

Abstract
Survivin is over-expressed in most hematologic malignancies but the prognostic significance of the subcompartmental distribution of wild-type or splicing variants in acute myeloid leukemia has not been addressed yet. Using western blotting, we assessed the expression of wild-type survivin and survivin splice variants 2B and Delta-Ex3 in nuclear and cytoplasmic protein extracts in samples taken from 105 patients at the time of their diagnosis of acute myeloid leukemia. Given that survivin is a downstream effector of the PI3K/Akt signaling pathway, survivin expression was also correlated with pSer473-Akt. Wild-type survivin and the 2B splice variant were positive in 76.3% and 78.0% of samples in the nucleus, cytoplasm or both, whereas the Delta-Ex3 isoform was only positive in the nucleus in 37.7% of samples. Cytoplasmic localization of wild-type survivin was significantly associated with the presence of high levels of pSer473-Akt (P<0.001). Inhibition of the PI3K/Akt pathway with wortmannin and Ly294002 caused a significant reduction in the expression of cytoplasmic wild-type survivin. The presence of cytoplasmic wild-type survivin and pSer473-Akt was associated with a lower fraction of quiescent leukemia stem cells (P=0.02). The presence of cytoplasmic wild-type survivin and pSer473-Akt were favorable independent prognostic factors. Moreover, the activation of the PI3K/Akt pathway with expression of cytoplasmic wild-type survivin identified a subgroup of acute myeloid leukemia patients with an excellent outcome (overall survival rate of 60.0±21.9% and relapse-free survival of 63.0±13.5%). Our findings suggest that cytoplasmic wild-type survivin is a critical downstream effector of the PI3K/Akt pathway leading to more chemosensitive cells and a more favorable outcome in acute myeloid leukemia.
Introduction
Survivin is a member of the inhibitor of apoptosis protein (IAP) family and plays an important role in inhibiting cell death by apoptosis.1,2 Survivin shuttles from the nucleus to the cytoplasm after a direct interaction with the nuclear export receptor Crm1, and is essential to cell division by acting as a chromosomal passenger protein and mitotic spindle. Survivin regulates the quality-control checkpoint and ensures the correct completion of cytokinesis with a peak level in G2/M.3 In addition to wild-type (WT) survivin, different splicing variants of survivin have been described such as survivin-2B with 165 amino acids, which has a pro-apoptotic function and is localized in both the nucleus and the cytoplasm, and survivin Delta-Ex3 with 137 amino acids, which has an anti-apoptotic function and is localized exclusively in the nucleus.4 Survivin is over-expressed in most human solid cancers2 and hematologic malignancies, including acute myeloid leukemia (AML).5–9 Some studies have addressed the prognostic impact of the detection of high levels of WT survivin by flow cytometry, western blotting or immunofluorescence in AML, but the results have been discordant.6,9 High levels of the splicing variants 2B and Delta-Ex3, as determined by reverse transcriptase polymerase chain reaction, could have an adverse prognostic impact in AML8. However, considering that survivin is a multifunctional protein compartmentalized in specialized subcellular microenvironments, it is plausible that different survivin isoforms and distinct subcellular pools can affect prognosis differently.2 Very recently it was reported that total survivin levels, assessed by reverse-phase protein array in whole cell lysates, were higher in the CD34+CD38−AML compartment and predicted poor outcome.9 However, it has not yet been investigated whether different subcellular pools of WT survivin and survivin isoforms might influence prognosis in AML.
In addition to the unknown prognostic role of subcellular pools of survivin in AML, the precise mechanisms underlying the up-regulation of this protein have not been clarified yet. In this sense, constitutive activation of the class I-A phospho-inositide3-kinase/serine-threonine kinase (PI3K/Akt) pathway, as the result of autocrine insulin-like growth factor-1 (IGF-1) receptor signaling, occurs in up to 50–80% of AML patients.10,11 This PI3K/Akt up-regulation leads to over-expression of several downstream substrates increasing blast cell proliferation.12–14 It has been reported that granulocyte-macrophage colony-stimulating factor exerts anti-apoptotic and proliferative effects partially by increasing survivin levels through PI3K activation in AML.15 Additionally, it has been shown in a mouse model that internal tandem duplication of Flt-3 regulates survivin expression via PI3K/Akt leading to aberrant progenitor cell proliferation.16 Although the importance of the PI3K/Akt/survivin pathway has been reported in lung and colon cancer,17,18 its incidence and prognostic impact in AML have not been described.
In this study we analyzed the subcellular compartment distribution of WT survivin and the splicing variants survivin-2B and survivin-Delta-Ex3 in AML patients, the correlation with PI3K/Akt pathway activation, and the possible impact on outcome.
