

Abstract
We present the sequencing and annotation of the Leishmania (Leishmania) amazonensis genome, an etiological agent of human cutaneous leishmaniasis in the Amazon region of Brazil. L. (L.) amazonensis shares features with Leishmania (L.) mexicana but also exhibits unique characteristics regarding geographical distribution and clinical manifestations of cutaneous lesions (e.g. borderline disseminated cutaneous leishmaniasis). Predicted genes were scored for orthologous gene families and conserved domains in comparison with other human pathogenic Leishmania spp. Carboxypeptidase, aminotransferase, and 3′-nucleotidase genes and ATPase, thioredoxin, and chaperone-related domains were represented more abundantly in L. (L.) amazonensis and L. (L.) mexicana species. Phylogenetic analysis revealed that these two species share groups of amastin surface proteins unique to the genus that could be related to specific features of disease outcomes and host cell interactions. Additionally, we describe a hypothetical hybrid interactome of potentially secreted L. (L.) amazonensis proteins and host proteins under the assumption that parasite factors mimic their mammalian counterparts. The model predicts an interaction between an L. (L.) amazonensis heat-shock protein and mammalian Toll-like receptor 9, which is implicated in important immune responses such as cytokine and nitric oxide production. The analysis presented here represents valuable information for future studies of leishmaniasis pathogenicity and treatment.
Keywords: genome, Leishmania amazonensis, interactome, amastin, heat-shock protein
1. Introduction
Leishmaniases are neglected infectious diseases caused by parasites belonging to the Trypanosomatidae family and the Leishmania genus. Leishmaniases are prevalent in tropical countries; ∼12 million people are affected by these diseases worldwide with 350 million people at risk of infection and an estimated yearly incidence of 2 million cases.1 Leishmania spp. are digenetic parasites that develop as promastigotes in the gut of phlebotominae sandflies and as intracellular amastigotes in the macrophages of vertebrate hosts. The Lainson and Shaw classification2 subdivides the Leishmania genus into two subgenera based on the localization of promastigotes in the insect alimentary tract. The subgenus Leishmania comprises species limited to the midgut and foregut of the sand fly, whereas the subgenus Viannia includes species that develop a prolonged phase in the hindgut with later migration of flagellates to the midgut and foregut of the vector's alimentary tract. More recently, a third subgenus has been included in Leishmania classification, the subgenus Sauroleishmania, which comprises species that exclusively parasitize lizards.3 A brief classification of Leishmania subgenera and species associated with the diverse array of leishmaniasis clinical manifestations (cutaneous, mucocutaneous, and visceral forms) is provided in Fig. 1A.
Figure 1.

Overview of the L. (L.) mexicana complex. (A) Classification of the Leishmania genus, subgenus and species complex (adapted from the WHO reports and Bates, 2007). Leishmania (L.) amazonensis and L. (L.) mexicana belong to the L. (L.) mexicana complex, subgenus Leishmania, and are causative agents of New World cutaneous leishmaniasis in which diffuse or disseminated lesions are hallmarks. The genomes of the species marked in red were employed in the present comparative analyses [*L. (S.) tarentolae was employed only in the amastin phylogenetic study]. (B) Large parasitophorous vacuoles (PVs) of L. (L.) amazonensis. Phase contrast microscopy image (left) of a bone marrow-derived macrophage containing a spacious PV (asterisk) lined with rounded amastigotes. Bar = 10 μm. Field-emission scanning electron micrograph (right) of an amastigote-hosting macrophage. The fractured sample indicated that amastigote forms (in red) were contained in a spacious PV. Bar = 5 μm.
Leishmaniases are primarily zoonotic diseases, and a variety of mammals acts as reservoirs of Leishmania species. Specifically, rodents, edentates, and marsupials typically harbor cutaneous leishmaniasis, whereas wild canines and domestic dogs are the main reservoirs of zoonotic visceral leishmaniases. In human hosts, disease outcomes are determined by a combination of parasitic properties (dermotropic versus viscerotropic species) and host factors, such as genetic variability and immune responses to infection.4,5 Among the causative species of cutaneous leishmaniases in Brazil, recent data indicate that 8% are attributed to Leishmania (L.) amazonensis.6 This species can cause simple and diffuse forms of cutaneous leishmaniasis (DCL) and was implicated recently in borderline disseminated cutaneous leishmaniasis, an intermediate form of disease.7
Infections with Leishmania species belonging to the Leishmania (L.) mexicana complex involve the dermal infiltration of macrophages that harbor parasites in large parasitophorous vacuoles (PVs). Most Leishmania species including Leishmania (L.) major, Leishmania (L.) donovani, and Leishmania (V.) braziliensis lodge intracellularly within small membrane-bound PVs that typically contain a single parasite and undergo fission as the amastigotes divide.8 In contrast, amastigotes of L. (L.) amazonensis and L. (L.) mexicana are housed in large numbers within spacious PVs (Fig. 1B) that fuse together.9 These enlarged PVs may subvert host cell defenses by facilitating conditions of relatively diluted hydrolytic enzymes.10,11 The mechanistic basis of spacious PV development remains unknown and likely is triggered by unidentified parasitic factors produced by L. (L.) amazonensis and other species from the L. (L.) mexicana complex, such as L. (L.) mexicana and Leishmania (L.) pifanoi.12
During the past decade, several reports have attempted to elucidate the factors used by Leishmania to interact with its vertebrate host and establish an infection. Like other kinetoplastids, gene expression in Leishmania is regulated mainly at the post-transcriptional level by RNA stability, rather than by promoters.13 Genes are organized into polycistronic transcriptional units, and protein-encoding genes are co-transcribed by RNA polymerase II. Precursor mRNAs subsequently are trans-spliced and polyadenylated.14–16
The number of chromosomes has been established for several Leishmania species.17–21 The molecular karyotypes of Old World Leishmania species [L. (L.) infantum, L. (L.) donovani, L. (L.) major, and L. (S.) tarentolae] each comprise 36 chromosomes,17 whereas the New World species, L. (V.) braziliensis, and L. (L.) mexicana have 35 and 34 chromosomes, respectively, due to fusion events involving 2–4 chromosomes.18,19
The genomes of two Old World Leishmania species, L. (L.) major Friedlin, and L. (L.) infantum JPCM5, and one New World species, L. (V.) braziliensis M2904, have been sequenced and annotated.19,22 Recently, the genomes of L. (L.) mexicana, 16 clinical isolates of L. (L.) donovani, and the lizard parasite, L. (S.) tarentolae, were sequenced and assembled using high-throughput DNA sequencing technologies.20,23,24 Despite evolutionary divergence within the Leishmania genus, Leishmania comparative genomics suggests a high degree of synteny.19,20,23,24 Leishmania spp. from the Leishmania and Viannia subgenera exhibit highly conserved gene sequences with remarkably few genes or paralog groups that are unique to any given species. However, L. (S.) tarentolae lacks genes associated with the intracellular life stages of human pathogenic Leishmania spp.20 On the other hand, the L. (V.) braziliensis genome includes features that are lacking in the genomes of Old World Leishmania spp., such as transposable elements and RNA interference (RNAi) machinery.19,25,26
To obtain a broader understanding of the pathogenesis of leishmaniasis, we sequenced the genome of the New World species, L. (L.) amazonensis. Using a comparative bioinformatics approach with other available Leishmania genomes, we searched for conserved domains and orthologous gene families among predicted proteins of L. (L.) amazonensis. In addition, we inferred the phylogeny of the surface glycoprotein, amastin, and generated a hybrid protein interactome to identify potential interactions between L. (L.) amazonensis secreted proteins and mammalian host factors.
2. Materials and methods
All the procedures employed in this study, except for phylogenetic analyses, are summarized in the workflow presented in Fig. 2. The workflow was divided into genome assembly and annotation steps, and functional and extended analyses of gene models.
Figure 2.

Bioinformatics analysis workflow used in the present study. Sequenced reads from the L. (L.) amazonensis genome were assembled into 2627 scaffolds and 8100 genes were predicted using comparative and ab initio prediction tools. The functional analysis of these predicted genes included: (i) AutoFACT functional annotation, which revealed that 45% of the predicted genes were unclassified or with unassigned function; (ii) screening for orthologous families of genes among Leishmania spp. (OrthoMCL); and (iii) screening for information about conserved protein domains deposited in CDD and PFAM databases. Expanded or exclusive orthologous proteins, or those conserved domains detected in the L. (L.) amazonensis genome were selected for interactome analysis with mammalian host proteins. This selection involved screening for possibly secreted proteins (using SecretomeP and TargetP) that also were orthologous to immune function-related proteins in humans and mice.
2.1. Genomic DNA extraction
Leishmania (L.) amazonensis (MHOM/BR/71973/M2269 strain) was extracted from the cutaneous lesions of a patient from Cafezal city, in the state of Pará, Brazil, in 1973. Since then, this strain has been maintained in the laboratory by inoculating hamsters and mice and by axenic culture. Parasites were cultivated in M199 culture medium supplemented with 10% fetal bovine serum. Genomic DNA was extracted from 109 promastigotes after the parasites were incubated in lysis buffer [50 mM Tris–HCl (pH 8.0), 62.5 mM EDTA (pH 9.0), 2.5 M LiCl, 4% Triton X-100] at 37°C for 5 min. DNA was purified using phenol–chloroform extraction (1:1 v/v) and ethanol precipitation. The resulting pellets were resuspended in 50 μl of 10 mM TE [Tris–HCl (pH 8.0), 1 mM EDTA] containing 0.6 μg/μl of RNase A (Life Technologies Corporation, USA), and were incubated at 37°C for 30 min. Genomic DNA was precipitated with 2.5 v of 100% ethanol and 0.3M sodium acetate, centrifuged at 15 700g at 4°C for 15 min, and resuspended in DNase-free water.
