

Abstract
Glycosaminoglycans (GAGs) participate in a variety of processes in the kidney, and evidence suggests that gender-related hormones participate in renal function. The aim of this study was to analyze the relationship of GAGs, gender, and proteinuria in male and female rats with chronic renal failure (CRF). GAGs were analyzed in total kidney tissue and 24-h urine of castrated (c), male (M), and female (F) Wistar control (C) rats (CM, CMc, CF, CFc) and after 30 days of CRF induced by 5/6 nephrectomy (CRFM, CRFMc, CRFF, CRFFc). Total GAG quantification and composition were determined using agarose and polyacrylamide gel electrophoresis, respectively. Renal GAGs were higher in CF compared to CM. CRFM presented an increase in renal GAGs, heparan sulfate (HS), and proteinuria, while castration reduced these parameters. However, CRFF and CRFFc groups showed a decrease in renal GAGs concomitant with an increase in proteinuria. Our results suggest that, in CRFM, sex hormones quantitatively alter GAGs, mainly HS, and possibly the glomerular filtration barrier, leading to proteinuria. The lack of this response in CRFMc, where HS did not increase, corroborates this theory. This pattern was not observed in females. Further studies of CRF are needed to clarify gender-dependent differences in HS synthesis.
Keywords: Castration, Chronic renal failure, Gender, Glycosaminoglycans, Proteinuria
Introduction
Glycosaminoglycans (GAGs) are anionic polysaccharides composed of repeating disaccharide units of amino sugar and hexuronic acid. Six types of GAGs are described in mammalian tissues: chondroitin sulfate (CS), dermatan sulfate (DS), keratan sulfate, heparan sulfate (HS), heparin, and hyaluronic acid. Their characteristics include the presence of anionic charges due to the sulfate and carboxyl groups of the hexuronic acid. Exceptions are hyaluronic acid, which is not sulfated, and keratan sulfate, which contains D-galactose instead of hexuronic acid. In tissues, besides hyaluronic acid, all occurring GAGs are covalently attached to proteins, forming the proteoglycans; they are widely distributed on the cell surface and in intracellular granules, and are essential components of the extracellular matrix (ECM) (1).
GAGs have been implicated in several pathophysiological processes: tissue organization, cell growth and maturation, and modulation of the action of growth factors and collagen fibrillogenesis, among others. In the kidney, HS is the most abundant GAG, mainly present in the glomerular ECM, which is formed by the mesangium and basement membrane of the glomerular capillary. Besides its ability to bind to other components of the glomerular ECM, especially laminin and collagen, and so helping to maintain the molecular architecture of this matrix, HS is also suggested to play an important role in glomerular permselectivity (2). Changes in this selectivity may be responsible for proteinuria (PTN), which has been considered the main marker of glomerular injury and is directly correlated to loss of renal function (3- 5).
Moreover, glomerular ECM fluidity is a consequence of the negative charge of HS, which retains sodium ions and water molecules, thus allowing the anchoring of growth factors and cytokines, which ultimately modulates mesangial expansion. These mechanisms indicate early steps in the progression of chronic renal failure (CRF) that, together with PTN, precede glomerulosclerosis (4, 6). GAGs excreted in the urine (uGAGs) have been used as a marker of the severity of glomerular diseases because they may reflect biochemical changes in the glomerular ECM (7, 8). However, the precise role of GAGs in the mechanisms that compromise the glomerular filtration barrier is not totally clear.
Additionally, some studies point to sex hormones as being responsible for a faster progression of CRF in males (9- 13). However, studies focusing on GAG composition and content in the kidney and urine, when there is deprivation of sex hormones, are lacking. The aim of the present study was to evaluate the composition of GAGs in the kidney and urine of male and female rats with CRF undergoing castration, in order to analyze possible gender-dependent changes between GAGs and renal function. We found quantitative differences in HS and PTN related to sex hormones in the CRF model.
Material and Methods
Experimental model
Male (M) and female (F) Wistar rats were randomly divided into castrated (c) and noncastrated (control, C) groups at 5 weeks of age (Figure 1). After castration, animals were separated into 4 groups: castrated (CMc, n=5 and CFc, n=5) and controls (CM, n=5 and CF, n=4). At 7 weeks of age, animals randomly underwent 5/6 nephrectomy (CRF) and were again separated in CRF castrated (CRFMc, n=5 and CRFFc, n=6) and CRF controls (CRFM, n=5 and CRFF, n=6). Eight animals were given a sham nephrectomy (CM Sham, n=4 and CF Sham, n=4).
