

Abstract
Bacilli with fibrinolytic activities were isolated from traditionally-prepared Meju and some of these strains showed strong antifungal activities. One isolate, MJ1-4, showed the strongest antifungal activity. MJ1-4 and other isolates were identified as B. amyloliquefaciens strains by recA gene sequencing and RAPD-PCR results. B. amyloliqufaciens MJ1-4 efficiently inhibited an Aspergillus spp.-producing aflatoxin B1 (AFB1) and a Penicillium spp.-producing ochratoxin (OTA) in addition to other fungi. Antifungal activity of B. amyloliquefaciens MJ1-4 culture reached its maximum (40 AU/mg protein) in LB or TSB medium around 48 hr at 37°C. Antifungal activity of the concentrated culture supernatant was not decreased significantly by protease treatments, implying that the antifungal substance might not be a simple peptide or protein. Considering its antifungal and fibrinolytic activities together, B. amyloliquefaciens MJ1-4 can serve as a starter for fermented soyfoods such as Cheonggukjang and Doenjang.
Keywords: antifungal agents, Meju, Bacillus amyloliquefaciens, food preservatives
INTRODUCTION
Fungal infection of foods and agricultural products such as fruits and vegetables is a serious problem causing huge economical losses throughout the world (1). Fungal growth on foods causes undesirable changes in the appearance and flavor of foods, making them inappropriate for consumption. In some cases, toxigenic fungi produce mycotoxins in foods and consumption of contaminated foods might cause serious mycoses (2). Some well-known examples of aflatoxins are produced by certain Aspergillus species such as Aspergillus flavus and Aspergillus parasiticus; these aflatoxins are some of the most potent agents causing liver cancer (3). Ochratoxins are other well-known mycotoxins produced by some Aspegillus species and Penicillium species such as Aspergillus ochraceus and Penicillium viridicatum(4). Fresh vegetables and fruits are infected by a variety of plant pathogenic fungi, resulting in huge monetary losses worldwide (1); therefore, effective measures to prevent fungal growth on agricultural products and foods are urgently needed. Chemical preservatives have been used for the control of fungi but because of their often toxicities to men and farm animals, they must be used carefully. These days, consumers are more aware of the adverse effects of chemical preservatives and prefer foods containing natural and safe preservatives (5). It is desirable if generally recognized as safe (GRAS) organisms or metabolites from GRAS organisms that effectively inhibit fungi are used as preservatives against fungal infection. In this respect, bacilli and their metabolites have great potential. Bacilli, natural soil residents, have been known to produce more than two dozen different antimicrobial compounds and, thus, effectively compete with other microorganisms, such as fungi, in the same environment (6). Bacillus subtilis has been known to devote a significant portion of its genome (average 4∼5%) for genes involved in the production of antimicrobial compounds (6). Among the various antimicrobial compounds produced by B. subtilis, antibiotic peptides are the most important ones. They include ribosomally synthesized bacterocins and lantibiotics and nonribosomally synthesized peptide-containing compounds such as surfactins and itulins. Other classes of compounds include polyketides, amino sugars and phospholipids (6). B. amyloliquefaciens and B. licheniformis also produce antimicrobial substances but compared to B. subtilis, few studies have been reported (7–9). Bacilli, with antifungal activities and belonging to B. subtilis group, can be used as starters for fermented foods such as Cheonggukjang and Doenjang. Safety of fermented foods can be improved by using such bacilli as starters. Those bacilli also might be useful as biocontrol agents against fungi infecting fruits and vegetables. In this report, we isolated Bacillus amyloliquefaciens strains with antifungal activities from Meju and one of them, MJ1-4, was further studied. We report some properties of MJ1-4 and its antifungal substances.