Methods
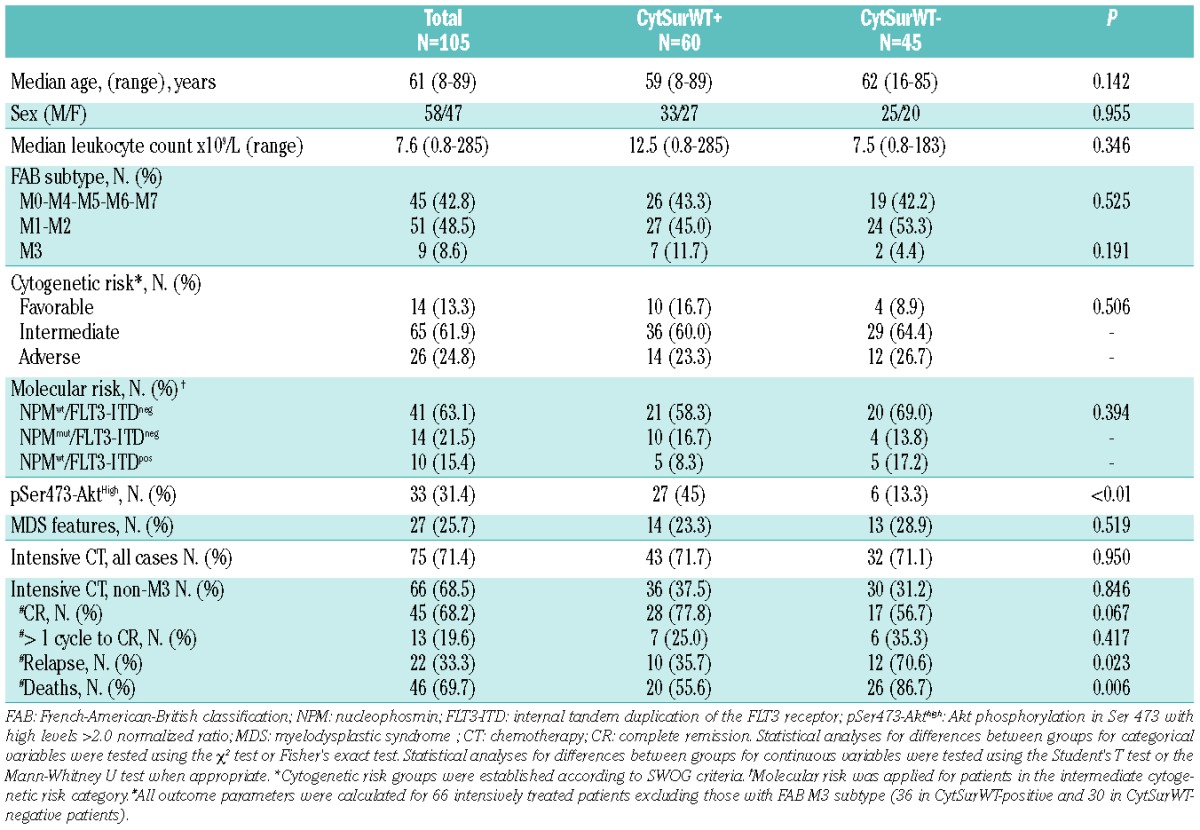
We included 105 consecutive patients diagnosed with AML between February 2006 and October 2010 in our institution. These patients’ clinical features, biological characteristics and outcomes are listed in Table 1. Conventional cytogenetic investigations, fluorescence in situ hybridization and molecular studies for NPM1 mutations and FLT3-ITD were performed as previously described.19 Seventy-five patients suitable for intensive treatment received chemotherapy induction regimens based on PETHEMA Spanish Cooperative group protocols. Samples had been acquired during routine diagnostic assessments and were analyzed in accordance with the regulations and protocols approved by the “Reina Sofia” University Hospital. Informed consent had been obtained in accordance with the Declaration of Helsinki. For in vitro experiments, K562, HL-60 and MV4-11 leukemia cell lines were used (purchased from American Tissue Cell Culture, ATCC).
Table 1.
Characteristics of the 105 AML patients according to cytoplasmic WT survivin status.
Preparation of cytosolic and nuclear protein extracts
Cytosolic, membrane and nuclear proteins were sequentially isolated from mononuclear cells from bone marrow aspirates obtained at diagnosis, using a Q-proteome Cell Compartment kit (Qiagen Iberia SL, Madrid, Spain) according to the manufacture’s instructions.
Western blotting
WT survivin and splicing variants were clearly distinguished by their different molecular weights: 16.5 KDa for the WT protein, 18 KDa for the 2B variant and 14 KDa for the Delta-Ex3 variant. The specificity of each isoform was further confirmed by blocking peptide experiments (Online Supplementary Figure S1). Cytosolic and nuclear specific fractions were tested for purity using anti-β actin (1:1000, Cell Signaling) and lamin-A (H-102, 1:500; Santa Cruz Biotechnology, Santa Cruz, CA, USA) (Online Supplementary Figure S2). Proteins were detected by chemiluminescence using a Chemigenius-2 device and quantified using Gene-Tools 5 analysis software (both from Syngene, Cambridge, UK) using β-actin or laminin as the loading control for cytoplasmic and nuclear extracts, respectively.
Analysis of G0 quiescent and cell cycle
Fresh leukemic cells were first stained with fluorescein isothiocyanate (FITC)- conjugated CD34 phycoerythrin (PE)-conjugated CD38 [Becton Dickinson Immunocytometry Systems (BDIS), San Jose, CA, USA]. To facilitate identification of different phases of cell cycle, leukemic cells (n= 37) were then stained with Hoechst 33342 (Hst; Molecular Probes, Eugene, OR, USA) and pyronin Y (PY; Polysciences, Warrington, PA, USA) as previously described.20,21
Statistical analysis
Normalized values of WT survivin and isoforms were categorized as positive if they were >1.0 (more than the expression in normal bone marrow) or negative if they were ≤ 1 (equal to or less than the expression in normal bone marrow). pSer473-Akt values were considered positive when normalized values were >1.0; normalized values from 1.0 to <2.0 were considered to indicate low expression, whereas normalized values ≥ 2.0 were considered to indicate high expression. Correlations between the expression of survivin isoforms and with pSer473-Akt were assessed with linear regression analysis. χ2 and Fisher’s exact tests were used to compare survivin expression with various clinicopathological parameters. The Mann-Whitney U test was chosen for non-parametric correlation of survivin expression with continuous variables. Response criteria were those reported by Cheson et al.22 Survival measures were estimated using the Kaplan-Meier method. The statistical independence between prognostic variables was evaluated by multivariate analysis using Cox’s proportional hazards model. All tests were two-sided and P values <0.05 were considered statistically significant. This study was approved by the Institutional Review Board of the Instituto Maimonides Investigation Biomedica (IMIBIC), Cordoba.
More details are available in the Online Supplementary Methods.
Results
Subcellular localization of wild-type survivin and isoforms 2B and Delta-Ex3 in acute myeloid leukemia
Using western-blot analysis of compartmental protein extracts, 76.3% of the 105 AML samples were positive for WT survivin and 78.0% were positive for the survivin 2B isoform. Three patterns of subcellular localization of WT survivin were observed: in 19.2% of the samples the protein was confined to the nucleus, in 26.3% it was present only in the cytoplasm and in 30.8% of the cases it was found in both the nucleus and the cytoplasm. Very similar findings were observed for the survivin 2B isoform: in 20% of cases the isoform was restricted to the nuclear protein extracts, in 26% of cases it was found only in the cytoplasm and in 32.0% it was detected in both the nucleus and the cytoplasm. The Delta-Ex3 isoform of survivin was found exclusively in nuclear protein extracts and was present in 37.7% of the 105 samples (Figure 1).
Figure 1.