2.2. Genome sequencing and assembly
Leishmania (L.) amazonensis DNA sequences were obtained using the whole-genome shotgun strategy27 on a combination of 454 GS-FLX Titanium (Roche) and Solexa (Illumina, Inc.) instruments from the University of North Carolina (UNC, USA) sequencing facility. The GS-FLX sequencer generated single-end fragment reads (454 reads) with a mean length of 315 bp. The Illumina Genome Analyzer generated 76-bp paired-end fragment reads (Solexa reads) with an average insert size of 400 bp. Using an Perl script developed in-house, Illumina reads were filtered out if the average Phred quality score was lower than 20. For 454 sequences, reads comprising fewer than 100 bp or more than 500 bp (5% of each side of the normal distribution of read sizes) and reads with more than 1 unknown nucleotide (N) also were filtered out. The Solexa reads were assembled into longer scaffolds using the Velvet 0.7.56 de novo assembler28 with a k-mer parameter of 43. This value was calculated using the Velvet Optimizer script (Victorian Bioinformatics Consortium, Monash University, Australia), which tested a range of k-mers from 31 to 69. Newbler software29 was then used to assemble the 454 reads. The N50 scaffold and contig lengths of the L. (L.) amazonensis assembly were 22 275 and 17 272 bp, respectively. Solexa scaffolds and 454 contigs were combined by the Zorro assembler (Laboratório de Genômica e Expressão, UNICAMP, Brasil, http://lge.ibi.unicamp.br/zorro/downloads/Zorro2.2/DOC) to create the final hybrid genome assembly. The hybrid assembly was performed by combining the 4441 Illumina scaffolds and the 27 856 contigs from the 454 reads. Briefly, the Zorro pipeline consisted of (i) masking repeat regions in the contigs, (ii) detecting overlaps, (iii) unmasking repeat regions, and (iv) assembling hybrid contigs. In Phase 1, the repeat regions were determined by counting the occurrences of k-mers in the 454 reads. The assembler then masked k-mers in the contigs that occurred at high frequency. The absence of repeats enabled accurate overlap detection in Phase 2. The contigs were then unmasked in Phase 3, and the correct hybrid assembly was obtained in Phase 4 by merging all overlapping contigs into hybrid contigs. Overlap detection and consensus generation were performed using the Minimus package.30 The hybrid contigs were ordered and oriented with the Bambus program31 using paired-end information and manual verification, which yielded 2627 scaffolds.
2.3. Gene identification and annotation
2.3.1. Alignment of Leishmania spp. proteins with the L. (L.) amazonensis genome
Predicted proteins for L. (V.) braziliensis (8153 proteins), L. (L.) infantum (8154 proteins), L. (L.) major (8298 proteins), and L. (L.) mexicana (8007 proteins) were mapped onto the L. (L.) amazonensis genome sequence using the Exonerate program (v. 2.2.0),32 which performs genomic searches and spliced alignments in a single run.
2.3.2. Ab initio prediction of gene models
Glimmer, v. 3.02 33 and Genemark.hmm, v. 3.3 34 programs were used to perform ab initio gene predictions. The Glimmer long-orfs program was trained on non-overlapping open reading frames (ORFs) exceeding 200 bp in length. Genemark.hmm was executed in self-training mode by considering ORFs of at least 200 bp in length.
2.3.3. Combined gene models
The above resources were used to automatically create L. (L.) amazonensis gene models using EVidenceModeler (EVM, v. r03062010) software.35 For individual outputs generated by Exonerate, Glimmer and Genemark.hmm were given values reflecting our data confidence to define gene structure. We considered the following confidence values: Glimmer = 3, Genemark = 3, nucleotide-to-protein match = 6. The gene model was considered reliable only if it was predicted by all ab initio software or if it had similarity with one of the compared species. A final set of predicted gene models was then selected and manually inspected. Incomplete genes or genes overlapping gap regions were inspected with input from blastx alignments against protein databases and scaffold edges.
2.3.4. Automatic annotation and analysis of protein domains
The automatic annotation software, AutoFACT (v. 3.4),36 was used for functional annotation of gene models. AutoFACT enables transitive annotation based on sequence similarity searches in several databases. We used the blastp algorithm (e-value 1 × 10–5) to model gene alignments against the following protein databases: non-redundant proteins (NCBI RefSeq database, downloaded 8/6/2010), Swiss-Prot (only manually curated proteins, downloaded 4/15/2010),37 UniRef90, and UniRef100 (UniProt databases of clustered protein families, downloaded 4/15/2010), CDD (conserved protein domains, downloaded 4/15/2010),38 PFAM (protein domains, downloaded 4/15/2010),39 and KEGG (metabolic pathways, downloaded 4/15/2010).40 We set AutoFACT to consider the following order of importance for annotation: UniRef100, UniRef90, KEGG, non-redundant proteins, and CDD. Data from CDD–PFAM analyses were extracted from AutoFACT and were used for comparative analyses of CDD–PFAM protein domains among L. (L.) amazonensis, L. (V.) braziliensis, L. (L.) infantum, L. (L.) major, and L. (L.) mexicana. These data also were evaluated using reverse PSI-BLAST (RPS-BLAST, e-value cutoff of 1 × 10−5).
2.3.5. Orthologous gene analysis
A data set composed of all the Leishmania spp. gene models was created and compared all-against-all using blastp (e-value cutoff of 1 × 10−5). The results were submitted to OrthoMCL (v. 1.4) software,41 which clustered the proteins into orthologous and paralogous families. We applied the default software parameters, including an inflation index of 1.5. The inflation index regulates the cluster tightness (granularity) associated with sensitivity and selectivity.42 Clusters of proteins that presented bidirectional similarities between at least two Leishmania species were considered orthologs.
2.3.6. Pseudogene identification
Leishmania and Trypanosoma spp. proteins were aligned against the L. (L.) amazonensis genome using blastx with an e-value threshold of 1 × 10−5. The coordinates of the first hit alignment for all proteins were converted to GFF file format using an in-house Perl script. The BEDTools package43 was used to identify L. (L.) amazonensis regions exceeding 200 bp that showed similarity with Leishmania or Trypanosoma spp. proteins without overlapping with the gene model predictions. These regions were compared against the NCBI non-redundant database using the blastx program and manual annotation.
2.3.7. Calculation of the codon adaptation index
The CodonW v. 1.4.4 software (http://bioweb.pasteur.fr/seqanal/interfaces/codonw.html) was employed to calculate the codon usage indices of each L. (L.) amazonensis predicted gene. The codon adaptation index (CAI) estimates the extent of bias toward codons known to be preferred in highly expressed genes.44 This index ranges from 0 to 1.0 with higher values indicating stronger codon usage bias and a higher expression level. The frequency of codon usage in highly expressed genes defines the relative fitness values for each synonymous codon. These values were calculated from the relative synonymous codon usage rather than from the raw codon usage and therefore were essentially independent of amino acid composition. Because fitness values are highly species specific, we first identified a set of highly expressed genes in L. (L.) amazonensis. This set was input into the calculation of the CAI. The effective number of codons (Nc) also was used to quantify the codon usage bias of each gene. The Nc ranges from 20 for a gene with extreme bias (using only 1 codon per amino acid) to 61 for a gene with no bias (using synonymous codons equally).45,46 Sequences for which Nc values are less than 30 were considered highly expressed, whereas sequences with Nc values exceeding 55 were considered poorly expressed genes.
2.4. Phylogeny of amastin surface proteins
A phylogeny of Leishmania amastin proteins was built from a set of L. (L.) amazonensis amastins predicted by our assembled genome and from all predicted amastin proteins in the T. cruzi, T. brucei, L. (L.) major, L. (L.) infantum, L. (V.) braziliensis, L. (L.) mexicana, and L. (S.) tarentolae genomes. These amastin sequences [except L. (L.) amazonensis amastins] were extracted from TriTrypDB47 (accessed 6/28/2012) by searching for ‘amastin’ or ‘amastin-like’ entries.
The 181 amastin/amastin-like protein sequences encoded by L. (L.) braziliensis, L. (L.) infantum, L. (L.) major, L. (L.) mexicana, and L. (S.) tarentolae and 24 amastin/amastin-like protein sequences from L. (L.) amazonensis were aligned using Geneious software (v. 5.6.3),48 in which an embedded MUSCLE software was applied.49 The alignment was performed using default parameters. The phylogenetic tree was inferred by Bayesian methods using MrBayes v. 3.1.250 with tree parameter optimization during the generations. A Bayesian tree was inferred based on 1 × 107 generations with a burn-in value of 75 000. Data were saved every 100 generations and were run in 4 chains during 2 runs. The Whelan and Goldman (WAG) substitution matrix was used for the protein alignment.51
2.5. Interactome of mammalian proteins and Leishmania secreted proteins
From the CDD and OrthoMCL analyses, protein families that were found to be expanded in L. (L.) amazonensis [i.e. more gene/domain copies when compared with L. (V.) braziliensis, L. (L.) infantum, and L. (L.) major] were also screened for orthologous in human protein databases. To identify human proteins that are orthologous to L. (L.) amazonensis proteins, we performed a blastp search (e-value cutoff of 1 × 10−10) against the human protein databases using the parasite's expanded proteins as query. These selected gene models were then analyzed using TargetP 1.1 and SecretomeP 2.0 prediction tools.52,53 Our objective was to identify proteins that could be secreted to the extracellular compartment or exposed for interaction with host cell components. We considered as putative secreted proteins those gene products for which TargetP returned a signal peptide value exceeding 0.8 and for which other values, such as the mitochondrial targeting peptide and the chloroplast transit peptide, were below 0.2. L. (L.) amazonensis proteins implicated in non-classical secretion or ectodomain shedding were predicted using SecretomeP v. 2.0 with the recommended threshold of 0.5.
Considering that amastigote is the developmental form which maintains a durable relationship with the mammalian host cell, an additional step was included in the screening. Using the screened gene models above mentioned, we performed a blastp analysis (e-value cutoff of 1 × 10−5) against L. (L.) mexicana proteins expressed by amastigotes and identified in proteomic databases.54 L. (L.) amazonensis gene models that attend to these four criteria were considered for hybrid interactomes: (i) expanded or exclusive in L. (L.) amazonensis; (ii) orthologous to human proteins; (iii) possibly secreted (start codon was manually verified); and (iv) possibly expressed by amastigotes.