MATERIALS AND METHODS
Bacterial strains and cultivation conditions
Bacterial strains used in this study are listed in Table 1. Luria-Bertani medium (LB, 1% tryptone, 0.5% yeast extract, 1% NaCl) was used for the propagation of bacilli and bacilli were cultivated at 37°C with aeration. Fungi were grown on PDA agar (Difco, Becton Dickinson and company, Franklin Lakes, NJ, USA) and fungal spores were recovered from mycelia after 5∼7 days incubation at 25°C. Mycelia were collected by using sterile wood-sticks and resuspended in sterile deionized water. Resuspended spores were filtered using cheese cloth and the number of spores was counted by using a haemocytometer (Marienfeld lab glassware, Lauda-Königshofen, Germany).
Table 1.
Bacterial strains used in this study
| Species | Strain | Source and relevant information |
|---|---|---|
| Bacillus amyloliqueficiens | MJ1-4 | This study |
| Bacillus amyloliqueficiens | MJ3-6 | This study |
| Bacillus amyloliqueficiens | MJ5-48 | This study |
| Bacillus amyloliqueficiens | MJ7-66 | This study |
| Bacillus amyloliqueficiens | CH51 | 10 |
| Bacillus amyloliqueficiens | CH86-1 | 10 |
| Bacillus subtilis | CH3-5 | 10 |
| Bacillus subtilis | CH3-25 | 10 |
| Bacillus subtilis | CH97 | 10 |
| Bacillus licheniformis | CH3-17 | 10 |
| Rhizopus microsporus var. oligosporus | ATCC 22959 (KACC 41363) | |
| Aspergillus awamori | ATCC 14331 (KACC 41844) | |
| Aspergillus niger | ATCC 4695 (KACC 40280) | |
| Aspergillus oryzae | ATCC 11493 (KCCM 60166) | |
| Aspergillus sojae | ATCC 9362 (KCCM 60354) | |
| Aspergillus spp. | Producing AFB1 | |
| Penicillium spp. | Producing OTA |
Isolation of Bacillus species with antifungal activities from Meju
Meju, prepared by traditional methods in Sunchang county, Jeonbuk, was homogenized using a stomacher (Seward Ltd., Worthing, West Sussex, UK). Twenty-five g of Meju was mixed with 225 mL of 0.1% peptone water for homogenization. Homogenate was serially diluted with sterile water and each dilution was spread on LB agar plates containing 0.7% skim milk. Plates were incubated for 2 days at 37°C and colonies showing protease activities were selected. They were inoculated into fresh LB media and stored frozen after overnight growth. These bacilli were screened for the antifungal activities as follows: each isolate was cultivated in LB broth for 24 hr and then 2 μL of culture was spotted on a PDA agar plate near the rim, while the center of the plate was inoculated with fungal spores. Growth inhibition of mycelia near the spotted areas was examined after 5 days incubation at 25°C. Agar well diffusion method was used for the antifungal activity assays. PDA top agar (0.7%, w/v) containing 107 fungal spores was overlayed on an LB plate. Holes were made using a capillary glass tube and culture supernatant or concentrated culture supernatant (ammonium sulfate precipitated) of each Bacillus strain was added into each hole. Then the plate was incubated for 5 days at 25°C.
Identification of Bacillus species with antifungal activities
Identification of isolates was done by recA gene sequencing and a RAPD-PCR method developed at our lab (10). For recA gene amplification, a primer pair was designed based on the conserved regions in recA sequences from Bacillus sp.: recA-F (5′-TGAGTGATCGTCAGGCAGCCTTTAG-3′) and recA-R (5′-CYTBRGATAAGARTACCAWGMACCGC-3′). RAPD-PCR was done using S-30 (5′-GTGATCGCAG-3′) primer and GoTaq green master mix (Promega, Madison, WI, USA), as described previously (10). Fibrinolytic activity of the isolates was measured by fibrin plate method (11). Plasmid DNA was prepared by a method described by Voskuil and Chambliss (12). The morphology of MJ1-4 was examined by SEM (scanning electron microscopy) as follows: cells were grown in LB broth for 24 hr and fixed in paraformaldehyde-glutaraldehyde (5%) at 4°C. Fixing buffer was removed and washed twice with 0.1 M phosphate buffer (pH 7.0). Then cells were fixed in 2% osmium tetroxide for 2 hr and washed twice with 0.1 M phosphate buffer (pH 7.0). Then cells were dehydrated using 95% ethanol and dried in hexamethyldisilazane for l hr. Prepared sample was observed using a SEM (XL30 S FEG, Philips, Eindhoven, Netherlands) at the magnification of 50,000×.