Figure 1. Schematic representation of the study design combining castration and CRF. C: control; c: castration; CRF: chronic renal failure; F: female rat; M: male rat; dashed line: no surgical procedure.
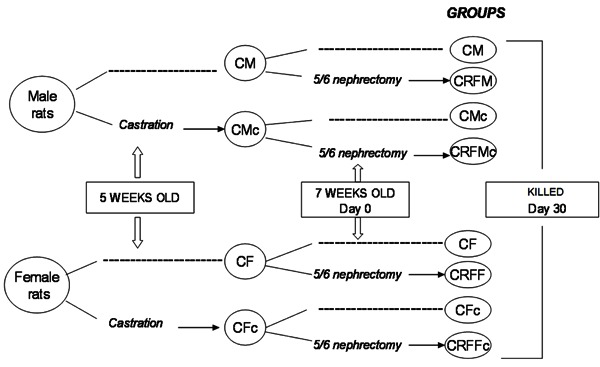
Surgical procedures were performed with animals anesthetized intraperitoneally with sodium thiopental (50 mg/kg body weight). In male rats, castration was performed by removal of the testicles; in female rats, the ovaries were removed through a bilateral lumbar incision. Reduction of total renal mass was performed by making an abdominal incision, isolating the left renal artery, ligating two or three branches and performing a right nephrectomy through a lumbar incision. This procedure was performed on day 0, and the animals were maintained on a standard rat chow (57% carbohydrate, 22% protein, and 4% fat; Nuvilab¯ CR1 Nuvital Nutrientes S/A, Brazil) and water ad libitum until the end of the study (after 30 days). All procedures were carried out in accordance with the conventional guidelines for the Care and Use of Laboratory Animals (US National Institutes of Health 85-23, revised 1996). Experimental protocols were approved by the Committee for the Use and Care of Experimental Animals of the Instituto de Biologia, Universidade do Estado do Rio de Janeiro, Brazil.
PTN and glomerular filtration rate
At day 30, animals were placed in metabolic cages (Nalgene, USA), where they had previously been trained, and received only water ad libitum, prior to collecting 24-h urine. Two samples were collected from each animal: one for measurement of urinary creatinine and PTN, and the other for extraction of uGAGs. Urine volumes were measured and urine stored at -20°C for subsequent evaluation. At the end of the collection, animals were anesthetized and killed by exsanguination. A blood sample was taken for creatinine determination. The kidneys were removed, weighed, and preserved in acetone. PTN was measured by the sulfosalicylic method (14). Urinary and serum creatinine were determined by the kinetic modified Jaffé method in the Central Laboratory of the Pedro Ernesto Hospital. Renal function was determined by creatinine clearance (CreCl) corrected to 100 g body weight (CreCl/100 g) as follows: CreCl = [(CreU×VolU/CreS)×100]/BW (mL·min-1·100 g-1), where CreU is urinary creatinine (mg/dL), VolU is urine volume in 24 h (mL/min), CreS is serum creatinine (mg/dL), and BW is body weight (g).
Isolation and purification of total GAGs
These methods have been described previously (15). The kidneys were rehydrated in ∼5 mL 0.1 M sodium acetate buffer, containing 5 mM cysteine and 5 mM EDTA, pH 5.0, at 4°C for 24 h. Thereafter, total renal GAGs (rGAGs) were isolated by proteolysis with papain followed by subsequent cetylpiridinium and ethanol precipitation. All pellets were previously treated with 2 mg/mL deoxyribonuclease (37°C/24 h). The final pellet was dried under vacuum and dissolved in 2.0 mL distilled water.
After a 1/2 dilution, urine was filtered and the pH adjusted to 6.0. The precipitation of uGAGs was performed by adding 5% N-cethyl-N,N,N-trimethyl-ammonium bromide (overnight; Sigma, USA) and absolute ethanol. After centrifugation, the pellet was kept at room temperature until completely dry. It was then eluted in 10% sodium acetate, adjusted to pH 9.0, and mixed vigorously. After centrifugation, the supernatant was collected and the pH adjusted to 6.0. A second precipitation was performed by adding 1.5 volumes of absolute ethanol at 0°C (overnight). After centrifugation, the final pellet containing the uGAGs was dried under vacuum.
Quantification of total GAGs
The total GAG samples were dissolved in ∼2.0 mL distilled water and assayed by the metachromasia produced by hexuronic acid using the carbazole reaction (16). Under these conditions more than 90% of total GAGs were recovered. Concentrations of rGAGs were expressed in micrograms per milligram dry tissue and uGAGs in milligrams per 24 h and milligrams per milligram creatinine.