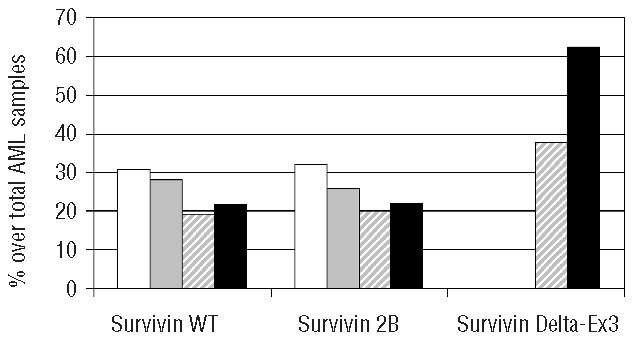
Sub-compartmental localization of WT survivin and isoforms in AML. Y axis represents percentages of cases over total AML samples. White bars: expression in both cytoplasm and nucleus; gray bars: cytoplasmic expression only; gray-shaded bars: nuclear expression only, and black bars: no survivin expression.
Cytoplasmic localization of wild-type survivin is associated with the activated PI3K-Akt pathway
Of the 105 AML samples, 73% were positive for pSer473-Akt with low levels in 41.6% and high-levels (normalized value ≥ 2.0) in 31.4%. We did not find a statistical correlation by linear regression analysis of normalized values of cytoplasmic survivin-2B isoform or nuclear localization of WT survivin, or the 2B or Delta-Ex3 isoforms with the expression of pSer473-Ak. Interestingly, however, linear regression analysis of normalized values obtained from all samples showed a statistical correlation between cytoplasmic wild-type survivin (CytSurWT) and pSer473-Akt (adjusted R2=0.48, P<0.001). In fact, 90% of samples positive for CytSurWT were also positive for pSer473-Akt with high levels in 45%, whereas only 13.3% of samples lacking CytSurWT were positive for high levels of pSer473-Akt. Western blots analyses of sub-cellular distribution of survivin and pSer473-Akt from representative patients are displayed in Figure 2.
Figure 2.
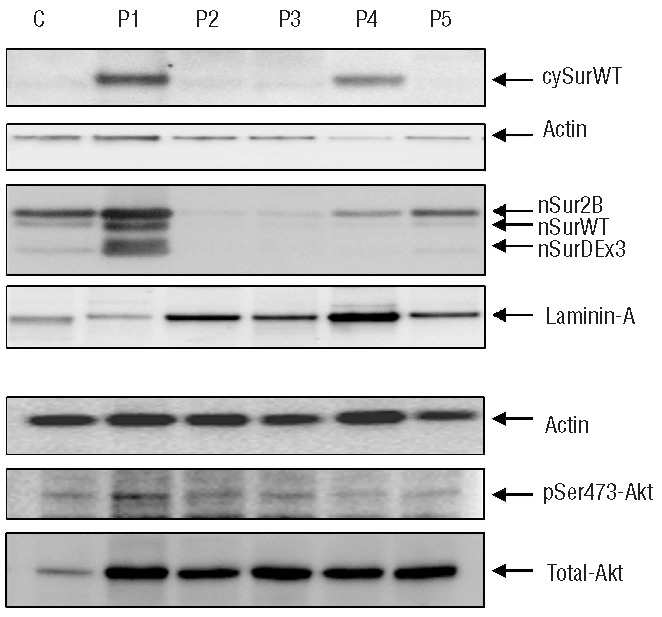
Western blot analysis of total Akt, pSer473-Akt, WT survivin and survivin isoforms in cytoplasmic and nuclear protein extracts of AML samples. Bone marrow aspirates from five representative AML patients at diagnosis (P1 to P5) and a control (C) from a healthy bone marrow donor. Cells were starved and maintained in a serum-free medium without cytokines for 4–5 h before protein extraction. Splicing variants of nuclear survivin are identified by different molecular weights: 2B 18 KDa, and Delta-Ex3 14 KDa and confirmed with blocking peptide experiments. Laminin-A and actin were used as controls for nuclear and cytoplasmic extracts, respectively. CytSurWT: WT cytoplasmic survivin; nSurWT: WT nuclear survivin; nSur2B: nuclear survivin 2B and nSurDEx3: nuclear survivin Delta-Ex3.
Irreversible inhibition of the PI3K/Akt pathway down-regulates the expression of cytoplasmic wild-type survivin and induces apoptosis in acute myeloid cell lines
In order to confirm that CytSurWT is a down-stream effector of the PI3K/Akt pathway we performed inhibition experiments. HL-60, MV4-11 and K562 leukemic cell lines expressed CytSurWT at measurable levels. The PI3K/Akt pathway inhibitor Ly294002 decreased pSer473-Akt levels in the three cell lines but the irreversible action of wortmannin (alone or in combination with Ly294002) caused a more marked down-regulation of CytSurWT expression in the MV4-11 and HL-60 AML cell lines. This was associated with an increase in the apoptotic fraction measured by DioC6. This increase was much greater when the combination of both inhibitors (Ly294002 and wortmannin) was used, increasing from 4.9±0.9% at basal conditions to 13.7±0.81% in the MV4-11 cell line and from 6.1±0.8% to 32±0.7% in the HL-60 cell line (Figure 3). Inhibition of CytSurWT expression by PI3K/Akt blockers was induced even in the presence of cytokines such as stem-cell factor and Flt-3 ligand (Online Supplementary Figure S3). By contrast, inhibition of the mammalian target of rapamycin (mTOR) pathway by rapamycin neither decreased CytSurWT expression nor increased the apoptotic fraction, as shown in Figure 3C and Online Supplementary Figures S4 and S5. Using the Bcr-Abl-positive K562 cell line, inhibition of the PI3K/Akt pathway with Ly294002 and wortmannin did not translate into suppression of CytSurWT suppression.
Figure 3.

Inhibition of the PI3K/Akt pathway using Ly294002 (Ly) and wortmannin (wort) in AML cell lines. Western blot analysis of pSer473-Akt and CytSurWT after culture for 24 h with different inhibitors using MV4-11 (A) and HL-60 (B) cell lines. (C) Mean plus SD of raw volumes of WT survivin of quadruplicate experiments in MV4-11 and HL-60; control, white bars; Ly, gray bars; wortmannin, gray shaded bars; and black bars, Ly+wortmannin. (D) Percentage of DiOC6 loss in the same experiments in MV4-11 (white bars) and HL-60 (gray bars). Ly: Ly294002, Wort: wortmannin, Rap: rapamycin.
Correlation of cytoplasmic wild-type survivin and pSer473-Akt with cell cycle status
The presence of CytSurWT and activation of PI3K/Akt through pSer473-Akt were associated with a significantly lower percentage of quiescent G0 cells in 37 evaluable samples. Thus, in samples negative for both high levels of pSer473-Akt and CytSurWT, the percentage (±SD) of G0 cells was 7.3% (±5.8), while the percentage of G0 cells in samples positive for both was 2.9% (±2.1), (P=0.02, Mann-Whitney U test). The percentage of cells in S/G2/M phase was significantly higher in samples with CytSurWT and high levels of pSer473-Akt (4.5±2.3% versus 3.4±3.2%; P=0.007, Mann-Whitney U test) (Figure 4).