Some of the screened gene products could allow for interactions and interferences with native mammalian interactomes. Interactome networks for parasite-secreted proteins and for human and mouse proteins were created using IPA software (Ingenuity Systems, Redwood City, CA, USA) which was configured to build interactomes considering only those proteins expressed by cells of the immune system.
3. Results and discussion
3.1. Annotation of Leishmania (L.) amazonensis gene models
Approximately 37 million 76-bp paired-end reads (average insert size, 400 bp) were assembled into 4411 scaffolds (coverage, ∼94×) using Velvet de novo assembler software. The 454 reads (179 112 reads; average read length, 312 bp) were assembled using Newbler software into 27 856 contigs (coverage, ∼2×. The final assembly was performed using Zorro, which combined scaffolds from Solexa and contigs from 454 sequencing to generate 2627 scaffolds (≥1000 bp in length) that specified an (L.) amazonensis genome size of 29.6 Mb.
Ab initio gene prediction was conducted using Glimmer and Genemark.hmm programs, which gave 8032 and 11 641 gene models (gene lengths ≥150 nt), respectively. Comparative gene predictions consisted of proteins from L. (V.) braziliensis, L. (L.) infantum, L. (L.) mexicana, and L. (L.) major aligned against the final L. (L.) amazonensis genome assembly. The final set of 8168 gene models was created using a combination of ab initio and comparative gene model analyses, EVM software to identify consensus gene structures supported by these two approaches, and manual annotation. Incomplete genes and genes overlapping gap regions were discarded from further analyses. Following manual annotation, 8100 gene models remained with an average length of 1793 bp; this is consistent with other sequenced Leishmania species. These data are summarized and compared with other Leishmania genomes in Table 1.
Table 1.
Summary of the information obtained from the genome sequences of Leishmania spp.
| L. (L.) amazonensis | L. (L.) mexicana | L. (L.) major | L. (L.) infantum | L. (V.) braziliensis | L. (L.) donovani | L. (S.) tarentolae | |
|---|---|---|---|---|---|---|---|
| Contigs | 3199 | 35 | 36 | 37 | 1041 | 2154 | N/A |
| Genome length (Mb) | 29.6 | 32.1 | 32.8 | 32.1 | 33.7 | 32.4 | 30.4 |
| chromosomes | 34* | 34 | 36 | 36 | 35 | 36 | 36 |
| Number of predicted genes | 8100 | 8007 | 8298 | 8216 | 8153 | 8252 | 8201 |
| Gene density (genes/Mb) | 273 | 256 | 260 | 252 | 228 | 254 | 270 |
| G + C content (%) | 58.5 | 60.5 | 59.7 | 59.3 | 57.8 | >60 | 57.2 |
| CDS G + C content (%) | 61 | 61.23 | 62.5 | 62.45 | 60.38 | 61 | 58.4 |
| References | Current study | Rogers et al.24 | Ivens et al.22 | Peacock et al.19 | Peacock et al.19 | Downing et al.23 | Raymond et al.20 |
The number of L. (L.) amazonensis chromosomes (*) was inferred by mapping against L. (L.) mexicana chromosomes using the software SSAHA2 with all the L. (L.) amazonensis reads generated by Illumina.
N/A, not available.
The final set of gene models were annotated automatically by comparing them against protein databases (blastp) and summarizing with AutoFACT software. The results indicated that 55% of the gene models showed high sequence similarity (e-value 1 × 10−10) to functionally annotated proteins, whereas 42% of the models were similar to unassigned proteins (i.e. proteins with no functional annotation). The remaining 3% yielded no hits with any databases and were considered unclassified (Fig. 2).
In agreement with previous reports,19,20,22–24 our genome sequence analysis indicated that more than 90% of the 8100 L. (L.) amazonensis genes are shared with other human pathogenic Leishmania spp. with little variation in orthologous gene content. Despite varying clinical manifestations and features of lesions, Leishmania spp. harbor a conserved genomic core encoding functions ranging from fundamental biological processes to complex host–parasite interaction networks.
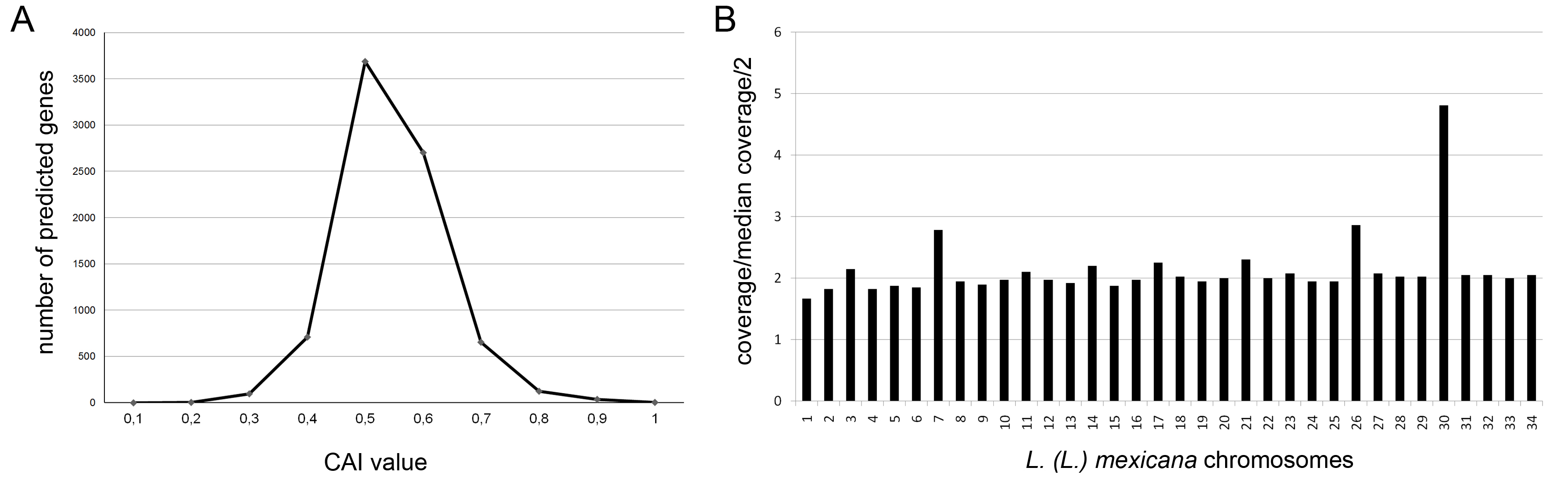
We performed a comparative annotation of tRNAs from the L. (L.) amazonensis, L. (L.) infantum, L. (V.) braziliensis, and L. (L.) major genomes. Using tRNAscan-SE software,55 the annotation yielded very similar numbers among the studied Leishmania species (Supplementary data, Table S1). Given the number of tRNAs for each codon in L. (L.) amazonensis, we calculated the CAI for all annotated gene models (Supplementary data, Table S2). The index measures the codon usage bias presented by highly expressed genes and can be comparatively employed to score native genes (higher CAI, more adapted to the pattern of codon usage) and possibly foreign or transferred genes (lower CAI, less adapted to the pattern of codon usage).56 Ranging from 0 (non-expressing gene, likely pseudogene) to 1 (highly expressed gene), we found that L. (L.) amazonensis predicted genes have a mean CAI value of 0.49 and median of 0.48 (Supplementary data, Fig. S1A). These data will be useful for future analyses of recent events of horizontal gene transfer in Leishmania.
Additionally, we scored regions in the L. (L.) amazonensis genome that showed similarities with predicted genes in other Leishmania and trypanosomatid species but were not identified as ORFs due to premature stop codons or frame shifts. This approach yielded 36 genomic regions corresponding to potential pseudogenes (Supplementary data, Table S3). One L. (L.) amazonensis pseudogene identified in our analysis was a fragment of argonaute 1 (AGO1), which is involved in the RNAi machinery. In the genome of L. (L.) amazonensis, we did not detect the known trypanosomatid argonaute and dicer variants, AGO1, DCL1 or DCL2. In addition, proteins containing two RNAse III domains (characteristic of dicer) or PAZ and Piwi domains (characteristic of argonaute) were not detected in this analysis. Our results suggest that RNAi pathways are absent in L. (L.) amazonensis, corroborating the hypothesis that RNAi via dicer and argonaute has been lost from the Leishmania subgenus following its divergence from the Viannia subgenus.26
Leishmania generally is considered to be a diploid organism because it carries two copies of most of its homologous chromosomes.19,22,24,57,58 However, there is increasing evidence suggesting that aneuploidy can occur in Leishmania species.23,24,58,59 Chromosome copy numbers can vary considerably among strains and species from different geographic regions, even among recent isolates.23,24 We mapped the L. (L.) amazonensis reads generated by Illumina against the L. (L.) mexicana chromosomes using the software SSAHA2.60 The median of coverage along each chromosome indicated a probable extra copy of chromosomes 7 and 26 and 3 extra copies of chromosome 30 in L. (L.) amazonensis (Supplementary data, Fig. S1B). In L. (L.) amazonensis, the exact number of chromosomes has not been defined. Preliminary studies using pulsed field gel electrophoresis have reported that the L. (L.) amazonensis karyotype consists of 25 chromosomal bands ranging in size from 0.2 to 2.2 Mb.61 Some bands exhibited variable ethidium bromide staining intensities possibly due to co-migration of chromosomes of similar sizes. Further studies will be needed to define the number of chromosomes and ploidy in L. (L.) amazonensis.
3.2. Functional analysis of gene models
In our functional analysis of gene models, we focused on the common factors, rather than the species-specific factors, predicted in L. (L.) amazonensis and L. (L.) mexicana genomes and not predicted in the genomes of the other species. We chose one genome data set for each disease outcome to compare with L. (L.) amazonensis and L. (L.) mexicana. Specifically, we chose leishmaniasis causative agents representative of cutaneous [L. (L.) major], mucocutaneous [L. (V.) braziliensis], and visceral [L. (L.) infantum] infections.
