

Abstract
In humans and other species, long-term hypoxia (LTH) during pregnancy can lead to intrauterine growth restriction with reduced body/brain weight, dysregulation of cerebral blood flow (CBF), and other problems. To identify the signal transduction pathways and critical molecules, which may be involved in acclimatization to high altitude LTH, we conducted microarray with advanced bioinformatic analysis on carotid arteries (CA) from the normoxic near-term ovine fetus at sea-level and those acclimatized to high altitude for 110+ days during gestation. In response to LTH acclimatization, in fetal CA we identified mRNA from 38 genes upregulated >2 fold (P<0.05) and 9 genes downregulated >2-fold (P<0.05). The major genes with upregulated mRNA were SLC1A3, Insulin-like growth factor (IGF) binding protein 3, IGF type 2 receptor, transforming growth factor (TGF) Beta-3, and genes involved in the AKT and BCL2 signal transduction networks. Most genes with upregulated mRNA have a common motif for Pbx/Knotted homeobox in the promoter region, and Sox family binding sites in the 3′ un translated region (UTR). Genes with downregulated mRNA included those involved in the P53 pathway and 5-lipoxygenase activating proteins. The promoter region of all genes with downregulated mRNA, had a common 49 bp region with a binding site for DOT6 and TOD6, components of the RPD3 histone deacetylase complex RPD3C(L). We also identified miRNA complementary to a number of the altered genes. Thus, the present study identified molecules in the ovine fetus, which may play a role in the acclimatization response to high-altitude associated LTH.
Introduction
The cellular and molecular mechanisms by which an organism acclimatizes to high altitude long-term hypoxia (LTH) are complex and not well understood. In human pregnancy, antenatal LTH, as a consequence of high altitude residence, maternal smoking, anemia, heart and/or lung diseases, placental pathologies (such as pre-eclampsia, placental insufficiency, abruptio placenta), or other factors may be associated with a generalized intrauterine growth restriction (IUGR) reduced organ weights (including brain and heart), and/or persistent pulmonary hypertension of the newborn [1]. In rodents, antenatal hypoxia also is known to cause reduced litter size and low birth weight pups [2].
In previous studies we have demonstrated that in response to prolonged maternal antenatal hypoxic exposure at high altitude (3,801 m). The sheep fetus undergoes successful acclimatization without the LTH-induced pathological conditions observed in many other species [3], [4]. This also has been reported in the sheep by others [5]. In contrast, high-altitude (2,740 to 3,100 m) exposed human infants have been reported to demonstrate intrauterine growth restriction, reduced weight of brain and other organs, increased incidence of pulmonary hypertension, as well as other pathological conditions [6], [7]. Of note, studies suggest 100 g of birth-weight reduction per 1000 m of elevation [8]. Despite the higher elevation in our study (3,801 m), the influence of hypoxic stress on the sheep fetus is attenuated, representing a unique adaptive characteristic.
Furthermore, the fetuses acclimatized to high altitude LTH had near-normal [9] cerebral blood flow despite a significant 27±4% decrease in cardiac output, and a 49±6% decrease in blood flow to the carcass and most other organs [3]. Additionally, we have observed that the acclimatized ovine fetus can maintain normal brain weight, cerebral oxygenation, cerebral O2 metabolic rate, sagittal sinus PO2, cortical tissue PO2, and electro-encephalographic activity at a level similar to the sea-level normoxic fetus [9], [10]. These findings raise questions regarding the mechanisms whereby, in contrast to the human, the ovine fetus successfully maintains normal CBF and brain weight during prolonged hypoxic exposure.
Carotid arteries (CA) have been shown to play a crucial role in the regulation and maintenance of CBF [11]. During increased flow demand, there is a significant pressure gradient from CA to cerebral arteries [12].. Importantly, other studies suggest that much of the change in systemic pressure results in dilation/contraction of the large arteries that supply the brain [13]. These studies underscore the importance of CA in the regulation of CBF, and suggest that failure of CA to effectively regulate the pressure of the blood reaching delicate cerebral arteries may lead to rupture with hemorrhage. Moreover, evidence suggests that the large cranial arteries of premature as well as intrauterine growth restricted infants may be unable to regulate effectively their CBF, as opposed to the near-term newborn [14], [15].