Qualitative analysis of GAGs
Agarose gel electrophoresis
Total rGAGs (∼5 µg) were applied to a 0.5% agarose gel in 0.05 M 1,3-diaminopropane/acetate buffer, pH 9.0. After electrophoresis (120 V/h), the GAGs on the gel were fixed with 0.1% N-cetyl-N,N,N-trimethylammonium bromide in water, stained with 0.1% toluidine blue in acetic acid:ethanol:water (0.1:5:5, v/v/v), and washed for about 30 min in acetic acid:ethanol:water (0.1:5:5, v/v/v). GAGs on the agarose gel were quantified by the Molecular Analyst¯ Software 1.4 (Bio-Rad Laboratories, USA). One densitometry profile was obtained for each sample. Quantification of material represented by the peaks was made by estimating peak areas of these typical profiles. The gel was then stained with Stains-All (Sigma-Aldrich, USA), which intensifies the metachromasia of sulfated GAGs and reveals the presence of nonsulfated GAGs (17). Because of the low concentration of GAGs in the urinary excretion, total uGAGs were pooled (∼2 μg/sample) for electrophoretic analysis.
Polyacrylamide gel electrophoresis
The molecular weights of the GAG chains were estimated by polyacrylamide gel electrophoresis as previously described (18). Briefly, GAG samples (∼5 µg) were applied to a 6% 1-mm thick polyacrylamide gel slab in Tris-HCl buffer, pH 8.6. After electrophoresis (100 V/30 min), the gel was stained with 0.1% toluidine blue in 1% acetic acid and washed for about 4 h in 1% acetic acid. The gel was then stained with Stains-All and bleached in distilled water. Chondroitin 6-sulfate (60 kDa) from shark cartilage, chondroitin 4-sulfate (40 kDa) from whale cartilage (2 µg/mL), and dextran sulfate (8 kDa, 3 µg/mL) were used as molecular weight markers.
Statistical analysis
Data are reported as means±SE. For comparison of means, one-way analysis of variance (ANOVA) was used, with significant differences indicated by the Duncan test. Significance was set at P<0.05 (SPSS version 13.0 for windows, IBM, USA).
Results
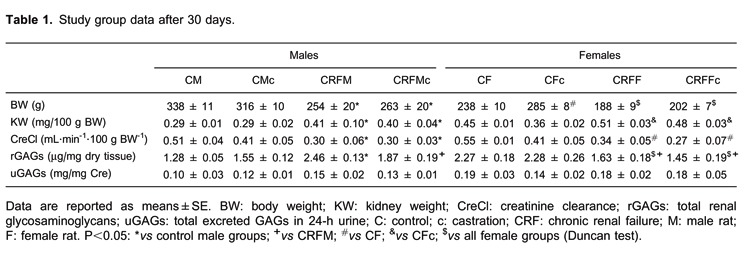
Characteristics of the groups 30 days after 5/6 nephrectomy are presented in Table 1. The death rate during surgical procedures was 5-8%, and survival after recovery from anesthesia was 100%. Data from the sham surgery group were similar to intact control groups (data not shown). CRF animals showed reduced CreCl. uGAGs were not different among the groups. However, rGAGs were almost two times higher in control females than control males (CF=2.27±0.18 vs CM=1.28±0.05; P<0.05). CRFM showed increased total rGAGs compared to all male groups and to the CRF females (P<0.05). After castration, rGAGs decreased in CRF males and remained unchanged in females.
CRF animals had increased PTN compared to their controls, and CRFM had higher PTN than CRFF in the same period of time (Figure 2). Castration resulted in different effects on PTN between genders, regardless of CRF. In control groups, castrated males showed decreased PTN compared to noncastrated males (CM=33.3±2.6 vs CMc=6.8±0.7 mg/24 h; P<0.0.5), but in the castrated animals there was no significant decrease in PTN in female rats (CF=7.1±1.8 vs CFc=4.8±0.6 mg/24 h). In castrated CRF males, PTN was decreased (CRFM=105.8±17.3 vs CRFMc=68.3±14.6 mg/24 h; P<0.05) but did not change in female castrated rats (CRFF=53.0±17.5 vs CRFFc=56.9±7.3 mg/24 h).
Figure 2. Proteinuria (mg/24 h) after 30 days. C: control; c: castration; CRF: chronic renal failure; F: female rat; M: male rat. P<0.05: + vs all; *vs CM; † vs CMc; # vs CF; & vs CFc.

