

Abstract
Societies of highly social animals feature vast lifespan differences between closely related individuals. Among social insects, the honey bee is the best established model to study how plasticity in lifespan and aging is explained by social factors.
The worker caste of honey bees includes nurse bees, which tend the brood, and forager bees, which collect nectar and pollen. Previous work has shown that brain functions and flight performance senesce more rapidly in foragers than in nurses. However, brain functions can recover, when foragers revert back to nursing tasks. Such patterns of accelerated and reversed functional senescence are linked to changed metabolic resource levels, to alterations in protein abundance and to immune function. Vitellogenin, a yolk protein with adapted functions in hormonal control and cellular defense, may serve as a major regulatory element in a network that controls the different aging dynamics in workers.
Here we describe how the emergence of nurses and foragers can be monitored, and manipulated, including the reversal from typically short-lived foragers into longer-lived nurses. Our representative results show how individuals with similar chronological age differentiate into foragers and nurse bees under experimental conditions. We exemplify how behavioral reversal from foragers back to nurses can be validated. Last, we show how different cellular senescence can be assessed by measuring the accumulation of lipofuscin, a universal biomarker of senescence.
For studying mechanisms that may link social influences and aging plasticity, this protocol provides a standardized tool set to acquire relevant sample material, and to improve data comparability among future studies.
Keywords: Developmental Biology, Issue 78, Insects, Microscopy, Confocal, Aging, Gerontology, Neurobiology, Insect, Invertebrate, Brain, Lipofuscin, Confocal Microscopy
Introduction
The complex colony structures of highly social animals are maintained through the interaction of a reproductive caste, and a helper caste of typically non-reproducing workers with different social task behaviors. In the different workers, specific physiological adaptations enable distinct sib care behaviors, and are also linked to extreme lifespan differences. Honey bees and mole rats represent the best-developed animal models to study how sociality is linked to patterns of accelerated, negligible or reversed aging1-3.
In honey bee colonies, a single egg-laying queen is assisted by thousands of workers that tend the brood, forage for food, and engage in guarding, thermoregulation or hygienic behaviors4. Among these workers are the extremely short-lived foragers, nurse bees with intermediate, and winter (diutinus) bees with longest lifespans. Individuals, however, are not permanently bound to a certain worker-type, but display a flexible behavioral ontogeny: they change from one social task behavior to another ("temporal castes"). Callow bees can change to brood tending nurse bees, which eventually may change to outside foraging. However, callow nest bees can also transform into longest-lived winter bees, and short-lived foragers can even revert into typically longer-lived nurses. Workers with extreme (winter bees) and intermediate (nurse bees) lifespan have well-developed food production and storage organs with copious resources - as opposed to short-lived foragers (reviewed in1,5). However, that the regulation of individual lifespan goes beyond simple changes in an individual's resource balance is suggested by research on a yolk protein, which has diverse adapted functions in the non-reproducing worker caste, such as jelly production6, hormonal control7, immune8 and anti-oxidant defense9.
Patterns of functional decline (senescence) mirror lifespan disparities among workers, as established for olfactory, and also for other brain or motor functions10-13. Specifically, the significant decline in learning function after only two weeks of foraging matches a similar mortality progression in foragers14, as opposed to the lack of detectable decline (negligible senescence) in long-lived winter bees15.
To identify the molecular fingerprints of flexible aging we adapted established experimental paradigms that allow for monitoring and manipulating aging-type transitions8,16,17. Experiment 1 details how to obtain samples in which the effects of chronological age and worker-type specific social behaviors on aging can be separated. Experiment 2 describes the reversal of foragers with accelerated into nurse bees with slowed aging dynamics. Experiment 3 provides an approach for probing effects of cellular senescence by anatomical quantification of an established biomarker for cellular aging (lipofuscin)18.