Antifungal activity changes during growth of MJ1-4 in different media
The effect of growth medium on the antifungal activity was tested using four different media: LB, nutrient broth (NB, Difco), tryptic soy broth (TSB, Difco), and brain heart infusion (BHI, Difco). Each medium, 1% (v/ v) inoculated with B. amyloliquefaciens MJ1-4, was incubated at 37°C up to 72 hr with aeration. Thirty mL of culture was taken at intervals and absorbance at 600 nm, viable cell counts and antifungal activities were measured. Culture was centrifuged at 5,000×g for 10 min at 4°C and supernatant was filtered using a 0.45 μm syringe filter (Minisart CE, Satorius Stedim, Aubagne, France). Ammonium sulfate (Amresco, Solon, OH, USA) was added to the culture supernatant to the final concentration of 80% saturation (w/v) and stored at 4°C overnight. An ammonium sulfate pellet was obtained by centrifugation (12,000×g, 30 min at 4°C) and resuspended in small volume of 100 mM Tris-HCI (pH 8.0). Dialysis was done against the same buffer for 24 hr at 4°C using regenerated cellulose membrane (molecular weight cutoff, 12,000∼14,000 Da, Membrane filtration products, Seguin, Guadalupe, TX, USA). Dialyzate was freeze-dried and resuspended in a small volume of sterile water. This 80% ammonium sulfate sample (called the AS sample) was used for the activity measurements, stability tests and inhibition spectrum studies. AS samples were serially diluted two-fold with sterile water and then anti-fungal activity was examined. Antifungal activity was calculated as the reciprocal of the maximum dilution still showing inhibition and defined as an activity unit (AU). The antifungal activity was expressed as AU/mg protein. A Penicillium species producing ochratoxin was used as an indicator. Protein concentration was measured by the Bradford method using bovine serum albumin (BSA) as a standard (13).
Inhibition spectrum
The inhibition spectrum of the AS sample from B. amyloliquefaciens MJ1-4 was examined using various fungal species. PDA soft agar (0.7%, w/v) containing 105 fungal spores (Table 1) was overlayed on LB plates. A paper disk was placed on the overlay and AS sample (102 μg) was applied onto each disk. Then plates were incubated at 25°C. Amphotericin B (Sigma, St. Louis, MO, USA) was applied at the same time as a positive control.
Stability of antifungal substance against proteolytic enzymes, pH and heat
B. amyloliquefaciens MJ1-4 was cultivated for 48 hr in LB at 37°C and AS sample was prepared as described above. 58.4 μg of AS sample was treated with proteolytic enzymes for 30 min at 37°C: 354 units of proteinase K (Sigma P2308, from Tritirachium album), 42 units of protease (Sigma P5147, from Streptomyces griseus), 4,690 units of pepsin (Sigma P-6887, from porcine stomach mucosa), and 14,730 units of trypsin (Sigma T-8918, from bovine pancreas). Proteolytic enzymes except pepsin were resuspended in 5 mM sodium phosphate buffer (pH 7.0). Pepsin was resuspended in 0.02 N HCl. Treated samples were applied onto paper disks on PDA top agar previously overlayed on an LB plate. Twelve mL of PDA top agar was mixed with 1 mL of spore suspension (105) of a Penicillium species producing OTA. Plates were incubated for 5 days at 26°C. For measuring the heat stability, 50 μg of AS sample was suspended in sodium phosphate buffer (pH 7.0) and exposed to 50°C, 70°C, and 100°C for 1 hr and 121°C for 15 min. The remaining activities were measured using a Penicillium species as an indicator as described above. pH stability of AS sample (50 μg) was examined by measuring the remaining activity after 2 hr incubation at pH 3∼10. Citrate-NaOH buffer (pH 3∼5), sodium phosphate buffer (pH 5∼8), and Tris-HCl buffer (pH 8∼10) were used and a Penicillum species producing OTH was used as an indicator.
