

Abstract
Object
Antiangiogenic treatments are beginning to give promising outcomes in many vascular diseases including tumor angiogenesis. In this current study the antiangiogenic and pro-apoptotic actions of α1(IV)NC1 and its N- and C- peptides α1S1(IV)NC1, α1S2(IV)NC1 were investigated in-vitro and in-vivo.
Study Method
Endothelial cells (ECs) were treated with α1(IV)NC1, α1S1(IV)NC1, α1S2(IV)NC1 and in-vitro proliferation, migration, tube formation and apoptotic assays were executed. FasL, Fas, Caspase-8, -3 and PARP activations were studied using immunoblotting analysis using specific antibodies. Also the in-vivo antiangiogenic and pro-apoptotic effects were tested using α1(IV)NC1 in a mice model.
Results
Like α1(IV)NC1, its N- and C- terminal α1S2(IV)NC1 and α1S1(IV)NC1 domains posses anti-proliferative, pro-apoptotic activity and inhibit ECs migration and tube formation in-vitro. Both α1S1(IV)NC1 and α1S2(IV)NC1 domains promote apoptosis by activating FasL and down stream apoptotic events including activation of caspase-8, -3 and PARP cleavage in a dose dependent manner in-vitro in ECs. Tumors in mice showed apoptotic TUNEL positive microvasculature upon α1(IV)NC1 treatment, indicating inhibition of tumor angiogenesis and tumor growth. Further, the antitumor activity of α1(IV)NC1 was abrogated when caspase-3 inhibitor was used. These results conform additional properties of α1(IV)NC1 as an endogenous angioinhibitor that induces apoptosis in-vitro and in-vivo by activating FasL mediated caspase-3.
Significance
α1(IV)NC1 and its N- and C- terminal α1S1(IV)NC1 and α1S2(IV)NC1 domains also posses pro-apoptotic and angioinhibitory activity in-vitro and in-vivo. α1(IV)NC1 regulates tumor angiogenesis by activating FasL mediated apoptosis in-vitro and in-vivo. These results demonstrate that α1(IV)NC1 and its peptides inhibit neo-vascular diseases.
Introduction
Angiogenesis, the formation of new blood vessels from preexisting blood vessels, is a very stringently controlled program and normally does not occur, except during development and wound repair processes [1], [2]. This stringent regulation of angiogenesis is manifested by a balance between pro-and anti-angiogenic factors, which keep angiogenesis in check [2]. However, the dynamic equilibrium between pro-angiogenic and anti-angiogenic factors are controlled under many pathological settings, including tumor angiogenesis in cancer progression and other incidents like as age-related macular degeneration, retinopathy of prematurity and diabetic retinopathy resulting in the growth of abnormal new blood vessels [3]–[5].
Vascular basement membranes (VBM) constitute an important component of blood vessels [6]. Makeover of VBM can provide vital pro- and anti-angiogenic molecules to control formation of new blood vessels [7]–[9]. Type IV collagen is a major component of VBM and plays a critical role in new blood vessel development [6]. Proteolytic degradation of type IV collagen in the VBM generates numerous antiangiogenic molecules [7], [10]–[12]. One such antiangiogenic molecule derived from type IV collagen non-collagenous (NC1) domain α1 chain, α1(IV)NC1, has been tested in variety of tumor angiogenesis studies in mice [13]–[15]. However, the molecular and cellular mechanism(s) responsible for inhibition of angiogenesis is not yet clearly understood. The in-vitro and in-vivo studies have demonstrated that α1(IV)NC1 can directly affect endothelial cell migration and impact their proliferation and sprouting [14]. Earlier we have demonstrated that α1(IV)NC1 promotes apoptosis via activation of caspase-3 and PARP cleavage by inhibiting FAK/p38-MAPK/Bcl-2 and Bcl-xL signaling cascade [15]. These results provide a clear understanding about the apoptotic signaling and therapeutic potential of α1(IV)NC1 molecule in neovascular diseases. However, the effects of α1(IV)NC1 and its N- and C-terminal domains α1S1(IV)NC1 and α1S2(IV)NC1 on endothelial cell apoptosis and neo-vascularization have not been previously studied.
In the present study, we demonstrate that α1(IV)NC1 and its N- and C-terminal domains α1S1(IV)NC1 and α1S2(IV)NC1 are potent inhibitors of endothelial cell proliferation, migration and tube formation in-vitro and tumor angiogenesis in-vivo. α1(IV)NC1 promotes apoptosis via activation of caspase-3 and PARP cleavage, presumably by inhibiting FAK/p38-MAPK/Bcl-2 and Bcl-xL signaling cascade [15]. Here in this study, we show that N- and C-terminal domains of α1(IV)NC1 cross talk with FasL and activate FasL and its downstream apoptotic missionary including caspase-8, caspase-3 and PARP cleavage in-vitro. Furthermore, we identified that α1(IV)NC1 promotes apoptosis in tumor vasculature and inhibits angiogenesis and this effect was reversed by a caspase-3 specific inhibitor DEVD in-vivo. These findings contribute significantly towards understanding the apoptotic activation in proliferating ECs and therapeutic potential of endogenous angioinhibitor α1(IV)NC1 and its N- and C-terminal α1S1(IV)NC1 and α1S2(IV)NC1 domains in tumor growth and tumor angiogenesis.
Materials and Methods
Fetal calf serum (FCS), Endothelial basal medium (EBM-2) and Endothelial cell growth medium (EGM-2) were obtained from Fischer Scientific Inc. Penicillin and streptomycin and low melting agarose were purchased from Sigma-Aldrich and cell stains hematoxylin and eosin (H&E) were purchased from Fischer Scientific Inc. Sephadex™-G 100, -G 25 and -G 200 were purchased from GE Healthcare Bio-Sciences AB. BD Matrigel™ Matrix (14.6 mg/ml) was purchased from BD Biosciences Discovery Laboratory. T4-DNA ligase (bacteriophage ligase), different restriction enzymes and polymerases were purchased from New England Biolabs. SCC-PSA1 tumor cells were purchased from ATCC. Caspase inhibitors z-DEVD-fmk was from Enzyme System Products. H&E staining kit form Fisher Scientific Inc. CD31 antibody was purchased from Upstate. Caspase-3, -8, PARP, FasL and Fas antibodies were purchased from Cell Signaling. MTT assay kit was purchased from Chemicon.
