

Abstract
The group of nuclei within the basal ganglia of the forebrain is central to the control of movement. We present data showing that the structure and function of the basal ganglia have been conserved throughout vertebrate evolution over some 560 million years. The interaction between the different nuclei within the basal ganglia is conserved as well as the cellular and synaptic properties and transmitters. We consider the role of the conserved basal ganglia circuitry for basic patterns of motor behaviour controlled via brainstem circuits. The output of the basal ganglia consists of tonically active GABAergic neurones, which target brainstem motor centres responsible for different patterns of behaviour, such as eye and locomotor movements, posture, and feeding. A prerequisite for activating or releasing a motor programme is that this GABAergic inhibition is temporarily reduced. This can be achieved through activation of GABAergic projection neurons from striatum, the input level of the basal ganglia, given an appropriate synaptic drive from cortex, thalamus and the dopamine system. The tonic inhibition of the motor centres at rest most likely serves to prevent the different motor programmes from becoming active when not intended. Striatal projection neurones are subdivided into one group with dopamine 1 receptors that provides increased excitability of the direct pathway that can initiate movements, while inhibitory dopamine 2 receptors are expressed on neurones that instead inhibit movements and are part of the ‘indirect loop’ in mammals as well as lamprey. We review the evidence showing that all basic features of the basal ganglia have been conserved throughout vertebrate phylogeny, and discuss these findings in relation to the role of the basal ganglia in selection of behaviour.
 |
Sten Grillner is professor in neuroscience at the Karolinska Institutet. He is known for his work on the intrinsic function of the brainstem–spinal cord networks controlling locomotion in mammals and a lower vertebrate model, the lamprey. Most recently he has been working on the neural mechanisms underlying selection of behaviour using the lamprey as a model, which has had a focus on the role of the basal ganglia and has shown that the organisation of the basal ganglia is conserved throughout the vertebrate phylum. The co-authors, Marcus Stephenson-Jones and Brita Robertson, have contributed very importantly to this work.
Introduction
To generate the basic motor repertoire allowing any vertebrate to move, breath and swallow, or to perform eye movements or fight and flight behaviour, each species has a set of motor programmes that are primed and can be activated at a given point of time (Fig. 1). The programmes underlying these integrated patterns of behaviour are generally conserved from cyclostomes (lamprey) to man, although specific details of the movements may vary. These motor programmes are for the most part generated at the brainstem and spinal cord levels (see Grillner, 2003, 2006). In this review, we will consider the mechanisms that determine when a given motor programme should be recruited and in particular, consider the role of the basal ganglia. We will first review the structure and function of the basal ganglia and focus on the important direct projections from the output nuclei to different brainstem motor centres. We will show that the detailed organisation of the basal ganglia is practically identical in mammals and the lamprey, representing the oldest group of vertebrates, and thus it has been conserved throughout vertebrate phylogeny.
Figure 1. Common motor infrastructure from lamprey to man.
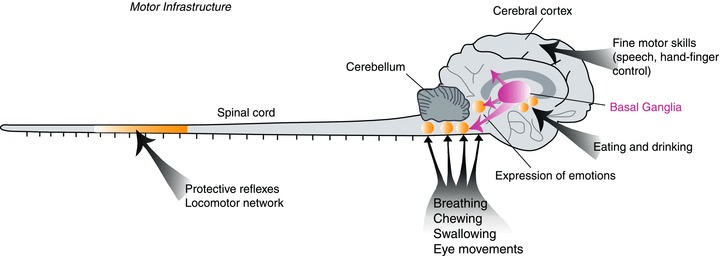
Basic patterns of motor behaviour are controlled by neuronal networks (CPGs) located in the brainstem (e.g. swallowing, breathing) or the spinal cord (e.g. locomotion) (indicated as yellow circles), and the organisation is very similar throughout vertebrate phylogeny. The organisation of the basal ganglia is conserved from lamprey to primates. The basal ganglia control the activity in different brainstem motor centres and play a crucial role in the selection of motor behaviours. In primates and man a well-developed cerebral cortex provides a locus for networks controlling fine motor skills, and it also receives input from the basal ganglia via the thalamus.