Thus, we tested the hypothesis that in fetal sheep, antenatal maternal high altitude long-term hypoxia is associated with changes in the gene expression in carotid arteries, which may be important in successful cerebrovascular acclimatization response and the maintenance of CBF, as well as normal for brain growth. By use of novel ovine oligonucleotide microarrays and signal pathway analysis, in near-term (140 days gestation) fetuses from both normal sea-level controls and those acclimatized to high altitude during antenatal development. We examined changes in carotid arteries gene expression pathways. We also conducted advanced bioinformatic analysis to identify cis-regulatory elements, trans-acting factors, and microRNA (miRNA), which can regulate gene expression in the CA.
Methods
Experimental animals and tissues
All experimental procedures were performed within the regulations of the Animal Welfare Act, the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and - the present study was approved by the Loma Linda University Animal Care and Use Committee.
Eight “Western breed” ewes were obtained from a single supplier (Nebeker Ranch, Lancaster, CA) and were allocated to two groups of four each, control or long-term hypoxic groups. For control (normoxic) group, sheep were maintained at the suppliers ranch (a 300 m elevation) on alfalfa pellets ad libitum. For the hypoxia study, at 30 days gestation, ewes were transported to the Barcroft Laboratory, White Mountain Research Station (WMRS, CA; elevation 3,801 m, barometric pressure −480 Torr), where they were kept until 135 days gestation. They were maintained in a sheltered outdoor pen and were fed with alfalfa pellets ad libitum. Sheep in both groups were kept in natural day-night conditions. In previous studies, we have obtained mean maternal arterial blood gas values from 12 adult sheep at WMRS. These were pO2 = 60±5 Torr, PCO2 = 30.0±2.5 Torr, and pH = 7.36±0.06. In contrast normoxic control sheep (elevation 346 m) had pO2 of 100±5 Torr, pCO2 35.2±0.9, pH = 7.44±0.1. Importantly, with LTH exposure, fetal arterial PO2 fell from the normoxic value of 29.7±2.1 to 19.1±2.1 Torr. At 135 days gestation, ewes from both groups were transported (4 to 7 hrs trip) to our laboratory at Loma Linda University. Soon after arrival, surgeries were conducted to isolate fetal carotid arteries. In the hypoxic group, shortly following arrival, a tracheal catheter was placed in the ewe, through which N2 flowed at a rate adjusted to maintain its PaO2 at ∼60 Torr, similar to the levels achieved at high altitude. For both groups, pregnant ewes were anesthetized with thiopental sodium (10 mg.kg−1, i.v.), and anesthesia was maintained with inhalation of 1% isoflurane in oxygen throughout surgery. The fetus was delivered by hysterotomy and following the removal of carotid arteries, the fetus and ewe was euthanized with an overdose of the proprietary euthanasia solution, Euthasol (pentobarbital sodium 100 mg.Kg−1 and phenytoin sodium 10 mg.Kg−1; Virbac, Ft. Worth, TX). Studies were performed in isolated mid-carotid artery segments cleaned of endothelium, adipose, and connective tissue.
Tissue Collection and Microarray Processing
We have described this technique in detail [16]. Ovine oligonucleotide microarrays were obtained from Agilent Technologies (Santa Clara, CA) and analysis was conducted by utilizing the commercial services of GenUs Biosystems (Northbrook, IL). Briefly, tissue samples were lysed in Tri-reagent (Ambion, Austin, TX) and total RNA was isolated using phenol/chloroform extraction, followed by purification over spin columns (Ambion). The concentration and purity of total RNA were measured by spectrophotometry at OD260/280, and the quality of the total RNA sample was assessed using an Agilent Bioanalyzer with the RNA6000 Nano Lab Chip (Agilent Technologies).