Ethics Statement
Boys Town National Research Hospital IACUC committee approved all mice studies.
Tissue culture procedure
SCC-PSA1 cells were maintained in 10% FCS with penicillin/streptomycin (100units/ml each) and maintained at 37°C incubator with 5% CO2. Primary mouse choroidal endothelial cells (EC) were maintained as describe previously [16], [17]. EC were maintained in 40% HAM's F-12, 40% DME-Low Glucose, 20% FCS supplemented with heparin (50 mg/L), endothelial mitogen (50 mg/L), L-glutamine (2.0 mM), penicillin/streptomycin (100units/ml each), Na Pyruvate (2.5 mM), NEAA (1X), 5.0 µg/L of murine INF-γ and cultured on 0.8% gelatin coated plates at 33°C with 5% CO2.
Cloning, Expression and Purification of α1(IV)NC1 and its N- and C- terminal domains α1S1(IV)NC1 and α1S2(IV)NC1
The coding sequence corresponding to the N- and C-terminal non-collagenous domains (NC1) from human Collagen type IV α1 chain was isolated from the placental cDNA using one-step reverse transcriptase-PCR (RT-PCR) (Invitrogen, CA). The N- terminal subunit 330-bp from full length α1(IV)NC1 was amplified using the forward primer: ATACATATGGGCTTCCTTGTGACCAGGCATA and the reverse primer: CACAAGCTTAGGCGCCTCACACACAGCAC and cloned between ‘NdeI’ and ‘XhoI’ sites of pET22b. The C- terminal subunit 330-bp from full length α1(IV)NC1 was amplified using the forward primers: GTGCATATGGTGCACAGCCAGACCAT and the reverse primer: GTGGCAGCAGCCAACTCA and cloned between ‘NdeI’ and ‘XhoI’ sites of pET22b. Amplification and cloning was carryout similarly as reported in our earlier publication [18]. The positive clone was used to transform E. coli strain BL21 for protein expression and purification was performed similarly as reported earlier [18].
Purification of α1(IV)NC1 and α1S1(IV)NC1 and α1S2(IV)NC1 domains
Inclusion bodies of α1(IV)NC1, α1S1(IV)NC1 and α1S2(IV)NC1 were prepared with minor modifications as reported [18]. In addition to the renaturation by stirring method, on-column renaturation was performed for simultaneous renaturation and purification of the α1(IV)NC1, α1S1(IV)NC1 and α1S2(IV)NC1 domains. Denatured α1(IV)NC1, α1S1(IV)NC1 and α1S2(IV)NC1 protein in 800 µl aliquots was loaded onto the Sephadex G-100, Superdex-200 followed by Sephadex G-25 columns similarly as reported [18]. The fractions containing α1(IV)NC1, α1S1(IV)NC1 and α1S2(IV)NC1 were pooled and further concentrated by lyophilization as reported. Endotoxin levels in the final purified α1(IV)NC1, α1S1(IV)NC1 and α1S2(IV)NC1 domains samples were estimated using the Limulus Amebocyte Lysate (LAL) QCL-1000 assay kit (Lonza) according to the manufacturer instructions and also similarly reported in earlier publication [18].
Proliferation assay
A suspension of 7000-cells/well mouse choroidal endothelial cells (ECs) in a 96 well plate was used in proliferation assay. Cells were grown in 96 well plate under 0.5% FBS supplemented with heparin, endothelial mitogen, glutamine and penicillin/streptomycin. After 24-hrs, medium was replaced with ECs medium containing 10% FCS and different concentrations of α1(IV)NC1 or its N- and C-terminal α1S1(IV)NC1 and α1S2(IV)NC1 domains (0.25 and 2.0 µM) and after 48-hrs relative levels of methylene blue incorporation was measured as reported [14], [17].
Migration assay
About 1.0×104 cells/well of ECs were seeded in serum free medium with and without recombinant α1S1(IV)NC1 and α1S2(IV)NC1 (1.0 µM). Medium containing 10 ηg/ml of VEGF was placed into the bottom wells of the Boyden chamber and incubated for about 48-hrs at 37°C with 5% CO2. The numbers of ECs that were migrated and attached to the bottom side of the Boyden chamber membrane were counted as reported earlier [14], [19].
Tube formation assay
Briefly, Matrigel matrix about 250 µl was thawed overnight on ice-cold room and added to each well of a 24-well plate and allowed to solidify for 30-min at 37°C culture incubator. A suspension of about 50×103 ECs in medium without antibiotic was plated on top of the Matrigel matrix. The ECs were than incubated with and without α1(IV)NC1 or its N- and C-terminal α1S1(IV)NC1 and α1S2(IV)NC1 domains (1.0 µM) for 48-hrs at 37°C and viewed using a CK2 Olympus microscope [14], [20], [21].
Cell viability assay
Endothelial cell viability was assessed by MTT assay [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazoliumbromide] following the manufacturer's protocol instructions. About 7.0×103 EC cells/well were plated on a 96-well plate, over night serum starved and stimulated with 10% FCS containing medium. After 24-hrs different concentration of α1(IV)NC1 or its N- and C-terminal α1S1(IV)NC1 and α1S2(IV)NC1 domains (0.25–2.0 µM) were added and incubated for 48-hrs. Apoptosis was monitored by trypan blue exclusion using the cell death detection ELISA kit [22].
FasL and caspase activation assay
About 1.0×106 serum starved EC were collected and suspended in serum free medium. These ECs were pretreated with α1(IV)NC1 N- and C-terminal α1S1(IV)NC1 and α1S2(IV)NC1 domains (1.0 µM) and incubated on 10-cm2 dishes for 6 and 18-hrs. After 6 and 18-hrs, the floating ECs were collected, adherent cells washed once with cold PBS. Adhering and floating cells were lysed on ice in RIPA lysis buffer and centrifuged at 4°C for 30-min at 13000-rpm. About 30 µg of cytosolic extract/lane was separated using 10% SDS-PAGE followed by immunoblotting with anti-FasL, Fas, caspase-8, caspase-3 and PARP antibodies. Immunoreactivity of FasL, Fas, cleaved caspase-8, caspase-3 and PARP proteins were visualized using ECL detection kit as reported [15].