Protocol
1. Decoupling Senescence from Chronological Age
This section describes the setup of double cohort colonies, which consist of a cohort of identified individuals that share the same chronological age ("single age cohort") and a cohort of nest bees. Same aged individuals of the single age cohort will eventually separate into different worker-types with different aging dynamics - these are nurse bees with slowed and forager bees with accelerated functional decline. All procedures are described for one experimental colony. We advise, however, to perform experiments for at least two colony replicates so that colony effects can be controlled for (two-replicate-design).
Preparing hive boxes for the double cohort colonies: Prepare one regular hive box that receives two food combs with honey, another food comb with pollen, and two empty combs. Make sure to locate a mated queen as well as a donor colony with more than 3,000 nest bees. Both will be introduced later (1.3).
- Obtaining and marking individuals with similar chronological age: Collect combs with sealed brood that is about to emerge. For one replicate expect to collect combs with a total of 3,000-5,000 capped brood cells. For each replicate use a balanced amount of brood from at least three different hive sources to avoid skewed distributions of maternal genotypes (hive origin).
- Place brood combs in an incubator set to 34 °C with 60-70% relative humidity. Make sure to store combs in such a way that emerging brood cannot escape.
- Let bees emerge for two days and mark these bees with a small paint tag on the thorax (e.g. Uni POSCA, Mitsubishi Pencil Co. Ltd.). The paint mark will allow identifying the bees of the single age cohort (day of emergence), and to distinguish them from other replicate colonies.
- Setting up a double cohort colony, which includes the cohort of identified, single age bees: On the day young bees have been marked, collect about 2,500-3,000 nest bees from a donor colony (compare section 1. in Discussion), and add these unmarked bees to the hive box that was prepared before (see step 1.1). The latter individuals will constitute the unidentified nest bee cohort.
- Add the queen, which will be initially confined to a queen cage (commercially available). Seal the cage with edible candy (e.g. Apifonda, Südzucker AG, Mannheim/Ochsenfurt, Germany) to make worker bees slowly release the queen.
- Add the newly emerged and marked bees, which will constitute the single age cohort. These bees are the only marked individuals, and are the focus group for all the following steps.
- Monitoring foraging onset and marking foragers: To assess onset and dynamics of the nurse-to-forager transition in the single age cohort, monitor the demographical development of foraging activity every other day for each colony. Begin counting five days after colonies were set up (Figure 1).
- Count the total number of bees returning from foraging flights (entrance counts) within 3 x 20 min observation periods at fixed times. Make sure to not count bees during periods of orientation flights (see Discussion).
- When entrance counts indicate considerable foraging activity to commence (>100 entrance counts/day), begin marking foragers. To do so, foragers of the single age cohort (single marked individuals) receive a second paint mark upon returning from their first foraging flights. This paint mark will specify the day of foraging onset, and will allow to later identify the foraging age for each forager.
- Repeat daily markings until a sufficient number of bees has been marked. For estimating a sufficient number of marked foragers, expect a retrieval rate of no more than 5-10% after these bees had been aged, typically after 14 days of foraging.
- Sampling: Since all initially marked bees have a similar chronological age, age-matched groups of nurses and old foragers can be collected simultaneously, when foragers have foraged for ≥14 days.
- Single marked nurse bees are collected within the hive, and are identified by nursing behavior (feeding and cleaning of larvae with heads put down in open brood cells).
- Double marked foragers are also collected within the hive before daily foraging activity begins.
- Collect bees in cages (tubes, boxes) that provide sufficient ventilation, and keep dark until further processing. Alternatively, for transcriptomic, epigenetic or proteomic studies, directly snap freeze bees in liquid nitrogen. Collect balanced numbers of individuals from all test groups and replicate colonies.
2. Reversal of Workers with Rapid to Workers with Slowed Aging by Changing the Hive's Demography
This section details how the reversal from workers with accelerated aging (foragers) to workers with slowed aging (nurse bees) is performed. Such behavioral reversal is induced, when foragers experience a lack of nurse bees, which normally engage in brood care. The reversion procedure will separate a single colony replicate into two hives: one hive with the nurse bee fraction ("nurse-derived"), and another one with the forager fraction ("forager-derived"). After successful reversal, possible symptoms of plastic and reversed aging can be studied in the single age cohort with reverted workers, continuing foragers, continuing nurse bees and newly recruited foragers. As before, identified bees of the single age cohort, not the cohort of unidentified nest bees, constitute the experimental focus group.