We searched for L. (L.) amazonensis genes that could be expanded or contracted in terms of gene or domain copies compared with the other species, particularly L. (L.) mexicana. We present a discussion of some of these expanded genes/domains that could participate in: (i) parasite tropism in host organisms via adhesion molecules or amastin surface proteins; (ii) the development of large PVs by lipid synthesis; and (iii) intracellular establishment by enzymes related to nutritional acquisition and resistance to host intracellular defenses, such as oxidative burst.
3.2.1. Expanded and contracted orthologous gene families
The OrthoMCL software was applied to genome data sets from L. (L.) amazonensis, L. (L.) mexicana, L. (L.) infantum, L. (V.) braziliensis, and L. (L.) major to identify Leishmania orthologous gene families. We identified 7826 orthologous gene families [7488 orthologous gene families containing ≥1 L. (L.) amazonensis protein] with 468 families comprising at least 7 members. Most (6784) of the orthologous gene families were shared by other Leishmania spp. Eight families were found only in L. (L.) amazonensis and L. (L.) mexicana and 23 families were unique to L. (L.) amazonensis (Fig. 3A, Supplementary data, Table S4). The five largest families identified across human pathogenic Leishmania spp. using OrthoMCL also were identified in L. (L.) amazonensis: dyneins (Family 1), glycoprotein GP63 (Family 3, leishmanolysins), histone H4 (Family 4), ABC transporters (Family 8), and amastin proteins (Family 14; Supplementary data, Table S4). We identified several families associated with 2-fold or more gene copies in L. (L.) amazonensis than in any of the other human pathogenic species. These families include aminotransferases (family 256), 60S ribosomal protein L37 (family 216), and hypothetical proteins (families 323, 5508, and 7732). Among the families with at least 2-fold fewer gene copies in L. (L.) amazonensis we highlighted the GP63 gene family. The list of contracted gene families requires further investigation because some of them could be present in unassembled regions.
Figure 3.

Diagrammatic representation of (A) species-specific orthologous gene families (OrthoMCL analysis) and (B) conserved domains (CDD–PFAM analysis). A core of 6784 orthologous families and 1881 domains was conserved in all studied Leishmania species [L. (L.) amazonensis, L. (L.) mexicana, L. (L.) major, L. (L.) infantum, and L. (V.) braziliensis]. We detected 8 orthologous families and 20 conserved domains that were exclusive to L. (L.) mexicana complex. A complete list of orthologous families and conserved domains is presented in Supplementary data, Table S4 and S5, respectively.
Comparative genome analyses of L. (L.) amazonensis and L. (L.) mexicana indicated an expansion in the gene family encoding a class-IV branched-chain amino acid aminotransferase (OrthoMCL family 256) that consisted of 270–415 amino acid residues and shared few regions of sequence similarity.62 Branched-chain aminotransferases catalyze the synthesis of leucine, isoleucine, and valine, and may be used to fulfill the parasite's nutritional requirements. They also may be involved in parasite sterol and prenol lipid synthesis because leucine is the main isoprenoid precursor for L. (L.) mexicana promastigotes and amastigotes.63,64
A 3′-nucleotidase/nuclease (OrthoMCL family 7761) gene was predicted as being exclusive to the L. (L.) amazonensis and L. (L.) mexicana genomes. This gene encodes an enzyme responsible for nucleic acid hydrolysis that was found to be dramatically up-regulated on the cell surface of the trypanosomatid, Chritidia luciliae, under purine starvation conditions.65 An ecto-3′-nucleotidase/nuclease was detected experimentally in L. (L.) amazonensis; this component has important implications for parasite nutrition, adhesion to host cells, and infectivity.66
Substantial differences in the gene copy numbers between certain Leishmania spp. may account for the observed phenotypic variability in terms of pathogenesis and virulence. Several genome features could contribute to quantitative variation in gene copies among Leishmania spp. The expansion and contraction of genes in tandem arrays could result in up- or down-regulation of gene expression associated with copy-number variation. In addition, extensive variation in aneuploidy frequencies within parasite populations has been reported for several Leishmania spp. and for different Leishmania isolates within the same species.23,24,59
3.2.2. Expanded and contracted conserved domains
Another comparison between Leishmania genomes was based on the identification of predicted protein domains in CDD–PFAM databanks. The result of CDD–PFAM analysis, included in the AutoFACT annotation, was retrieved and manually evaluated. In total, 2509 protein domains were identified; 2186 of these domains were detected in at least one L. (L.) amazonensis protein (Supplementary data, Table S5). Most (1881) of the identified domains were shared by other Leishmania spp.; 20 domains were found exclusive to L. (L.) mexicana complex [L. (L.) amazonensis, and L. (L.) mexicana], and 26 domains were unique to L. (L.) amazonensis (Fig. 3B, Supplementary data, Table S5). The expanded and contracted protein domains in L. (L.) amazonensis were evaluated by the same criteria applied in OrthoMCL (i.e. variations in gene copies when compared with other Leishmania genomes). Among the most prevalent domains were the heat-shock protein (HSP)70 chaperone (CDD: 143803) and the vacuolar protein sorting-associated protein MRS6 (CDD: 34648). The following protein domains were detected only in L. (L.) amazonensis and L. (L.) mexicana: thioredoxin domain (CDD: 32932), tat-binding protein 1 (CDD: 148614), sterile alpha motif (SAM) domain of bicaudal C homolog protein 1 (BCC1, CDD: 188919), hydrolase (CDD: 188206), and ATPase (CDD: 190944).
As examples of contracted protein domains identified in L. (L.) amazonensis, we highlight: amastins (CDD: 140228), UDP-GlcNAc-dependent glycosyltransferase (CDD: 140237), leishmanolysin peptidase M8 (CDD: 189994), cathepsin (CDD: 185513), tryparedoxin peroxidase (CDD: 140280), non-long terminal repeat reverse transcriptases (RTs-nLTR, CDD: 73156), rim ABC transporters (CDD: 185513), adenylyl/guanylyl cyclase (CDD: 128359), and paraflagellar rod protein (CDD: 140353).
The present study detected a thioredoxin domain unique to L. (L.) amazonensis and L. (L.) mexicana (NCBI accession COG3118). Thioredoxin functions as a hydrogen donor or disulfide reductase and is involved in the response to oxidative stress and in protein folding.67 Reactive oxygen species can be scavenged directly by thioredoxin or by thioredoxin-related dehydrogenases. Parasitic thiol and dithiol proteins may buffer the redox environment of PVs; this could account for the resistance of L. (L.) amazonensis to nitric oxide (NO) production in interferon (IFN)-γ-activated macrophages.68–70
3.3. Extended gene model analysis
3.3.1. Amastin phylogeny suggests specialized amastins in the Leishmania (L.) mexicana complex
Previous Leishmania phylogenetic analyses, based on comparisons of isoenzymes, DNA sequences, and HSP profiles among species agreed with the adopted Linnean classification and with complexes proposed by Lainson and Shaw in 1987.4,71,72 The phylogenies from these studies indicated that L. (L.) amazonensis has an evolutionary proximity to L. (L.) mexicana, a finding that was interpreted as these species comprising a monophyletic clade. These studies also indicate that parasites responsible for cutaneous/mucocutaneous lesions could be as different and divergent from one another as they are from the parasites that cause visceral leishmaniasis. For instance, the phylogenetic distance between the cutaneous-associated species, L. (L.) amazonensis and L. (L.) major, is similar to the distance between L. (L.) amazonensis/L. (L.) major and L. (L.) donovani, which causes visceral leishmaniasis. Therefore, the same disease outcomes in Leishmania mammalian hosts can result from a variety of evasive strategies and factors distinctively featured by Leishmania spp.
Amastin belongs to a multi-gene family in Leishmania that encodes small surface proteins of ∼200 amino acids. Several members of the amastin gene family are dispersed throughout the genomes of all Leishmania species and exhibit various expression patterns.73 Phylogenetic analysis of trypanosomatid amastins defined four subfamilies of amastin (α, β, γ, and δ) with distinct genomic organizations and expression patterns during the cell cycles of T. cruzi and Leishmania spp.74 δ-amastins comprise the largest and most diverse amastin subfamily. In T. cruzi, δ-amastin expression was associated with parasite infectivity to host cells.75 In Leishmania, the amastin N-terminal signature peptides are among the most immunogenic of all leishmanial surface antigens in mice76 and generate strong immune responses in humans with visceral leishmaniasis.77 DNA microarray analyses have implicated amastin in the intracellular survival of the parasite.78 Amastin gene expression was detected predominantly in amastigotes of several L. (L.) donovani strains isolated from patients with visceral and post-kala-azar dermal leishmaniasis.79 The roles of amastin genes in parasite homeostasis and growth inside acidic PVs also have been addressed.73,80 As transmembrane proteins, amastins could contribute to proton or ion trafficking across the membrane to adjust cytoplasmic pH under the harsh conditions of the phagolysosome. As a surface epitope, amastin may be recognized by opsonizing host IgG antibodies and could promote parasite uptake by host macrophages (via Fc receptors) and subsequent release of interleukin (IL)-10.81 We speculate that amastins could be involved in certain peculiar characteristics of L. (L.) amazonensis, such as its propensity to induce DCL, its development inside spacious PVs, and its resistance to the highly oxidative phagolysosomal environment in host cells.