Purification of antifungal substances
AS sample was applied onto a Sephadex G-75 column (1.5×26.5 cm, GE Healthcare, Uppsala, Sweden) and eluted with 100 mM Tris-HCl (pH 8.0). Fractions were collected and the absorbance of each fraction (1 mL) was monitored at 280 nm using a UV-visible spectrophotometer. Ten μL of each fraction was taken out and examined for antifungal activity.
RESULTS AND DISCUSSION
Identification of Bacillus species with antifungal activity from Meju
A total of 448 colonies showing protease activities were isolated from Meju, Korean traditional fermented soyfood using LB plates with skim milk. These isolates were screened for antifungal activities and several isolates were obtained. Four isolates (MJ 1–4, 3–6, 5–48, and 7–66) showing higher activities were selected for further studies. They were identified by recA gene sequence analyses to be Bacillus amyloliquefaciens strains. A recA gene fragment was amplified from each isolate and the sequence was determined. The size of the amplified fragment was in the range of 666 and 685 base pairs (bp). A BLAST search showed that each sequence was 99% homologous to recA from Bacillus amyloliquefaciens FZB42, a reference strain for which genome sequencing was completed (results not shown). Genbank accession numbers are JN813530, JN813531, JN813532, and JN813533 for the partial recA gene from MJ1-4, MJ3-6, MJ5-48, and MJ7-66, respectively. RAPD-PCR profiles of the isolates were also obtained using S30 (5′-GTGATCGCAG-3′) as a single primer (10). MJ1-4 generated 1.1 and 1.5 kb bands which are common bands among B. amyloliquefaciens strains including CH51 and CH86-1, two strains previously isolated from Cheonggukjang (Fig. 1, lane 8, 9) (10). Three other isolates generated only 1.1 kb band and failed to generate a 1.5 kb band but their RAPD-PCR profiles were similar to those of B. amyloliqufaciens strains. From the recA sequence and RAPD-PCR profiles, four strains were identified to be B. amyloliquefaciens and named B. amyloliquefaciens MJ1-4, MJ3-6, MJ5-48, and MJ7-66, respectively. Except for MJ3-6, three strains had fibrinolytic activities when the culture supernatant (10 μL) of each strain was examined by fibrin plate method. B. amyloliquefaciens MJ7-66 showed the strongest fibrinolytic activity followed by MJ1-4 and then MJ5-48 (data not shown). Other isolates with antifungal activities did not possess fibrinolytic activities (data not shown). Considering the antifungal and fibrinolytic activities, three B. amyloliquefaciens strains seem qualified as starters for fermented soyfoods. When plasmid profiles of these strains were examined by agarose gel electrophoresis, no plasmid was detected, indicating that the gene(s) responsible for antifungal activity was chromosomally located (data not shown). All four strains produced sticky material when grown on LB agar plate. MJ3-6 was the strain producing sticky materials most profusely, while MJ1-4 produced little sticky material (data not shown). It was difficult to isolate single colonies of MJ3-6 on LB agar plates due to the sticky material. Considering these observations together, four strains differ from each other in certain specific properties. Thus, each strain needs to be studied individually if strains are selected for any specific applications such as fibrinolytic activity or exopolysaccharide production. B. amyloliquefaciens MJ1-4 is rod-shaped organism with rough outer surfaces as shown by scanning electron microscopy (Fig. 2).
Fig. 1.