- Preparation: Replicate hives with nurses (single marked) and foragers (double marked) are made available as described in the previous section. Make sure to not begin reversion with less than 500 marked foragers per replicate colony to ensure sufficient retrieval after the reversion has been completed.
- For safe identification of test groups after reversal it is crucial that the entire forager population in the original hive has been marked off before reversion. The following procedure is described for one replicate.
- The day before reversion, prepare one additional hive box for the forager-derived hive (see step 1.1). Locate two queens and two brood combs from donor hives. Before transfer to the experimental colonies brush away all adult bees from these combs. One caged queen (see step 1.3) and one brood comb will replace queen and brood combs in the original hive box. The other set of queen and brood comb will be used the next day for the new hive box. Similar allocation of new brood combs and alien queens for both, the original and the new hive is advised to make sure that separated foragers and nurses will equally experience changed hive cues ("hive smell").
- Reversion: In the morning, just before the reversion, add the caged queen and the brood comb to the new box that will receive the forager-derived fraction. Wait until the peak foraging time begins. Then move the original colony with marked foragers and nurse bees at least 100 m away from the original location.
- At the original location, set up the new box for the forager-derived hive with brood and queen only.
- Foragers will leave the dislocated original hive box, and head back to the original location. Allow foragers to return to the original location for 2 hr in order to achieve next to complete separation of the forager population from nest bees.
- Then, to terminate separation, close off the original, now "nurse-derived" hive, and transfer it to an apiary at least 3 km away.
- Hive maintenance and monitoring for successful social task reversal: Check the experimental hives regularly for healthy, open brood.
- During the first days after colony manipulation replace unattended and dead brood to reduce potential pathogen load.
- To validate successful reversal within the forager-derived hive, take pictures of brood combs before introducing these, and again when combs are replaced or when the reversion experiment is completed (Figure 2). Areas with previously uncapped brood and with open, live brood are reliable markers of nursing activity in forager-derived colonies.
- Sampling: Physiological effects that accompany social reversal can be detected 3-8 days after foragers and nurses were separated.
- We advise sampling all test groups, i.e. reverted workers and continuing foragers (forager-derived hive), as well as continuing nurses and newly-recruited foragers (nurse-derived hive) 8 days after reversal is initiated.
- Collect samples as described in step 1.5.
3. Analyzing Worker-type Specific Cellular Senescence Patterns by Quantification of Lipofuscin
Lipofuscin is a universal biomarker of cellular senescence. As an intrinsic accumulation product, lipofuscin's specific autofluorescence (emissionmax = 530-650 nm) can be used for detection18.
- Dissection and fixation: Chill bees on ice until motionless; remove and dissect out the desired tissue sample.
- Transfer into fixative (4% paraformaldehyde in phosphate-buffered saline, PBS; pH 7.2) for overnight incubation at 4 °C.
- Wash samples 3 times in PBS.
- Tissue processing and mounting: Cut tissue samples into sections with no more than 40 μm thickness, for example by using a vibrating blade microtome, e.g. Leica VT 1000S (Leica Biosystems, Nussloch, Germany).
- For clearing, incubate tissue sections overnight in 30% glycerol (PBS), and another 2 hr in 50% glycerol (PBS).
- Mount sections on microscopic slides in 50% glycerol (PBS). For long-term storage seal cover slips with nail polish.
- Image acquisition: To detect lipofuscin, we suggest using a laser scanning confocal microscope that provides laser lines with λ = 514, 561nm or similar for excitation, and with the detector bandwidth set to 570-650 nm.
- For better identification of lipofuscin, include a second channel, and do a simultaneous scan at shorter wavelength spectra (excitation = 405 nm; emission = 410-450 nm). The longer wavelength channel will reveal both, granular lipofuscin, but also unspecific "background" due to autofluorescent trachea and other non-granular structures. The second, shorter wavelength channel will only reveal unspecific autofluorescence, but not lipofuscin. Thus, lipofuscin identification can be facilitated by comparing the signals in both channels, with only one of them revealing the granules with lipofuscin specific fluorescence.
