

Abstract
Circadian rhythms in mammals are generated by a feedback loop in which the three PERIOD (PER) proteins, acting in a large complex, inhibit the transcriptional activity of the CLOCK-BMAL1 dimer, repressing their own expression. Although fundamental, the mechanism of negative feedback in the mammalian clock, or any eukaryotic clock, is unknown. We analyzed protein constituents of PER complexes purified from mouse tissues and identified PSF (polypyrimidine tract binding protein-associated splicing factor). Our analysis indicates that PSF within the PER complex recruits SIN3A, a scaffold for assembly of transcriptional inhibitory complexes, and that the PER complex thereby rhythmically delivers histone deacetylases to the Per1 promoter, repressing Per1 transcription. These findings provide a function for the PER complex and a molecular mechanism for circadian clock negative feedback.
Circadian clocks are oscillators that drive daily rhythms of physiology and behavior (1). In mammals, circadian clocks are found in most or all tissues (2), where they play important roles in local and systemic physiology (3,4).
The mammalian clock is built at least in part on a conserved transcriptional feedback loop (1). The three PERIOD (PER) and two CRYPTOCHROME (CRY) proteins form a nuclear complex (PER complex) of ~1 megadalton (5), associate with CLOCK-BMAL1 (6,7), the dimeric transcription factor activating their expression, and repress its transcriptional activity (8–10). Turnover of PERs and CRYs (11,12) de-represses CLOCK-BMAL1 activity, initiating a new cycle.
Negative feedback is fundamental to the clock, but little is known about its mechanism. Apart from binding to CLOCK-BMAL1, PERs and CRYs have no known activity. Besides PERs and CRYs, only two constituents of a purified PER complex have been identified, NONO and WDR5 (5). NONO is a poorly understood RNA-binding protein important for clock function, and WDR5 is a histone methyltransferase subunit of plausible but uncertain relevance to the clock (5).
To search for unrecognized proteins important for circadian negative feedback, we set out to purify PER complexes from mouse tissues and identify constituent proteins by mass spectrometry. For efficient purification, we generated two mouse lines, one in which endogenous PER1 was replaced by a PER1 fusion protein tagged at the N-terminus with FLAG-Hemagglutinin (FH) (13) (FH-PER1) and another in which endogenous PER2 was replaced by a PER2 fusion protein tagged at the C-terminus (PER2-FH) (fig. S1; SOM text, in vivo epitope tag strategy). FH-PER1 and PER2-FH were similar in function to wild-type PERs in cells and in vivo in mice (fig. S1A-D). Because PER1 and PER2 are present in the same complexes (5), this strategy provided two ways of purifying PER complexes from mouse tissues, allowing cross-validation of results.
Tissue from littermate controls (with only wildtype PERs) was processed in parallel to that from FH-Per1 or Per2-FH mice. Livers were obtained at circadian time (CT) 20, a time when nuclear PER complex represses CLOCK-BMAL1 activity. PER complexes were isolated from nuclear extracts by anti-FLAG immunoaffinity purification, and PER-associated proteins were resolved by SDS-polyacrylamide gel electrophoresis (fig. S1E). Known PER-associated clock proteins co-purified with FH-PER1 and PER2-FH (fig. S1F).
After excluding proteins detected in controls, we found by mass spectrometry that in both the FH-PER1 and PER2-FH samples, known clock proteins accounted for ten of the fifteen proteins from which the most numerous unique peptides were identified. This set consisted of PER1, PER2, PER3, CRY1, CRY2, CLOCK, BMAL1, Casein Kinase-1ε and δ, and NONO. Others were RNA-binding proteins, to be described elsewhere (14). We focused on one particular RNA-binding protein, PSF (also called PTB-associated splicing factor), a pre-mRNA splicing factor related to NONO (15) that also acts as a transcriptional co-repressor (16).
The presence of PSF in endogenous PER complexes was confirmed by co-immunoprecipitation. Antibodies to PSF co-immunoprecipitated PER2, and antibodies to PER2 co-immunopreciptated PSF from mouse lung and liver nuclear extracts (CT18) (Fig. 1A). The abundance of Psf mRNA did not appear to oscillate (fig. S2A), but there was an apparent oscillation of nuclear PSF protein, peaking shortly after PER2 (fig. S2B). PSF over-expression repressed CLOCK-BMAL1 transcriptional activity, consistent with a function in the PER complex (fig. S2C,D).
Fig. 1.