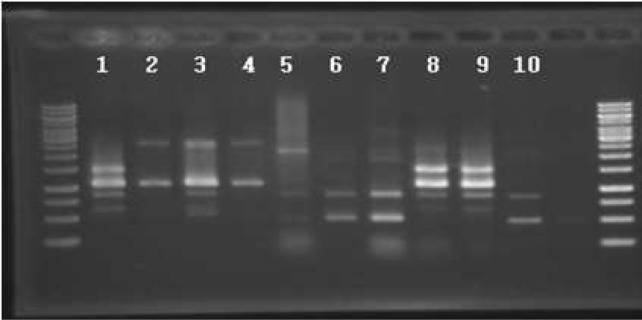
RAPD-PCR profiles of selected Bacillus isolates from Meju. Lane 1, MJ1-4; 2, MJ3-6; 3, MJ5-48; 4, MJ7-66; 5, B. subtilis CH3-5; 6, B. licheniformis CH3-17; 7, B. subtilis CH3-25; 8, B. amyloliquefaciens CH51; 9, B. amyloliquefaciens CH86-1; 10, B. subtilis CH97. Bacillus CH strains were previously isolated from Cheonggukjang(10).
Fig. 2.

Electron microgram of B. amyloliquefaciens MJ1-4 observed by scanning electron microscopy (XL30 S FEG, Philips, Netherland). The magnification was ×50,000 and a scale corresponds to 500 nm.
Inhibition spectrum of AS sample from B. amyloliquefaciens MJ1-4
Inhibition of fungal species by AS sample from B. amyloliquefaciens MJ1-4 was examined. AS sample strongly inhibited a Penicillium spp. producing OTA, A. niger ATCC 4695, and A. awamori ATCC 14331 (Table 2). An Aspergillus spp. producing AFB1 was also inhibited significantly. Penicillum and related fungi cause spoilage of stored cereals, bread and flour, peanuts and nuts such as walnuts, resulting in significant economical losses. Thus B. amyloliqufaciens MJ1-4 and/or its anti-fungal agent(s) might be useful for the control of spoilage and toxigenic fungi.
Table 2.
Antifungal activities of Bacillus sp.
| Test strain | MJ1-4 | MJ3-6 | MJ5-48 | MJ7-66 | CH51 | CH86-1 | B. subtilis 168 |
|---|---|---|---|---|---|---|---|
| Aspergillus oryzae ATCC 11493 | +1) | − | − | − | + | − | − |
| Aspergillus sojae ATCC 9362 | + | − | − | − | + | − | − |
| Aspergillus niger ATCC 4695 | +++ | − | +++ | − | +++ | +++ | − |
| Aspergillus awamori ATCC 14331 | +++ | − | +++ | − | ++ | ++ | − |
| Rhizopus microsporus var. oligosporus ATCC 22959 | + | − | − | − | − | − | − |
| Aspergillus spp., AFB1 producer | ++ | − | ++ | − | ++ | ++ | − |
| Penicillium spp., OTA producer | +++ | − | ++ | − | ++ | ++ | − |
Inhibition zone size: +, 0.5 to 2 mm2); ++, 2 to 4 mm; +++, more than 4 mm.
The value equals to (diameter of an inhibition zone–diameter of a colony)/2.
Stability of antifungal substances against proteolytic enzymes, pH variation and heat
The antifungal activity of the AS sample from B. amyloliquefaciens MJ1-4 was not affected by protease treatments (Fig. 3). Proteinase K treatment with the AS sample reduced the size of the inhibition zone a little whereas other protease treatments did not affect the size of the inhibition zone. These results indicated that the major antifungal compound was not a simple protein or peptide. Rather, small compounds such as iturin or surfactin might be responsible for the antifungal activity. When PCR was done using a primer pair targeting for an ituB gene involved in the iturin synthesis, an amplicon with expected size was observed (results not shown) for B. amyloliquefaciens MJ1-4 and MJ3-6, indicating the likely production of iturin in these strains. Another possibility is that the substance might be a lantibiotic with heavily modified amino acids, which is difficult for proteases to digest. It is also possible that more than two different compounds are present in the AS sample, including one bacteriocin-like substance and another non-proteinaceous compound. During the preparation of AS sample, dialysis was done using dialysis membrane with molecular weight cut-off 12,000∼14,000 Da. Thus, it can be expected that most small-sized antifungal substance(s) might be lost during dialysis unless they were aggregated. Since the AS sample showed antifungal activity after dialysis and the activity was insensitive to proteases, the compound(s) somehow remained during dialysis. Detailed characterization of the antifungal substances in the AS sample might answer all of these questions, including the nature of the major antifungal compounds produced by B. amyloliquefaciens MJ1-4. Bacillus species are well-known to produce a vast array of different antimicrobial substances, including bacteriocins, lipopeptides, and other small-sized molecules (6,14,15). Bacilli with antifungal activities could be isolated from natural environments, including fermented foods, plants, and soil. Those bacilli and their antifungal substances are the subjects of recent studies because of their potential as biopreservatives for foods and feed against fungal infection (16,17).