In-vivo tumor studies
About 1.0×106 SCC-PSA1 teratocarcinoma tumor cells were implanted subcutaneously into each mouse at dorso-lateral sides. Six male 129/Sv mice were used in each treatment and control groups. Once the tumors reached around 100-mm3 sizes, 30 µg of α1(IV)NC1 or in combination with 10 µg of caspase-3 specific inhibitor DEVD in 100 µl total volume was intravenously injected to each mouse daily for 15-days. All the experimental mice were scarified and tumors and other organs were collected [21], [31].
Immunohistochemical staining
Tumors from control and α1(IV)NC1 or in combination with DEVD treated mice were embedded in OTC compound and snap-frozen in liquid nitrogen. Tumor sections were incubated for 2-hrs with 5% BSA in PBS to block any nonspecific binding and then incubated with anti-CD-31 antibody for 1-hr followed by incubation with peroxidase-labeled goat anti-rabbit secondary antibody at 37°C. The number of CD-31 positive blood vessels per 10-microscopic fields from four tumor sections per condition was counted [14], [23].
TUNEL assay for apoptosis
To evaluate the relative levels of CD-31 positive vascular or cellular apoptosis, and to quantify cellular apoptosis in tumors from control and α1(IV)NC1 or in combination with DEVD treated mice with and without α1(IV)NC1 treatment, we used ApopTag apoptosis detection kit. Briefly the tumor sections were incubated with 5% BSA in PBS to block any nonspecific binding and stained with ApopTag apoptosis detection reagents following the manufacturer's instructions. To determine whether CD-31 positive blood vessels were undergoing apoptosis, control and α1(IV)NC1 or in combination with DEVD treated tumor sections were co-incubated with both ApopTag reagents and anti-CD-31 antibody. Tumor sections were washed twice and incubated with rhodamine-conjugated goat anti-rabbit secondary antibody for 1-hr at 37°C. The fluorescence staining was analyzed using a fluorescence microscope [14], [23].
Results
Distinct anti-angiogenic activities of α1(IV)NC1 and its N- and C- terminal domains α1S1(IV)NC1 and α1S2(IV)NC1
α1(IV)NC1 is an endogenous metabolite generated from the non-collagenous (NC1) domain of α1 chain of type IV collagen by matrix metalloproteinases-9 (MMP-9) [24], [25]. It was discovered as an angioinhibitory protein with substantial anti-tumor activities [13], [14]. The present study was aimed at understanding the molecular mechanism(s) underlying angioinhibition by α1(IV)NC1 and its N- and C- terminal α1S1(IV)NC1, α1S2(IV)NC1 domains and its implications in prevention of tumor angiogenesis. Our studies tried to identify the angioinhibitory and pro-apoptotic activities of α1(IV)NC1 and its N- and C- terminal α1S1(IV)NC1, α1S2(IV)NC1 domains in mouse choroidal endothelial cells (ECs).
We have performed different angiogenesis experiments to determine the angioinhibitory potential of α1(IV)NC1 and its N- and C-terminal domains α1S1(IV)NC1 and α1S2(IV)NC1 using ECs. Many serum proteins are major pro-angiopromoting factors, elevated levels or imbalance of serum proteins (example VEGF and bFGF) are responsible for the majority of ocular angiogenesis driven by ischemia and also tumor angiogenesis. We first determined serum stimulated angioinhibitory activity of α1(IV)NC1 and its N- and C-terminal domains α1S1(IV)NC1 and α1S2(IV)NC1 by measuring ECs proliferation. The anti-proliferative effect of α1(IV)NC1 and its N- and C-terminal domains were tested in ECs using methylene blue incorporation. Fetal calf serum (10% FCS) stimulated proliferation of ECs were significantly inhibited by α1(IV)NC1 its N-and C-terminal α1S1(IV)NC1 and α1S2(IV)NC1 domains in a dose dependent manner ( Figure 1A ). Interestingly, both N- and C-terminal α1S1(IV)NC1 and α1S2(IV)NC1 domains were showing similar proliferation inhibitory activity when compared to its parent molecule α1(IV)NC1.
Figure 1. Proliferation (A).

Graph summarizes relative levels of methylene blue incorporation in mouse choroidal endothelial cells (ECs) treating with different concentrations of α1(IV)NC1 and its N- and C terminal α1S1(IV)NC1 and α1S2(IV)NC1 domains compared with and without FCS controls after 48-hrs. All groups represent triplicate samples and data in the graphs are represented as mean±SD. Migration (B). Number of ECs with and without α1S1(IV)NC1 and α1S2(IV)NC1 domains migrated towards VEGF on the underside of Boyden chamber membrane were shown and cells viewed using a light microscope after 48-hrs representative fields (100× magnification) shown. Tube formation (C). ECs were plated on Matrigel coated plates in endothelial cell medium as control or with 1.0 µM α1(IV)NC1, α1S1(IV)NC1 and α1S2(IV)NC1 proteins. Tube formation was assessed using a light microscope after 48-hrs, and representative fields at 100× magnification were shown.
Migration of endothelial cells is basically essential during neo-vascularization [14], [26]. Migration of ECs across a PVD membrane towards VEGF in a Boyden chamber was inhibited by 1.0 µM N-and C-terminal α1S1(IV)NC1 and α1S2(IV)NC1 domains ( Figure 1B ). We additionally confirmed the angioinhibitory action of α1(IV)NC1 and its two domains α1S1(IV)NC1 and α1S2(IV)NC1 by a functional assay of ECs tube formation [20]. Tube formation on Matrigel is associated with ECs proliferation, migration and survival [1]. Treatment of ECs with α1(IV)NC1 and its domains α1S1(IV)NC1 and α1S2(IV)NC1 inhibited tube formation equally on Matrigel matrix ( Figure 1C ). Our previous study reported that the angioinhibitory activity of α1(IV)NC1 is mediated through α1β1 integrin [14], [21]. Surprisingly, in this study we noticed that ECs when treated with N- and C-terminal domains and full length α1(IV)NC1 (1.0 µM) were appeared rounded (apoptosis-like) and some of the cells detach from the Matrigel matrix (data not shown). This might be due to activation caspases and apoptosis in ECs treated with α1(IV)NC1 or its N- and C- terminal domains α1S1(IV)NC1 and α1S2(IV)NC1 similarly as reported earlier [15].