Figure 4.
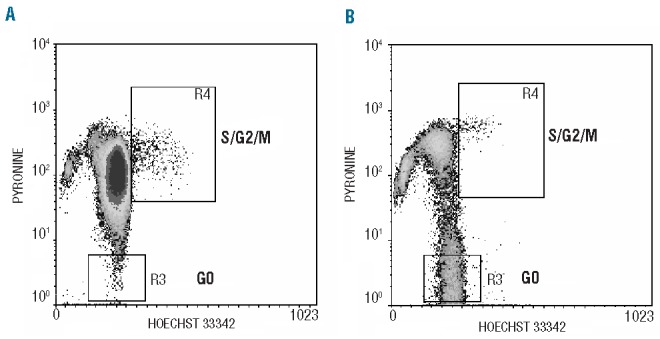
Cell cycle analysis of leukemic cells at diagnosis. In each dot plot, R3 displays G0 cells that are Hoechst 3342 positive but low/negative for the RNA-binding dye pyronin Y and R4 displays S/G2/M cells. Events were gated on forward and side scatter signals (R1) and CD34-CD33/CD38 expression (R2). To set up regions lymphocytes from chronic lymphoid leukemia were used. (A) A representative patient expressing cytoplasmic survivin WT and high levels of pSer473-Akt. (B) A representative patient lacking cytoplasmic WT survivin and negative for pSer473-Akt.
Impact of subcellular localization of wild-type survivin and isoforms on the outcome patients with acute myeloid leukemia
We investigated the impact of subcellular localization of WT suvivin and survivin isoforms in 66 intensively treated AML patients, excluding those with M3 subtype. No statistical differences in overall survival or relapse-free survival were observed when comparing patients with different subcellular localizations of survivin isoform 2B (P=0.097 and P=0.098, respectively). Likewise, we did not observe statistical differences comparing outcomes of patients with or without nuclear Delta-Ex3 expression (P=0.08 and P=0.148 for overall and relapse-free survival, respectively) (Online Supplementary Figure S6). Interestingly, overall and relapse-free survival were significantly better in those patients with cytoplasmic localization of WT survivin (P=0.048 and P=0.02, respectively). Kaplan-Meier curves are shown in Figure 5. For patients positive for CytSurWT, there were not statistical differences in overall or relapse-free survival comparing those positive or negative for nuclear WT survivin expression.
Figure 5.
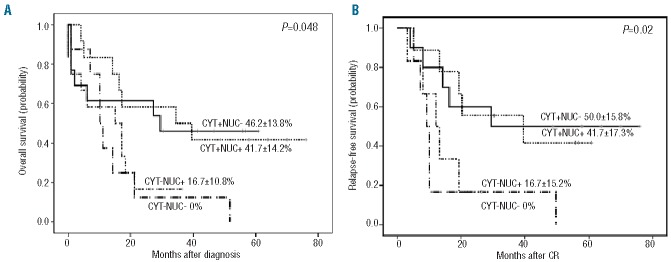
Outcomes of AML patients according to subcellular WT survivin localization. Overall survival was calculated with Kaplan-Meier curves for all 66 intensively treated patients and relapse-free survival was calculated for intensively treated patients achieving complete remission. (A) Kaplan-Meier curves for overall survival according to subcompartmental WT survivin expression. (B) Kaplan-Meier curves for relapse-free survival according subcompartmental WT survivin expression.
With a median follow up of 39 months (range, 18.2–76.0), overall survival probabilities were 39.6±8.9% (median 32.5 months; 95% CI: 12.4–52.5) and 0% (median 14.2 months; 95% CI: 9.2–19.2) for CytSurWT positive and negative patients, respectively (P=0.005), log-rank test). Likewise, the relapse-free survival probability was significantly superior in CytSurWT-positive patients (45.02±10.3 versus 0%, P=0.010).
The expression of CytSurWT, with or without simultaneous nuclear expression, did, therefore, identify a subgroup of intensively treated AML patients with significantly better outcomes. The median values of the Cyt/Nuc ratio (normalized values of cytoplasmic WT survivin/normalized values of nuclear WT survivin) was 0.68 (range, 0 to 6.5). When analyzing outcomes, patients with ratios more than 0.68 had a better relapse-free survival, although the difference was not statistically significant (P=0.054), and no difference in overall survival (P=0.242)
The clinical characteristics of positive (n=60) and negative CytSurWT (n=45) AML patients are summarized in Table 1. The expression of CytSurWT was not statistically associated with clinical features at presentation, such as white blood cell count, age, percentage of blast cells in the bone marrow, myelodysplastic changes, cytogenetic risk groups or molecular profile. CytSurWT was positive in seven out of nine M3-AML cases. Most (90%) CytSurWT positive cases were positive for pSer473-Akt with high levels in 45% of them, in sharp contrast to the CytSurWT-negative cases (P<0.01).
Impact of the PI3K/Akt activation pathway and cytoplasmic wild-type survivin on outcome of patients with acute myeloid leukemia
We analyzed the impact of low and high pSer473-Akt expression on outcomes and found that high levels (normalized values >2.0) were associated with significantly better overall and relapse-free survival. Thus, overall survival probabilities were 57.9±13.1% (median 52.1 months, 95% CI: 37.9.–66.2) for pSer473-Akt-positive patients whereas it was 0% and 17.1±7.4% for negative ones (normalized valued <1.0) and ones with low expression (normalized values >1.0 and < 2.0), respectively (P<0.01). Likewise, relapse-free survival was 56.4±11.9% (median 48.7 months, 95% CI: 34.2–63.3) for pSer473-Akt-positive patients whereas it was 0% and 28.1±12.2% for those with negative and low expression, respectively (P=0.038) (Figure 6A,B).
Figure 6.