- For acquisition of high-resolution images use an objective with 40X magnification or higher, and preferably a numerical aperture of 1.25 or higher. Scan image stacks with dimensions of about 100 x 100 x 10 μm3. Every individual and tissue sample has to be represented by multiple image stacks.
- To reduce intra-individual and inter-individual variation caused by technical variation, always keep laser power and detector sensitivity constant.
- To reduce bias by day-to day technical variations scan equal sample numbers of all test groups in each of the several scanning sessions.
- Image processing: Use software packages with modules that allow for advanced processing of microscopic image stacks, for example ImageJ (U.S. National Institutes of Health, Bethesda, Maryland, USA, http://imagej.nih.gov/ij/).
- Generate a 2D maximum projection for each of the 3D image stacks.
- Apply a Gauss filter with a modest kernel size in order to attenuate high frequency noise, and to preserve structures with dimensions of lipofuscin granules.
- Merge both color channels to facilitate identification of lipofuscin (see step 3.2).
- Image analyses: Make sure that the subject performing the quantification steps will be blind to test group identity.
- For all images, first choose a region of interest (ROI) that covers the relevant structures, and has similar dimensions as the ROIs from other images.
- Then select the desired number of lipofuscin granules that represent each ROI. When selecting lipofuscin granules within a ROI, the application of the following rules will reduce subjective bias.
- Choose a consistent location for selecting the first granule. This can be, for example, the leftmost edge of a ROI, and the granule that is closest to the edge will always be the first selection.
- One after another, granules are chosen that are closest to the previous selection (next neighbor).
- When choosing the next neighbor, move only into one direction, for example only search right from the previous selection. This rule prevents that the selection is dominated by occasional clusters of densely packed granules.
- When selection is completed, assess the size of each lipofuscin particle by outlining and measuring the respective granule area. Use appropriate statistical tests to compare individuals of the different test groups.
Representative Results
Protocol sections 1 and 2 detail how test groups can be obtained to study attributes of accelerated, slowed and reversed aging in colonies with a single age cohort. To monitor worker-type differentiation that accompanies the normal ontogeny we assessed forager counts ("entrance counts") for 6 colonies (Figure 1, compare section 1). The graphs show that considerable change from nurse to the forager state is typically not observed before individuals are more than 10 days old. Marked variability in forager counts was observed with regards to the timing of foraging onset among different colonies, and as a marked day-to-day variation within each colony. Apart from colony specific demographic factors, such as different brood load, much variability is explained by changing weather conditions (time points marked in red in Figure 1). Close monitoring of foraging dynamics therefore is advised to optimize marking and collection efforts during the experiment.
The reversed ontogeny (section 2) from foraging back to nursing tasks can be validated by inspecting brood combs that are introduced into the forager-derived colonies (see steps 2.2 and 2.3). For three replicates Figures 2A, C, and E show brood combs before introduction into forager-derived colonies. Figures 2B, D, and F show the respective combs after removal. Patches of newly capped brood, healthy larvae, and increased pollen storage around brood cells indicate that former foragers now had successfully performed typical nest, including nursing tasks.
Lipofuscin (section 3) is a highly conserved symptom of cellular senescence, and can be readily assessed for post-experimental analyses in the various bee tissues. Figure 3 contrasts lipofuscin accumulation, measured as granule size (Figure 3E), in the hypopharyngeal glands of age matched nurse and forager bees. The difference in chronological age between the two young and the two old groups was ≥17 days, with only one group (foragers) spending these ≥17 days with outside flight and food collection activities. Representative microscopic images (Figures 3A-D) show increased lipofuscin accumulation only for the group of older foragers after more than 17 days of foraging (Figure 3D), not for older nurse bees of similar chronological age (37-43 days; Figure 3B). A two-factorial ANOVA with the fixed main factors worker-type (foragers, nurses) and age difference (Δage ≥17 days) revealed significant effects for worker-type, age difference and the interaction between both factors (Ftype = 33.67, P<0.001; FΔage = 21.93, P<0.001; Ftype x Δage = 22.07, P<0.001). However, post-hoc tests showed significant effects only when contrasting older foragers (≥17 days of foraging) to younger foragers, or to both nurse groups (PF17d vs. F1d/N1d/N17d<0.001, Fisher's LSD; Figure 3E). No difference was detected among the latter three groups, including chronologically young and old nurse groups (all tests with P >0.5, Fisher's LSD; Figure 3E). This suggests that lipofuscin accumulation depends on forager specific activities (foraging age), rather than being function of chronological age only per se.