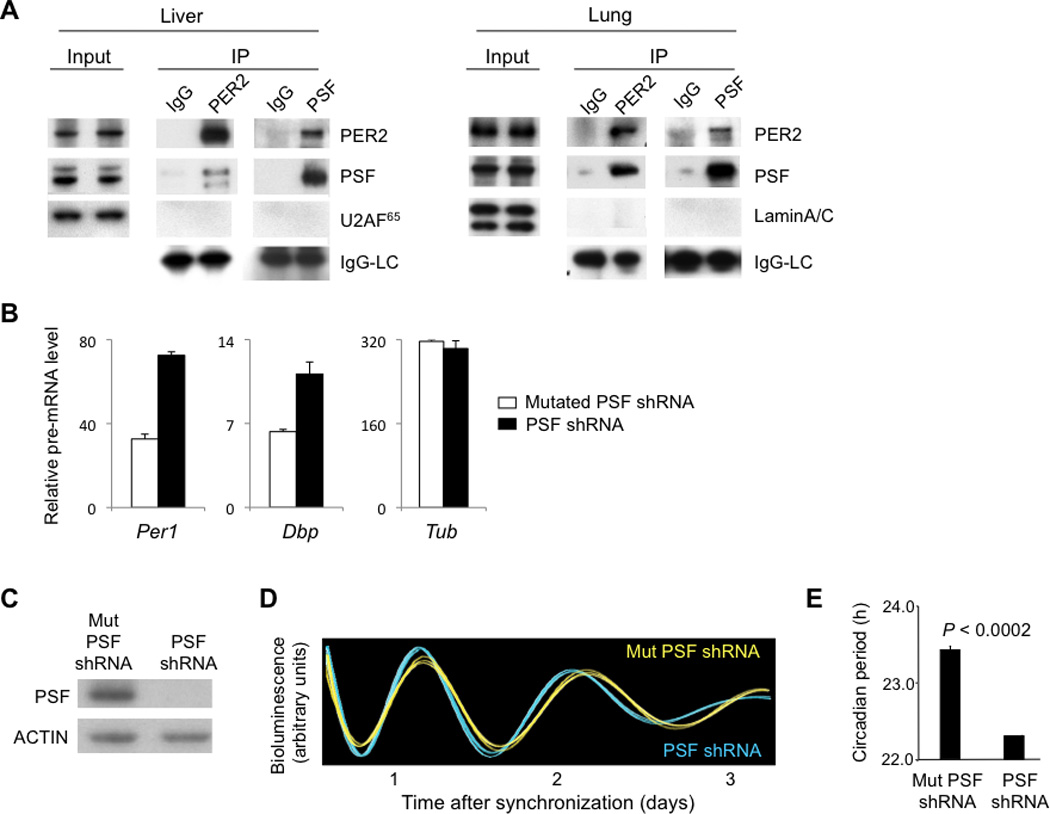
PSF is a constituent of endogenous PER complexes and is important for clock function. (A) Co-immunoprecipitation of PSF and PER2. Nuclear extracts (CT18) from liver or lung (input) and immunoprecipitates (IP, antibodies at top) from the extracts were probed with antibodies at right. U2AF65 and LaminA/C, negative controls; IgG-LC (light chain), positive control. (B) Depletion of PSF increases Per1 transcription. Quantitative RT-PCR assays showing steady-state abundance of indicated pre-mRNAs (normalized to Gapdh mRNA) in mouse fibroblasts after introduction of point-mutant control PSF shRNA (white) or after depletion of PSF by PSF shRNA (black). Shown are mean +/− SEM of triplicate experiments; representative of 3 experiments. (C-E) Short circadian period length caused by depletion of endogenous PSF from fibroblasts. (C) Western blot showing the effect of point-mutant control (Mut) shRNA or PSF shRNA on steady-state level of endogenous PSF. ACTIN, loading control. (D) Circadian oscillations of bioluminescence in synchronized reporter fibroblasts after delivery of control Mut PSF shRNA (yellow) or PSF shRNA (blue). Traces from three independent cultures are shown for each. (E) Circadian periods of fibroblasts in (D) (mean +/− SEM; N = 3 for each condition; t-test, two-tailed).
To determine whether PSF plays an important role in the transcriptional action of the PER complex, we introduced a small hairpin RNA construct (shRNA) into unsynchronized fibroblasts to deplete endogenous PSF. We then measured steady-state abundance of pre-mRNAs from two CLOCK-BMAL1 circadian target genes, Per1 and Dbp, and an arbitrary control gene. Depletion of PSF caused an increase in the average transcription of Per1 and Dbp, but it had no evident effect on transcription of the control (Fig. 1B). Thus endogenous PSF has a PER-like action in selectively repressing the transcriptional activity of CLOCK-BMAL1.