Further we also tested the angioinhibitory activity at different doses of α1(IV)NC1 and its N-and C- terminal domains α1S1(IV)NC1 and α1S2(IV)NC1 (0, 0.25, 0.5, 1.0, 1.5, 2.0 µM) by MTT cell viability assay after 10% FCS stimulation in ECs. The results reveal that ECs proliferation was significantly increased by FCS stimulation which was inhibited by α1(IV)NC1 and its N- and C-terminal domains dose dependently after 24-hrs ( Figure 2 ).
Figure 2. Cell viability.
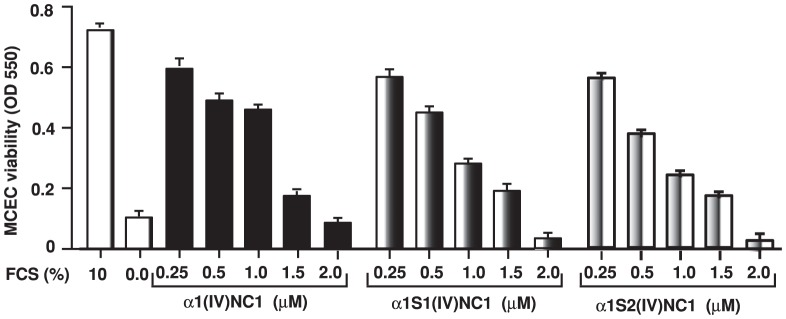
MTT assay was performed to evaluate mouse choroidal endothelial cells (ECs) viability after treatment with different concentrations of α1(IV)NC1 and its N- and C- terminal domains α1S1(IV)NC1 and α1S2(IV)NC1. ECs grown with and without FCS as positive and negative controls. Experiments were performed with three replicates and data in the graphs are represented as mean±SD.
FasL mediated in-vitro apoptotic activity of α1(IV)NC1 and its N- and C- terminal α1S1(IV)NC1 and α1S2(IV)NC1 domains
Caspase-3 is a pivotal molecule mediating cellular apoptosis [27]. Earlier we demonstrated that α1(IV)NC1 induces apoptosis in endothelial cells by activating Caspase-3 [15]. Here in this study, we tested, whether α1(IV)NC1 and its N- and C-terminal domains posses pro-apoptotic activity or not? Interestingly, ECs incubated with α1(IV)NC1 and its N- and C- terminal domains showed dose and time dependent activation of FasL without affecting Fas expression compared to control untreated cells ( Figure 3A & B ). These results suggest both α1S1(IV)NC1 and α1S2(IV)NC1 domains may cross talk with death receptor, activating FasL mediated apoptosis in ECs. These results demonstrate that α1(IV)NC1 and its two domains α1S1(IV)NC1 and α1S2(IV)NC1 promoted apoptosis through FasL in addition to MAPK signaling inhibition [15].
Figure 3. FasL activation (A and B).
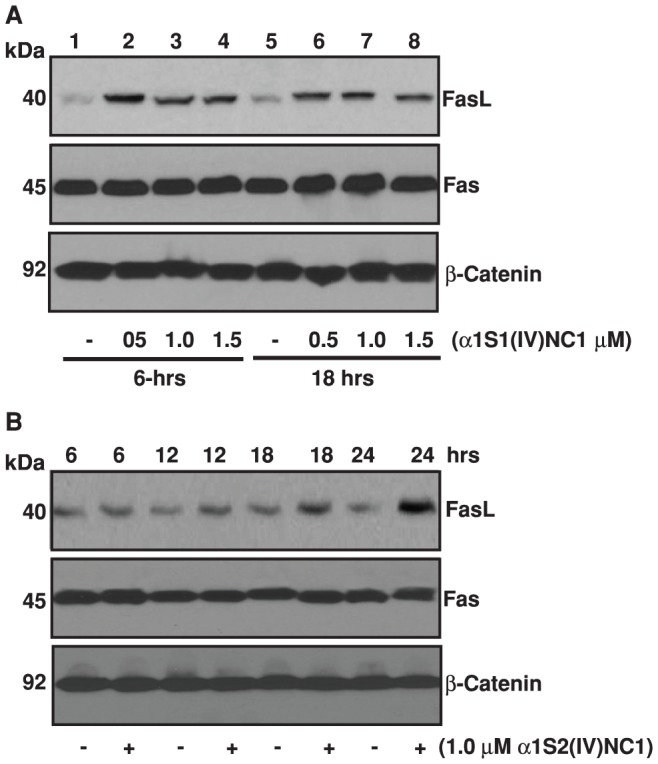
Mouse choroidal endothelial cells were incubated with and without different doses of α1S1(IV)NC1 and α1S2(IV)NC1 domains for 6 and 18-hrs, and total cells were collected, lysed for 30-min in ice-cold RIPA lysis buffer and about 25 µg of total cytosolic extract per lane was separated and immunoblotted with primary antibodies against FasL and Fas. In panel A and B, β-catenin was shown as loading control.
We further assessed whether α1S1(IV)NC1 and α1S2(IV)NC1 induces activation of FasL downstream different caspases such as caspase-8 and caspase-3. We treated ECs with α1S1(IV)NC1 and α1S2(IV)NC1 and observed that activation of caspase-8 ( Figure 4A and B ). These results demonstrate that full length α1(IV)NC1, both its N- and C- terminal domains α1S1(IV)NC1 and α1S2(IV)NC1 induced cellular apoptosis by activating FasL and activating downstream caspase-8. Among the known caspases, caspase-3 is an important effector molecule for most cellular apoptosis [27]. To study whether caspase-3 could be activated by α1S1(IV)NC1 and α1S2(IV)NC1 domains, we treated ECs with different doses of α1S1(IV)NC1 and α1S2(IV)NC1 and observed activation of caspase-3 in a time dependent manner ( Figure 4C and D ). Here, we also identified that activation PARP cleavage which is further down stream to FasL, caspase-8 and caspase-3 in ECs treated with α1S1(IV)NC1 and α1S2(IV)NC1 time and dose dependently ( Figure 5A and B ). These results demonstrate that activation of ECs apoptosis by α1S1(IV)NC1 and α1S2(IV)NC1 domains by activating FasL and its down stream caspases and PARP cleavage.