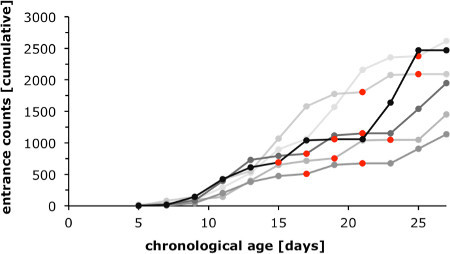 Figure 1. Worker-type differentiation during normal ontogeny. The graph displays entrance counts of foragers returning from foraging flights counted for 6 different colonies beginning 5 days after they were established (for details compare Protocol section 1.4). Considerable transition from nest to foraging activities was first observed when marked individuals of the single age cohort were about 10 days old. Varying slopes for the cumulative entrance counts indicate that the dynamics of the nurse bee to forager transition differ between colonies, and are affected by climatic factors. For example, on days with rain and less than two hours of foraging, the increase in entrance counts typically flattened out (data points in red).
Figure 1. Worker-type differentiation during normal ontogeny. The graph displays entrance counts of foragers returning from foraging flights counted for 6 different colonies beginning 5 days after they were established (for details compare Protocol section 1.4). Considerable transition from nest to foraging activities was first observed when marked individuals of the single age cohort were about 10 days old. Varying slopes for the cumulative entrance counts indicate that the dynamics of the nurse bee to forager transition differ between colonies, and are affected by climatic factors. For example, on days with rain and less than two hours of foraging, the increase in entrance counts typically flattened out (data points in red).
 Figure 2. Validating behavioral reversion. To test if foragers have successfully reverted to nursing tasks, we compared brood combs before they were introduced into forager-derived hives, and after they were removed from these hives. Representative images show brood combs before introduction (A, C, E) and after removal (B, D, F) from three different forager-derived hives, respectively. Brood care by previous forager bees is indicated by an increasing number of cells with capped brood (B, D, F; black arrow, inset in D), sustained survival of larvae in open cells (red arrow) and increased storage of pollen close to brood cells (white arrow). Note that forager-derived-colonies initially are typically less efficient in tending the brood than nurse-derived colonies. This can lead to higher larval mortality in the forager-derived-colonies. Pictures in B, D, F were taken 5, 4 and 7 days after brood combs were introduced into forager-derived colonies.
Figure 2. Validating behavioral reversion. To test if foragers have successfully reverted to nursing tasks, we compared brood combs before they were introduced into forager-derived hives, and after they were removed from these hives. Representative images show brood combs before introduction (A, C, E) and after removal (B, D, F) from three different forager-derived hives, respectively. Brood care by previous forager bees is indicated by an increasing number of cells with capped brood (B, D, F; black arrow, inset in D), sustained survival of larvae in open cells (red arrow) and increased storage of pollen close to brood cells (white arrow). Note that forager-derived-colonies initially are typically less efficient in tending the brood than nurse-derived colonies. This can lead to higher larval mortality in the forager-derived-colonies. Pictures in B, D, F were taken 5, 4 and 7 days after brood combs were introduced into forager-derived colonies.