To determine whether PSF is important for clock function, we depleted PSF from reporter fibroblasts (17) and monitored circadian rhythms of bioluminescence in the synchronized cells. Three non-overlapping PSF shRNAs each caused a substantial depletion of endogenous PSF, and each caused a shortening of circadian period length that was evident in the real-time bioluminescence traces and highly significant in the group data (Fig. 1C-E; fig. S3). PSF thus plays a role in the clock mechanism.
PSF acts as a transcriptional co-repressor by recruiting the SIN3-HDAC complex (16, 18), a negative regulator of transcription (19). We therefore tested for SIN3A, the scaffold of the SIN3-HDAC complex (19), in endogenous PER complexes. An antibody to SIN3A co-immunoprecipitated PSF and PER2 from mouse lung and liver nuclear extracts (CT18) (Fig. 2A), and an antibody to PER2 co-immunoprecipitated SIN3A (fig. S4A).
Fig. 2.

PERs, PSF, and the SIN3-HDAC complex. (A) Co-immunoprecipitation of endogenous PER2 and PSF with SIN3A. Nuclear extracts (CT18) from liver or lung (input) and immunoprecipitates (IP, antibodies at top) from the extracts were probed with antibodies at right. U2AF65 and LaminA/C, negative controls; IgG-LC (light chain), positive control. (B) Synchronous circadian cycle of PER2, CRY2, PSF, SIN3A, and HDAC1 at Per1 proximal E-box site. ChIP assays from lungs sampled across a circadian cycle (bottom) performed with antibodies at the top left of each panel. ChIP values are relative to the signal at an arbitrary internal control genomic region (SOM methods). (C) Top, diagram of mouse Per1 gene showing positions of proximal E-box and control sites C1, C2, and C3. Bottom, ChIP assays from lungs harvested at CT10-14, performed with antibodies against the indicated proteins. ChIP data show mean +/− SEM of triplicate experiments; representative of 2-4 experiments.
To test whether PERs, CRYs, PSF, SIN3A, and HDAC1 (a histone deacetylase of the SIN3-HDAC complex) (19) are coordinately targeted to the Per1 promoter, as expected of proteins acting in the PER complex, we performed chromatin immunoprecipitations (ChIP) from lung and liver. PER2, CRY2, PSF, SIN3A, and HDAC1 exhibited a synchronous circadian rhythm at the Per1 proximal E-box (a CLOCK-BMAL1 binding site) (Fig. 2B; fig. S4B). We obtained similar results from liver (fig. S5). Peak occupancy occurred at CT 10-14, a time when PERs first appear in the nucleus (14) and when Per1 pre-mRNA begins to decline (fig. S5B). Enrichment of PER complex proteins was observed at the E-box but not other sites in the Per1 gene (Fig. 2C, fig. S5C), mirroring BMAL1 (Fig. 2C) and CLOCK (fig. S5C). As reported for Dbp E-box sites (20), we observed a circadian cycle of BMAL1 at the Per1 proximal E-box (Fig. 2B, fig. S5A). The BMAL1 cycle overlapped with the cycle of PER complex proteins, but it appeared to lead by a few hours (Fig. 2B).
We performed ChIP on lungs (CT 10) from wild-type littermates and Per1−/−; Per2−/− double mutants (21). Per1−/−; Per2−/− samples showed a reduction in PSF, SIN3A, and HDAC1 at the Per1 proximal E-box but little or no change at a control promoter (Fig. 3A). We obtained similar results from liver (fig. S5D). Thus PERs are important for recruiting PSF and the SIN3-HDAC complex to the Per1 promoter.
Fig. 3.

Presence of SIN3-HDAC1 at Per1 promoter depends on PER and PSF. (A) ChIP assays from lung (CT 10) comparing PSF, SIN3A, and HDAC1 at the Per1 proximal E-box or control promoter in wild-type littermates (white) and Per1−/−; Per2−/− mice (black). (B) ChIP assays, as in (A), from mouse fibroblasts after introduction of point-mutant control PSF shRNA (white) or after depletion of PSF by effective PSF shRNA (black). (C) ChIP assays, as in (A), from lung (CT14) comparing acetylation of HDAC1 targets (at bottom) at the Per1 E-box site or control promoter in wild-type littermates (white) and Per1−/−; Per2−/− mice (black). (D) ChIP assays, as in (C), from fibroblasts after introduction of point-mutant control PSF shRNA (white) or after depletion of PSF by effective PSF shRNA (black). ChIP values are plotted relative to the signal at an arbitrary internal control genomic region (SOM methods). All show mean +/− SEM of triplicate experiments and are representative of 2-4 experiments.