The output of the basal ganglia controls brainstem motor programmes
The output level of the basal ganglia (see Fig. 2A) consists of tonically active GABAergic neurones originating from two nuclei, globus pallidus interna (GPi) and substantia nigra pars reticulata (SNr). Subpopulations of GABAergic output neurones from both structures have prominent projections to different motor centres in the brainstem in rodents, cats and primates as well as lower vertebrates (superior colliculus, mesencephalic locomotor region, postural centres, swallowing central pattern generators (CPGs); Fig. 1) (see Swanson 2000). These neurons are tonically active at rest and therefore maintain a continuous inhibitory drive (Grillner et al. 1997, 2005; Hikosaka et al. 2000; Takakusaki et al. 2004; Ménard et al. 2007; Ménard & Grillner, 2008; Takakusaki, 2008, 2010; Stephenson-Jones et al. 2011, 2012a). Triggering a motor programme will depend on removal of this tonic inhibition and therefore the pallidal output neurones in turn need to be inhibited from the input layer of the basal ganglia, the striatum (see Brudzynski & Mogenson, 1985; Mogenson, 1991; Hikosaka et al. 2000; Grillner et al. 2005, 2008; Ménard et al. 2007; Kozlov et al. 2009). One important role of the basal ganglia output in this context is to keep the different motor centres under inhibitory control during resting conditions, so that they only are released when intended.
Figure 2. The organisation of the basal ganglia is almost identical throughout vertebrate phylogeny – from lamprey to primates.
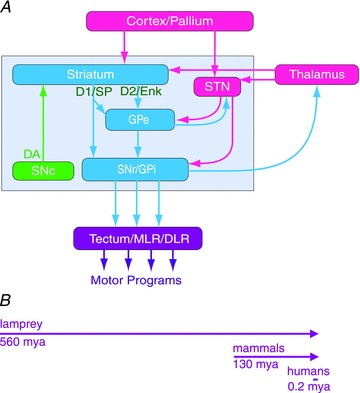
A, the striatum consists of GABAergic neurones (blue), as also globus pallidus externa (GPe), globus pallidus interna (GPi) and substantia nigra pars reticulata (SNr). SNr and GPi represent the output level of the basal ganglia, and it projects via different subpopulations of neurones to tectum (superior colliculus), the mesencephalic (MLR) and diencephalic (DLR) locomotor command regions and other brainstem motor centres, and also back to thalamus and cortex (pallium in lower vertebrates). The indirect loop is represented by the GPe, the subthalamic nucleus (STN) and the output level (SNr/GPi). The striatal neurones of the direct pathway to SNr/GPi express D1R and substance P (D1/SP), while the indirect pathway neurones in striatum express D2R and enkephalin (D2/Enk). Excitatory glutamatergic neurones are in pink. Also indicated is the dopamine supply from the substantia nigra pars compacta (SNc; green). B, the vertebrate lineage is represented. The lamprey had already diverged from the main vertebrate line 560 million years ago (mya); mammals emerged only some 130 mya and humans some 0.2 mya. Yet the design of the basal ganglia is conserved from lamprey to primates. In mammals there is a well-described pallido-thalamo-cortico projection that has not yet been investigated in the lamprey.