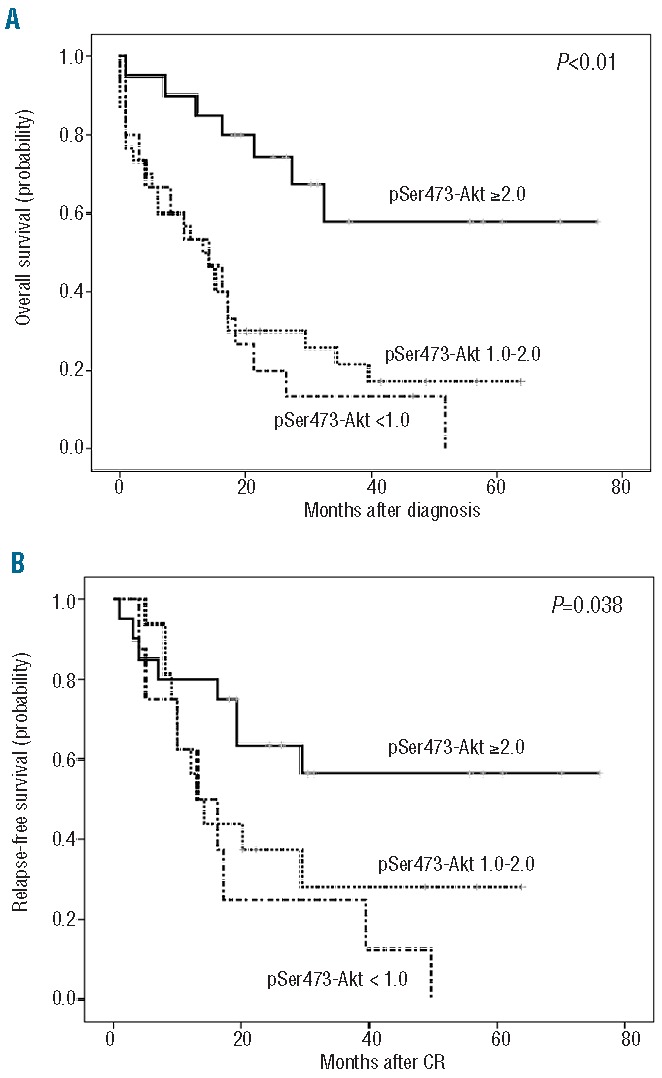
Outcomes of AML patients according to normalized values of pSer473-Akt. Overall survival was calculated with Kaplan-Meier curves for all 66 intensively treated patients and relapse-free survival was calculated for intensively treated patients achieving complete remission. (A) Kaplan-Meier curves for overall survival according to pSer473-Akt normalized values. (B) Kaplan-Meier curves for relapse-free survival according to pSer473-Akt normalized values.
Altogether, when we defined four PI3K/Akt/CytSurWT activation pathway groups, we found that patients positive for CytSurWT and with high levels of pSer473-Akt had an excellent prognosis with overall and relapse-free probabilities of 60.1±21.9% and 63.0±13.5%, respectively, compared to those lacking an activated PI3K/Akt/CytSurWT pathway, whose prognosis was dismal (5-year overall and relapse-free survival, 0%) (Figure 7A,B).
Figure 7.
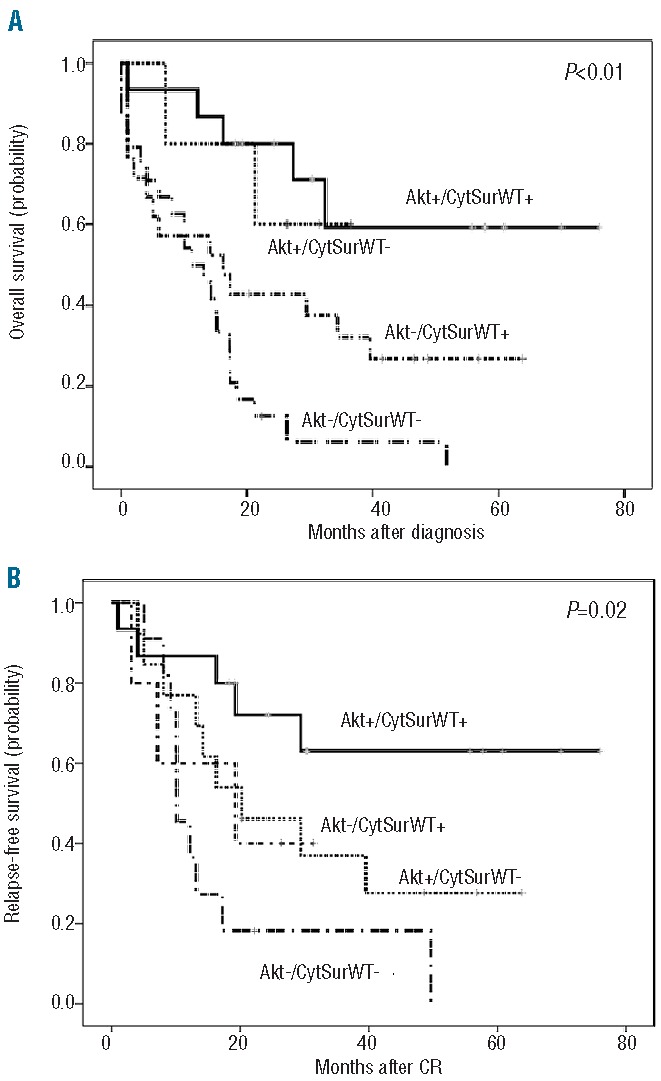
Outcomes of AML patients according to the PI3K/Akt/CytSurWT activation pathway. Overall survival was calculated with Kaplan-Meier curves for all 66 intensively treated patients and relapse-free survival was calculated for intensively treated patients achieving complete remission. (A) Kaplan-Meier curves for overall survival according to PI3K/Akt/CytSurWT activation pathway status. (B) Kaplan-Meier curves for relapse-free survival according to PI3K/Akt/CytSurWT activation pathway status.
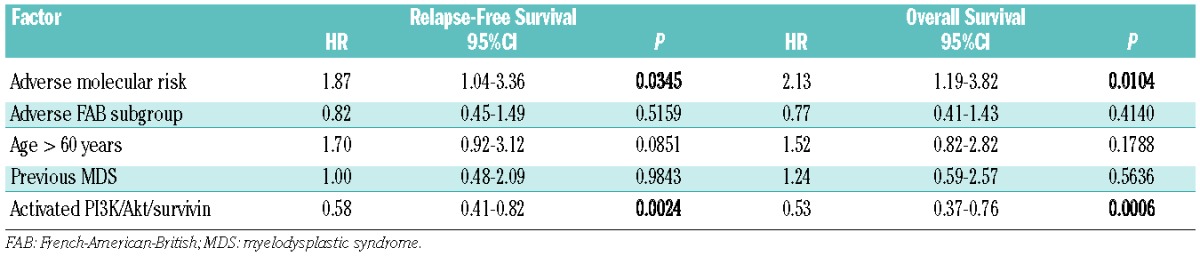
Additionally, age <60 years and low-cytogenetic/molecular risk group were favorable prognosis factors in univariate analyses. In the multivariate Cox regression analysis only the presence of an activated PI3K/Akt/CytSurWT pathway remained statistically significant for favorable outcomes in terms of overall and relapse-free survival, whereas poor molecular risk (FLT3-ITD-positive) remained statistically significant for inferior outcomes (Table 2). Cox multivariate analysis for overall survival (in the whole series and the 66 intensively treated patients) and relapse-free survival (66 intensively treated patients) including categorical variables obtained for nuclear/cytoplasmic localization of survivin isoforms was also performed and no statistical significance was observed for the isoforms 2B and Del-Ex3.
