacerbate L-DOPA-induced dyskinesias in a rat model of Parkinson’s disease. J Neurosci. 2007;27:8011–8022. doi: 10.1523/JNEUROSCI.2079-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carneiro AM, Airey DC, Thompson B, Zhu CB, Lu L, Chesler EJ, Erikson KM, Blakely RD. Functional coding variation in recombinant inbred mouse lines reveals multiple serotonin transporter-associated phenotypes. Proc Natl Acad Sci U S A. 2009;106:2047–2052. doi: 10.1073/pnas.0809449106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, Harrington H, McClay J, Mill J, Martin J, Braithwaite A, Poulton R. Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science. 2003;301:386–389. doi: 10.1126/science.1083968. [DOI] [PubMed] [Google Scholar]
- Cervo L, Canetta A, Calcagno E, Burbassi S, Sacchetti G, Caccia S, Fracasso C, Albani D, Forloni G, Invernizzi RW. Genotype-dependent activity of tryptophan hydroxylase-2 determines the response to citalopram in a mouse model of depression. J Neurosci. 2005;25:8165–8172. doi: 10.1523/JNEUROSCI.1816-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen JJ, Li Z, Pan H, Murphy DL, Tamir H, Koepsell H, Gershon MD. Maintenance of serotonin in the intestinal mucosa and ganglia of mice that lack the high-affinity serotonin transporter: abnormal intestinal motility and the expression of cation transporters. J Neurosci. 2001;21:6348–6361. doi: 10.1523/JNEUROSCI.21-16-06348.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crowley JJ, Brodkin ES, Blendy JA, Berrettini WH, Lucki I. Pharmacogenomic evaluation of the antidepressant citalopram in the mouse tail suspension test. Neuropsychopharmacology. 2006;31:2433–2442. doi: 10.1038/sj.npp.1301065. [DOI] [PubMed] [Google Scholar]
- Duman RS, Monteggia LM. A neurotrophic model for stress-related mood disorders. Biol Psychiatry. 2006;59:1116–1127. doi: 10.1016/j.biopsych.2006.02.013. [DOI] [PubMed] [Google Scholar]
- Fiskerstrand CE, Lovejoy EA, Quinn JP. An intronic polymorphic domain often associated with susceptibility to affective disorders has allele dependent differential enhancer activity in embryonic stem cells. FEBS Lett. 1999;458:171–174. doi: 10.1016/s0014-5793(99)01150-3. [DOI] [PubMed] [Google Scholar]
- Gaglani SM, Lu L, Williams RW, Rosen GD. The genetic control of neocortex volume and covariation with neocortical gene expression in mice. BMC Neurosci. 2009;10:44. doi: 10.1186/1471-2202-10-44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garattini S, Bizzi A, Codegoni AM, Caccia S, Mennini T. Progress report on the anorexia induced by drugs believed to mimic some of the effects of serotonin on the central nervous system. Am J Clin Nutr. 1992;55:160S–166S. doi: 10.1093/ajcn/55.1.160s. [DOI] [PubMed] [Google Scholar]
- Gaspar P, Cases O, Maroteaux L. The developmental role of serotonin: news from mouse molecular genetics. Nat Rev Neurosci. 2003;4:1002–1012. doi: 10.1038/nrn1256. [DOI] [PubMed] [Google Scholar]
- Glatt CE, DeYoung JA, Delgado S, Service SK, Giacomini KM, Edwards RH, Risch N, Freimer NB. Screening a large reference sample to identify very low frequency sequence variants: comparisons between two genes. Nat Genet. 2001;27:435–438. doi: 10.1038/86948. [DOI] [PubMed] [Google Scholar]
- Guilluy C, Eddahibi S, Agard C, Guignabert C, Izikki M, Tu L, Savale L, Humbert M, Fadel E, Adnot S, Loirand G, Pacaud P. RhoA and Rho kinase activation in human pulmonary hypertension: role of 5-HT signaling. Am J Respir Crit Care Med. 2009;179:1151–1158. doi: 10.1164/rccm.200805-691OC. [DOI] [PubMed] [Google Scholar]
- Hahn MK, Blakely RD. The functional impact of SLC6 transporter genetic variation. Annu Rev Pharmacol Toxicol. 2007;47:401–441. doi: 10.1146/annurev.pharmtox.47.120505.105242. [DOI] [PubMed] [Google Scholar]