Labeled cRNA was prepared by linear amplification of the Poly(A)+ RNA population within the total RNA sample. Briefly, 1 µg of total RNA was reverse transcribed after priming with a DNA oligonucleotide containing the T7 RNA polymerase promoter 5′ to a d(T)24 sequence. After second-strand cDNA synthesis and purification of double-stranded cDNA, in vitro transcription was performed using T7 RNA polymerase. The quantity and quality of the labeled cRNA were assayed by spectrophotometry and the Agilent Bioanalyzer.
One µg of purified cRNA was fragmented to uniform size and applied to Agilent Sheep Gene Expression Microarray, 8×15K (Design ID 019921, Agilent Technologies) in hybridization buffer. Arrays were hybridized at 65°C for 17 hrs. in a shaking incubator and washed at 37°C for 1 min. Rinsed and dried arrays were scanned with an Agilent G2565 Microarray Scanner (Agilent Technologies) at 5 µm resolution. Agilent Feature Extraction software was used to process the scanned images from arrays (grid and feature intensity extraction) and the data generated for each probe on the array was analyzed with Gene Spring GX v7.3.1 software (Agilent Technologies). Annotations are based on the Agilent eArray annotation file dated January 2010. The data set has been submitted in GEO database, the accession no. Is GSE49920.
Pathway/Network Analysis
Each gene was annotated/checked manually using NCBI Blast Search, Unigene, Entrez or other databases. Genes with unknown sequences for Ovis aries were annotated if more than 90% sequence homology was identified with Bos taurus. We then analyzed the annotated genes using an Ingenuity Pathway Analysis Program (Ingenuity Systems, Redwood City, CA). Group of genes were analyzed by Meme Software Suite to examine common motifs and cis-regulatory elements [17]. Common motifs and cis-regulatory elements were analyzed by Transfac Public 6.0 to examine transcription factors [18]. The 3′ un-translated region (3′UTR) of the group of upregulated and downregulated regions were examined by Diana Lab Softwares (Athens, Greece) [19].
Real Time PCR Validation
To validate the results of the microarray analysis, we examined expression of IGFBP3, AKT1, CRABP2, ERK1, and ERK2 genes by using real time PCR. Using the same probe sequences as those on the microarray chip, we designed primers with the use of Primer 3 web-based software (Primer3 website. Available: http://frodo.wi.mit.edu/primer3/. Accessed 2013 November 1). The primers were synthesized by Integrated DNA technologies (Coralville, CA). The total RNA (1 ug per reaction) was reverse transcribed using Quantitect reverse transcriptase kit (Qiagen, Valencia, CA). Relative expression was normalized to 18S RNA and fold-changes were calculated using the ΔΔCt method with normalization of individual PCR efficiencies [20]. Samples (n = 4 from each group) were analyzed on the Roche LightCycler 1.5 (Roche, Indianapolis, IN).
Western Immunoblot Validation
Isolated carotid arteries from normoxic and hypoxic fetuses (n = 4 in each group) were cleaned of adventitia and the endothelium was denuded. These arteries were homogenized with a tissue grinder in ice-cold cell lysis buffer (Cell Signaling Technology, Danvers, MA), as we have described [21]. Protein concentrations were measured using a protein assay kit (Bio-Rad Laboratories, Hercules, CA) with bovine serum albumin (BSA) as a reference protein. A Mini Trans-Blot Electrophoretic Transfer Cell system (Bio-Rad Laboratories) was used to transfer proteins from the gel to a nitrocellulose membrane at 100 V for 3 h. We then performed an overnight incubation of AKT and IGFBP3 specific primary antibodies (1∶500 dilution). We used the total ERK as an internal control for equal protein loading, as well as the blocking peptide for each subtype specific antibody as a negative control. All antibodies were obtained from Abcam Inc. (Cambridge, MA). Earlier, we have demonstrated that total ERK levels do not change in carotid arteries in response to long-term high altitude acclimatization [22]. The membrane then was incubated in chemiluminescence luminol reagent (Pierce, Rockford, IL) for 1 min, and the protein band detected using an Alpha Innotech Chemiluminescent imaging system (San Leandro, CA).