Table 2.
Results of multivariate Cox regression analysis of overall survival and relapse-free survival risk of 66 intensively treated non-M3 AML patients.
Discussion
The findings of this study can be summarized in three main areas. First, we described the distribution of subcellular pools of WT survivin and the isoforms 2B and Delta-Ex3 in AML. Second, we demonstrated that CytSurWT is a downstream effector of activated PI3-K/Akt, independent of mTOR, in AML. Finally, and most importantly, we identified the activated PI3K/Akt/CytSurWT pathway as an independent predictor of a better outcome in patients diagnosed with AML.
Survivin has been consistently described to be overex-pressed in most human malignancies, independently of mitotic index,2,23 and identified as a risk factor for poor prognosis. However, considering that survivin is a multi-functional protein compartmentalized in specialized sub-cellular microenvironments, it is plausible that different survivin isoforms and distinct subcellular pools can affect prognosis differently.1,2.24 In fact, some studies in solid tumors have demonstrated that nuclear survivin expression assessed by immunohistochemistry could be considered a favorable prognostic marker.25 In hematologic malignancies, a differential compartmental localization of survivin has been assessed in mantle cell lymphoma26 and lymphoid leukemia (acute and chronic)27 by western blot and immunohistochemical studies. However, to the best of our knowledge only two studies have analyzed the global cellular expression of WT survivin and isoforms by reverse transcriptase polymerase chain reaction analysis in AML8,28 and this is, therefore, the first description of the distribution of WT survivin and isoforms in nuclear and cytoplasmic compartments. Our data indicate a substantial proportion of AML samples overexpressed WT survivin and the 2B isoform in the nuclear and/or cytoplasmic compartments, whereas the Delta-Ex3 isoform is confined to the nucleus and was expressed in only one third of cases. There are inconsistencies in the literature regarding the prognostic significance of nuclear or cytoplasmic survivin expression in solid tumors with the potential pitfall of the misinterpretation or inappropriate processing of samples for immunohistochemistry. In fact, western blotting of subcellular protein extracts is recommended as the most reliable method to confirm the expression of these proteins.24 Our results using this well-proven method are, therefore, a step forward in the understanding of the sub-cellular distribution of WT survivin and isoforms, enabling assessment of the potential prognostic impact in AML. Additional experiments are warranted to determine whether the subcompartmental distribution for each patient is stable due to mutational changes or whether there could still be shuttling between the nucleus and cytoplasm. It is likely that accumulation of cytoplasmic survivin via PI3k/AKT activation occurs in a phosphorylation-dependent manner, acting directly on its nuclear exclusion upon activation of nuclear export signals such as Thr-34 phosphorylation or increasing survivin stability at mitosis by CDK1 phosphorylation.
Mechanisms leading to survivin over-expression in cancer are not completely understood and several oncogenic pathways have been identified which independently turn on survivin gene expression, including TCF4/β-catenin and Akt. Contrariwise, many tumor suppressor networks such as p53, FHIT and APC have also been shown to silence transcription of the survivin gene.1,2 In our study, the presence of CytSurWT was correlated significantly with the levels of pSer473-Akt and the inhibition of PI3K blocked CytSurWT expression even in the presence of exogenous cytokines. By contrast, mTOR pathway inhibition by rapamycin did not modify CytSurWT expression. Importantly, a body of evidence suggests that constitutive activation of mTORC1 in AML is independent of PI3K which in turn can be constitutively fully activated in an mTOR-independent manner.29 Previous studies have demonstrated that PI3K/Akt inhibitors caused decreased expression of many proteins including survivin17,18,30–32 in solid tumors and hematologic malignancies. Additionally, it has been shown in a mouse model that internal tandem duplication of Flt3 regulates survivin expression via PI3K/Akt leading to aberrant progenitor cell proliferation and promoting leukemogenesis.16 Our results demonstrate a clear correlation of constitutive activation of PI3k/Akt/CytSurWT pathway in a significant proportion of AML patients. In strong agreement with our findings, Carter et al., who analyzed survivin and 206 other proteins, recently reported that survivin levels are significantly correlated with the levels of proteins in the PI3K signaling pathway9 but not with the MAPK signaling pathway. The precise network by which Akt recruits CytSurWT is not completely understood but it has been demonstrated that p70S6K1 activation and overexpression of p70S6K1 alone was sufficient to induce survivin expression in embryonic fibroblast cells.33
In our study the activation of PI3K/Akt/CytSurWT in AML samples was associated with a more proliferative cell fraction and low proportion of G0 quiescent cells. In accordance with this biological finding, our results demonstrate that activation of this pathway identifies a subgroup of patients with a significantly more favorable outcome. Importantly, the presence of more proliferative AML blast cells translates into greater susceptibility to chemotherapy. In this sense, constitutive activation of the PI3K/Akt pathway was initially thought to represent an unfavorable marker in AML,12 but Tamburini et al.13 subsequently demonstrated that this constitutive activation of PI3K pathway leads leukemic cells to S-phase being more susceptible to conventional cell-cycle chemotherapy drugs. Our findings confirm those of Tamburini et al. and, more importantly, identify CytSurWT as a critical downstream target of the PI3K/Akt pathway that further refines the prognosis. In fact, patients with PI3K/Akt/CytSurWT activated pathway have an excellent prognosis compared to those lacking PI3K/Akt and cytoplasmic WT survivin with dismal overall and relapse-free survival. This finding is in sharp contrast to the poorer clinical outcome related to total survivin expression in AML reported by other groups.7–9 Different methodologies of measuring survivin levels (protein versus RNA), cohort sizes or detection in specific cellular pools (total cells, CD34+ cells or CD34+CD38− cells) can result in discrepancies. We used AML bone marrow samples taken at diagnosis only enriched by gradient centrifugation to avoid the use of antibodies that could modify protein expression after binding.34 It is, however, important to note that those patients positive for CytSurWT did not have a significantly higher percentage of CD34+CD38− stem cells at diagnosis than those negative in our series (data not shown). However, due to the unique biological features of survivin, detection in specific subcellular compartments (nuclear or cytoplasmic) seems to be crucial when investigating an impact on prognosis. Our findings suggest that cytoplasmic over-expression of survivin in AML may be coincident with cell cycle progression and can, therefore, be considered as a surrogate marker of cell proliferation.2,3,35
The prognostic impact of the PI3K/Akt/CytSurWT pathway described in this study provides important information for the clinical management of AML patients. PI3K/Akt inhibitors (MK2206, BKM120, LY2181308) or dual PI3K/mTOR inhibitors, as well as survivin antagonists (terameprocol EM-1421, CWP232291) are currently being evaluated in clinical trials, in order to improve results obtained with conventional chemotherapy in AML29. As a consequence of our findings, the PI3K/Akt/CytSurWT activated pathway represents a favorable prognostic factor identifying patients likely to achieve a long-term cure with conventional chemotherapy regimens. Moreover it seems plausible that inhibition of this pathway could result in only a selective depletion of the more proliferative cell fraction. Thus, it could be important to address whether targeting the PI3K/Akt/CytSurWT pathway could eliminate quiescent, putative leukemic stem cells in vivo in AML.