In addition to the pallidal control of motor centres, the pallidum also projects back to groups of cells within the thalamus that in turn project back to the cortex and its counterpart pallium in lower vertebrates, which lack a layered neocortex. The pallido-thalamo-cortical loops have received considerable attention in the basal ganglia literature and loops focused on emotions, motor and cognitive functions (Alexander et al. 1986), although the role of these pallido-thalamo-cortical loops is not yet clear. A disinhibition of thalamocortical neurones caused by a cessation of GPi/SNr discharge is thought to increase the level of excitation of the different cortical target areas including motor cortex. In humans, unilateral lesions of the thalamic relay nucleus or of the pallidum are performed to alleviate some of the motor symptoms of Parkinson's disease (e.g. Duval et al. 2006). These lesions decrease the tremor, but leave the Parkinsonian gait and posture unchanged. It is remarkable that no cognitive or emotional symptoms are reported. It would be of considerable interest to further explore the effects of such lesions in order to acquire a better understanding of the role of these pallido-thalamo-cortical loops. Under these conditions, when cortex is out of the loop, all actions of the basal ganglia will be exerted directly over brainstem targets. In this context, it may be important to note that mammals (e.g. cat) devoid of neocortex, but with all other parts of the forebrain intact, display basic goal-directed movements like searching for food and eating (Bjursten et al. 1976), which is most likely dependent on the basal ganglia control of brainstem circuits.
Mechanisms underlying striatal control of action
We next need to consider the intrinsic function of striatum and the intrinsic nuclei of the basal ganglia that ultimately control the output level we have just discussed. The projection neurones in the striatum are of two types (see Fig. 2A), one of which projects directly to subpopulations of neurones at the output level (GPi, SNr) and express dopamine receptors of the D1 type (D1R), which mediate additional excitation. The other type of projection neurones express dopamine D2 receptors (D2R), which instead lower the excitability of this striatal neurone subtype that projects to globus pallidus externa (GPe), and this in turn interacts with the subthalamic nucleus (STN), which projects to the output level. The D1R projection neurones are referred as a direct pathway (to GPi and SNr) and are involved in the initiation of different motor programmes, while the D2R projection neurones are part of the indirect pathway, which inhibit movements and serve as a NO-GO pathway (deLong, 1990; Kravitz et al. 2010). There is also a ‘hyperdirect’ NO-GO pathway from cortex to the subthalamic nucleus that will enhance the inhibitory output. All these components of the basal ganglia are conserved from lamprey to mammals (see below).
Subpopulations of D1R projection neurones can thus be thought of as controlling basic aspects of motor behaviour. They are in turn dependent on excitatory input from thalamus and cortex/pallium, which determine whether they become activated or not. If activated they will take part in the initiation of a given motor programme. The cortical input to striatum has received much attention in the literature, but there is also a direct input from the thalamostriatal neurones (Lacey et al. 2007; Doig et al. 2010), which is almost as large as the cortical input and it will presumably have a similar weight in determining the activity level in striatal neurones.
The rich dopamine innervation of striatum is of fundamental importance (Gerfen & Surmeier, 2011). The tonic level of dopamine release determines the responsiveness of the striatal neurons, and so too little dopamine makes it difficult to initiate movements in Parkinson's disease. Similar motor symptoms occur in all vertebrates investigated from lamprey to man (Thompson et al. 2008). Essentially, too little dopamine will make the D1R direct pathway more difficult to activate, while the D2R NO-GO pathway instead will become more active due to lack of D2R depression. Conversely, when too much dopamine is available, for instance due to medication, the threshold for initiation of movement is lowered to the extent that involuntary movements may arise, hyperkinesias, as opposed to the hypokinesia of Parkinson's disease. Together these observations testify to the paramount importance of the basal ganglia in motor control.
The tonic level of dopamine release is thus a very important feature for normal function of the basal ganglia and the motor system. The dopamine neurones also have another prominent role: to respond with short-lasting bursts of activity during novelty perception or when a reward is anticipated (Redgrave & Gurney, 2006; Schultz, 2007), a feature that can be important in promoting motor learning. Conversely, a short-lasting decline in dopamine activity occurs during aversive behaviour. The control of activity in the dopamine system is thus another critical element in the control of the striatal microcircuits. The neural circuits responsible for the value-based changes in dopamine discharge are not fully understood. The lateral habenula plays an important role, in particular for the phasic decline of dopamine activity during aversive behaviour in both mammals and in the lamprey (Hikosaka, 2010; Shabel et al. 2012; Stephenson-Jones et al. 2012b). These circuits that control the striatum via the lateral habenula and the dopamine system are also exceedingly well conserved (Stephenson-Jones et al. 2012b).