- Hansson SR, Mezey E, Hoffman BJ. Serotonin transporter messenger RNA in the developing rat brain: early expression in serotonergic neurons and transient expression in non-serotonergic neurons. Neuroscience. 1998;83:1185–1201. doi: 10.1016/s0306-4522(97)00444-2. [DOI] [PubMed] [Google Scholar]
- Hansson SR, Mezey E, Hoffman BJ. Serotonin transporter messenger RNA expression in neural crest-derived structures and sensory pathways of the developing rat embryo. Neuroscience. 1999;89:243–265. doi: 10.1016/s0306-4522(98)00281-4. [DOI] [PubMed] [Google Scholar]
- Haydon PG, McCobb DP, Kater SB. Serotonin selectively inhibits growth cone motility and synaptogenesis of specific identified neurons. Science. 1984;226:561–564. doi: 10.1126/science.6093252. [DOI] [PubMed] [Google Scholar]
- Hendricks TJ, Fyodorov DV, Wegman LJ, Lelutiu NB, Pehek EA, Yamamoto B, Silver J, Weeber EJ, Sweatt JD, Deneris ES. Pet-1 ETS gene plays a critical role in 5-HT neuron development and is required for normal anxiety-like and aggressive behavior. Neuron. 2003;37:233–247. doi: 10.1016/s0896-6273(02)01167-4. [DOI] [PubMed] [Google Scholar]
- Henry LK, Defelice LJ, Blakely RD. Getting the message across: a recent transporter structure shows the way. Neuron. 2006a;49:791–796. doi: 10.1016/j.neuron.2006.03.002. [DOI] [PubMed] [Google Scholar]
- Henry LK, Field JR, Adkins EM, Parnas ML, Vaughan RA, Zou MF, Newman AH, Blakely RD. Tyr-95 and Ile-172 in transmembrane segments 1 and 3 of human serotonin transporters interact to establish high affinity recognition of antidepressants. J Biol Chem. 2006b;281:2012–2023. doi: 10.1074/jbc.M505055200. [DOI] [PubMed] [Google Scholar]
- Henry LK, Meiler J, Blakely RD. Bound to be different: neurotransmitter transporters meet their bacterial cousins. Mol Interv. 2007;7:306–309. doi: 10.1124/mi.7.6.4. [DOI] [PubMed] [Google Scholar]
- Holmes A, Wrenn CC, Harris AP, Thayer KE, Crawley JN. Behavioral profiles of inbred strains on novel olfactory, spatial and emotional tests for reference memory in mice. Genes Brain Behav. 2002;1:55–69. doi: 10.1046/j.1601-1848.2001.00005.x. [DOI] [PubMed] [Google Scholar]
- Jacobson LH, Cryan JF. Feeling strained? Influence of genetic background on depression-related behavior in mice: a review. Behav Genet. 2007;37:171–213. doi: 10.1007/s10519-006-9106-3. [DOI] [PubMed] [Google Scholar]
- Keiser MJ, Setola V, Irwin JJ, Laggner C, Abbas AI, Hufeisen SJ, Jensen NH, Kuijer MB, Matos RC, Tran TB, Whaley R, Glennon RA, Hert J, Thomas KL, Edwards DD, Shoichet BK, Roth BL. Predicting new molecular targets for known drugs. Nature. 2009;462:175–181. doi: 10.1038/nature08506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koutnikova H, Laakso M, Lu L, Combe R, Paananen J, Kuulasmaa T, Kuusisto J, Haring HU, Hansen T, Pedersen O, Smith U, Hanefeld M, Williams RW, Auwerx J. Identification of the UBP1 locus as a critical blood pressure determinant using a combination of mouse and human genetics. PLoS Genet. 2009;5:e1000591. doi: 10.1371/journal.pgen.1000591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozikowski AP, Araldi GL, Prakash KR, Zhang M, Johnson KM. Synthesis and biological properties of new 2beta-alkyl- and 2beta-aryl-3-(substituted phenyl)tropane derivatives: stereochemical effect of C-3 on affinity and selectivity for neuronal dopamine and serotonin transporters. J Med Chem. 1998;41:4973–4982. doi: 10.1021/jm9802564. [DOI] [PubMed] [Google Scholar]
- Kranzler H, Lappalainen J, Nellissery M, Gelernter J. Association study of alcoholism subtypes with a functional promoter polymorphism in the serotonin transporter protein gene. Alcohol Clin Exp Res. 2002;26:1330–1335. doi: 10.1097/01.ALC.0000030840.48315.40. [DOI] [PubMed] [Google Scholar]
- Kulikov AV, Osipova DV, Naumenko VS, Popova NK. Association between Tph2 gene polymorphism, brain tryptophan hydroxylase activity and aggressiveness in mouse strains. Genes Brain Behav. 2005;4:482–485. doi: 10.1111/j.1601-183X.2005.00145.x. [DOI] [PubMed] [Google Scholar]