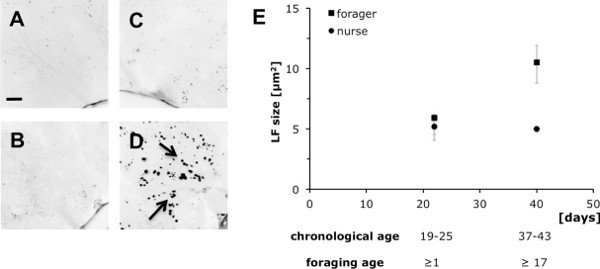 Figure 3. Accumulation of lipofuscin, a biomarker of cellular senescence, can indicate worker-type specific tissue deterioration. Representative microscopic images of hypopharyngeal glands in young (A) and old nurse bees (B), as well as in age-matched forager bees with ≥1 day (C), respectively ≥17 days foraging experience (scale bar in A = 20 μm). Lipofuscin accumulation was measured as granule size, and is given as medians and quartiles for N = 5 individuals for each age and worker-type (E). Foraging for 17 days resulted in significant lipofuscin accumulation, while the same period did not lead to lipofuscin changes in nurse bees (for statistics see Results).
Figure 3. Accumulation of lipofuscin, a biomarker of cellular senescence, can indicate worker-type specific tissue deterioration. Representative microscopic images of hypopharyngeal glands in young (A) and old nurse bees (B), as well as in age-matched forager bees with ≥1 day (C), respectively ≥17 days foraging experience (scale bar in A = 20 μm). Lipofuscin accumulation was measured as granule size, and is given as medians and quartiles for N = 5 individuals for each age and worker-type (E). Foraging for 17 days resulted in significant lipofuscin accumulation, while the same period did not lead to lipofuscin changes in nurse bees (for statistics see Results).
Discussion
We here adopt previously described approaches8,16,17,19,20, and integrate them into a single workflow that will facilitate studying flexible aging in honey bees. Our aim is to provide scientists that are novice to this field with a standardized tool set to obtain relevant sample material, and to improve experimental reproducibility among different research teams. While our procedures are simplified and do not require special equipment as in earlier descriptions (compare for example8), some measures of precaution are advised and are collected below.
Decoupling senescence from chronological age. A most critical aspect is to avoid false identification of forager bees during initial confirmation of foraging behaviors (2nd marking). Therefore, when foragers are to be monitored ("entrance counts") or marked, do strictly avoid daily periods with orientation flights. During these periods many pre-foraging stage bees will depart from or enter the hive. These bees do not display typical physiological characteristics of mature foragers, but build up a spatial map of the hive surroundings by readily identifiable circular flight patterns21.
While most bees change to foraging with the age of 2 weeks and older, sporadic foraging is observed already at very young ages (Figure 1). Extremely precocious foragers typically develop directly from callow nest bees without having passed through the nurse stage. To not include individuals with such an aberrant ontogeny (compare22 and references therein), individuals that begin foraging with the age of 10 days or less are not considered for further analyses.
To further avoid overrepresentation of precocious foragers, we do not make use of classical "single cohort colonies" that consist only of the single age cohort17,23. Instead, when setting up colonies we add random nest bees ("nest bee cohort") to the marked single age cohort (see steps 1.1 and 1.3). Since random nest bees are typically older, they can reduce the pressure on very young bees to develop into extremely precocious foragers17. Such double cohort colonies, therefore, may better resemble a natural hive demography with individuals that slowly progress from nursing to foraging.
When long-term worker specific adaptations are to be studied, collect all test groups outside foraging hours. This is advised to reduce bias by more acute metabolic adjustments due to recent locomotor activities, for example exhausting flight.
Reversal of workers with rapid to slowed aging by changing the hive's demography. After foragers had flown back to the original location it is essential to move away the nurse-derived hive (>3 km). This is to avoid that pre-foraging stage bees are recruited and guided to the old location by other bees, respectively through pheromone communication24.
To further prevent any nurse or other pre-foraging individuals from entering the forager-derived hive, we advise keeping with the following rules: (I) Terminate the separation procedure before daily orientation flights begin. (II) Only attempt reversal on days when strong foraging activity is observed. (III) During and after the initial translocation of the original hive, avoid unnecessary agitation of bees, in particular do not open the hive.