The lamprey basal ganglia
Work in primates, rodents and lamprey have shown that the different components of the basal ganglia in terms of striatum, GPi, SNr, GPe and STN and their connectivity, as well as molecular phenotypes, are all conserved from lamprey to mammals (Reiner et al. 1998; Hikosaka et al. 2000; Ericsson et al. 2011; Stephenson-Jones et al. 2011, 2012a; Ericson, 2012; Robertson et al. 2012). Also in birds the indirect and direct pathways are present, although the GPi and GPe neurones are located within the same nucleus (Reiner et al. 1998).
The lamprey represents the oldest extant species, which diverged from the main vertebrate line around 560 million years ago (Kumar & Hedges, 1998; see Fig. 2B). Common mechanisms within the organisation of the lamprey and mammalian basal ganglia should thus be expected to have been present from the beginning of vertebrate evolution.
During the last few years we have explored the structure and function of the lamprey basal ganglia (Ericsson et al. 2011, 2012; Stephenson-Jones et al. 2011, 2012a; Robertson et al. 2012). At the onset of this work, we knew that the lamprey had a striatum (Pombal et al. 1997a,b), but there were reports in the literature that the pallidum was absent (Murakami et al. 2005). We have since shown that the striatum contains two types of spiny projection neurones that express D1 and D2 receptors and that these neurones have similar membrane properties to those of mammals, characterised by inwardly rectifying potassium channels (Kir), which serve to keep the neurones at a hyperpolarised level during rest, but which are inactivated when the neurones are depolarised by excitatory input (Ericsson et al. 2011; Ericsson, 2012). The D1R spiny neurones also express substance P. They project directly to the lamprey SNr and GPi (Stephenson-Jones et al. 2011, 2012a). These output nuclei both contain tonically active GABAergic neurones, which project to the different brainstem centres. The D2R (enkephalin)-expressing neurones instead project to GPe neurones that in turn project to the glutamatergic lamprey STN, which projects back to the globus pallidus. This means that the intrinsic connectivity within the basal ganglia, the transmitters, synaptic interaction, level of tonic activity, and subtypes of ion channels are conserved from lamprey to primates (Ericsson et al. 2011, 2012; Stephenson-Jones et al. 2011, 2012a; Robertson et al. 2012).
The extrinsic input to the striatum from thalamus and the lateral pallium is conserved even at the level of synaptic dynamics with both providing a glutamatergic excitatory input that activates AMPA and NMDA receptors on striatal neurones. During a stimulus train the EPSPs elicited from the pallium sum progressively at a facilitating synapse; while the initial EPSPs from thalamus are large, the subsequent EPSPs decline in amplitude, so sustained activity generates a short-term depression (Ericsson et al. 2012). The mammalian cortical input to the striatum is also facilitating, whereas it is depressing from thalamus (Ding et al. 2008). This similar arrangement of the synaptic properties between rodents and lamprey has been preserved throughout evolution, which may have functional implications. Furthermore, there is a pallial projection to the subthalamic nucleus (see Fig. 2A), which would correspond to the ‘hyperdirect pathway’ in mammals, a pathway postulated to rapidly terminate an ongoing movement by activating the GABAergic output neurones (GPi/SNr) (Ocaña et al. 2012).