Epiflorescence Imaging Studies Validation
By use of an Evos Florescence Microscope (Advanced Microscopy Group, Bothell, WA) using standard techniques, we examined the expression of PKNOX1. Alpha smooth muscle actin and nuclear stain Dapi (4′, 6-Diamindino-2-Phenylindole -di-Lactate) were used as controls as we have reported [23]. Briefly, arterial segments (n = 4 from each group) were sliced to 10 μ sections using a Leica Cryostat (Leica Microsystems Inc. Buffalo Grove, IL). Tissue sections were then sealed on microscopic slides with Vectashield mounting medium containing DAPI (H-1200, Vector Labs, Burlingame, CA). All antibodies were obtained from Abcam Inc. Images were analyzed using the ImageJ software (NIH), and protein expression was measured as fluorescent intensity/unit area normalized to the florescent intensity/unit area of the alpha smooth muscle actin control.
Statistics
To compare individual expression values across arrays, the raw intensity data from each gene was normalized to the 75th percentile intensity of each array. Only genes with values greater than background intensity for all samples within each group were used for further analysis. For the microarray data analysis, we used a parametric T-test that assumes variances are not equal (Welch T-test). This test also assumes that the sheep were selected from a population that follows a normal distribution of individual gene expression. To further ensure individual gene data fit a normal distribution, all T-test p-values were calculated using the means of log10 transformed values. Differentially expressed genes were identified by 2-fold change and Welch T-test [24] p-values<0.05 between each treatment group and its age-specific normoxic control. Statistical significance in the real-time PCR, western immunoblot and immuno histochemistry was determined by Students t-test (P<0.05).
Results
As a consequence of antenatal maternal long-term hypoxia, we did not observe a significant reduction in the fetal body or brain weight at 140 days of gestation (data not shown), and this was similar to our previous studies [4]. In terms of gene regulation, antenatal maternal high altitude long-term hypoxic exposure was associated with 38 genes being upregulated more than 2-fold (P<0.05; Table 1) and 9 genes downregulated more than 2-fold (P<0.05; Table 2).
Table 1. Top upregulated genes in high altitude acclimatized ovine fetal carotid arteries compared to normal control.
| Symbol | Entrez Gene Name | Fold Change | FH Mean | FH SD | FN Mean | FN SD | p-value |
| SLC16A3 | Solute carrier family 16, member 3 (monocarboxylic acid transporter 4) | 6.866 | 0.38 | 0.28 | 0.06 | 0.03 | 1.77E-02 |
| TNC | Tenascin C | 5.704 | 4.72 | 3.49 | 0.83 | 0.38 | 2.47E-02 |
| IGFBP3 | Insulin-like growth factor binding protein 3 | 5.288 | 1.20 | 0.12 | 0.23 | 0.13 | 3.04E-02 |
| TP53I11 | Tumor protein p53 inducible protein 11 | 5.133 | 0.13 | 0.08 | 0.03 | 0.02 | 4.46E-02 |
| IGF2R | Insulin-like growth factor 2 receptor | 4.906 | 0.59 | 0.41 | 0.12 | 0.04 | 2.99E-02 |