Figure 4. Caspase-8 activation (A and B).
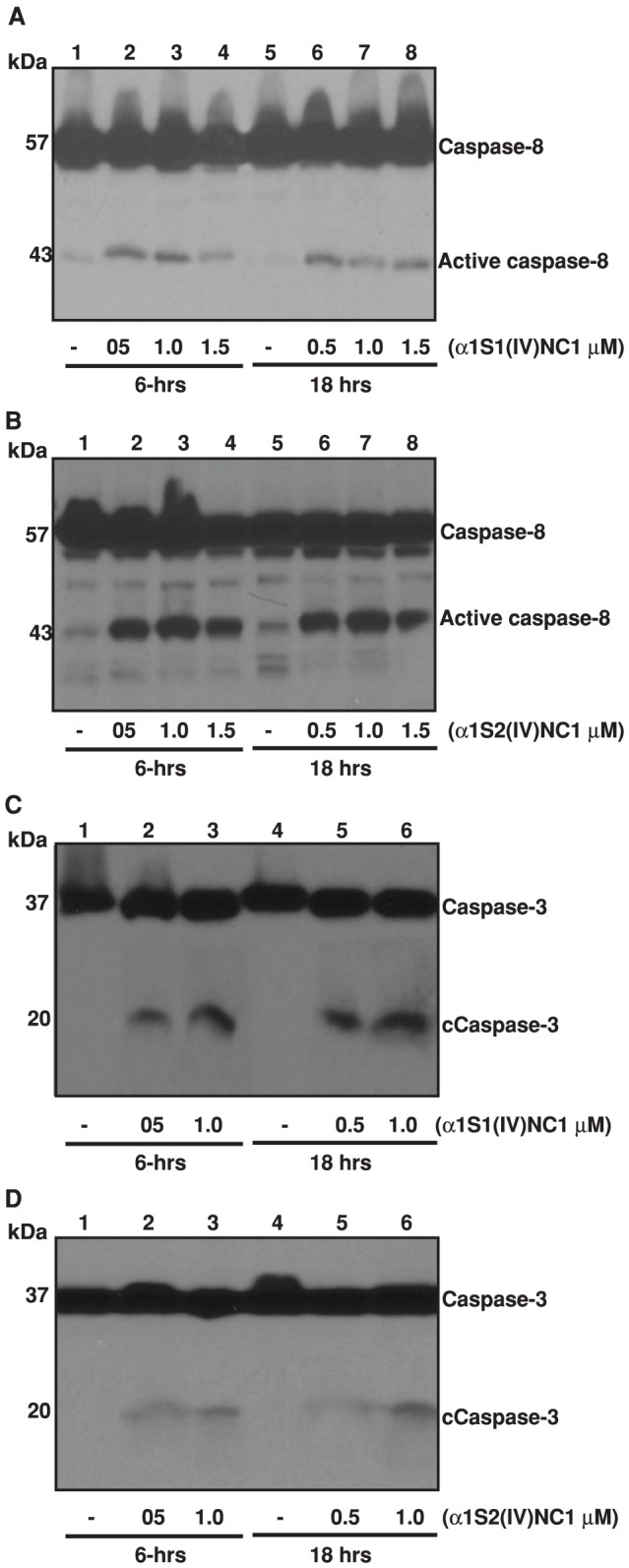
Mouse choroidal endothelial cells (ECs) were incubated with and without different doses of α1S1(IV)NC1 and α1S2(IV)NC1 domains for 6 and 18-hrs, and total cells lysed for 30-min in ice-cold RIPA lysis buffer and about 25 µg of cytosolic extract per lane was separated and immunoblotted with primary antibodies against caspase-8. Caspase-3 activation (C and D). ECs were incubated with and without different doses of α1S1(IV)NC1 and α1S2(IV)NC1 domains and total cytosolic extract immunoblotted with primary antibodies against caspase-3.
Figure 5. PARP activation (A and B).
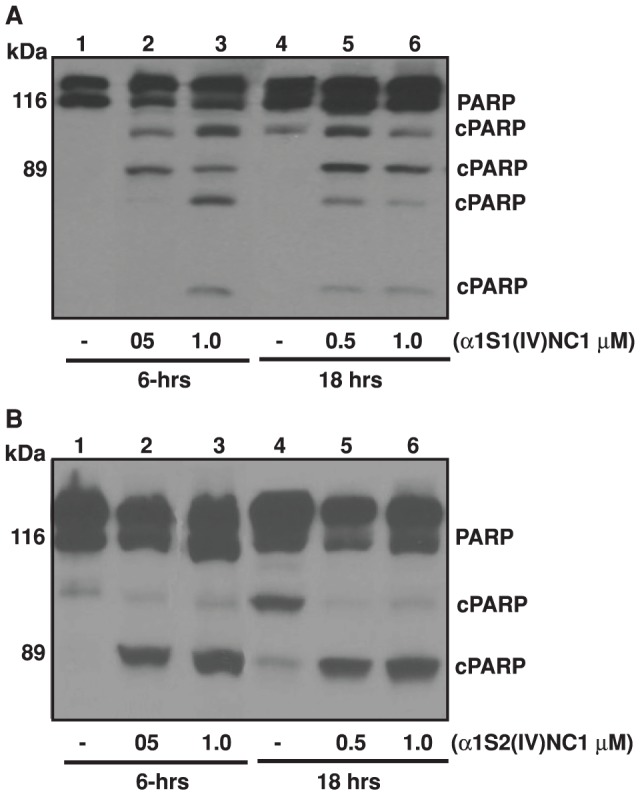
Mouse choroidal endothelial cells (ECs) were incubated with and without different doses of α1S1(IV)NC1 and α1S2(IV)NC1 domains for 6 and 18-hrs, and total cells lysed for 30-min in ice-cold RIPA lysis buffer and about 25 µg of cytosolic extract per lane was separated and immunoblotted with primary antibodies against PARP.