A possible modular structure conserved from lamprey to mammals
It thus appears that practically all details of the basal ganglia circuitry had developed some 560 million years ago (Ericsson et al. 2011, 2012; Stephenson-Jones et al. 2011, 2012a; Ericson, 2012; Robertson et al. 2012) and continued in mammals when they appeared some 130 million years ago (see Fig. 2B). The basal ganglia structure developed most likely to control basic patterns of behaviour, such as initiation of locomotion, steering, eye movements and feeding. Figure 3 illustrates the concept that different modules within the basal ganglia would be responsible for controlling different motor programmes such as locomotion, eye movements and so forth. Each such module would depend on input from pallium/cortex, thalamus and the dopamine system, and it would contain the different components of both the direct (D1) and indirect pathways (D2), while the output cells from GPi and SNr target different motor centres. Subpopulations of pallidal neurons are engaged in the control of different brainstem motor programmes and project to the respective output structures in both lamprey and rodents (Takakusaki, 2008; Stephenson-Jones et al. 2011, 2012a), and most likely there will be a corresponding specificity in the striatum. Whether a given module is selected for action will then depend on the excitatory input from thalamus and pallium/cortex and in addition the degree of tonic dopamine activity.
Figure 3. Conceptual scheme of a modular organisation of the basal ganglia, with one module for each type of motor programme.

Each module would contain the D1R and D2R projection neurones and the components of the direct and indirect pathway GPi, SNr, GPe and STN (see Fig. 2A). Each module would be activated, if sufficient drive occurs from neurones in pallium/cortex and thalamus. The responsiveness of the modules would be determined by the tonic dopaminergic drive. Whereas the lamprey would have a limited behavioural repertoire and few modules, mammals and particularly primates have a very varied and versatile motor repertoire and presumably a greater number of modules.
A conceptual scheme of this type could thus account for the selection of one pattern of behaviour. Some motor patterns can be combined, one can for instance walk and chew at the same time (Kozlov et al. 2009), or walk and turn at the same time; however, one can only turn left or right, not both at the same time. There must thus be a mechanism that gives priority to one behaviour over another. These mechanisms remain to be understood, but it is clear that the basal ganglia play an important role in this context, but turning left or right could also be handled at a downstream level through reciprocal action. It is clear, however, that in humans an intact function of the basal ganglia is required in order to generate a smooth sequence of movements. In Parkinsonian patients this ability is compromised and the patients tend only to be able to perform one motor pattern at a time. In one clinical test, the patient is asked to pick up an object from the floor and then take two steps forward and put it on a shelf (Johnels et al. 2001). A normal person combines these motor patterns in a smooth way, while the patient breaks up the sequence into discrete steps.
Evolutionary considerations: exaptation
Whereas the lamprey has a very limited behavioural repertoire and presumably a limited number of putative modules to control different motor programmes, the situation is different in mammals with an extensive and varied set of motor behaviours. Since all the different components of the basal ganglia are conserved, it would seem likely that during development the number of modules controlling behaviour has increased with each new pattern of behaviour that has evolved. The development would seem to have taken place through a multiplication of basal ganglia modules (Stephenson-Jones et al. 2011) rather than through a development of the intrinsic circuits of the basal ganglia, which had been generally assumed. Such a process of multiplication of existing types of modules to control new functions is referred to as exaptation.
Concluding remarks
In this brief account we have tried to convey some understanding of the basal ganglia and their role in determining which motor programme should be recruited at a given moment of time. We have reviewed the evidence showing that the basic features of the basal ganglia have been conserved throughout vertebrate phylogeny, from lamprey to primates. This indicates that this type of control circuit is of critical importance and illustrates how an understanding of conserved vertebrate physiology can provide insights into the integrated function of the neural circuits controlling behaviour in mammals and other vertebrates.
Acknowledgments
This study was supported by the European Union (FP5 ‘Neurobotics’ 001917, FP7 ‘Lampetra’ 216100, FP7 ‘Select-and-Act’ 201716), the Swedish Research Council (VR-M 3026, VR-NT 621-2007-6049), Karolinska Institutet's Research Funds, and the European Union Cortex Training Program.