The TriTrypDB provides a set of annotated amastin and amastin-like surface proteins in the Leishmania and Trypanosoma genomes. Our search of the TriTrypDB yielded 181 annotated genes encoding amastin or amastin-like proteins in L. (L.) braziliensis, L. (L.) infantum, L. (L.) major, L. (L.) mexicana, and L. (S.) tarentolae. The L. (L.) amazonensis genome presented in this study identified 12 orthologous groups annotated as amastin or amastin-like proteins (families 14, 19, 20, 3539, 3935, 5852, 6119, 6120, 6543, 7556, 7771, and 7778). All families corresponded to 1 representative gene model, except family 19 (3 gene models were associated with this orthologous group). Thus, our OrthoMCL analysis predicted 14 amastin/amastin-like proteins in the L. (L.) amazonensis genome. All of these proteins also were identified in the CDD–PFAM analysis. The CDD–PFAM list also identified gene models containing amastin domains that were not identified in the OrthoMCL analysis. We gathered all gene models identified as amastin/amastin-like proteins (OrthoMCL) or containing amastin domains (CDD–PFAM) and built a list of 24 L. (L.) amazonensis predicted amastin/amastin-like surface proteins to perform our phylogenetic analysis.
Alignment of the Leishmania-annotated amastins with 24 amastin/amastin-like proteins identified in the L. (L.) amazonensis genome (OrthoMCL and CDD–PFAM combined scoring) allowed us to build an amastin phylogenetic tree (Fig. 4). By placing the phylogenetic root halfway between the two most divergent/distant amastins (midpoint rooting) we identified clades composed of species-related amastins. Early branching clades could represent a class of amastin surface proteins conserved in Leishmania prior to its radiation (Fig. 4, blue branches). These Leishmania pre-speciation amastins are gathered in α, β, and γ subfamily clades.74 The presence of species-specific clades of δ-amastins at terminal tree branches (Fig. 4, red, green, yellow, and purple branches) suggests that several amastin surface proteins appeared because of environmental selective pressures or pathogen speciation. At least in part, this could be associated with the diverse leishmaniasis outcomes of different Leishmania species. It is possible to identify amastin subfamilies unique to L. (L.) major, L. (V.) braziliensis, and L. (L.) infantum, and three groups in which L. (L.) amazonensis and L. (L.) mexicana amastins represent a distinct clade of amastins (Fig. 4, red branches). These amastins could play a role in the unusual housing of these parasites within spacious PVs of infected macrophages.
Figure 4.

Bayesian consensus phylogeny of amastin surface proteins. The phylogram is represented by a consensus of 214 amastin sequences. The root was inferred using midpoint rooting. WAG was used as the substitution matrix for the protein alignment. Posterior probabilities exceeding 0.5 are shown in the branches. The tree topology suggests early branching of similar amastins shared by different species (blue). These branches were classified as Leishmania pre-speciation amastins, composed by α, β, and γ subfamily clades. We highlighted the terminal taxa (late branching or apomorphic) of species-specific δ-amastin clades of L. (L.) major (yellow), L. (V.) braziliensis (green), and L. (L.) infantum (purple). Complex-specific clades of L. (L.) amazonensis and L. (L.) mexicana amastin surface proteins are in red. The scale of the generated tree (see 0.4 bar) represents the number of substitutions per sequence position. The classification of amastin clades in subfamilies α, β, γ, and δ was based on the amastin phylogeny performed by Jackson et al. (2010).
3.3.2. Secreted Leishmania HSPs could interfere with native host interactomes
One of the most striking features of the L. (L.) mexicana complex is the development of giant PVs in infected macrophages that harbor amastigotes. We speculate that the formation of large PVs may be related to factors secreted by the parasite, the subversion of host native vesicular trafficking, and potentially the production/incorporation of parasitic components into PV membranes. L. (L.) amazonensis amastigotes interact with the internal membranes of PVs via their posterior poles.8 The posterior pole behaves like an adhesion site between the parasite and PV membranes, although no adhesion factors or junction components have been identified to date. De Souza Leão et al.82 suggested that in L. (L.) amazonensis-infected macrophages, the internalization and degradation of major histocompatibility complex (MHC) class II molecules by amastigotes occur through their posterior poles. This degradation could be performed by secreted components inserted into the PV membrane. The posterior pole also may be interpreted as part of a parasitic secretory pathway in which secreted proteins directly encounter the host cell cytosol, bypassing the acidic milieu of the PV. Once in the host cell cytosol, secreted factors may be transferred to the host cell nucleus and/or plasma membrane, affecting gene expression, cellular functions and metabolic processes. However, the classically described site for parasite exocytosis and endocytosis is the anterior pole where the flagellar pocket is located. The flagellar pocket faces the lumen of the PV, and most secreted Leishmania proteins, regardless of their association with exosomes, are expected to reach the acidic (pH 4.5–5.0) PV milieu from there.83
We hypothesized that Leishmania secreted factors could mimic mammalian factors, thus perturbing native host protein interactions. To identify possible interactions between parasitic and mammalian host factors, we constructed hybrid protein interaction networks in which human and mouse databases were compared against the list of proteins that are potentially secreted by L. (L.) amazonensis amastigotes. Our CDD–PFAM and OrthoMCL analyses identified nine conserved domains and three orthologous gene families that were exclusive to or expanded in L. (L.) amazonensis and/or L. (L.) mexicana and are also orthologous to human proteins (Supplementary data, Table S6). The CDD 143803, an HSP 70 domain, exists as six copies in L. (L.) amazonensis, four copies in L. (L.) mexicana, and three copies in the other species' genomes. We considered this as an expanded number of HSP70 domains in the L. (L.) mexicana complex. The six gene models in which these domains were identified (A42670, A6630, A68920, A73510, A30200, and A45910) were submitted to the TargetP and SecretomeP servers and gene models A30200 and A45910 were predicted for secretion. Additionally, A30200 and A45910 were similar to two L. (L.) mexicana proteins identified in a proteomic data analysis of L. (L.) mexicana amastigotes54 (LmxM.28.2770 and LmxM.34.4710, respectively; Supplementary data, Table S6). Although these genes are not exclusive to the L. (L.) mexicana complex, their similarity to two products from amastigote proteomic data is suggestive that, at least in L. (L.) amazonensis and L. (L.) mexicana, these products are expressed by the intracellular form of the parasite and could be secreted within host cells.
The A30200 and A45910 gene models are candidates for the construction of hybrid interactomes, given that they present a protein domain expanded in both L. (L.) amazonensis and L. (L.) mexicana (CDD 143803), are predicted to be secreted and are similar to proteins identified in the proteome of L. (L.) mexicana amastigotes. The A30200 and A45910 gene models present a considerable similarity with the mammalian hypoxia up-regulated protein 1 (HYOU1) and the HSP 70 kDa protein 5 (HSPA5), respectively. We created HYOU1 and HSPA5 interactome subsets to identify host components that could be affected by the secretion of both A30200 and A45910 gene products. This analysis led us to identify toll-like receptor (TLR) 9 and IL-6 as putative targets of the L. (L.) amazonensis A30200 and A45910 gene products (Fig. 5A and B). The HYOU1 and HSPA5 both directly interact with TLR9, a receptor implicated in the recognition of CpG DNA motifs and present in endolysosomal compartments where it is activated by proteolytic cleavage.84 TLR9 is preferentially expressed in the granulomas of human cutaneous leishmaniasis caused by L. (V.) braziliensis,85 and TLR9-deficient mice are more susceptible to L. (L.) major infection.86,87 Thus, TLR9 is implicated in the immune response against Leishmania. The interaction between host TLR9 and the putative secreted L. (L.) amazonensis HYOU1/HSPA5-mimic could block TLR9 function and favor intracellular establishment of the parasite. TLR9 also is implicated in the production of NO via NO synthase 2, tumor necrosis factor, IL-6, and IL-12B. The production of IL-6 is inhibited in dendritic cells differentiated from monocytes in the presence of L. (L.) amazonensis88 and is present at low levels in the sera of Chiclero's ulcer patients infected for 3–8 months.89 Linares et al.90 reported that in vitro infection with L. (L.) amazonensis amastigotes decreases NO production by macrophages stimulated with IFN-γ plus lipopolysaccharide. Thus, although hypothetical and genome based, our proposed interactome can be used to identify components implicated in the establishment of Leishmania infection of mammalian host cells. Moreover, the interactome provides a model for studying Leishmania-secreted proteins and their influence on important effectors of the host cell immune response.
Figure 5.
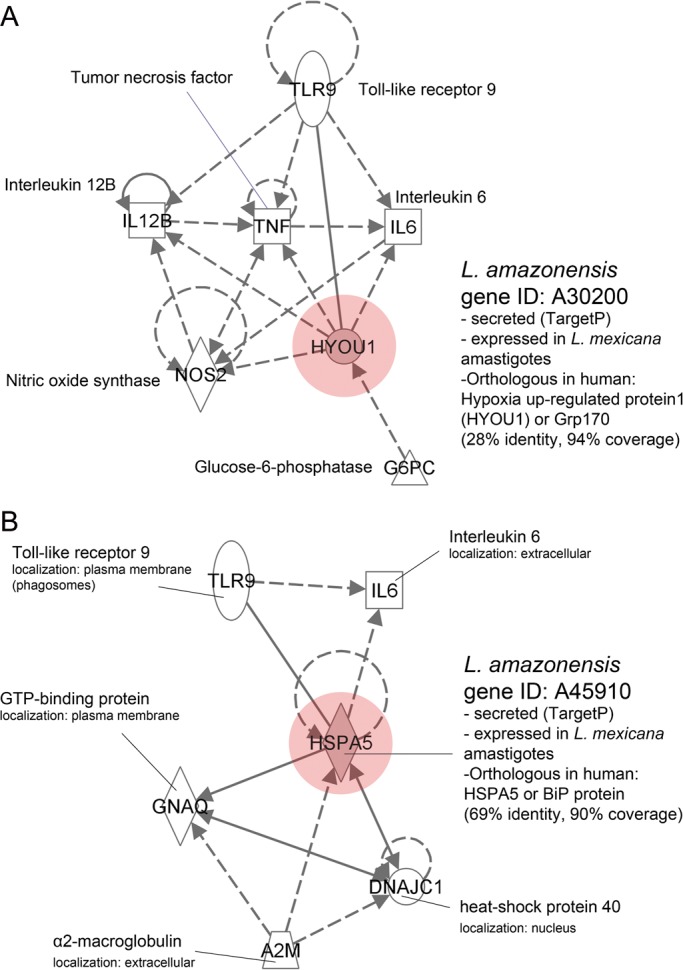
Interactomes of potentially secreted L. (L.) amazonensis [A30200 (A) and A45910 (B)] and mammalian immune cell proteins. The secreted parasite gene products are represented by red nodes in the interactome. The expression statuses of these parasite proteins during the amastigote stage were inferred using blastp with the proteomic database of L. (L.) mexicana amastigotes. The secreted components of L. (L.) amazonensis amastigotes share 28% identity and 94% coverage (A30200, A) and 69% identity and 90% coverage (A45910, B) with the mammalian HYOU1 and HSPA5 proteins, respectively. Both secreted components could directly interact with TLR9. We propose that orthologs of mammalian HYOU1 and HSPA5 are secreted by L. (L.) amazonensis amastigotes, interfering with host cell functions such as signaling and the production of NO and ILs. Arrows represent direct interactions and dashed arrows represent indirect interactions. The interactome was built using Ingenuity software, considering only proteins expressed in human and mouse immune cells and considering experimentally identified protein–protein interactions.