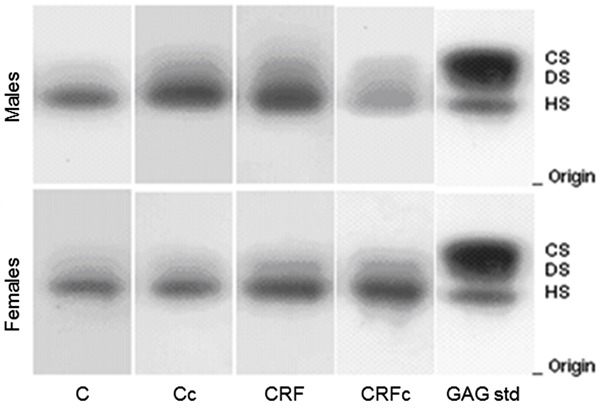
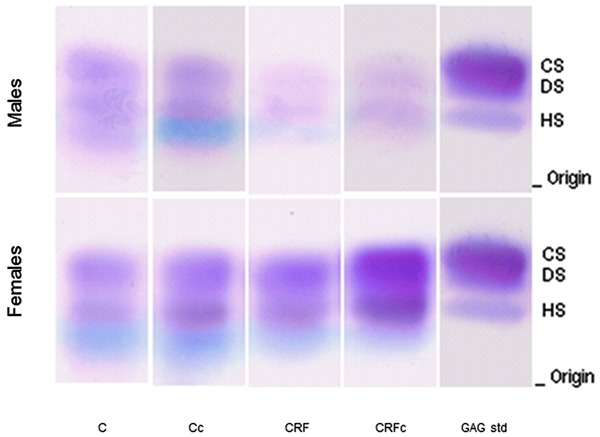
Agarose gel electrophoresis profiles of GAGs extracted from kidney after 30 days are shown in Figure 3. After staining the gels with Stains-All, there was no evidence of the presence of nonsulfated GAGs in the kidney. Densitometry following agarose gel electrophoresis showed one predominant band, corresponding to HS, in the kidneys of all study groups.
Figure 3. Agarose gel electrophoresis of total renal glycosaminoglycans (rGAGs) after toluidine blue staining. CS: chondroitin sulfate; DS: dermatan sulfate; HS: heparan sulfate; C: control; c: castration; CRF: chronic renal failure; GAG std: mixture of standard glycosaminoglycans.
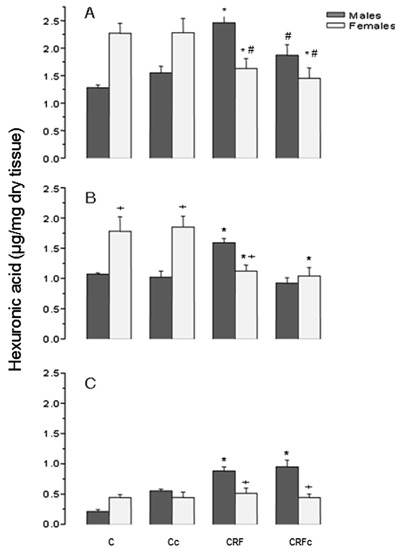
The quantification of rGAGs and hexuronic acid (µg/mg dry tissue) and the estimated content (µg/mg dry tissue) of HS and CS+DS from male and female rats are reported in Figure 4. Control females had higher concentrations of HS than control males. These data further emphasize the gender differences when comparing CRF groups: HS was reduced in females and increased in males. Castration reduced HS in males to levels similar to those found in females (Figure 4B). CS+DS (Figure 4C) were more homogeneous in females. CRFM presented significantly higher values of CS+DS compared to females and their respective controls (P<0.05).
Figure 4. Total glycosaminoglycans (A), heparan (B), and chondroitin+dermatan sulfate (C) in kidneys from male and female rats. C: control; c: castration; CRF: chronic renal failure. P<0.05: *vs respective control groups; + vs males; # vs male CRF.
Molecular weight analysis of total rGAGs showed no difference between male and female rats (∼40 kDa), while uGAGs did not show a defined pattern for molecular weight (Figure 5). Data for uGAGs showed a similar pattern among the groups, with predominance of CS+DS (Figure 6), except for CRF male groups that presented a decrease of CS+DS and HS. After staining the agarose gels by Stains-All, the presence of another clearer blue band was observed, suggesting the presence of hyaluronic acid or nonsulfated GAGs.
Figure 5. Polyacrylamide gel electrophoresis of total renal glycosaminoglycans (rGAGs) and total excreted GAGs in 24-h urine (uGAGs) from male and female rats. MW std: molecular weight standard; C: control; c: castration; CRF: chronic renal failure.