Glossary
- CPG
central pattern generator
- D1R
dopamine D1 receptor
- D2R
dopamine D2 receptor
- GPe
globus pallidus externa
- GPi
globus pallidus interna
- SNr
substantia nigra pars reticulata
- STN
subthalamic nucleus
Author contributions
S.G. wrote the manuscript in interaction with all authors, who also approved the final version of the manuscript.
References
- Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Ann Rev Neurosci. 1986;9:357–381. doi: 10.1146/annurev.ne.09.030186.002041. [DOI] [PubMed] [Google Scholar]
- Bjursten LM, Norrsell K, Norrsell U. Behavioural repertory of cats without cerebral cortex from infancy. Exp Brain Res. 1976;25:115–130. doi: 10.1007/BF00234897. [DOI] [PubMed] [Google Scholar]
- Brudzynski SM, Morgenson GJ. Association of the mesencephalic locomotor region with locomotor activity induced by injections of amphetamine into the nucleus accumbens. Brain Res. 1985;334:77–84. doi: 10.1016/0006-8993(85)90569-4. [DOI] [PubMed] [Google Scholar]
- DeLong MR. Primate models of movement disorders of basal ganglia origin. Trends Neurosci. 1990;13:281–285. doi: 10.1016/0166-2236(90)90110-v. [DOI] [PubMed] [Google Scholar]
- Ding J, Peterson JD, Surmeier DJ. Corticostriatal and thalamostriatal synapses have distinctive properties. J Neurosci. 2008;28:6483–6492. doi: 10.1523/JNEUROSCI.0435-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doig NM, Moss J, Bolam JP. Cortical and thalamic innervation of direct and indirect pathway medium-sized spiny neurons in mouse striatum. J Neurosci. 2010;30:14610–14618. doi: 10.1523/JNEUROSCI.1623-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duval C, Panisset M, Strafella AP, Sadikot AF. The impact of ventrolateral thalamotomy on tremor and voluntary motor behaviour in patients with Parkinson's disease. Exp Brain Res. 2006;170:160–171. doi: 10.1007/s00221-005-0198-4. [DOI] [PubMed] [Google Scholar]
- Ericsson J. Cellular and synaptic properties in the lamprey striatum. Stockholm, Sweden: Karolinska Institutet, Dept of Neuroscience; 2012. PhD Thesis. [Google Scholar]
- Ericsson J, Silberberg G, Robertson B, Wikström MA, Grillner S. Striatal cellular properties conserved from lampreys to mammals. J Physiol. 2011;589:2979–2992. doi: 10.1113/jphysiol.2011.209643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ericsson J, Stephenson-Jones M, Kardamakis A, Robertson B, Silberberg G, Grillner S. Evolutionary conserved differences in pallial and thalamic short-term synaptic plasticity in striatum. J Physiol. 2012 doi: 10.1113/jphysiol.2012.236869. (in press; DOI: 10.1113/jphysiol.2012.236869) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerfen CR, Surmeier DJ. Modulation of striatal projection systems by dopamine. Annu Rev Neurosci. 2011;34:441–466. doi: 10.1146/annurev-neuro-061010-113641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grillner S. The motor infrastructure: From ion channels to neuronal networks. Nat Rev Neurosci. 2003;4:573–586. doi: 10.1038/nrn1137. [DOI] [PubMed] [Google Scholar]
- Grillner S. Biological pattern generation: The cellular and computational logic of networks in motion. Neuron. 2006;52:751–766. doi: 10.1016/j.neuron.2006.11.008. [DOI] [PubMed] [Google Scholar]
- Grillner S, Georgopoulos P, Jordan LM. Selection and initiation of motor behavior. In: Stein PSG, Grillner S, Selverston AI, Stuart DG, editors. Neurons, Networks and Motor Behavior. Cambridge, MA, USA: MIT Press; 1997. pp. 2–19. [Google Scholar]