3.4. Conclusions
We present the genome of the protozoan L. (L.) amazonensis together with functional annotations and extended analyses focused on host–parasite interactions. We examined the genome sequences of L. (L.) amazonensis and L. (L.) mexicana for potentially expressed genes at expanded copy numbers. Confirming that a few Leishmania species-specific genes may exist despite striking conservation at the gene level, we report conserved domains, orthologous gene families, and amastin surface proteins unique to L. (L.) amazonensis and L. (L.) mexicana. Additionally, we propose an innovative approach to interactome analysis that emphasizes the role of parasite secreted proteins in host interaction networks.
4. Availability
The Leishmania (Leishmania) amazonensis Genome Database is available at the URL http://www.lge.ibi.unicamp.br/leishmania. This Whole Genome Shotgun project was deposited at DDBJ/EMBL/GenBank under the accession APNT00000000 (SUBID SUB120161, BioProject PRJNA173202). The version described in this paper is the first version, APNT01000000. While revising this manuscript, we realized that another Brazilian group from Instituto Oswaldo Cruz (IOC)—Fiocruz, Rio de Janeiro, is sequencing the genome of L. (L.) amazonensis.
Supplementary Data
Supplementary Data are available at www.dnaresearch.oxfordjournals.org.
Funding
The authors acknowledge the financial support of Fundação de Amparo à Pesquisa do Estado de São Paulo, FAPESP (Jovem Pesquisador 07/50551-2). F.R. is recipient of a FAPESP post-doctorate fellowship (10/19335-4). D.B., J.F.S., R.A.M., M.S.B., G.A.P. are recipients of a Conselho Nacional de Desenvolvimento Científico e Tecnológico, CNPq, fellowship.
Supplementary Material
Acknowledgements
F.R. and D.B. would like to thank Dr Michel Rabinovitch for stimulating advice and Dr Andrew Jackson for providing amastin alignments. The authors also thank Dr Angela Kaysel Cruz and Dr Colin Bowles for kindly revising the manuscript and response to reviewers and BioMed Proofreading (http://www.biomedproofreading.com) for English editing services.
Footnotes
Edited by Naotake Ogasawara
References
- 1.WHO. Geneva: WHO Press; 2010. Control of the Leishmaniasis. WHO Technical Report Series. [Google Scholar]
- 2.Lainson R., Shaw J.J. 1987. The leishmaniases in biology and medicine. Evolution, classification and geographical distribution.
- 3.Bates P.A. Transmission of Leishmania metacyclic promastigotes by phlebotomine sand flies. Int. J. Parasitol. 2007;37:1097–106. doi: 10.1016/j.ijpara.2007.04.003. doi:10.1016/j.ijpara.2007.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dedet J.P., Pratlong F., Lanotte G., Ravel C. Cutaneous leishmaniasis. The parasite. Clin. Dermatol. 1999;17:261–8. doi: 10.1016/s0738-081x(99)00044-9. doi:10.1016/S0738-081X(99)00044-9. [DOI] [PubMed] [Google Scholar]
- 5.Murray H.W., Berman J.D., Davies C.R., Saravia N.G. Advances in leishmaniasis. Lancet. 2005;366:1561–77. doi: 10.1016/S0140-6736(05)67629-5. doi:10.1016/S0140-6736(05)67629-5. [DOI] [PubMed] [Google Scholar]
- 6.Camara Coelho L.I., Paes M., Guerra J.A., et al. Characterization of Leishmania spp. causing cutaneous leishmaniasis in Manaus, Amazonas, Brazil. Parasitol. Res. 2011;108:671–7. doi: 10.1007/s00436-010-2139-9. doi:10.1007/s00436-010-2139-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Silveira F.T., Lainson R., Corbett C.E. Further observations on clinical, histopathological, and immunological features of borderline disseminated cutaneous leishmaniasis caused by Leishmania (Leishmania) amazonensis. Mem Inst Oswaldo Cruz. 2005;100:525–34. doi: 10.1590/s0074-02762005000500013. doi:10.1590/S0074-02762005000500013. [DOI] [PubMed] [Google Scholar]
- 8.Real F., Mortara R.A. The diverse and dynamic nature of Leishmania parasitophorous vacuoles studied by multidimensional imaging. PLoS Negl. Trop. Dis. 2012;6:e1518. doi: 10.1371/journal.pntd.0001518. doi:10.1371/journal.pntd.0001518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Real F., Pouchelet M., Rabinovitch M. Leishmania (L.) amazonensis: fusion between parasitophorous vacuoles in infected bone-marrow derived mouse macrophages. Exp. Parasitol. 2008;119:15–23. doi: 10.1016/j.exppara.2007.12.013. doi:10.1016/j.exppara.2007.12.013. [DOI] [PubMed] [Google Scholar]
- 10.Alpuche-Aranda C.M., Racoosin E.L., Swanson J.A., Miller S.I. Salmonella stimulate macrophage macropinocytosis and persist within spacious phagosomes. J. Exp. Med. 1994;179:601–8. doi: 10.1084/jem.179.2.601. doi:10.1084/jem.179.2.601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Real F., Mortara R.A., Rabinovitch M. Fusion between Leishmania amazonensis and Leishmania major parasitophorous vacuoles: live imaging of coinfected macrophages. PLoS Negl. Trop. Dis. 2010;4:e905. doi: 10.1371/journal.pntd.0000905. doi:10.1371/journal.pntd.0000905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ndjamen B., Kang B.H., Hatsuzawa K., Kima P.E. Leishmania parasitophorous vacuoles interact continuously with the host cell's endoplasmic reticulum; parasitophorous vacuoles are hybrid compartments. Cell Microbiol. 2010;12:1480–94. doi: 10.1111/j.1462-5822.2010.01483.x. doi:10.1111/j.1462-5822.2010.01483.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Clayton C., Shapira M. Post-transcriptional regulation of gene expression in trypanosomes and leishmanias. Mol. Biochem. Parasitol. 2007;156:93–101. doi: 10.1016/j.molbiopara.2007.07.007. doi:10.1016/j.molbiopara.2007.07.007. [DOI] [PubMed] [Google Scholar]
- 14.Martinez-Calvillo S., Yan S., Nguyen D., Fox M., Stuart K., Myler P.J. Transcription of Leishmania major Friedlin chromosome 1 initiates in both directions within a single region. Mol. Cell. 2003;11:1291–9. doi: 10.1016/s1097-2765(03)00143-6. doi:10.1016/S1097-2765(03)00143-6. [DOI] [PubMed] [Google Scholar]
- 15.Haile S., Papadopoulou B. Developmental regulation of gene expression in trypanosomatid parasitic protozoa. Curr. Opin. Microbiol. 2007;10:569–77. doi: 10.1016/j.mib.2007.10.001. doi:10.1016/j.mib.2007.10.001. [DOI] [PubMed] [Google Scholar]
- 16.Martinez-Calvillo S., Vizuet-de-Rueda J.C., Florencio-Martinez L.E., Manning-Cela R.G., Figueroa-Angulo E.E. Gene expression in trypanosomatid parasites. J. Biomed. Biotechnol. 2010;2010:525241. doi: 10.1155/2010/525241. doi:10.1155/2010/525241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wincker P., Ravel C., Blaineau C., et al. The Leishmania genome comprises 36 chromosomes conserved across widely divergent human pathogenic species. Nucleic Acids Res. 1996;24:1688–94. doi: 10.1093/nar/24.9.1688. doi:10.1093/nar/24.9.1688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Britto C., Ravel C., Bastien P., et al. Conserved linkage groups associated with large-scale chromosomal rearrangements between Old World and New World Leishmania genomes. Gene. 1998;222:107–17. doi: 10.1016/s0378-1119(98)00472-7. doi:10.1016/S0378-1119(98)00472-7. [DOI] [PubMed] [Google Scholar]
- 19.Peacock C.S., Seeger K., Harris D., et al. Comparative genomic analysis of three Leishmania species that cause diverse human disease. Nat. Genet. 2007;39:839–47. doi: 10.1038/ng2053. doi:10.1038/ng2053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Raymond F., Boisvert S., Roy G., et al. Genome sequencing of the lizard parasite Leishmania tarentolae reveals loss of genes associated to the intracellular stage of human pathogenic species. Nucleic Acids Res. 2012;40:1131–47. doi: 10.1093/nar/gkr834. doi:10.1093/nar/gkr834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rovai L., Tripp C., Stuart K., Simpson L. Recurrent polymorphisms in small chromosomes of Leishmania tarentolae after nutrient stress or subcloning. Mol. Biochem. Parasitol. 1992;50:115–25. doi: 10.1016/0166-6851(92)90249-j. doi:10.1016/0166-6851(92)90249-J. [DOI] [PubMed] [Google Scholar]