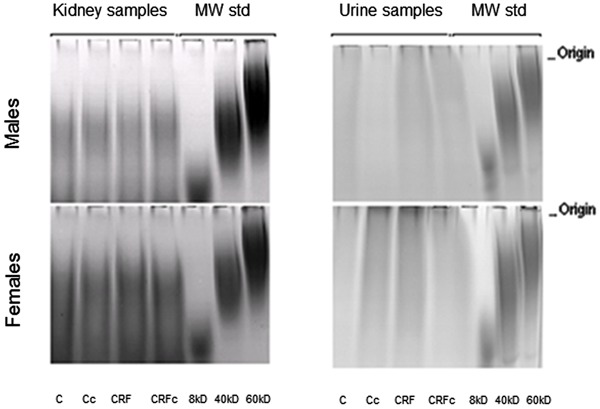
Figure 6. Agarose gel electrophoresis of pooled total excreted glycosaminoglycans (GAGs) in 24-h urine after toluidine blue and Stains-All stainings. CS: chondroitin sulfate; DS: dermatan sulfate; HS: heparan sulfate; C: control; c: castration; CRF: chronic renal failure; F: female rat; M: male rat; GAGs std: mixture of standard GAGs.
Discussion
In the present study, we analyzed the GAG profile in the kidneys of castrated and noncastrated male and female rats at an early stage of CRF. Reports relating sex hormones to GAGs and PTN in the CRF model are lacking. Animals in the study recovered well after surgery, and renal changes such as reduction of CreCl and renal hypertrophy were observed. Castration did not influence these parameters in either gender. Hormonal dosage was not performed after castration because hormonal reposition was not done. Animals with CRF developed PTN that was higher in males than in females and was reduced by castration. These data agree with our previous observations (19), suggesting that sex hormones may interfere with the appearance of PTN.
Evaluation of rGAGs showed higher values in CRF males, which were decreased after castration. Some authors have shown gender differences in GAG concentration, independent of castration, with adult females having higher total GAG content in the bladder compared to young females (4 weeks old) and male rats. This suggests that GAGs can vary with age and sex (20) and also with tissue type (21). However, studies evaluating rGAG profiles in female rats are lacking. The present data suggest that, in early stages of CRF (after 30 days), male sex hormones may interfere with the synthesis and/or degradation of rGAGs.
Regardless of gender, predominance of HS was found in the kidney. Furthermore, CRF males presented increased CS+DS. Females did not present differences in CS+DS among the study groups (Figure 4B and C). It is known that several growth factors and cytokines stored in the glomerular basement membrane interact with HS, and ultimately this interaction contributes to the tissue remodeling process (6). We observed an increase in kidney HS as well as PTN, and a decrease in HS in CRFM after castration, maintaining the same pattern observed with rGAGs.
Available data suggest that androgens may exacerbate, while estrogens may protect against, the development of mechanisms that ultimately lead to glomerulosclerosis. It has been suggested that the impact of gender on renal disease progression may reflect direct receptor-mediated effects of sex hormones (12, 13). Estradiol and testosterone receptors are expressed on the surface of mesangial cells (22). Endogenous estrogens have in general been considered to have anti-fibrotic and anti-apoptotic effects on the kidney (23). On the other hand, androgens present proapoptotic and profibrotic properties (24). Estrogen reduces proliferation of mesangial cells, collagen synthesis, renin-angiotensin system activity, and superoxide production, whereas testosterone potentiates these effects (10, 12, 25). These data also suggest that sex hormones could directly interfere with GAG synthesis.
uGAGs are considered mainly to result from glomerular metabolism, with some contribution of tubular and urinary tract cells, accounting for the higher concentration in urine compared to blood (26). Pooled urine samples from study groups were used to analyze uGAGs on agarose gels. No gender-related differences in uGAGs were observed in accordance with data reported by Lee et al. (26) and Michelacci et al. (27). Molecular weights of uGAGs were similar for all groups. In females, however, regions of metachromasia were observed near the origin, which suggest the presence of higher molecular weight molecules (Figure 5), such as hyaluronic acid from the urogenital system (28).
Although uGAGs were similar in all groups, CRF males showed a reduction in sulfated GAGs and the presence of nonsulfated GAGs, reflecting a qualitative change of GAGs in this group (Figure 6). Conversely, the uGAG profile did not change in the female groups. A reduction in uGAGs has been found in diabetic rats and in experimental glomerulonephritis, suggesting that uGAG excretion can vary with renal baseline pathology (1, 29, 30).