Depletion of PSF from unsynchronized fibroblasts caused a reduction in the average occupancies of PSF itself, SIN3A, and HDAC1 at the Per1 promoter, but had little or no effect on SIN3A or HDAC1 at a control promoter (Fig. 3B), phenocopying Per1−/−; Per2−/−. Depletion of PSF did not reduce PER2 at the proximal Per1 E-box (fig. S6). PSF is thus important for PER-dependent recruitment of the SIN3-HDAC complex to the Per1 promoter.
To test the role of the SIN3-HDAC complex in PER function, we monitored acetylation of HDAC1 histone target residues (22) in the Per1 promoter in lung (CT14) in wild-type littermates and Per1−/−; Per2−/− mutants. The Per1 promoter of Per1−/−; Per2−/− mice, but not a control promoter, exhibited increased acetylation of histone 3, lysine 9 in histone 3 (H3K9), and, to a lesser extent, lysine 5 in histone 4 (H4K5) (Fig. 3C). Similar results were obtained in fibroblasts after depletion of PSF (Fig. 3D). Thus the PER complex promotes histone deacetylation at the Per1 promoter by the PSF-dependent recruitment of the SIN3-HDAC complex.
We tested the role of the SIN3-HDAC complex in circadian negative feedback and clock function. Depletion of SIN3A from unsynchronized fibroblasts by a small interfering RNA (siRNA) caused an increase in the average transcription of the CLOCK-BMAL1 target genes Per1 and Dbp but had little or no effect on control gene transcription (Fig. 4A). Depletion of SIN3A from synchronized fibroblasts caused a shortening of circadian period length that was evident in the real-time bioluminescence traces and highly significant in the group data (Fig. 4BD). Thus SIN3A depletion mimicked the effects of PSF depletion on both Per1 transcription and clock function.
Fig. 4.

Depletion of SIN3A increases Per1 transcription and shortens circadian period length. (A) Quantitative RT-PCR assays, as in Fig. 1B, showing steady-state abundance of indicated pre-mRNAs in fibroblasts after introduction of point-mutant control SIN3A shRNA (white) or after depletion of SIN3A by an effective SIN3A shRNA (black). Shown are means +/− of triplicate experiment; representative of 3 experiments. (B-D) Short circadian period length caused by depletion of endogenous SIN3A from fibroblasts. (B) Western blot showing the effect of mutated control (Mut) siRNA or SIN3A siRNA on steady-state level of endogenous SIN3A. ACTIN, loading control. (C) Circadian oscillations of bioluminescence in synchronized circadian reporter fibroblasts after delivery of control Mut SIN3A siRNA (yellow) or SIN3A siRNA (blue). Traces from three independent cultures are shown for each condition. (D) Circadian period of fibroblasts with control Mut SIN3A siRNA or SIN3A siRNA (mean +/− SEM; N = 3; t-test, two-tailed).
These results establish a specific function for the PER proteins and a molecular mechanism for circadian clock negative feedback, central aspects of clock function that have long been poorly understood. Our analysis indicates that a PER complex rhythmically associates with DNA-bound CLOCK-BMAL1 at the Per1 promoter and, by virtue of its constituent PSF, recruits the SIN3-HDAC complex, thereby deacetylating histones 3 and 4 and repressing transcription. CLOCK preferentially acetylates H3K9 (23), so PER- and PSF- dependent recruitment of the SIN3-HDAC complex could serve to reverse modifications produced by CLOCK, generating a circadian rhythm of H3K9 acetylation. Our ongoing work indicates that a PER complex has an additional role in negative feedback, acting to repress Per and Cry transcriptional elongation (14).
The function of the SIN3-HDAC complex in transcriptional repression is conserved across virtually all eukaryotes (19). Circadian cycles of histone 3 acetylation have been observed at the promoters of clock genes in extraordinarily diverse organisms, including mammals (20, 24–26), insects (27), plants (28), and fungi (29). Thus it is conceivable that the role of the SIN3-HDAC complex in circadian negative feedback dates back to the evolutionary origins of eukaryotic circadian clocks.
Supplementary Material
Acknowledgments
We thank Ming Liu for technical assistance, Pat Nakatani (Dana Farber Cancer Institute) for the FH tag, David Weaver and Steve Reppert (University of Massachusetts) for Per1−/−; Per2−/− mice, Neal Copeland (Institute of Molecular and Cell Biology, Singapore) for reagents, and Yasuyuki Kurihara (Yokohama National University) for PSF plasmid. This work was supported by the G. Harold and Leila Y. Mathers Charitable Foundation (C.J.W.), a NIH Training Grant in Fundamental Neurobiology fellowship (H.A.D.), an Edward R. and Anne G. Lefler Center fellowship (M.S.R.), and a Swiss National Science Foundation fellowship (D.K.).