Neutralizing anti-tumorigenic activity of α1(IV)NC1 by caspase-3 inhibitor
To further conform the pro-apoptotic activity of α1(IV)NC1 also partly regulated by caspase-3 activation and apoptosis, we carried in-vitro tumor studies with α1(IV)NC1, or α1(IV)NC1 co-administrated with caspase-3 inhibitor DEVD to tumor bearing mice. Consistent to our earlier findings, administration of α1(IV)NC1 significantly inhibited SCC-PSA1 tumor growth and tumor angiogenesis in-vivo [27]. In contrast, treatment with caspase-3 specific inhibitor DEVD alone showed significant effect on inhibition of tumor growth. The antitumor activity of α1(IV)NC1 was significantly attenuated when α1(IV)NC1 and DEVD were co-administrated to tumor bearing mice (data not shown).
To assess the antitumor effects of α1(IV)NC1 and its ability to activate microvasculature apoptosis, control and α1(IV)NC1 treated mice tumor sections were stained with anti-CD31 antibody ( Fig. 6 , middle panel). In control tumors, less TUNEL positive apoptotic staining was detected whereas elevated levels of apoptosis was observed in tumors that were treated with α1(IV)NC1 ( Fig. 6 ). DEVD treatment alone meagerly affected overall tumor cell apoptosis when compared with α1(IV)NC1. Further, a significant number of ECs in tumor vasculature underwent apoptosis upon α1(IV)NC1 treatment when compared to control tumors, as observed through CD31/TUNEL dual staining ( Fig. 6 , right panel yellow). Interestingly, co-administration of α1(IV)NC1 with DEVD to tumor bearing mice significantly reduced EC apoptosis in microvasculature that was induced by α1(IV)NC1. Further apoptotic tumor cells were observed in α1(IV)NC1 treated tumors using TUNEL staining. This could be the result of an indirect effect since the microvasculature is affected by α1(IV)NC1 treatment, and the tumor cells possibly suffer from oxygen supply and nourishment resulting in apoptosis. This effect was observed at elevated levels in cells undergoing apoptosis, other than CD-31 positive ECs ( Fig. 6 , left and right panels). We also quantified the microvascular density and number of apoptotic vessels through TUNEL staining and by counting the number of CD-31 positive blood vessels (data not shown). Collectively, our results demonstrate that α1(IV)NC1 and its -N and -C terminal domains induced activation of FasL mediated caspase-3 activation and apoptosis in proliferating endothelial cells.
Figure 6. Attenuation of antitumor activity and tumor vasculature apoptosis by caspase-3 inhibitor.

α1(IV)NC1 alone or together with DEVD was injected into SCC-PSA1 tumor bearing mice daily for 15-days. Frozen sections (4.0 µm) of tumors were examined through immunohistochemistry. Apoptotic TUNEL positive cells (green, left), and CD-31 positive tumor blood vessel counts were scored from 10-microscopic fields and four tumors for each experimental condition (red, middle) and co-localization (yellow, right) was shown at 100× magnification. Scale bar corresponds to 50 µm.
Discussion
Many endogenous metabolites generated from type IV collagen were identified as pro-apoptotic, antiangiogenic and anti-tumorigenic in nature [12]–[15], [21], [25], [28]–[32]. The non-collagenous (NC1) domain released by proteolytic activity from type IV collagen α1 chain (α1(IV)NC1) was identified as an antiangiogenic molecule, where as its N- and C terminal domains apoptotic activity is not yet studied. Antiangiogenic activity of α1(IV)NC1 is mediating through α1β1 integrin [14], [21]. However, it is critical to examine α1(IV)NC1 and its both its N- and C- terminal domains α1S1(IV)NC1 and α1S2(IV)NC1 effects in several well defined relevant in-vitro experiments before confirming that these domains are also antiangiogenic. In this study, we tested the antiangiogenic/pro-apoptotic effects of α1(IV)NC1 and its N- and C- terminal domains in different in-vitro and in-vivo experiments.
We demonstrate in this study for the first time, that α1(IV)NC1 and its N- and C- terminal domains inhibits serum induced EC proliferation, migration and tube formation. In addition, we also identified that both α1S1(IV)NC1 and α1S2(IV)NC1 domains activates caspase-8, caspase-3/PARP cleavage through FasL activation in proliferating endothelial cells. This is coherent with the earlier studies revealing that antiangiogenic activity of α1(IV)NC1 is mediated through α1β1 integrin signaling and apoptosis [14], [33]. α1(IV)NC1 and its N- and C- terminal α1S1(IV)NC1 and α1S2(IV)NC1 domains activated FasL mediated apoptosis in ECs. Understanding the mechanism(s) of action of α1S1(IV)NC1 and α1S2(IV)NC domains is crucial for their therapeutic development and use. Thus, α1(IV)NC1 and its N- and C- terminal domains promotes apoptosis in proliferating ECs and inhibits tumor angiogenesis, these endogenous molecules may be an effective therapeutic candidate for treatment of many neovascular diseases. Further evaluation through extensive laboratory studies on these molecules is needed to address the function of these angioinhibitors to be considered for the clinical trials.
Earlier lessons from preclinical trials of angiostatin, endostatin, Thrombospondin-1 (ABT-510) and 2-ME suggest that more basic laboratory studies are required to better understand the mechanism of actions associated with these angioinhibitor molecules. Presently, some of the angioinhibitory agents such as Bevacizumab and VEGFR tyrosine kinase inhibitors; Vatalanib (PTK787/ZK 222584), Semaxanib (SU5416), Sunitinib (SU11248), Sorafenib (BAY 43-9006) are in clinical trials [34], [35]. Many other angioinhibitory drugs [Macugen (pegaptanib sodium), Lucentis (formerly RhuFab V2), tryptophanyl-tRNA synthetase (TrpRS), VEGF-TRAP, AdPEDF, AG-013958, Avastin (bevacizumab), JSM6427 etc] inhibit ocular neovascularization and prevent leakiness of retinal blood vessels by preventing binding of VEGF to its receptors on endothelial cells [15], [36]–[42]. Endogenous metabolite, α2(IV)NC1/α3(IV)NC1 were also reported to laser induced CNV by promoting apoptosis of endothelial cells in-vivo [10].