In conclusion, WT survivin and the 2B isoform are frequently expressed in AML in the nucleus, cytoplasm or both. The PI3K/Akt activated pathway increases cytoplasmic WT survivin expression leading to a more proliferative cell fraction. Activated PI3K/Akt/CytSurWT in AML identifies patients with a clearly superior outcome, with overall and relapse-free survival rates of over 60%.
Footnotes
The online version of this article has a Supplementary Appendix.
Funding
This study was supported by a grant from Ministerio de Ciencia e Innovación BFU2009-11826, by the Asociación Medicina e Investigación (AMI), and by a donation from “Instituto Roche para las Soluciones Integrales en Salud” and was also supported in part by an unrestricted grant from Celgene, Madrid, Spain
Authorship and Disclosures
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Stauber RH, Mann W, Knauer SK. Nuclear and cytoplasmic survivin: molecular mechanism, prognostic and therapeutic potential. Cancer Res. 2007;67(13): 5999–6002 [DOI] [PubMed] [Google Scholar]
- 2.Altieri DC. Survivin, cancer networks and pathway-directed drug discovery. Nat Rev Cancer. 2008;8(1): 61–70 [DOI] [PubMed] [Google Scholar]
- 3.Knauer SK, Bier C, Habtemichael N, Stauber RH. The case survivin-Crm1 interaction is essential for chromosomal passenger complex localization and function. EMBO Rep. 2006;7(12): 1259–65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Caldas H, Jiang Y, Holloway MP, Fangusaro J, Mahotka C, Conway EM, et al. Survivin splice variants regulate the balance between proliferation and cell death. Oncogene. 2005;24(12): 1994–2007 [DOI] [PubMed] [Google Scholar]
- 5.Adida C, Recher C, Raffoux E, Daniel MT, Taksin AL, Rousselot P, et al. Expression and prognostic significance of survivin in de novo acute myeloid leukaemia. Br J Haematol. 2000;111(1): 196–203 [DOI] [PubMed] [Google Scholar]
- 6.Carter BZ, Kornblau SM, Tsao T, Wang RY, Schober WD, Milella M, et al. Caspase-independent cell death in AML: caspase inhibition in vitro with pan-caspase inhibitors or in vivo by XIAP or survivin does not affect cell survival or prognosis. Blood. 2003;102(12): 4179–86 [DOI] [PubMed] [Google Scholar]
- 7.Tamm I, Richter S, Oltersdorf D, Creutzig U, Harbott J, Scholz F, et al. High expression levels of x-Linked inhibitor of apoptosis protein and survivin correlate with poor overall survival in childhood de novo acute myeloid leukemia. Clin Cancer Res. 2004; 10(11): 3737–44 [DOI] [PubMed] [Google Scholar]
- 8.Wagner M, Schmeiz K, Wuchter C, Ludwig WD, Dürken B, Tamm I. In vivo expression of survivin and its splice variants survivin-2B: impact on clinical outcome in acute myeloid leukemia. Int J Cancer. 2006;119 (6): 1291–7 [DOI] [PubMed] [Google Scholar]
- 9.Carter BZ, Qiu Y, Huang X, Diao L, Zhang N, Coombes KR, et al. Survivin is highly expressed in CD34+38− leukemic stem/progenitor cells and predicts poor clinical outcomes in AML. Blood. 2012;120 (1): 173–80 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Park S, Chapuis N, Tamburini J, Bardet V, Cornillet-Lefebvre P, Willems L, et al. Role of the PI3K/AKT and mTOR signaling pathways in acute myeloid leukemia. Haematologica. 2010;95(5): 819–28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tamburini J, Chapuis N, Bardet V, Park S, Sujobert P, Willems L, et al. Mammalian target of rapamycin (mTOR) inhibition activates phosphatidylinositol 3-kinase/Akt by up-regulating insulin-like growth factor-1 receptor signaling in acute myeloid leukemia: rationale for therapeutic inhibition of both pathways. Blood. 2008;111 (1): 379–82 [DOI] [PubMed] [Google Scholar]
- 12.Kornblau SM, Womble M, Qiu YH, Jackson CE, Chen W, Konopleva M, et al. Simultaneous activation of multiple signal transduction pathways confers poor prognosis in acute myelogenous leukemia. Blood. 2006;108(7): 2358–65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tamburini J, Elie C, Bardet V, Chapuis N, Park S, Broët P, et al. Constitutive phosphoinositide 3-kinase/Akt activation represents a favorable prognostic factor in de novo acute myelogenous leukemia patients. Blood. 2007;110(3): 1025–8 [DOI] [PubMed] [Google Scholar]
- 14.Min YH, Eom JI, Cheong JW, Maeng HO, Kim JY, Jeung HK, et al. Constitutive phosphorylation of Akt/PKB protein in acute myeloid leukemia: its significance as a prognostic variable. Leukemia. 2003;17(5): 995–7 [DOI] [PubMed] [Google Scholar]
- 15.Carter BZ, Milella M, Altieri DC, Andreeff M. Cytokine-regulated expression of survivin in myeloid leukemia. Blood. 2001;97 (9): 2784–90 [DOI] [PubMed] [Google Scholar]
- 16.Fukuda S, Singh P, Moh A, Abe M, Conway EM, Boswell HS, et al. Survivin mediates aberrant hematopoietic progenitor cell proliferation and acute leukemia in mice induced by internal tandem duplication of Flt3. Blood. 2009;114(2): 394–403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Belyanskaya LL, Hopkins-Donaldson S, Kurtz S, Simões-Wüst AP, Yousefi S, Simon HU, et al. Cisplatin activates Akt in small cell lung cancer cells and attenuates apoptosis by survivin upregulation. Int J Cancer. 2005;117(5): 755–63 [DOI] [PubMed] [Google Scholar]