- 22.Ivens A.C., Peacock C.S., Worthey E.A., et al. The genome of the kinetoplastid parasite, Leishmania major. Science. 2005;309:436–42. doi: 10.1126/science.1112680. doi:10.1126/science.1112680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Downing T., Imamura H., Decuypere S., et al. Whole genome sequencing of multiple Leishmania donovani clinical isolates provides insights into population structure and mechanisms of drug resistance. Genome Res. 2011;21:2143–56. doi: 10.1101/gr.123430.111. doi:10.1101/gr.123430.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Rogers M.B., Hilley J.D., Dickens N.J., et al. Chromosome and gene copy number variation allow major structural change between species and strains of Leishmania. Genome Res. 2011;21:2129–42. doi: 10.1101/gr.122945.111. doi:10.1101/gr.122945.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Smith D.F., Peacock C.S., Cruz A.K. Comparative genomics: from genotype to disease phenotype in the leishmaniases. Int. J. Parasitol. 2007;37:1173–86. doi: 10.1016/j.ijpara.2007.05.015. doi:10.1016/j.ijpara.2007.05.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lye L.F., Owens K., Shi H., et al. Retention and loss of RNA interference pathways in trypanosomatid protozoans. PLoS Pathog. 2010;6:e1001161. doi: 10.1371/journal.ppat.1001161. doi:10.1371/journal.ppat.1001161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Messing J., Crea R., Seeburg P.H. A system for shotgun DNA sequencing. Nucleic Acids Res. 1981;9:309–21. doi: 10.1093/nar/9.2.309. doi:10.1093/nar/9.2.309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zerbino D.R., Birney E. Velvet: algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008;18:821–9. doi: 10.1101/gr.074492.107. doi:10.1101/gr.074492.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Quinn N.L., Levenkova N., Chow W., et al. Assessing the feasibility of GS FLX Pyrosequencing for sequencing the Atlantic salmon genome. BMC Genomics. 2008;9:404. doi: 10.1186/1471-2164-9-404. doi:10.1186/1471-2164-9-404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sommer D.D., Delcher A.L., Salzberg S.L., Pop M. Minimus: a fast, lightweight genome assembler. BMC Bioinformatics. 2007;8:64. doi: 10.1186/1471-2105-8-64. doi:10.1186/1471-2105-8-64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Pop M., Kosack D.S., Salzberg S.L. Hierarchical scaffolding with Bambus. Genome Res. 2004;14:149–59. doi: 10.1101/gr.1536204. doi:10.1101/gr.1536204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Slater G.S., Birney E. Automated generation of heuristics for biological sequence comparison. BMC Bioinformatics. 2005;6:31. doi: 10.1186/1471-2105-6-31. doi:10.1186/1471-2105-6-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Salzberg S.L., Delcher A.L., Kasif S., White O. Microbial gene identification using interpolated Markov models. Nucleic Acids Res. 1998;26:544–8. doi: 10.1093/nar/26.2.544. doi:10.1093/nar/26.2.544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lomsadze A., Ter-Hovhannisyan V., Chernoff Y.O., Borodovsky M. Gene identification in novel eukaryotic genomes by self-training algorithm. Nucleic Acids Res. 2005;33:6494–506. doi: 10.1093/nar/gki937. doi:10.1093/nar/gki937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Haas B.J., Salzberg S.L., Zhu W., et al. Automated eukaryotic gene structure annotation using EVidenceModeler and the program to assemble spliced alignments. Genome Biol. 2008;9:R7. doi: 10.1186/gb-2008-9-1-r7. doi:10.1186/gb-2008-9-1-r7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Koski L.B., Gray M.W., Lang B.F., Burger G. AutoFACT: an automatic functional annotation and classification tool. BMC Bioinformatics. 2005;6:151. doi: 10.1186/1471-2105-6-151. doi:10.1186/1471-2105-6-151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Suzek B.E., Huang H., McGarvey P., Mazumder R., Wu C.H. UniRef: comprehensive and non-redundant UniProt reference clusters. Bioinformatics. 2007;23:1282–8. doi: 10.1093/bioinformatics/btm098. doi:10.1093/bioinformatics/btm098. [DOI] [PubMed] [Google Scholar]
- 38.Marchler-Bauer A., Bryant S.H. CD-Search: protein domain annotations on the fly. Nucleic Acids Res. 2004;32:W327–31. doi: 10.1093/nar/gkh454. doi:10.1093/nar/gkh454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bateman A., Birney E., Cerruti L., et al. The Pfam protein families database. Nucleic Acids Res. 2002;30:276–80. doi: 10.1093/nar/30.1.276. doi:10.1093/nar/30.1.276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kanehisa M., Goto S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000;28:27–30. doi: 10.1093/nar/28.1.27. doi:10.1093/nar/28.1.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Chen F., Mackey A.J., Stoeckert C.J., Jr, Roos D.S. OrthoMCL-DB: querying a comprehensive multi-species collection of ortholog groups. Nucleic Acids Res. 2006;34:D363–8. doi: 10.1093/nar/gkj123. doi:10.1093/nar/gkj123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chen F., Mackey A.J., Vermunt J.K., Roos D.S. Assessing performance of orthology detection strategies applied to eukaryotic genomes. PloS One. 2007;2:e383. doi: 10.1371/journal.pone.0000383. doi:10.1371/journal.pone.0000383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Quinlan A.R., Hall I.M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics. 2010;26:841–2. doi: 10.1093/bioinformatics/btq033. doi:10.1093/bioinformatics/btq033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sharp P.M., Li W.H. The codon adaptation index—a measure of directional synonymous codon usage bias, and its potential applications. Nucleic Acids Res. 1987;15:1281–95. doi: 10.1093/nar/15.3.1281. doi:10.1093/nar/15.3.1281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sharp P.M., Tuohy T.M., Mosurski K.R. Codon usage in yeast: cluster analysis clearly differentiates highly and lowly expressed genes. Nucleic Acids Res. 1986;14:5125–43. doi: 10.1093/nar/14.13.5125. doi:10.1093/nar/14.13.5125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Comeron J.M., Aguade M. An evaluation of measures of synonymous codon usage bias. J. Mol. Evol. 1998;47:268–74. doi: 10.1007/pl00006384. doi:10.1007/PL00006384. [DOI] [PubMed] [Google Scholar]
- 47.Aslett M., Aurrecoechea C., Berriman M., et al. TriTrypDB: a functional genomic resource for the Trypanosomatidae. Nucleic Acids Res. 2010;38:D457–62. doi: 10.1093/nar/gkp851. doi:10.1093/nar/gkp851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Drummond A.J., Ashton B., Buxton S., et al. 2011. Geneious v5.6.3http://www.geneious.com/ (June 2012, date last accessed)
- 49.Edgar R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32:1792–7. doi: 10.1093/nar/gkh340. doi:10.1093/nar/gkh340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ronquist F., Huelsenbeck J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19:1572–4. doi: 10.1093/bioinformatics/btg180. doi:10.1093/bioinformatics/btg180. [DOI] [PubMed] [Google Scholar]
- 51.Whelan S., Goldman N. A general empirical model of protein evolution derived from multiple protein families using a maximum-likelihood approach. Mol. Biol. Evol. 2001;18:691–9. doi: 10.1093/oxfordjournals.molbev.a003851. doi:10.1093/oxfordjournals.molbev.a003851. [DOI] [PubMed] [Google Scholar]
- 52.Emanuelsson O., Brunak S., von Heijne G., Nielsen H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007;2:953–71. doi: 10.1038/nprot.2007.131. doi:10.1038/nprot.2007.131. [DOI] [PubMed] [Google Scholar]
- 53.Bendtsen J.D., Jensen L.J., Blom N., Von Heijne G., Brunak S. Feature-based prediction of non-classical and leaderless protein secretion. Protein Eng Des Sel: PEDS. 2004;17:349–56. doi: 10.1093/protein/gzh037. doi:10.1093/protein/gzh037. [DOI] [PubMed] [Google Scholar]
- 54.Paape D., Barrios-Llerena M.E., Le Bihan T., Mackay L., Aebischer T. Gel free analysis of the proteome of intracellular Leishmania mexicana. Mol. Biochem. Parasitol. 2010;169:108–14. doi: 10.1016/j.molbiopara.2009.10.009. doi:10.1016/j.molbiopara.2009.10.009. [DOI] [PubMed] [Google Scholar]
- 55.Lowe T.M., Eddy S.R. tRNAscan-SE: a program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997;25:955–64. doi: 10.1093/nar/25.5.955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Castillo-Ramirez S., Vazquez-Castellanos J.F., Gonzalez V., Cevallos M.A. Horizontal gene transfer and diverse functional constrains within a common replication-partitioning system in Alphaproteobacteria: the repABC operon. BMC Genomics. 2009;10:536. doi: 10.1186/1471-2164-10-536. doi:10.1186/1471-2164-10-536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Bastien P., Blaineau C., Pages M. Leishmania: sex, lies and karyotype. Parasitol. Today. 1992;8:174–7. doi: 10.1016/0169-4758(92)90016-u. doi:10.1016/0169-4758(92)90016-U. [DOI] [PubMed] [Google Scholar]
- 58.Mannaert A., Downing T., Imamura H., Dujardin J.C. Adaptive mechanisms in pathogens: universal aneuploidy in Leishmania. Trends Parasitol. 2012;28:370–6. doi: 10.1016/j.pt.2012.06.003. doi:10.1016/j.pt.2012.06.003. [DOI] [PubMed] [Google Scholar]
- 59.Sterkers Y., Lachaud L., Bourgeois N., Crobu L., Bastien P., Pages M. Novel insights into genome plasticity in Eukaryotes: mosaic aneuploidy in Leishmania. Mol. Microbiol. 2012;86:15–23. doi: 10.1111/j.1365-2958.2012.08185.x. doi:10.1111/j.1365-2958.2012.08185.x. [DOI] [PubMed] [Google Scholar]