Fig. 3.
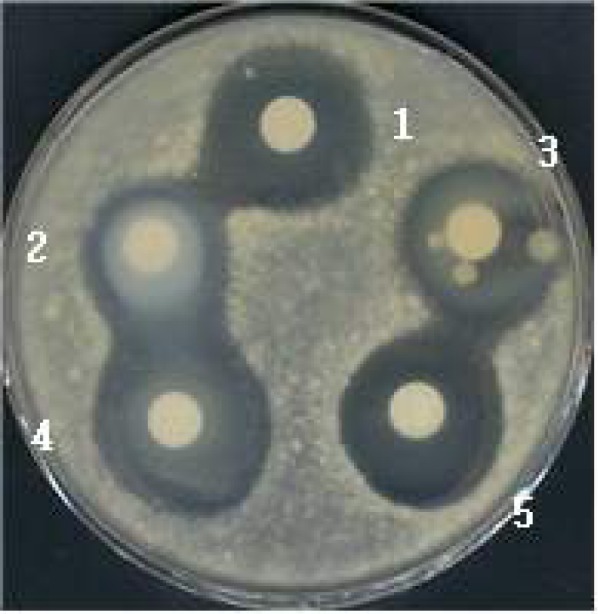
Stability of the antifungal activity of AS sample against proteolytic enzyme treatments. AS sample (58.4 μg) was treated with different proteases for 30 min at 37°C. A Penicillium spp. producing OTA was used as an indicator. 1, control (no enzyme treatment); 2, proteinase K (354 units) treated; 3, pro-tease (42 units) treated; 4, trypsin (14,730 units) treated; 5, pepsin (4,690 units) treated.
The antifungal activity of AS sample was most active at pH 6 and 7 and relatively stable at pH 8–10 but unstable at acidic pHs (pH 3∼5). The antifungal activity of the AS sample was stable after 1 hr exposure at 70°C and about half of the activity (48%) still remained at 100°C for 1 hr (Table 3), but the activity was destroyed by autoclaving (121°C, 15 min). The results showed that AS sample had significant heat resistance, supporting the idea that the antimicrobial substance(s) might be a nonproteinaceous, small-sized compound or a highly modified peptide.
Table 3.
Stability of antifungal activity against heat and pH
| Treatment | Remaining activity (%) |
|---|---|
| Control | 100 |
| Heat treatment (15 min exposure) | |
| 50°C, 1 hr | 100 |
| 70°C, 1 hr | 100 |
| 100°C, 1 hr | 48 |
| 121°C, 15 min | 0 |
| pH variation | |
| 3 | 37 |
| 4 | 34 |
| 5 | 54 |
| 6 | 100 |
| 7 | 100 |
| 8 | 69 |
| 9 | 66 |
| 10 | 67 |
Antifungal activity changes of B. amyloliquefaciens MJ1-4 during growth in different media
Among the four media tested, LB and TSB were better than NB and BHI in antifungal activities. Viable cell numbers were basically the same in all four media, reaching up to 109 cells/mL at 12∼18 hr except NB (36 hr). In LB and TSB, antifungal activity of the culture supernatant of MJ1-4 increased by stepwise fashion and reached the highest point (40 AU/mg protein) around 48 hr and then remained constant. In NB, antifungal activity increased sharply from basal level after 36 hr, reaching the maximum value (40 AU/mg protein) at 48 hr, and then decreased rapidly. In BHI, culture showed only half (20 AU/mg protein) of the highest activity of those in other media at 36 hr and then remained constant (Fig. 4).