Our findings suggest that α1(IV)NC1 and its N- and C- terminal domains α1S1(IV)NC1 and α1S2(IV)NC1 may also be effective in inhibition of tumor angiogenesis by activating caspase-3 in new blood vessels. This is further supported by earlier findings that another angioinhibitor, α3(IV)NC1 regulate angiogenesis in a number of in-vitro and in-vivo models [10]. Regression of tumor growth in mice upon α1(IV)NC1 treatment is associated with reduced tumor vasculature and increased TUNEL positive endothelial and tumor cells when compared to control tumors, indicating in-vitro activation of caspase-3. This suggests that α1(IV)NC1 activates caspase-3 and inhibits tumor angiogenesis and tumor growth. Thus, this work not only supports our efforts in development of α1(IV)NC1, its N- and C- terminal domains α1S1(IV)NC1 and α1S2(IV)NC as a potential candidate for tumor angiogenesis neovascular diseases.
Funding Statement
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Folkman J, Haudenschild C (1980) Angiogenesis by capillary endothelial cells in culture. Trans Ophthalmol Soc U K 100: 346–353. [PubMed] [Google Scholar]
- 2. Folkman J (2006) Angiogenesis. Annu Rev Med 57: 1–18. [DOI] [PubMed] [Google Scholar]
- 3. Garner A (1986) Ocular angiogenesis. Int Rev Exp Pathol 28: 249–306. [PubMed] [Google Scholar]
- 4. Jampol LM, Ebroon DA, Goldbaum MH (1994) Peripheral proliferative retinopathies: an update on angiogenesis, etiologies and management. Surv Ophthalmol 38: 519–540. [DOI] [PubMed] [Google Scholar]
- 5. Folkman J (1995) Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat Med 1: 27–31. [DOI] [PubMed] [Google Scholar]
- 6. Paulsson M (1992) Basement membrane proteins: structure, assembly, and cellular interactions. Crit Rev Biochem Mol Biol 27: 93–127. [DOI] [PubMed] [Google Scholar]
- 7. Ingber D, Folkman J (1988) Inhibition of angiogenesis through modulation of collagen metabolism. Lab Invest 59: 44–51. [PubMed] [Google Scholar]
- 8. Carmeliet P, Jain RK (2000) Angiogenesis in cancer and other diseases. Nature 407: 249–257. [DOI] [PubMed] [Google Scholar]
- 9. Maragoudakis ME, Missirlis E, Karakiulakis GD, Sarmonica M, Bastakis M, et al. (1993) Basement membrane biosynthesis as a target for developing inhibitors of angiogenesis with anti-tumor properties. Kidney Int 43: 147–150. [DOI] [PubMed] [Google Scholar]
- 10. Pedchenko V, Zent R, Hudson BG (2004) Alpha(v)beta3 and alpha(v)beta5 integrins bind both the proximal RGD site and non-RGD motifs within noncollagenous (NC1) domain of the alpha3 chain of type IV collagen: implication for the mechanism of endothelia cell adhesion. J Biol Chem 279: 2772–2780. [DOI] [PubMed] [Google Scholar]
- 11. Sudhakar A (2009) The matrix reloaded: new insights from type IV collagen derived endogenous angiogenesis inhibitors and their mechanism of action. J Bioequi Availab 1: 052–062. [Google Scholar]
- 12. Sudhakar A, Boosani CS (2007) Signaling Mechanisms of Endogenous Angiogenesis Inhibitors Derived from Type IV Collagen Gene Regulation and System Biology 1: 217–226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Colorado PC, Torre A, Kamphaus G, Maeshima Y, Hopfer H, et al. (2000) Anti-angiogenic cues from vascular basement membrane collagen. Cancer Res 60: 2520–2526. [PubMed] [Google Scholar]
- 14. Sudhakar A, Nyberg P, Keshamouni VG, Mannam AP, Li J, et al. (2005) Human alphα1 type IV collagen NC1 domain exhibits distinct antiangiogenic activity mediated by alphα1betα1 integrin. J Clin Invest 115: 2801–2810. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 15. Boosani CS, Nalabothula N, Munugalavadla V, Cosgrove D, Keshamoun VG, et al. (2009) FAK and p38-MAP kinase-dependent activation of apoptosis and caspase-3 in retinal endothelial cells by alphα1(IV)NC1. Invest Ophthalmol Vis Sci 50: 4567–4575. [DOI] [PMC free article] [PubMed] [Google Scholar] [Research Misconduct Found]
- 16. Su X, Sorenson CM, Sheibani N (2003) Isolation and characterization of murine retinal endothelial cells. Mol Vis 9: 171–178. [PubMed] [Google Scholar]
- 17. Gunda V, Verma RK, Sudhakar YA (2013) Inhibition of Elastin Peptide-Mediated Angiogenic Signaling Mechanism(s) in Choroidal Endothelial Cells by the α6(IV)NC1 Collagen Fragment. Invest Ophthalmol Vis Sci Accepted, unpublished paper “in Press”. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Gunda V, Boosani CS, Verma RK, Guda C, Sudhakar YA (2012) L-arginine mediated renaturation enhances yield of human, alpha6 Type IV collagen non-collagenous domain from bacterial inclusion bodies. Protein Pept Lett 19: 1112–1121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Boosani CS, Sudhakar A (2006) Cloning, purification, and characterization of a non-collagenous anti-angiogenic protein domain from human alpha1 type IV collagen expressed in Sf9 cells. Protein Expr Purif 49: 211–218. [DOI] [PubMed] [Google Scholar]
- 20. Chow J, Ogunshola O, Fan SY, Li Y, Ment LR, et al. (2001) Astrocyte-derived VEGF mediates survival and tube stabilization of hypoxic brain microvascular endothelial cells in vitro. Brain Res Dev Brain Res 130: 123–132. [DOI] [PubMed] [Google Scholar]