- Larsen MB, Sonders MS, Mortensen OV, Larson GA, Zahniser NR, Amara SG. Dopamine transport by the serotonin transporter: a mechanistically distinct mode of substrate translocation. J Neurosci. 2011;31:6605–6615. doi: 10.1523/JNEUROSCI.0576-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lenkey N, Karoly R, Kiss JP, Szasz BK, Vizi ES, Mike A. The mechanism of activity-dependent sodium channel inhibition by the antidepressants fluoxetine and desipramine. Mol Pharmacol. 2006;70:2052–2063. doi: 10.1124/mol.106.026419. [DOI] [PubMed] [Google Scholar]
- Lesch KP, Bengel D, Heils A, Sabol SZ, Greenberg BD, Petri S, Benjamin J, Müller CR, Hamer DH, Murphy DL. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science. 1996;274:1527–1531. doi: 10.1126/science.274.5292.1527. [DOI] [PubMed] [Google Scholar]
- Levkovitz Y, Ben-Shushan G, Hershkovitz A, Isaac R, Gil-Ad I, Shvartsman D, Ronen D, Weizman A, Zick Y. Antidepressants induce cellular insulin resistance by activation of IRS-1 kinases. Mol Cell Neurosci. 2007;36:305–312. doi: 10.1016/j.mcn.2007.05.009. [DOI] [PubMed] [Google Scholar]
- Liu Y, Wei L, Laskin DL, Fanburg BL. Role of protein transamidation in serotonin-induced proliferation and migration of pulmonary artery smooth muscle cells. Am J Respir Cell Mol Biol. 2011;44:548–555. doi: 10.1165/rcmb.2010-0078OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu L, Wei L, Peirce JL, Wang X, Zhou J, Homayouni R, Williams RW, Airey DC. Using gene expression databases for classical trait QTL candidate gene discovery in the BXD recombinant inbred genetic reference population: mouse forebrain weight. BMC Genomics. 2008;9:444. doi: 10.1186/1471-2164-9-444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mercado CP, Kilic F. Molecular mechanisms of SERT in platelets: regulation of plasma serotonin levels. Mol Interv. 2010;10:231–241. doi: 10.1124/mi.10.4.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy DL, Fox MA, Timpano KR, Moya PR, Ren-Patterson R, Andrews AM, Holmes A, Lesch KP, Wendland JR. How the serotonin story is being rewritten by new gene-based discoveries principally related to SLC6A4, the serotonin transporter gene, which functions to influence all cellular serotonin systems. Neuropharmacology. 2008;55:932–960. doi: 10.1016/j.neuropharm.2008.08.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navailles S, De Deurwaerdere P. Presynaptic control of serotonin on striatal dopamine function. Psychopharmacology (Berl) 2010;213:213–242. doi: 10.1007/s00213-010-2029-y. [DOI] [PubMed] [Google Scholar]
- Ozaki N, Goldman D, Kaye WH, Plotnicov K, Greenberg BD, Lappalainen J, Rudnick G, Murphy DL. Serotonin transporter missense mutation associated with a complex neuropsychiatric phenotype. Mol Psychiatry. 2003;8:933–936. doi: 10.1038/sj.mp.4001365. [DOI] [PubMed] [Google Scholar]
- Peirce JL, Lu L, Gu J, Silver LM, Williams RW. A new set of BXD recombinant inbred lines from advanced intercross populations in mice. BMC Genet. 2004;5:7. doi: 10.1186/1471-2156-5-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piscitelli CL, Krishnamurthy H, Gouaux E. Neurotransmitter/ sodium symporter ortholog LeuT has a single high-affinity substrate site. Nature. 2010;468:1129–1132. doi: 10.1038/nature09581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prasad HC, Steiner JA, Sutcliffe JS, Blakely RD. Enhanced activity of human serotonin transporter variants associated with autism. Philos Trans R Soc Lond B Biol Sci. 2009;364:163–173. doi: 10.1098/rstb.2008.0143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prasad HC, Zhu CB, McCauley JL, Samuvel DJ, Ramamoorthy S, Shelton RC, Hewlett WA, Sutcliffe JS, Blakely RD. Human serotonin transporter variants display altered sensitivity to protein kinase G and p38 mitogen-activated protein kinase. Proc Natl Acad Sci U S A. 2005;102:11545–11550. doi: 10.1073/pnas.0501432102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rainey MM, Korostyshevsky D, Lee S, Perlstein EO. The antidepressant sertraline targets intracellular vesiculogenic membranes in yeast. Genetics. 