Fig. 4.

Changes in absorbance, viable cell number and antifungal activity of B. amyloliquefaciens MJ1-4 culture during 72 hr cultivation. A, NB; B, TSB; C, BHI; D, LB.
 , absorbance (OD600);
, absorbance (OD600);
 , viable cell numbers (CFU/mL);
, viable cell numbers (CFU/mL);
 , antifungal activity (AU/mg protein).
, antifungal activity (AU/mg protein).
Antifungal substance(s) production and/or stability might be affected significantly by growth medium (18, 19), thus, growth medium should be selected carefully if maximum production of antifungal substance(s) is intended. From these results, LB or TSB is the medium of choice for the production of antifungal substance(s).
Partial purification of antifungal substance
Antifungal substance was purified from the AS sample using Sephadex G-75 column chromatography. Fig. 5 shows the elution profile and fractions 39∼50 contained the antifungal activity. Fractions 42∼43 and 47∼48 showed higher activities, indicating a possibility that two different antifungal compounds were present in the AS sample. Characterization of each compound is necessary to elucidate the nature of the antifungal activities of the AS sample. Studies of these two fractions are in progress now. Detection of antimicrobial substance was attempted by using SDS-PAGE and activity overlay, but no clear bands were observed (data not shown).
Fig. 5.

Purification of 80% ammonium sulfate precipitated substances (AS) by size exclusion chromatography using Sephadex G-75. Antifungal activity of AS sample was examined by using Penicillium spp. producing OTA as an indicator.
Further studies on the nature of these antifungal substances are necessary to develop efficient methods for using the strain and its antifungal agents. Genes responsible for the antifungal activity need to be cloned for the purpose of improving the stability or yield of the substance(s). Increase in the production yield also involves the optimization of cultivation methods, including medium optimization. In conclusion, the four isolates (MJ1-4, MJ3-6, MJ5-48, and MJ7-66) could be used as starters for the production of Cheonggukjang or Doenjang. Antifungal activities of bacilli, including the four strains in this study, should be studied for food safety purposes.
Acknowledgments
This research was supported by Youlchon Foundation (2009). It was also supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2010-0008048). H. A Lee was supported by a scholarship from the BK 21 program, the Ministry of Education, Science and Technology, Korea.
REFERENCES
- 1.Moss MO. Fungi, quality and safety issues in fresh fruits and vegetables. J Appl Microbiol. 2008;104:1239–1243. doi: 10.1111/j.1365-2672.2007.03705.x. [DOI] [PubMed] [Google Scholar]
- 2.Melin P, Sundh I, Håkansson S, Schnürer J. Biological preservation of plant derived animal feed with anti-fungal microorganisms: safety and formulation aspects. Biotechnol Lett. 2007;29:1147–1154. doi: 10.1007/s10529-007-9375-9. [DOI] [PubMed] [Google Scholar]
- 3.Bennett JW, Klich M. Mycotoxins. Clin Microbiol Rev. 2003;16:497–516. doi: 10.1128/CMR.16.3.497-516.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bayman P, Baker JL. Ochratoxins: A global perspective. Mycopathologia. 2006;162:215–223. doi: 10.1007/s11046-006-0055-4. [DOI] [PubMed] [Google Scholar]