| ATF3 | Activating transcription factor 3 | 4.384 | 0.43 | 0.14 | 0.10 | 0.05 | 1.82E-02 |
| SERPINH1 | Serpin peptidase inhibitor, clade H (HSP47; collagen binding protein 1) | 4.246 | 12.89 | 5.76 | 3.03 | 0.92 | 1.13E-02 |
| PIM2 | Pim-2 oncogene | 3.489 | 0.07 | 0.01 | 0.02 | 0.01 | 3.40E-03 |
| DCAF8 | DDB1 and CUL4 associated factor 8 | 3.374 | 0.45 | 0.22 | 0.13 | 0.06 | 3.13E-02 |
| TGFB3 | Transforming growth factor, beta 3 | 2.919 | 1.43 | 0.50 | 0.49 | 0.26 | 4.56E-02 |
| MYOT | Myotilin | 2.789 | 5.77 | 1.12 | 2.07 | 0.46 | 3.98E-03 |
| DNAJC21 | DnaJ (Hsp40) homolog, subfamily C, member 21 | 2.613 | 0.23 | 0.05 | 0.09 | 0.03 | 1.38E-02 |
| CRELD2 | Cysteine-rich with EGF-like domains 2 | 2.597 | 0.43 | 0.16 | 0.17 | 0.02 | 4.07E-02 |
| BCL2 | B-cell CLL/lymphoma 2 | 2.549 | 0.85 | 0.24 | 0.33 | 0.16 | 4.83E-02 |
| GNL1 | Guanine nucleotide binding protein-like 1 | 2.52 | 0.35 | 0.10 | 0.14 | 0.06 | 4.20E-02 |
| HR | Hairless homolog (mouse) | 2.499 | 0.17 | 0.02 | 0.07 | 0.02 | 9.53E-03 |
| SNRNP70 | Small nuclear ribonucleoprotein 70 kDa (U1) | 2.485 | 1.12 | 0.46 | 0.45 | 0.10 | 3.78E-02 |
| ATF6B | Activating transcription factor 6 beta | 2.386 | 0.29 | 0.10 | 0.12 | 0.04 | 3.48E-02 |
| SURF6 | Surfeit 6 | 2.351 | 0.21 | 0.06 | 0.09 | 0.04 | 4.88E-02 |
| KDM4A | Lysine (K)-specific demethylase 4A | 2.327 | 0.42 | 0.11 | 0.18 | 0.04 | 1.17E-02 |
| ACTN1 | Actinin, alpha 1 | 2.326 | 23.45 | 2.57 | 10.08 | 1.35 | 1.24E-03 |
| DYNC1H1 | Dynein, cytoplasmic 1, heavy chain 1 | 2.307 | 3.16 | 0.49 | 1.37 | 0.53 | 4.59E-02 |
| MED16 | Mediator complex subunit 16 | 2.264 | 0.14 | 0.05 | 0.06 | 0.02 | 4.97E-02 |
| ADIPOR1 | Adiponectin receptor 1 | 2.247 | 0.39 | 0.08 | 0.17 | 0.05 | 2.31E-02 |
| DNAJB5 | DnaJ (Hsp40) homolog, subfamily B, member 5 | 2.247 | 1.24 | 0.47 | 0.55 | 0.12 | 4.20E-02 |
| MTMR2 | Myotubularin related protein 2 | 2.231 | 0.22 | 0.08 | 0.10 | 0.03 | 4.78E-02 |
| MTCH1 | Mitochondrial carrier 1 | 2.224 | 5.12 | 1.75 | 2.30 | 0.35 | 3.63E-02 |
| CDV3 | CDV3 homolog (mouse) | 2.206 | 0.46 | 0.15 | 0.21 | 0.07 | 4.51E-02 |
| KHSRP | KH-type splicing regulatory protein | 2.153 | 0.27 | 0.07 | 0.13 | 0.02 | 1.34E-02 |
| RBM42 | RNA binding motif protein 42 | 2.153 | 1.91 | 0.50 | 0.89 | 0.30 | 3.58E-02 |
| ATG4B | ATG4 autophagy related 4 homolog B (S. cerevisiae) | 2.119 | 1.46 | 0.46 | 0.69 | 0.10 | 3.55E-02 |
| WASF2 | WAS protein family, member 2 | 2.054 | 0.40 | 0.09 | 0.19 | 0.05 | 1.88E-02 |
| COL18A1 | Collagen, type XVIII, alpha 1 | 2.05 | 6.00 | 0.60 | 2.93 | 0.54 | 1.45E-02 |
| PLXNB1 | Plexin B1 | 2.047 | 0.56 | 0.10 | 0.27 | 0.09 | 4.24E-02 |
| COX7B | Cytochrome c oxidase subunit VIIb | 2.045 | 0.64 | 0.15 | 0.31 | 0.09 | 3.01E-02 |
| ANXA11 | Annexin A11 | 2.026 | 0.93 | 0.21 | 0.46 | 0.16 | 4.46E-02 |
| FADS2 | Fatty acid desaturase 2 | 2.023 | 0.59 | 0.20 | 0.29 | 0.06 | 4.68E-02 |
| AKT1 | v-akt murine thymoma viral oncogene homolog 1 | 2.019 | 5.00 | 0.52 | 2.48 | 0.52 | 1.41E-02 |
Table 2. Top downregulated genes in high altitude acclimatized fetal carotid arteries compared to normal control.