2010;185:1221–1233. doi: 10.1534/genetics.110.117846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramamoorthy S, Bauman AL, Moore KR, Han H, Yang-Feng T, Chang AS, Ganapathy V, Blakely RD. Antidepressant- and cocaine-sensitive human serotonin transporter: molecular cloning, expression, and chromosomal localization. Proc Natl Acad Sci U S A. 1993a;90:2542–2546. doi: 10.1073/pnas.90.6.2542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramamoorthy S, Leibach FH, Mahesh VB, Ganapathy V. Partial purification and characterization of the human placental serotonin transporter. Placenta. 1993b;14:449–461. doi: 10.1016/s0143-4004(05)80465-5. [DOI] [PubMed] [Google Scholar]
- Ritz MC, Lamb RJ, Goldberg SR, Kuhar MJ. Cocaine receptors on dopamine transporters are related to self-administration of cocaine. Science. 1987;237:1219–1223. doi: 10.1126/science.2820058. [DOI] [PubMed] [Google Scholar]
- Rocha BA, Fumagalli F, Gainetdinov RR, Jones SR, Ator R, Giros B, Miller GW, Caron MG. Cocaine self-administration in dopamine-transporter knockout mice. Nat Neurosci. 1998;1:132–137. doi: 10.1038/381. [DOI] [PubMed] [Google Scholar]
- Schroeter S, Blakely RD. Drug targets in the embryo. Studies on the cocaine- and antidepressant-sensitive serotonin transporter. Ann N Y Acad Sci. 1996;801:239–255. doi: 10.1111/j.1749-6632.1996.tb17446.x. [DOI] [PubMed] [Google Scholar]
- Shan J, Javitch JA, Shi L, Weinstein H. The substrate-driven transition to an inward-facing conformation in the functional mechanism of the dopamine transporter. PLoS One. 2011;6:e16350. doi: 10.1371/journal.pone.0016350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sillaber I, Panhuysen M, Henniger MS, Ohl F, Kuhne C, Putz B, Pohl T, Deussing JM, Paez-Pereda M, Holsboer F. Profiling of behavioral changes and hippocampal gene expression in mice chronically treated with the SSRI paroxetine. Psychopharmacology (Berl) 2008;200:557–572. doi: 10.1007/s00213-008-1232-6. [DOI] [PubMed] [Google Scholar]
- Singh SK, Piscitelli CL, Yamashita A, Gouaux E. A competitive inhibitor traps LeuT in an open-to-out conformation. Science. 2008;322:1655–1661. doi: 10.1126/science.1166777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh SK, Yamashita A, Gouaux E. Antidepressant binding site in a bacterial homologue of neurotransmitter transporters. Nature. 2007;448:952–956. doi: 10.1038/nature06038. [DOI] [PubMed] [Google Scholar]
- Smith PA, Proks P, Ashcroft FM. Quantal analysis of 5-hydroxytryptamine release from mouse pancreatic beta-cells. J Physiol. 1999;521 (Pt 3):651–664. doi: 10.1111/j.1469-7793.1999.00651.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sora I, Hall FS, Andrews AM, Itokawa M, Li XF, Wei HB, Wichems C, Lesch KP, Murphy DL, Uhl GR. Molecular mechanisms of cocaine reward: combined dopamine and serotonin transporter knockouts eliminate cocaine place preference. Proc Natl Acad Sci U S A. 2001;98:5300–5305. doi: 10.1073/pnas.091039298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steiner JA, Carneiro AM, Wright J, Matthies HJ, Prasad HC, Nicki CK, Dostmann WR, Buchanan CC, Corbin JD, Francis SH, Blakely RD. cGMP-dependent protein kinase Ialpha associates with the antidepressant-sensitive serotonin transporter and dictates rapid modulation of serotonin uptake. Mol Brain. 2009;2:26. doi: 10.1186/1756-6606-2-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutcliffe JS, Delahanty RJ, Prasad HC, McCauley JL, Han Q, Jiang L, Li C, Folstein SE, Blakely RD. Allelic heterogeneity at the serotonin transporter locus (SLC6A4) confers susceptibility to autism and rigid-compulsive behaviors. Am J Hum Genet. 