- 5.Settanni L, Corsetti A. Application of bacteriocins in vegetable food biopreservation. Int J Food Microbiol. 2008;121:123–138. doi: 10.1016/j.ijfoodmicro.2007.09.001. [DOI] [PubMed] [Google Scholar]
- 6.Stein T. Bacillus subtilis antibioitic: structures, syntheses and specific functions. Mol Microbiol. 2005;56:845–857. doi: 10.1111/j.1365-2958.2005.04587.x. [DOI] [PubMed] [Google Scholar]
- 7.Lisboa MP, Bonatto D, Bizani D, Henriques JAP, Brandelli A. Characterization of a bacteriocin-like substance produced by Bacillus amyloliquefaciens isolated from the Brazilian Atlantic forest. Int Microbiol. 2006;9:111–118. [PubMed] [Google Scholar]
- 8.Benitez LB, Voltolini R, Lisboa MP, da Costa Medina LF, Brandelli A. Isolation and characterization of antifungal peptides produced by Bacillus amyloliquefaciens LBM 5006. J Microbiol. 2010;48:791–797. doi: 10.1007/s12275-010-0164-0. [DOI] [PubMed] [Google Scholar]
- 9.Wong JH, Hao J, Cao Z, Qiao M, Xu H, Bai Y, Ng TB. An antifungal protein from Bacillus amyloliquefaciens. J Appl Microbiol. 2008;105:1888–1898. doi: 10.1111/j.1365-2672.2008.03917.x. [DOI] [PubMed] [Google Scholar]
- 10.Kwon GH, Lee HA, Park JY, Kim JS, Lim J, Park CS, Kwon DY, Kim YS, Kim JH. Development of a RAPD-PCR method for identification of Bacillus species isolated from Cheonggukjang. Int J Food Microbiol. 2009;129:282–287. doi: 10.1016/j.ijfoodmicro.2008.12.013. [DOI] [PubMed] [Google Scholar]
- 11.Astrup T, Mullertz S. The fibrin method for estimating fibrinolytic activity. Arch Biochem Biophys. 1952;40:346–351. doi: 10.1016/0003-9861(52)90121-5. [DOI] [PubMed] [Google Scholar]
- 12.Voskuil MI, Chambliss GH. Rapid isolation and sequencing of purified plasmid DNA from Bacillus subtilis. Appl Environ Microbiol. 1993;59:1138–1142. doi: 10.1128/aem.59.4.1138-1142.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bradford MM. Rapid and sensitive methods for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 14.Schallmey M, Singh A, Ward OP. Developments in the use of Bacillus species for industrial production. Can J Microbiol. 2004;50:1–17. doi: 10.1139/w03-076. [DOI] [PubMed] [Google Scholar]
- 15.Riazi S, Wirawan RE, Badmaev V, Chikindas ML. Characterization of lactosporin, a novel antimicrobial protein produced by Bacillus coagulans ATCC 7050. J Appl Microbiol. 2009;106:1370–1377. doi: 10.1111/j.1365-2672.2008.04105.x. [DOI] [PubMed] [Google Scholar]
- 16.Gálvez A, López RL, Abriouel H, Valdivia E, Omar NB. Application of bacteriocins in the control of food-borne pathogenic and spoilage bacteria. Crit Rev Biotechnol. 2008;28:125–152. doi: 10.1080/07388550802107202. [DOI] [PubMed] [Google Scholar]
- 17.Hammami I, Rhouma A, Jaouadi B, Rebai A, Nesme X. Optimization and biochemical characterization of a bacteriocin from a newly isolated Bacillus subtilis strain 14B for biocontrol of Agrobacterium spp. strains. Lett Appl Microbiol. 2009;48:253–260. doi: 10.1111/j.1472-765X.2008.02524.x. [DOI] [PubMed] [Google Scholar]
- 18.Kumar A, Saini P, Shrivastava JN. Production of peptide antifungal antibiotic and biocontrol activity of Bacillus subtilis. Indian J Exp Biol. 2009;47:57–62. [PubMed] [Google Scholar]
- 19.Wang ZW, Liu XL. Medium optimization for anti-fungal active substances production from a newly isolated Paenibacillus sp. using response surface methodology. Bioresour Technol. 2008;99:8245–8251. doi: 10.1016/j.biortech.2008.03.039. [DOI] [PubMed] [Google Scholar]