- 60.Ning Z., Cox A.J., Mullikin J.C. SSAHA: a fast search method for large DNA databases. Genome Res. 2001;11:1725–9. doi: 10.1101/gr.194201. doi:10.1101/gr.194201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Gentil L.G., Lasakosvitsch F., Silveira J.F., Santos M.R., Barbieri C.L. Analysis and chromosomal mapping of Leishmania (Leishmania) amazonensis amastigote expressed sequence tags. Mem Inst Oswaldo Cruz. 2007;102:707–11. doi: 10.1590/s0074-02762007005000065. doi:10.1590/S0074-02762007005000065. [DOI] [PubMed] [Google Scholar]
- 62.Hutson S. Structure and function of branched chain aminotransferases. Prog Nucleic Acid Res. Mol. Biol. 2001;70:175–206. doi: 10.1016/s0079-6603(01)70017-7. [DOI] [PubMed] [Google Scholar]
- 63.Ginger M.L., Chance M.L., Goad L.J. Elucidation of carbon sources used for the biosynthesis of fatty acids and sterols in the trypanosomatid Leishmania mexicana. Biochem. J. 1999;342:397–405. doi:10.1042/0264-6021:3420397. [PMC free article] [PubMed] [Google Scholar]
- 64.Arruda D.C., D'Alexandri F.L., Katzin A.M., Uliana S.R. Leishmania amazonensis: biosynthesis of polyprenols of 9 isoprene units by amastigotes. Exp. Parasitol. 2008;118:624–8. doi: 10.1016/j.exppara.2007.11.009. doi:10.1016/j.exppara.2007.11.009. [DOI] [PubMed] [Google Scholar]
- 65.Neubert T.A., Gottlieb M. An inducible 3′-nucleotidase/nuclease from the trypanosomatid Crithidia luciliae. Purification and characterization. J. Biol. Chem. 1990;265:7236–42. [PubMed] [Google Scholar]
- 66.Paletta-Silva R., Vieira D.P., Vieira-Bernardo R., et al. Leishmania amazonensis: characterization of an ecto-3′-nucleotidase activity and its possible role in virulence. Exp. Parasitol. 2011;129:277–83. doi: 10.1016/j.exppara.2011.07.014. doi:10.1016/j.exppara.2011.07.014. [DOI] [PubMed] [Google Scholar]
- 67.Holmgren A., Lu J. Thioredoxin and thioredoxin reductase: current research with special reference to human disease. Biochem. Biophys. Res. Commun. 2010;396:120–4. doi: 10.1016/j.bbrc.2010.03.083. doi:10.1016/j.bbrc.2010.03.083. [DOI] [PubMed] [Google Scholar]
- 68.Scott P., Sher A. A spectrum in the susceptibility of leishmanial strains to intracellular killing by murine macrophages. J. Immunol. 1986;136:1461–6. [PubMed] [Google Scholar]
- 69.Krauth-Siegel R.L., Comini M.A. Redox control in trypanosomatids, parasitic protozoa with trypanothione-based thiol metabolism. Biochim. Biophys. Acta. 2008;1780:1236–48. doi: 10.1016/j.bbagen.2008.03.006. doi:10.1016/j.bbagen.2008.03.006. [DOI] [PubMed] [Google Scholar]
- 70.de Souza Carmo E.V., Katz S., Barbieri C.L. Neutrophils reduce the parasite burden in Leishmania (Leishmania) amazonensis-infected macrophages. PloS One. 2010;5:e13815. doi: 10.1371/journal.pone.0013815. doi:10.1371/journal.pone.0013815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Asato Y., Oshiro M., Myint C.K., et al. Phylogenic analysis of the genus Leishmania by cytochrome b gene sequencing. Exp. Parasitol. 2009;121:352–61. doi: 10.1016/j.exppara.2008.12.013. doi:10.1016/j.exppara.2008.12.013. [DOI] [PubMed] [Google Scholar]
- 72.Fraga J., Montalvo A.M., De Doncker S., Dujardin J.C., Van der Auwera G. Phylogeny of Leishmania species based on the heat-shock protein 70 gene. Infect. Genet. Evol. 2010;10:238–45. doi: 10.1016/j.meegid.2009.11.007. doi:10.1016/j.meegid.2009.11.007. [DOI] [PubMed] [Google Scholar]
- 73.Rochette A., McNicoll F., Girard J., et al. Characterization and developmental gene regulation of a large gene family encoding amastin surface proteins in Leishmania spp. Mol. Biochem. Parasitol. 2005;140:205–20. doi: 10.1016/j.molbiopara.2005.01.006. doi:10.1016/j.molbiopara.2005.01.006. [DOI] [PubMed] [Google Scholar]
- 74.Jackson A.P. The evolution of amastin surface glycoproteins in trypanosomatid parasites. Mol. Biol. Evol. 2010;27:33–45. doi: 10.1093/molbev/msp214. doi:10.1093/molbev/msp214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Cruz M.C., Souza-Melo N., da Silva C.V., et al. Trypanosoma cruzi: role of delta-Amastin on extracellular amastigote cell invasion and differentiation. PloS One. 2012;7:e51804. doi: 10.1371/journal.pone.0051804. doi:10.1371/journal.pone.0051804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Stober C.B., Lange U.G., Roberts M.T., et al. From genome to vaccines for leishmaniasis: screening 100 novel vaccine candidates against murine Leishmania major infection. Vaccine. 2006;24:2602–16. doi: 10.1016/j.vaccine.2005.12.012. doi:10.1016/j.vaccine.2005.12.012. [DOI] [PubMed] [Google Scholar]
- 77.Rafati S., Hassani N., Taslimi Y., Movassagh H., Rochette A., Papadopoulou B. Amastin peptide-binding antibodies as biomarkers of active human visceral leishmaniasis. Clin. Vaccine Immunol. 2006;13:1104–10. doi: 10.1128/CVI.00188-06. doi:10.1128/CVI.00188-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Salotra P., Duncan R.C., Singh R., Subba Raju B.V., Sreenivas G., Nakhasi H.L. Upregulation of surface proteins in Leishmania donovani isolated from patients of post kala-azar dermal leishmaniasis. Microbes Infect. 2006;8:637–44. doi: 10.1016/j.micinf.2005.08.018. doi:10.1016/j.micinf.2005.08.018. [DOI] [PubMed] [Google Scholar]
- 79.Rochette A., Raymond F., Ubeda J.M., et al. Genome-wide gene expression profiling analysis of Leishmania major and Leishmania infantum developmental stages reveals substantial differences between the two species. BMC Genomics. 2008;9:255. doi: 10.1186/1471-2164-9-255. doi:10.1186/1471-2164-9-255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Azizi H., Hassani K., Taslimi Y., Najafabadi H.S., Papadopoulou B., Rafati S. Searching for virulence factors in the non-pathogenic parasite to humans Leishmania tarentolae. Parasitology. 2009;136:723–35. doi: 10.1017/S0031182009005873. doi:10.1017/S0031182009005873. [DOI] [PubMed] [Google Scholar]
- 81.Naderer T., McConville M.J. The Leishmania-macrophage interaction: a metabolic perspective. Cell Microbiol. 2008;10:301–8. doi: 10.1111/j.1462-5822.2007.01096.x. doi:10.1111/j.1462-5822.2007.01096.x. [DOI] [PubMed] [Google Scholar]
- 82.De Souza Leao S., Lang T., Prina E., Hellio R., Antoine J.C. Intracellular Leishmania amazonensis amastigotes internalize and degrade MHC class II molecules of their host cells. J. Cell Sci. 1995;108:3219–31. doi: 10.1242/jcs.108.10.3219. [DOI] [PubMed] [Google Scholar]
- 83.Silverman J.M., Chan S.K., Robinson D.P., et al. Proteomic analysis of the secretome of Leishmania donovani. Genome Biol. 2008;9:R35. doi: 10.1186/gb-2008-9-2-r35. doi:10.1186/gb-2008-9-2-r35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Mouchess M.L., Arpaia N., Souza G., et al. Transmembrane mutations in Toll-like receptor 9 bypass the requirement for ectodomain proteolysis and induce fatal inflammation. Immunity. 2011;35:721–32. doi: 10.1016/j.immuni.2011.10.009. doi:10.1016/j.immuni.2011.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Tuon F.F., Fernandes E.R., Pagliari C., Duarte M.I., Amato V.S. The expression of TLR9 in human cutaneous leishmaniasis is associated with granuloma. Parasite Immunol. 2010;32:769–72. doi: 10.1111/j.1365-3024.2010.01243.x. doi:10.1111/j.1365-3024.2010.01243.x. [DOI] [PubMed] [Google Scholar]
- 86.Abou Fakher F.H., Rachinel N., Klimczak M., Louis J., Doyen N. TLR9-dependent activation of dendritic cells by DNA from Leishmania major favors Th1 cell development and the resolution of lesions. J. Immunol. 2009;182:1386–96. doi: 10.4049/jimmunol.182.3.1386. [DOI] [PubMed] [Google Scholar]
- 87.Carvalho L.P., Petritus P.M., Trochtenberg A.L., et al. Lymph node hypertrophy following Leishmania major infection is dependent on TLR9. J. Immunol. 2012;188:1394–401. doi: 10.4049/jimmunol.1101018. doi:10.4049/jimmunol.1101018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Favali C., Tavares N., Clarencio J., Barral A., Barral-Netto M., Brodskyn C. Leishmania amazonensis infection impairs differentiation and function of human dendritic cells. J. Leukoc. Biol. 2007;82:1401–6. doi: 10.1189/jlb.0307187. doi:10.1189/jlb.0307187. [DOI] [PubMed] [Google Scholar]
- 89.Lezama-Davila C.M., Isaac-Marquez A.P. Systemic cytokine response in humans with chiclero's ulcers. Parasitol Res. 2006;99:546–53. doi: 10.1007/s00436-006-0203-2. doi:10.1007/s00436-006-0203-2. [DOI] [PubMed] [Google Scholar]
- 90.Linares E., Augusto O., Barao S.C., Giorgio S. Leishmania amazonensis infection does not inhibit systemic nitric oxide levels elicited by lipopolysaccharide in vivo. J. Parasitol. 2000;86:78–82. doi: 10.1645/0022-3395(2000)086[0078:LAIDNI]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}