2005;77:265–279. doi: 10.1086/432648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tapocik JD, Letwin N, Mayo CL, Frank B, Luu T, Achinike O, House C, Williams R, Elmer GI, Lee NH. Identification of candidate genes and gene networks specifically associated with analgesic tolerance to morphine. J Neurosci. 2009;29:5295–5307. doi: 10.1523/JNEUROSCI.4020-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor BA, Bedigian HG, Meier H. Genetic studies of the Fv-1 locus of mice: linkage with Gpd-1 in recombinant inbred lines. J Virol. 1977;23:106–109. doi: 10.1128/jvi.23.1.106-109.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor BA, Wnek C, Kotlus BS, Roemer N, MacTaggart T, Phillips SJ. Genotyping new BXD recombinant inbred mouse strains and comparison of BXD and consensus maps. Mamm Genome. 1999;10:335–348. doi: 10.1007/s003359900998. [DOI] [PubMed] [Google Scholar]
- Torres GE, Gainetdinov RR, Caron MG. Plasma membrane monoamine transporters: structure, regulation and function. Nat Rev Neurosci. 2003;4:13–25. doi: 10.1038/nrn1008. [DOI] [PubMed] [Google Scholar]
- Veenstra-Vanderweele J, Blakely RD. Networking in autism: leveraging genetic, biomarker and model system findings in the search for new treatments. Neuropsychopharmacol Rev. doi: 10.1038/npp.2011.185. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Veenstra-Vanderweele J, Jessen TN, Thompson BJ, Carter M, Prasad HC, Steiner JA, Sutcliffe JS, Blakely RD. Modeling rare gene variation to gain insight into the oldest biomarker in autism: construction of the serotonin transporter Gly56Ala knock-in mouse. J Neurodev Disord. 2009;1:158–171. doi: 10.1007/s11689-009-9020-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walther DJ, Peter JU, Winter S, Höltje M, Paulmann N, Grohmann M, Vowinckel J, Alamo-Bethencourt V, Wilhelm CS, Ahnert-Hilger G, Bader M. Serotonylation of small GTPases is a signal transduction pathway that triggers platelet alpha-granule release. Cell. 2003;115:851–862. doi: 10.1016/s0092-8674(03)01014-6. [DOI] [PubMed] [Google Scholar]
- Wankerl M, Wüst S, Otte C. Current developments and controversies: does the serotonin transporter gene-linked polymorphic region (5-HTTLPR) modulate the association between stress and depression? Curr Opin Psychiatry. 2010;23:582–587. doi: 10.1097/YCO.0b013e32833f0e3a. [DOI] [PubMed] [Google Scholar]
- Wylie CJ, Hendricks TJ, Zhang B, Wang L, Lu P, Leahy P, Fox S, Maeno H, Deneris ES. Distinct transcriptomes define rostral and caudal serotonin neurons. J Neurosci. 2010;30:670–684. doi: 10.1523/JNEUROSCI.4656-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamashita A, Singh SK, Kawate T, Jin Y, Gouaux E. Crystal structure of a bacterial homologue of Na+/Cl−-dependent neurotransmitter transporters. Nature. 2005;437:215–223. doi: 10.1038/nature03978. [DOI] [PubMed] [Google Scholar]
- Zhang X, Beaulieu JM, Sotnikova TD, Gainetdinov RR, Caron MG. Tryptophan hydroxylase-2 controls brain serotonin synthesis. Science. 2004;305:217. doi: 10.1126/science.1097540. [DOI] [PubMed] [Google Scholar]
- Zhao Y, Terry DS, Shi L, Quick M, Weinstein H, Blanchard SC, Javitch JA. Substrate-modulated gating dynamics in a Na+-coupled neurotransmitter transporter homologue. Nature. 2011;474:109–113. doi: 10.1038/nature09971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Z, Zhen J, Karpowich NK, Goetz RM, Law CJ, Reith ME, Wang DN. LeuT-desipramine structure reveals how antidepressants block neurotransmitter reuptake. Science. 2007;317:1390–1393. doi: 10.1126/science.1147614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Z, Zhen J, Karpowich NK, Law CJ, Reith ME, Wang DN. Antidepressant specificity of serotonin transporter suggested by three LeuT-SSRI structures. Nat Struct Mol Biol. 2009;16:652–657. doi: 10.1038/nsmb.1602. [DOI] [PMC free article] [PubMed] [Google Scholar]