There is evidence that alterations in the glomerular basal membrane are the main cause of PTN (2, 3). Charge selectivity of the glomerular basal membrane has been widely associated with the electrostatic property of HS, and an inverse correlation between HS and PTN has been suggested (2, 31, 32). The mechanisms that link changes in GAGs and especially HS, compromising glomerular permeability and leading to PTN, are still unclear (1, 2, 33, 34). Regardless of its functional involvement with the charge of the glomerular barrier, HS is also involved in other biological functions. Conformational changes of the HS molecule are able to maintain constant activation of resident (epithelial and mesangial) and recruited cells, and these mechanisms are also linked to sex hormones (22). Changes caused by endosulfatases and/or heparanases (35, 36) and reactive oxygen species may remove a few epitopes essential to the biological functions of HS (10), contributing to the dysfunction of the glomerular filtration barrier, and causing PTN.
Early structural changes in the acetylation and sulfation of HS disaccharides, such as those occurring at glomerular maturation (37), give rise to an enormous structural diversity, which corresponds to the variety of biological functions mediated by HS including its role in inflammation (38). These modifications may have occurred in CRF males, altering the functionality of the de novo synthesized HS molecule in the glomerular filtration barrier. However, the present study did not analyze rGAG structure.
Apparent paradoxical data from CRF males (i.e., simultaneous increase in HS and PTN) were observed, but the reversion of this pattern in castrated animals may favor the hypothesis of a functionally compromised HS molecule. The behavior of CRF females contributes to this conclusion, in which a reduction in HS and an expected increase in PTN were observed and, in addition, these data remained unchanged after castration.
In conclusion, it is feasible to hypothesize that male sex hormones may play a role in the synthesis of rGAGs at early stages of CRF, interfering in renal remodeling of the ECM. Thus, gender-dependent quantitative differences were observed in the CRF model when measuring HS and PTN. Further studies analyzing rGAG structure in both sexes are needed to clarify how sex hormones and GAGs contribute to gender-dependent differences in CRF.
Footnotes
First published online July 30, 2013.
References
- 1.Cadaval RA, Kohlman O, Michelacci YM. Urinary excretion of glycosaminoglycans and albumin in experimental diabetes mellitus. Glycobiology. 2000;10:185–192. doi: 10.1093/glycob/10.2.185. [DOI] [PubMed] [Google Scholar]
- 2.Raats CJ, van den Born J, Berden JH. Glomerular heparan sulfate alterations: mechanisms and relevance for proteinuria. Kidney Int. 2000;57:385–400. doi: 10.1046/j.1523-1755.2000.00858.x. [DOI] [PubMed] [Google Scholar]
- 3.Remuzzi G, Bertani T. Is glomerulosclerosis a consequence of altered glomerular permeability to macromolecules? Kidney Int. 1990;38:384–394. doi: 10.1038/ki.1990.217. [DOI] [PubMed] [Google Scholar]
- 4.Burton C, Harris KP. The role of proteinuria in the progression of chronic renal failure. Am J Kidney Dis. 1996;27:765–775. doi: 10.1016/S0272-6386(96)90512-0. [DOI] [PubMed] [Google Scholar]
- 5.Remuzzi G, Benigni A, Remuzzi A. Mechanisms of progression and regression of renal lesions of chronic nephropathies and diabetes. J Clin Invest. 2006;116:288–296. doi: 10.1172/JCI27699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Parish CR. The role of heparan sulphate in inflammation. Nat Rev Immunol. 2006;6:633–643. doi: 10.1038/nri1918. [DOI] [PubMed] [Google Scholar]
- 7.Mitsuhashi H, Tsukada Y, Ono K, Yano S, Naruse T. Urine glycosaminoglycans and heparan sulfate excretions in adult patients with glomerular diseases. Clin Nephrol. 1993;39:231–238. [PubMed] [Google Scholar]