- 18.Wang J, Yang L, Yang J, Kuropatwinski K, Wang W, Liu XQ, et al. Transforming growth factor beta induces apoptosis through repressing the phosphoinositide 3-kinase/AKT/survivin pathway in colon cancer cells. Cancer Res. 2008;68(9): 3152–60 [DOI] [PubMed] [Google Scholar]
- 19.Serrano-López J, Sanchez-Garcia J, Serrano J, Alvarez-Rivas MA, Garcia-Castellano JM, Roman-Gomez J, et al. Non-leukemic myeloid dendritic cells obtained from autologous stem cell products elicit antileukemia responses in patients with acute myeloid leukemia. Transfusion. 2011; 51(7): 1546–55 [DOI] [PubMed] [Google Scholar]
- 20.Guan Y, Gerhard B, Hogge DE. Detection, isolation and stimulation of quiescent primitive leukemia progenitor cells from patients with acute myeloid leukemia (AML). Blood. 2003; 101(8): 3142–9 [DOI] [PubMed] [Google Scholar]
- 21.Sanchez-Garcia J, Torres A, Herrera C, Alvarez MA. Cell cycle kinetic changes induced by interleukin-3 and interleukin-6 during ex vivo expansion of mobilized peripheral blood CD34 cells. Haematologica. 2006;91(1): 121–4 [PubMed] [Google Scholar]
- 22.Cheson BD, Bennett JM, Kopecky KJ, Büchner T, Willman CL, Estey EH, et al. Revised recommendations of the International Working Group for Diagnosis, Standardization of Response Criteria, Treatment Outcomes, and Reporting Standards for Therapeutic Trials in Acute Myeloid Leukemia. J Clin Oncol. 2003; 21(24): 4642–9 [DOI] [PubMed] [Google Scholar]
- 23.Andersen MH, Svane IM, Becker JC, Straten PT. The universal character of the tumor-associated antigen survivin. Clin Cancer Res. 2007;13:(20)5991–4 [DOI] [PubMed] [Google Scholar]
- 24.Li F, Yang J, Ramnath N, Javie MM, Tan D. Nuclear of cytoplasmic expression of survivin: what is the significance? Int J Cancer. 2005;114(4): 509–12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nakano J, Huang C, Liu D, Masuya D, Yokomise H, Ueno M, et al. The clinical significance of splice variants and subcellular localization of survivin in non-small cell lung cancers. Br J Cancer. 2008;98(6): 1109–17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Martinez A, Bellosillo B, Bosch F, Ferrer A, Marce S, Villamor N, et al. Nuclear survivin expression in mantle cell lymphoma is associate with cell proliferation and survival. Am J Pathol. 2004;164(2): 501–10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Nakagawa Y, Yamaguchi S, Hasegawa M, Nemoto T, Inoue M, Suzuki K, et al. Differential expression of survivin in bone marrow cells from patients with acute lymphocytic leukemia and chronic lymphocytic leukemia. Leuk Res. 2004;28(5): 487–94 [DOI] [PubMed] [Google Scholar]
- 28.Lopes R, Castro I, Pontes P, Candeias J, Lemoine NR, Sambade C. Expression profile of survivin in acute leukaemias: the importance of differential splicing. Leukemia. 2005;19(7): 1284–6 [DOI] [PubMed] [Google Scholar]
- 29.Martelli AM, Chiarini F, Evangelisti C, Cappellini A, Buontempo F, Bressanin D, et al. Two hits are better than one: targeting both phosphatidylinositol 3-kinase and mammalian target of rapamycin as a therapeutic strategy for acute leukemia treatment. Oncotarget. 2012;3(4): 371–94 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hideshima T, Catley L, Raje N, Chauhan D, Podar K, Mitsiades C, et al. Inhibition of Akt induces significant downregulation of survivin and cytotoxicity in human multiple myeloma cells. Br J Haematol. 2007;138 (6): 783–91 [DOI] [PubMed] [Google Scholar]
- 31.Liu JJ, Liu WD, Yang HZ, Zhang Y, Fang ZG, Liu PQ, et al. Inactivation of PI3k/Akt signaling pathway and activation of caspase-3 are involved in tanshinone I-induced apoptosis in myeloid leukemia cells in vitro. Ann Hematol. 2010;89(11): 1089–97 [DOI] [PubMed] [Google Scholar]
- 32.Kojima K, Shimanuki M, Shikami M, Samudio IJ, Ruvolo V, Corn P, et al. The dual PI3 kinase/mTOR inhibitor PI-103 prevents p53 induction by Mdm2 inhibition but enhances p53-mediated mitochondrial apoptosis in p53 wild type AML. Leukemia. 2008;22(9): 1728–36 [DOI] [PubMed] [Google Scholar]
- 33.Zhao P, Meng Q, Liu LZ, You YP, Liu N, Jiang BH. Regulation of survivin by PI3K/Akt/p70S6K1 pathway. Biochem Biophys Res Commun. 2010; 395(2): 219–24 [DOI] [PubMed] [Google Scholar]
- 34.Todisco E, Suzuki T, Srivannaboon K, Coustan-Smith E, Raimondi SC, Behm FG, Kitanaka A. CD38 ligation inhibits normal and leukemic myelopoiesis. Blood. 2000; 95(2): 535–42 [PubMed] [Google Scholar]
- 35.Fukuda S, Foster RG, Porter SB, Pelus LM. The antiapoptosis protein survivin is associated with cell cycle entry of normal cord blood CD34(+) cells and modulates cell cycle and proliferation of mouse hematopoietic progenitor cells. Blood. 2002;100(7): 2463–71 [DOI] [PubMed] [Google Scholar]