- 21. Boosani CS, Mannam AP, Cosgrove D, Silva R, Hodivala-Dilke KM, et al. (2007) Regulation of COX-2 mediated signaling by alpha3 type IV noncollagenous domain in tumor angiogenesis. Blood 110: 1168–1177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Borza CM, Pozzi A, Borza DB, Pedchenko V, Hellmark T, et al. (2006) Integrin alpha 3beta 1: a novel receptor for alpha3 (IV) noncollagenous domain and a trans-dominant inhibitor for integrin alpha vbeta 3. J Biol Chem [DOI] [PubMed] [Google Scholar]
- 23. Boosani CS, Mannam AP, Cosgrove D, Silva R, Hodivala-Dilke KM, et al. (2007) Regulation of COX-2 mediated signaling by alpha3 type IV noncollagenous domain in tumor angiogenesis. Blood 110: 1168–1177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. McCawley LJ, Matrisian LM (2001) Matrix metalloproteinases: they're not just for matrix anymore!. Curr Opin Cell Biol 13: 534–540. [DOI] [PubMed] [Google Scholar]
- 25. Hamano Y, Zeisberg M, Sugimoto H, Lively JC, Maeshima Y, et al. (2003) Physiological levels of tumstatin, a fragment of collagen IV alpha3 chain, are generated by MMP-9 proteolysis and suppress angiogenesis via alphaV beta3 integrin. Cancer Cell 3: 589–601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Huang Q, Sheibani N (2008) High glucose promotes retinal endothelial cell migration through activation of Src, PI3K/Akt1/eNOS, and ERKs. Am J Physiol Cell Physiol 295: C1647–1657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Nunez G, Benedict MA, Hu Y, Inohara N (1998) Caspases: the proteases of the apoptotic pathway. Oncogene 17: 3237–3245. [DOI] [PubMed] [Google Scholar]
- 28. Maeshima Y, Sudhakar A, Lively JC, Ueki K, Kharbanda S, et al. (2002) Tumstatin, an endothelial cell-specific inhibitor of protein synthesis. Science 295: 140–143. [DOI] [PubMed] [Google Scholar]
- 29. Maeshima Y, Colorado PC, Kalluri R (2000) Two RGD-independent alpha vbeta 3 integrin binding sites on tumstatin regulate distinct anti-tumor properties. J Biol Chem 275: 23745–23750. [DOI] [PubMed] [Google Scholar]
- 30. Sudhakar A, Boosani CS (2008) Inhibition of tumor angiogenesis by tumstatin: insights into signaling mechanisms and implications in cancer regression. Pharm Res 25: 2731–2739. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 31. Sudhakar A, Sugimoto H, Yang C, Lively J, Zeisberg M, et al. (2003) Human tumstatin and human endostatin exhibit distinct antiangiogenic activities mediated by alphaV beta3 and alpha5 beta1 integrins. Proc Natl Acad Sci U S A 100: 4766–4771. [DOI] [PMC free article] [PubMed] [Google Scholar] [Research Misconduct Found]
- 32. Lima ESR, Kachi S, Akiyama H, Shen J, Aslam S, et al. (2006) Recombinant non-collagenous domain of alpha2(IV) collagen causes involution of choroidal neovascularization by inducing apoptosis. J Cell Physiol 208: 161–166. [DOI] [PubMed] [Google Scholar]
- 33. Nyberg P, Xie L, Sugimoto H, Colorado P, Sund M, et al. (2008) Characterization of the anti-angiogenic properties of arresten, an alpha1 beta1 integrin-dependent collagen-derived tumor suppressor. Exp Cell Res 314: 3292–305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Hurwitz H, Fehrenbacher L, Novotny W, Cartwright T, Hainsworth J, et al. (2004) Bevacizumab plus irinotecan, fluorouracil, and leucovorin for metastatic colorectal cancer. N Engl J Med 350: 2335–2342. [DOI] [PubMed] [Google Scholar]
- 35. Morabito A, De Maio E, Di Maio M, Normanno N, Perrone F (2006) Tyrosine kinase inhibitors of vascular endothelial growth factor receptors in clinical trials: current status and future directions. Oncologist 11: 753–764. [DOI] [PubMed] [Google Scholar]
- 36. Deissler HL, Lang GE (2008) [Effect of VEGF165 and the VEGF aptamer pegaptanib (Macugen) on the protein composition of tight junctions in microvascular endothelial cells of the retina]. Klin Monatsbl Augenheilkd 225: 863–867. [DOI] [PubMed] [Google Scholar]
- 37. Mantel I, Zografos L, Ambresin A (2008) Early clinical experience with ranibizumab for occult and minimally classic neovascular membranes in age-related macular degeneration. Ophthalmologica 222: 321–323. [DOI] [PubMed] [Google Scholar]
- 38. Konstantinidis L, Mantel I, Pournaras JA, Zografos L, Ambresin A (2008) Intravitreal ranibizumab (Lucentis(R)) for the treatment of myopic choroidal neovascularization. Graefes Arch Clin Exp Ophthalmol [DOI] [PubMed] [Google Scholar]
- 39. Siemann DW, Shi W (2008) Dual targeting of tumor vasculature: combining Avastin and vascular disrupting agents (CA4P or OXi4503). Anticancer Res 28: 2027–2031. [PMC free article] [PubMed] [Google Scholar]
- 40. Nguyen QD, Shah SM, Hafiz G, Quinlan E, Sung J, et al. (2006) A phase I trial of an IV-administered vascular endothelial growth factor trap for treatment in patients with choroidal neovascularization due to age-related macular degeneration. Ophthalmology 113: 1522 e1521–1522 e1514. [DOI] [PubMed] [Google Scholar]
- 41. Campochiaro PA, Nguyen QD, Shah SM, Klein ML, Holz E, et al. (2006) Adenoviral vector-delivered pigment epithelium-derived factor for neovascular age-related macular degeneration: results of a phase I clinical trial. Hum Gene Ther 17: 167–176. [DOI] [PubMed] [Google Scholar]
- 42. Lu F, Adelman RA (2009) Are intravitreal bevacizumab and ranibizumab effective in a rat model of choroidal neovascularization? Graefes Arch Clin Exp Ophthalmol 247: 171–177. [DOI] [PubMed] [Google Scholar]