References
- 1.Takahashi JS, Hong HK, Ko CH, McDearmon EL. Nat Rev Genet. 2008;9:764. doi: 10.1038/nrg2430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Schibler U, Ripperger J, Brown SA. J Biol Rhythms. 2003;18:250. doi: 10.1177/0748730403018003007. [DOI] [PubMed] [Google Scholar]
- 3.Lamia KA, Storch KF, Weitz CJ. Proc Natl Acad Sci U S A. 2008;105:15172. doi: 10.1073/pnas.0806717105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Storch KF, et al. Cell. 2007;130:730. doi: 10.1016/j.cell.2007.06.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Brown SA, et al. Science. 2005;308:693. [Google Scholar]
- 6.Lee C, Etchegaray JP, Cagampang FR, Loudon AS, Reppert SM. Cell. 2001;107:855. doi: 10.1016/s0092-8674(01)00610-9. [DOI] [PubMed] [Google Scholar]
- 7.Chen R, et al. Mol Cell. 2009;36:417. doi: 10.1016/j.molcel.2009.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sangoram AM, et al. Neuron. 1998;21:1101. doi: 10.1016/s0896-6273(00)80627-3. [DOI] [PubMed] [Google Scholar]
- 9.Kume K, et al. Cell. 1999;98:193. doi: 10.1016/s0092-8674(00)81014-4. [DOI] [PubMed] [Google Scholar]
- 10.Griffin EA, Jr, Staknis D, Weitz CJ. Science. 1999;286:768. doi: 10.1126/science.286.5440.768. [DOI] [PubMed] [Google Scholar]
- 11.Siepka SM, et al. Cell. 2007;129:1011. doi: 10.1016/j.cell.2007.04.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Godinho SI, et al. Science. 2007;316:897. doi: 10.1126/science.1141138. [DOI] [PubMed] [Google Scholar]
- 13.Nakatani Y, Ogryzko V. Methods Enzymol. 2003;370:430. doi: 10.1016/S0076-6879(03)70037-8. [DOI] [PubMed] [Google Scholar]
- 14.Padmanabhan K, Robles MS, Weitz CJ. unpublished observations [Google Scholar]
- 15.Shav-Tal Y, Zipori D. FEBS Lett. 2002;531:109. doi: 10.1016/s0014-5793(02)03447-6. [DOI] [PubMed] [Google Scholar]
- 16.Mathur M, Tucker PW, Samuels HH. Mol Cell Biol. 2001;21:2298. doi: 10.1128/MCB.21.7.2298-2311.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Robles MS, Boyault C, Knutti D, Padmanabhan K, Weitz CJ. Science. 2010;327:463. doi: 10.1126/science.1180067. [DOI] [PubMed] [Google Scholar]
- 18.Dong X, Sweet J, Challis JR, Brown T, Lye SJ. Mol Cell Biol. 2007;27:4863. doi: 10.1128/MCB.02144-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Grzenda A, Lomberk G, Zhang JS, Urrutia R. Biochim Biophys Acta. 2009;1789:443. doi: 10.1016/j.bbagrm.2009.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ripperger JA, Schibler U. Nat Genet. 2006;38:369. doi: 10.1038/ng1738. [DOI] [PubMed] [Google Scholar]
- 21.Bae K, et al. Neuron. 2001;30:525. doi: 10.1016/s0896-6273(01)00302-6. [DOI] [PubMed] [Google Scholar]
- 22.Rundlett SE, et al. Proc Natl Acad Sci U S A. 1996;93:14503. doi: 10.1073/pnas.93.25.14503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Nakahata Y, et al. Cell. 2008;134:329. doi: 10.1016/j.cell.2008.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Etchegaray JP, Lee C, Wade PA, Reppert SM. Nature. 2003;421:177. doi: 10.1038/nature01314. [DOI] [PubMed] [Google Scholar]
- 25.Curtis AM, et al. J Biol Chem. 2004;279:7091. doi: 10.1074/jbc.M311973200. [DOI] [PubMed] [Google Scholar]
- 26.Naruse Y, et al. Mol Cell Biol. 2004;24:6278. doi: 10.1128/MCB.24.14.6278-6287.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Taylor P, Hardin PE. Mol Cell Biol. 2008;28:4642. doi: 10.1128/MCB.01612-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Perales M, Mas P. Plant Cell. 2007;19:2111. doi: 10.1105/tpc.107.050807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Belden WJ, Loros JJ, Dunlap JC. Mol Cell. 2007;25:587. doi: 10.1016/j.molcel.2007.01.010. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.