| Symbol | Entrez Gene Name | Fold Change | FH Mean | FH SD | FN Mean | FN SD | p-value |
| BRB/BRN-2 | Brain Ribonucleas | −3.63 | 4.10 | 0.73 | 14.91 | 6.33 | 0.01 |
| ALOX5AP | Arachidonate 5-lipoxygenase-activating protein | −3.247 | 0.11 | 0.03 | 0.37 | 0.04 | 0.007 |
| RNASE6 | Ribonuclease, RNase A family, k6 | −2.924 | 0.38 | 0.04 | 1.12 | 0.44 | 0.039 |
| TRA@ | T cell receptor alpha locus | −2.625 | 0.11 | 0.04 | 0.30 | 0.11 | 0.023 |
| CRABP2 | Cellular retinoic acid binding protein 2 | −2.584 | 0.16 | 0.07 | 0.42 | 0.08 | 0.039 |
| RPL35 | Ribosomal protein L35 | −2.257 | 1.87 | 0.19 | 4.21 | 0.42 | 0.003 |
| RGS10 | Regulator of G-protein signaling 10 | −2.123 | 0.16 | 0.06 | 0.34 | 0.07 | 0.043 |
| BATF3 | Basic leucine zipper transcription factor, ATF-like 3 | −2.096 | 0.10 | 0.03 | 0.20 | 0.06 | 0.038 |
| FAM35A | Family with sequence similarity 35, member A | −2.008 | 0.30 | 0.06 | 0.59 | 0.11 | 0.013 |
As shown in Table 3, based on Ingenuity Pathway Analysis, the major functional networks altered were those associated with cell survival, cellular development, cellular growth and proliferation, cell morphology, cardiovascular system development and function, and hematological system development and function. Also, shown are the five major canonical pathways significantly altered because of antenatal maternal LTH acclimatization.
Table 3. Chief Functional and Canonical Pathways Altered by Long-Term Hypoxia.
| Functional Pathways | Molecules |
| Cell Survival | ATF3, ATG4B, IGF2R, ADIPOR1, BCL2, DYNC1H1, AKT1, CRABP2, RGS10, IGFBP3, TGFB3, PLXNB1, COL18A1, TRA@, PIM2 |
| Cellular Development | ATF3, TNC, IGF2R, BCL2, DYNC1H1, AKT1, BATF3, RGS10, TGFB3, IGFBP3, PLXNB1, WASF2, COL18A1, TRA@ |
| Cellular Growth and Proliferation | ATF3, TNC, SURF6, TP53I11, MTCH1, HR, ADIPOR1, IGF2R, BCL2, DYNC1H1, AKT1, ANXA11, SERPINH1, CRABP2, TGFB3, FADS2, IGFBP3, PLXNB1, WASF2, COL18A1, ACTN1, TRA@, PIM2 |
| Cell Morphology | ATF3, TNC, AKT1, SERPINH1, ATG4B, TGFB3, IGFBP3, WASF2, COL18A1, TRA@, PIM2, BCL2 |
| Cardiovascular System Development and Function | TNC, ATF3, AKT1, SERPINH1, TGFB3, IGFBP3, PLXNB1, WASF2, COL18A1, IGF2R, PIM2, BCL2 |
| Hematological System Development and Function | TNC, ATF3, AKT1, BATF3, RGS10, PLXNB1, WASF2, IGF2R, TRA@, PIM2, BCL2 |