- Grillner S, Hellgren J, Ménard A, Saitoh K, Wikström M. Mechanisms for selection of basic motor programs – roles for the striatum and pallidum. Trends Neurosci. 2005;28:364–370. doi: 10.1016/j.tins.2005.05.004. [DOI] [PubMed] [Google Scholar]
- Grillner S, Wallén P, Saitoh K, Kozlov A, Robertson B. Neural bases of goal-directed locomotion in vertebrates – an overview. Brain Res Rev. 2008;57:2–12. doi: 10.1016/j.brainresrev.2007.06.027. [DOI] [PubMed] [Google Scholar]
- Hikosaka O. The habenula: from stress evasion to value-based decision-making. Nat Rev Neurosci. 2010;11:503–513. doi: 10.1038/nrn2866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hikosaka O, Takikawa Y, Kawagoe R. Role of the basal ganglia in the control of purposive saccadic eye movements. Physiol Rev. 2000;80:953–978. doi: 10.1152/physrev.2000.80.3.953. [DOI] [PubMed] [Google Scholar]
- Johnels B, Ingvarsson PE, Steg G, Olsson T. The posturo-locomotion-manual test. A simple method for the characterization of neurological movement disturbances. Adv Neurol. 2001;87:91–100. [PubMed] [Google Scholar]
- Kozlov A, Huss M, Lansner A, Hellgren Kotaleski J, Grillner S. Simple cellular and network control principles govern complex patterns of motor behavior. Proc Natl Acad Sci U S A. 2009;106:20027–20032. doi: 10.1073/pnas.0906722106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kravitz AV, Freeze BS, Parker PR, Kay K, Thwin MT, Deisseroth K, Kreitzer AC. Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry. Nature. 2010;466:622–626. doi: 10.1038/nature09159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar S, Hedges SB. A molecular timescale for vertebrate evolution. Nature. 1998;392:917–920. doi: 10.1038/31927. [DOI] [PubMed] [Google Scholar]
- Lacey CJ, Bolam JP, Magill PJ. Novel and distinct operational principles of intralaminar thalamic neurons and their striatal projections. J Neurosci. 2007;27:4374–4384. doi: 10.1523/JNEUROSCI.5519-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ménard A, Auclair F, Bourcier-Lucas C, Grillner S, Dubuc R. Descending GABAergic projections to the mesencephalic locomotor region in the lamprey Petromyzon marinus. J Comp Neurol. 2007;501:260–273. doi: 10.1002/cne.21258. [DOI] [PubMed] [Google Scholar]
- Ménard A, Grillner S. Diencephalic locomotor region in the lamprey – afferents and efferent control. J Neurophysiol. 2008;100:1343–1353. doi: 10.1152/jn.01128.2007. [DOI] [PubMed] [Google Scholar]
- Morgenson GJ. The role of mesolimbic dopamine projections to the ventral striatum in response initiation. In: Shimamura M, Grillner S, Edgerton VR, editors. Neurobiological Basis of Human Locomotion. Tokyo: Japan Scientific Society; 1991. pp. 33–44. [Google Scholar]
- Murakami Y, Uchida K, Rijli FM, Kuratani S. Evolution of the brain developmental plan: Insights from agnathans. Dev Biol. 2005;280:249–259. doi: 10.1016/j.ydbio.2005.02.008. [DOI] [PubMed] [Google Scholar]
- Ocaña FM, Saitoh K, Ericsson J, Robertson B, Grillner S. The lamprey pallium controls motion via projections to striatum and brainstem structures similar to those of the mammalian cortex. FENS Abstract. 2012;6 p155.07, 2012. [Google Scholar]