- 8.De Muro P, Faedda R, Satta A, Finetti D, Masala A, Cigni A, et al. Urinary glycosaminoglycan composition in chronic glomerulonephritis. J Nephrol. 2005;18:154–160. [PubMed] [Google Scholar]
- 9.Verhagen AM, Attia DM, Koomans HA, Joles JA. Male gender increases sensitivity to proteinuria induced by mild NOS inhibition in rats: role of sex hormones. Am J Physiol Renal Physiol. 2000;279:F664–F670. doi: 10.1152/ajprenal.2000.279.4.F664. [DOI] [PubMed] [Google Scholar]
- 10.Ji H, Zheng W, Menini S, Pesce C, Kim J, Wu X, et al. Female protection in progressive renal disease is associated with estradiol attenuation of superoxide production. Gend Med. 2007;4:56–71. doi: 10.1016/S1550-8579(07)80009-X. [DOI] [PubMed] [Google Scholar]
- 11.Sullivan JC, Semprun-Prieto L, Boesen EI, Pollock DM, Pollock JS. Sex and sex hormones influence the development of albuminuria and renal macrophage infiltration in spontaneously hypertensive rats. Am J Physiol Regul Integr Comp Physiol. 2007;293:R1573–R1579. doi: 10.1152/ajpregu.00429.2007. [DOI] [PubMed] [Google Scholar]
- 12.Sandberg K. Mechanisms underlying sex differences in progressive renal disease. Gend Med. 2008;5:10–23. doi: 10.1016/S1550-8579(08)80004-6. [DOI] [PubMed] [Google Scholar]
- 13.Silbiger S, Neugarten J. Gender and human chronic renal disease. Gend Med. 2008;5(Suppl A):S3–S10. doi: 10.1016/j.genm.2008.03.002. [DOI] [PubMed] [Google Scholar]
- 14.Schwartz MM, Bidani AK. Mesangial structure and function in the remnant kidney. Kidney Int. 1991;40:226–237. doi: 10.1038/ki.1991.204. [DOI] [PubMed] [Google Scholar]
- 15.Guimarães MA, Mourão PA. Urinary excretion of sulfated polysaccharides administered to Wistar rats suggests a renal permselectivity to these polymers based on molecular size. Biochim Biophys Acta. 1997;1335:161–172. doi: 10.1016/S0304-4165(96)00133-X. [DOI] [PubMed] [Google Scholar]
- 16.Taylor KA, Buchanan-Smith JG. A colorimetric method for the quantitation of uronic acids and a specific assay for galacturonic acid. Anal Biochem. 1992;201:190–196. doi: 10.1016/0003-2697(92)90194-C. [DOI] [PubMed] [Google Scholar]
- 17.Volpi N, Maccari F, Titze J. Simultaneous detection of submicrogram quantities of hyaluronic acid and dermatan sulfate on agarose-gel by sequential staining with toluidine blue and Stains-All. J Chromatogr B Analyt Technol Biomed Life Sci. 2005;820:131–135. doi: 10.1016/j.jchromb.2005.03.028. [DOI] [PubMed] [Google Scholar]
- 18.Tovar AM, Cesar DC, Leta GC, Mourao PA. Age-related changes in populations of aortic glycosaminoglycans: species with low affinity for plasma low-density lipoproteins, and not species with high affinity, are preferentially affected. Arterioscler Thromb Vasc Biol. 1998;18:604–614. doi: 10.1161/01.ATV.18.4.604. [DOI] [PubMed] [Google Scholar]
- 19.Lemos CC, Mandarim-de-Lacerda CA, Dorigo D, Coimbra TM, Bregman R. Chronic renal failure in male and female rats. J Nephrol. 2005;18:368–373. [PubMed] [Google Scholar]
- 20.Cabral CA, Sampaio FJ, Cardoso LE. Analysis of the modifications in the composition of bladder glycosaminoglycan and collagen as a consequence of changes in sex hormones associated with puberty or oophorectomy in female rats. J Urol. 2003;170:2512–2516. doi: 10.1097/01.ju.0000088776.17126.93. [DOI] [PubMed] [Google Scholar]
- 21.Fortepiani LA, Yanes L, Zhang H, Racusen LC, Reckelhoff JF. Role of androgens in mediating renal injury in aging SHR. Hypertension. 2003;42:952–955. doi: 10.1161/01.HYP.0000099241.53121.7F. [DOI] [PubMed] [Google Scholar]
- 22.Pawluczyk IZ, Tan EK, Harris KP. Rat mesangial cells exhibit sex-specific profibrotic and proinflammatory phenotypes. Nephrol Dial Transplant. 2009;24:1753–1758. doi: 10.1093/ndt/gfn714. [DOI] [PubMed] [Google Scholar]
- 23.Elliot SJ, Berho M, Korach K, Doublier S, Lupia E, Striker GE, et al. Gender-specific effects of endogenous testosterone: female alpha-estrogen receptor-deficient C57Bl/6J mice develop glomerulosclerosis. Kidney Int. 2007;72:464–472. doi: 10.1038/sj.ki.5002328. [DOI] [PubMed] [Google Scholar]