| Canonical Pathways | Molecules |
| VEGF Signaling | AKT1,ACTN1,BCL2 |
| PTEN Signaling | AKT1, IGF2R, BCL2 |
| PI3K Signaling in B Lymphocytes | AKT1, ATF3, ATF6B |
| TGF-β Signaling | TGFB3, BCL2 |
| PI3K/AKT Signaling | AKT1, BCL2 |
As shown in Figure 1, analysis of the genes involved in the altered functional and canonical pathways belong to two major networks of genes namely AKT1 and B-cell lymphoma 2 (BCL2) pathways. To confirm the findings of microarray data, few genes with moderate changes in fold-change were examined with real-time PCR analysis. Real-time PCR analysis, demonstrated that Insulin like growth factor binding protein 3 (IGFBP3) was upregulated 3.6±1.2 and AKT was upregulated 2.1±0.3 - fold and were similar to the findings in the microarray analysis. ERK1 and 2 did not show any significant changes with LTH with real-time PCR or microarray analysis, this was further confirmed with western immunoblot analysis and used a loading control. Also, with real-time PCR, CRABP2 showed 2.3±0.4 -fold downregulation on long-term hypoxic exposure. Similarly, total ERK (TERK) and α-smooth muscle actin showed no significant change on western immunoblot (Figure 2) and immuno histochemistry (Figure 3) analysis. Further validation of AKT (Figure 2A) and IGFBP3 (Figure 2B) demonstrated an increased level of expression by western immunoblot. Additionally, PKNOX analysis by immuno histochemistry (Figure 3) analysis demonstrated a significantly increased expression.
Figure 1. Demonstrates the network of genes in ovine fetal carotid arteries extra-cellular, plasma membrane, cytoplasm, and nucleus altered as a consequence of antenatal maternal long-term hypoxia.

Red color represents the upregulated genes; the higher intensity of color denotes greater fold-change; green represents the downregulated genes.
Figure 2. Demonstrates protein expression of (A) AKT and (B) IGFBP3. N = 4 in each group; Total extracellular regulated kinase (TERK) served as an internal control to which AKT and IGFBP3 were normalized to provide relative integrated density (IDV).
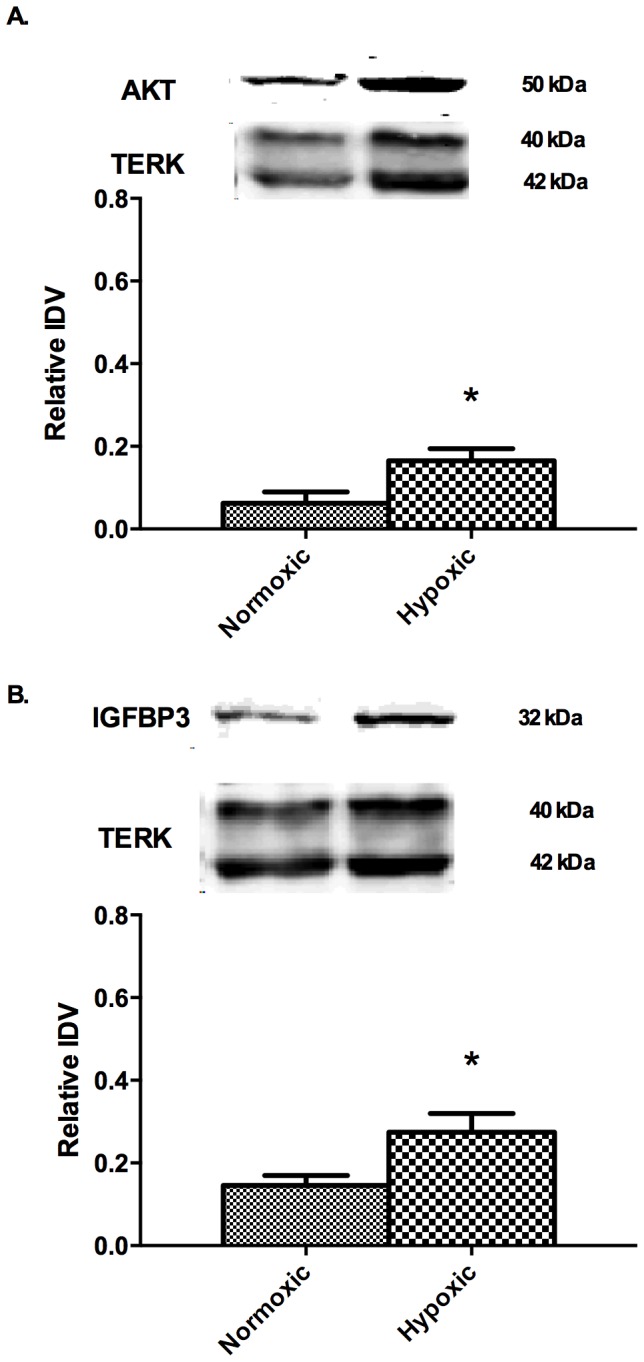
* Denotes significant changes in protein expression by Students t-test (P<0.05); FN - Carotid arteries from control (normoxic) fetus, FH - Carotid arteries from antenatal long-term hypoxia acclimatized animals.