- Pombal MA, El Manira A, Grillner S. Afferents of the lamprey striatum with special reference to the dopaminergic system: a combined tracing and immunohistochemical study. J Comp Neurol. 1997a;386:71–91. [PubMed] [Google Scholar]
- Pombal MA, El Manira A, Grillner S. Organization of the lamprey striatum – transmitters and projections. Brain Res. 1997b;766:249–254. doi: 10.1016/s0006-8993(97)00701-4. [DOI] [PubMed] [Google Scholar]
- Redgrave P, Gurney K. The short-latency dopamine signal: a role in discovering novel actions. Nat Rev Neurosci. 2006;12:967–975. doi: 10.1038/nrn2022. [DOI] [PubMed] [Google Scholar]
- Reiner A, Medina L, Veenman CL. Structural and functional evolution of the basal ganglia in vertebrates. Brain Res Rev. 1998;28:235–285. doi: 10.1016/s0165-0173(98)00016-2. [DOI] [PubMed] [Google Scholar]
- Robertson B, Huerta-Ocampo I, Ericsson J, Stephenson-Jones M, Pérez-Fernández J, Bolam JP, Diaz-Heijtz R, Grillner S. The dopamine D2 receptor gene in lamprey, its expression in the striatum and cellular effects of D2 receptor activation. PLoS One. 2012;7:e35642. doi: 10.1371/journal.pone.0035642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schultz W. Multiple dopamine functions at different time courses. Annu Rev Neurosci. 2007;30:259–288. doi: 10.1146/annurev.neuro.28.061604.135722. [DOI] [PubMed] [Google Scholar]
- Shabel SJ, Proulx CD, Trias A, Murphy RT, Malinow R. Input to the lateral habenula from the basal ganglia is excitatory, aversive, and suppressed by serotonin. Neuron. 2012;74:475–481. doi: 10.1016/j.neuron.2012.02.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stephenson-Jones M, Ericsson J, Robertson B, Grillner S. Evolution of the basal ganglia: dual-output pathways conserved throughout vertebrate phylogeny. J Comp Neurol. 2012a;520:2957–2973. doi: 10.1002/cne.23087. [DOI] [PubMed] [Google Scholar]
- Stephenson-Jones M, Floros O, Robertson B, Grillner S. Evolutionary conservation of the habenular nuclei and their circuitry controlling the dopamine and 5-hydroxytryptophan (5-HT) systems. Proc Natl Acad Sci U S A. 2012b;109:E164–173. doi: 10.1073/pnas.1119348109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stephenson-Jones M, Samuelsson E, Ericsson J, Robertson B, Grillner S. Evolutionary conservation of the basal ganglia as a common vertebrate mechanism for action selection. Curr Biol. 2011;21:1081–1091. doi: 10.1016/j.cub.2011.05.001. [DOI] [PubMed] [Google Scholar]
- Swanson LW. Cerebral hemisphere regulation of motivated behaviour. Brain Res. 2000;886:113–164. doi: 10.1016/s0006-8993(00)02905-x. [DOI] [PubMed] [Google Scholar]
- Takakusaki K. Forebrain control of locomotor behaviors. Brain Res Rev. 2008;57:192–198. doi: 10.1016/j.brainresrev.2007.06.024. [DOI] [PubMed] [Google Scholar]
- Takakusaki K. Auditory pathway in the brainstem contributes to the basal ganglia control of swallowing. J Physiol Sci. 2010;60(supplement):S15. Abstract 87th Meeting of the Physiological Society of Japan. [Google Scholar]
- Takakusaki K, Saitoh K, Harada H, Kashiwayanagi M. Role of basal ganglia–brainstem pathways in the control of motor behaviors. Neurosci Res. 2004;50:137–151. doi: 10.1016/j.neures.2004.06.015. [DOI] [PubMed] [Google Scholar]
- Thompson RH, Ménard A, Pombal M, Grillner S. Forebrain dopamine depletion impairs motor behaviour in lamprey. Eur J Neurosci. 2008;27:1452–1460. doi: 10.1111/j.1460-9568.2008.06125.x. [DOI] [PubMed] [Google Scholar]