- 24.Metcalfe PD, Leslie JA, Campbell MT, Meldrum DR, Hile KL, Meldrum KK. Testosterone exacerbates obstructive renal injury by stimulating TNF-alpha production and increasing proapoptotic and profibrotic signaling. Am J Physiol Endocrinol Metab. 2008;294:E435–E443. doi: 10.1152/ajpendo.00704.2006. [DOI] [PubMed] [Google Scholar]
- 25.Park KM, Kim JI, Ahn Y, Bonventre AJ, Bonventre JV. Testosterone is responsible for enhanced susceptibility of males to ischemic renal injury. J Biol Chem. 2004;279:52282–52292. doi: 10.1074/jbc.M407629200. [DOI] [PubMed] [Google Scholar]
- 26.Lee EY, Kim SH, Whang SK, Hwang KY, Yang JO, Hong SY. Isolation, identification, and quantitation of urinary glycosaminoglycans. Am J Nephrol. 2003;23:152–157. doi: 10.1159/000070156. [DOI] [PubMed] [Google Scholar]
- 27.Michelacci YM, Cadaval RA, Rovigatti RM, Kohlman O. Renal and urinary glycosaminoglycans in an experimental model of chronic renal failure in rats. Exp Nephrol. 2001;9:40–48. doi: 10.1159/000020706. [DOI] [PubMed] [Google Scholar]
- 28.de Deus JM, Girao MJ, Sartori MG, Baracat EC, Rodrigues de Lima G, Nader HB, et al. Glycosaminoglycan profile in bladder and urethra of castrated rats treated with estrogen, progestogen, and raloxifene. Am J Obstet Gynecol. 2003;189:1654–1659. doi: 10.1016/S0002-9378(03)00867-6. [DOI] [PubMed] [Google Scholar]
- 29.de Lima CR, Aguiar JA, Michelacci YM. Reduced urinary excretion of sulfated polysaccharides in diabetic rats. Biochim Biophys Acta. 2005;1741:30–41. doi: 10.1016/j.bbadis.2004.10.001. [DOI] [PubMed] [Google Scholar]
- 30.Kaznowska-Bystryk I. Changes in the excretion of glycosaminoglycans with urine in patients with kidney stones. Ann Univ Mariae Curie Sklodowska Med. 1999;54:395–400. [PubMed] [Google Scholar]
- 31.van den Born J, van den Heuvel LP, Bakker MA, Veerkamp JH, Assmann KJ, Weening JJ, et al. Distribution of GBM heparan sulfate proteoglycan core protein and side chains in human glomerular diseases. Kidney Int. 1993;43:454–463. doi: 10.1038/ki.1993.67. [DOI] [PubMed] [Google Scholar]
- 32.Celie JW, Reijmers RM, Slot EM, Beelen RH, Spaargaren M, Ter Wee PM, et al. Tubulointerstitial heparan sulfate proteoglycan changes in human renal diseases correlate with leukocyte influx and proteinuria. Am J Physiol Renal Physiol. 2008;294:F253–F263. doi: 10.1152/ajprenal.00429.2007. [DOI] [PubMed] [Google Scholar]
- 33.Harvey SJ, Miner JH. Revisiting the glomerular charge barrier in the molecular era. Curr Opin Nephrol Hypertens. 2008;17:393–398. doi: 10.1097/MNH.0b013e32830464de. [DOI] [PubMed] [Google Scholar]
- 34.Morita H, Yoshimura A, Kimata K. The role of heparan sulfate in the glomerular basement membrane. Kidney Int. 2008;73:247–248. doi: 10.1038/sj.ki.5002659. [DOI] [PubMed] [Google Scholar]
- 35.Guimarães MA, Nikolovski J, Pratt LM, Greive K, Comper WD. Anomalous fractional clearance of negatively charged Ficoll relative to uncharged Ficoll. Am J Physiol Renal Physiol. 2003;285:F1118–F1124. doi: 10.1152/ajprenal.00370.2002. [DOI] [PubMed] [Google Scholar]
- 36.van den Hoven MJ, Wijnhoven TJ, Li JP, Zcharia E, Dijkman HB, Wismans RG, et al. Reduction of anionic sites in the glomerular basement membrane by heparanase does not lead to proteinuria. Kidney Int. 2008;73:278–287. doi: 10.1038/sj.ki.5002706. [DOI] [PubMed] [Google Scholar]
- 37.Lauer ME, Hascall VC, Wang A. Heparan sulfate analysis from diabetic rat glomeruli. J Biol Chem. 2007;282:843–852. doi: 10.1074/jbc.M608823200. [DOI] [PubMed] [Google Scholar]
- 38.Rops AL, van der Vlag J, Lensen JF, Wijnhoven TJ, van den Heuvel LP, van Kuppevelt TH, et al. Heparan sulfate proteoglycans in glomerular inflammation. Kidney Int. 2004;65:768–785. doi: 10.1111/j.1523-1755.2004.00451.x. [DOI] [PubMed] [Google Scholar]