In principle, more artificial setups that confine foragers in a nurse-deprived environment also may lead to reversion. However, such setups only have limited informative value as the forager-derived fraction experiences other stressful environments, thus precluding a direct comparison with control groups from nurse-derived colonies.
Confirming different senescence patterns by quantification of lipofuscin, a biomarker of cellular senescence. Here we exemplified lipofuscin assessment with images and statistical data of hypopharyngeal glands because lipofuscin is most easily detectable in this tissue. This, we believe, is important to help the inexperienced observer setting up the correct protocols for microscopic detection. However, unlike other tissues, hypopharyngeal glands do display significant apoptosis and necrosis during nurse to forager transition25. Such processes may interact with accumulation of the senescence marker, even though we did not detect increased levels of lipofuscin in young foragers that recently had changed from nursing tasks (Figures 3C, E). However, to assess senescence measures in other bee tissues, the microscopy-based methods described here can be easily adapted.
Alternatively, flow-cytometric approaches are less time consuming26. Microscopy-based analyses have the advantage that cellular aging symptoms can be assessed for different regions or even for cells within a single organs27. For studies in brain and other complex organs with spatial heterogeneity in cellular aging28, we therefore recommend the microscopy based approach.
Disclosures
We have nothing to disclose.
Acknowledgments
We thank Osman Kaftanoglu for helpful advice and assistance during filming. We would like to thank the anonymous reviewers for insightful comments. This work was supported by the Research Council of Norway (grants 180504, 191699, and 213976), Marie Curie/FP7 (project ref. 238665), the National Institute on Aging (grant NIA P01 AG22500), and the Pew Charitable Trusts.
References
- Munch D, Amdam GV. The curious case of aging plasticity in honey bees. FEBS Lett. 2010;584:2496–2503. doi: 10.1016/j.febslet.2010.04.007. [DOI] [PubMed] [Google Scholar]
- Buffenstein R. Negligible senescence in the longest living rodent, the naked mole-rat: insights from a successfully aging species. J Comp Physiol B. 2008;178:439–445. doi: 10.1007/s00360-007-0237-5. [DOI] [PubMed] [Google Scholar]
- Parker JD. What are social insects telling us about aging? Myrmecological News. 2010;13:103–110. [Google Scholar]
- Seeley TD. The Wisdom of the Hive. Harvard University Press; 1995. [Google Scholar]
- Amdam GV, Omholt SW. The regulatory anatomy of honeybee lifespan. J Theor Biol. 2002;216:209–228. doi: 10.1006/jtbi.2002.2545. [DOI] [PubMed] [Google Scholar]
- Amdam GV, Norberg K, Hagen A, Omholt SW. Social exploitation of vitellogenin. Proc Natl Acad Sci U S A. 2003;100:1799–1802. doi: 10.1073/pnas.0333979100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guidugli KR, et al. Vitellogenin regulates hormonal dynamics in the worker caste of a eusocial insect. FEBS Lett. 2005;579:4961–4965. doi: 10.1016/j.febslet.2005.07.085. [DOI] [PubMed] [Google Scholar]
- Amdam GV, et al. Social reversal of immunosenescence in honey bee workers. Exp Gerontol. 2005;40:939–947. doi: 10.1016/j.exger.2005.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seehuus SC, Norberg K, Gimsa U, Krekling T, Amdam GV. Reproductive protein protects functionally sterile honey bee workers from oxidative stress. Proc Natl Acad Sci U S A. 2006;103:962–967. doi: 10.1073/pnas.0502681103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheiner R, Amdam GV. Impaired tactile learning is related to social role in honeybees. J Exp Biol. 2009;212:994–1002. doi: 10.1242/jeb.021188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrends A, Scheiner R, Baker N, Amdam GV. Cognitive aging is linked to social role in honey bees (Apis mellifera. Exp Gerontol. 2007;42:1146–1153. doi: 10.1016/j.exger.2007.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Münch D, Baker N, Kreibich CD, Braten AT, Amdam GV. In the laboratory and during free-flight: old honey bees reveal learning and extinction deficits that mirror mammalian functional decline. PLoS One. 2010;5:e13504. doi: 10.1371/journal.pone.0013504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vance JT, Williams JB, Elekonich MM, Roberts SP. The effects of age and behavioral development on honey bee (Apis mellifera) flight performance. J Exp Biol. 2009;212:2604–2611. doi: 10.1242/jeb.028100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dukas R. Mortality rates of honey bees in the wild. Insect Soc. 2008;55 [Google Scholar]
- Behrends A, Scheiner R. Learning at old age: a study on winter bees. Front Behav Neurosci. 2010;4:15. doi: 10.3389/fnbeh.2010.00015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang Z-Y, Robinson GE. Honeybee colony integration: Worker-worker interactions mediate hormonally regulated plasticity in division of labor. Proc Natl Acad Sci USA. 1992;89:11726–11729. doi: 10.1073/pnas.89.24.11726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang ZY, Robinson GE. Regulation of honey bee division of labor by colony age demography. Behavioral Ecology and Sociobiology. 1996;39:147–158. [Google Scholar]
- Double KL, et al. The comparative biology of neuromelanin and lipofuscin in the human brain. Cell Mol Life Sci. 2008;65:1669–1682. doi: 10.1007/s00018-008-7581-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fonseca DB, Brancato CL, Prior AE, Shelton PM, Sheehy MR. Death rates reflect accumulating brain damage in arthropods. Proc Biol Sci. 2005;272:1941–1947. doi: 10.1098/rspb.2005.3192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker N, Wolschin F, Amdam GV. Age-related learning deficits can be reversible in honeybees Apis mellifera. Exp Gerontol. 2012;47:764–772. doi: 10.1016/j.exger.2012.05.011. [DOI] [PubMed] [Google Scholar]
- Capaldi EA, et al. Ontogeny of orientation flight in the honeybee revealed by harmonic radar. Nature. 2000;403:537–540. doi: 10.1038/35000564. [DOI] [PubMed] [Google Scholar]
- Marco Antonio DS, Guidugli-Lazzarini KR, do Nascimento AM, Simoes ZL, Hartfelder K. RNAi-mediated silencing of vitellogenin gene function turns honeybee (Apis mellifera) workers into extremely precocious foragers. Naturwissenschaften. 2008;95:953–961. doi: 10.1007/s00114-008-0413-9. [DOI] [PubMed] [Google Scholar]
- Whitfield CW, Cziko A-M, Robinson GE. Gene expression profiles in the brain predict behavior in individual honey bees. Science. 2003;302:296–299. doi: 10.1126/science.1086807. [DOI] [PubMed] [Google Scholar]
- Schmidt JO. Attraction of reproductive honey bee swarms to artificial nests by Nasonov pheromone. Journal of Chemical Ecology. 1994;20:1053–1056. doi: 10.1007/BF02059741. [DOI] [PubMed] [Google Scholar]
- De Moraes R, Bowen ID. Modes of cell death in the hypopharyngeal gland of the honey bee (Apis mellifera L) Cell Biol Internat. 2000;24:737–743. doi: 10.1006/cbir.2000.0534. [DOI] [PubMed] [Google Scholar]
- Sheehy MR. A flow-cytometric method for quantification of neurolipofuscin and comparison with existing histological and biochemical approaches. Arch Gerontol Geriatr. 2002;34:233–248. doi: 10.1016/s0167-4943(01)00217-5. [DOI] [PubMed] [Google Scholar]
- Hsieh YS, Hsu CY. Honeybee trophocytes and fat cells as target cells for cellular senescence studies. Exp Gerontol. 2011;46:233–240. doi: 10.1016/j.exger.2010.10.007. [DOI] [PubMed] [Google Scholar]
- Wolschin F, Munch D, Amdam GV. Structural and proteomic analyses reveal regional brain differences during honeybee aging. J Exp Biol. 2009;212:4027–4032. doi: 10.1242/jeb.033845. [DOI] [PMC free article] [PubMed] [Google Scholar]