Figure 3. Demonstrates immunohistochemistry images of PKNOX1 expression in carotid arteries from normoxic fetus (FN) and hypoxic fetuses (FH).
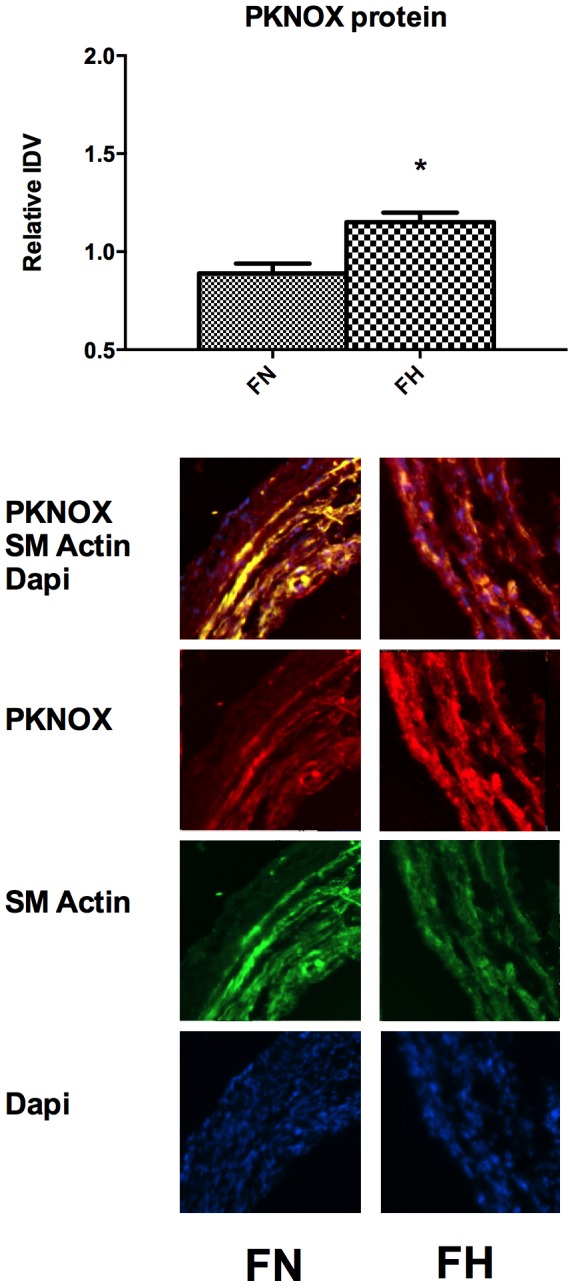
*Denotes statistically significant difference by Students t-test (P<0.05).
Analysis of 1800 bp upstream of transcription start site (TSS) of up- and downregulated genes
As shown in Figure 4, we found a 41 bp common motif in the upregulated gene sequence. Notably, TGFB and Pbx/Knotted 1 Homeobox binding sites are present in the promoters of a number of upregulated genes. The promoter regions of the 9 downregulated genes demonstrate a common motif sequence of 49 bp (Figure 5). In this 49 bp region, we identified binding sites for transcription factors DOT6 and TOD6, components of the RPD3 histone deacetylase complex RPD3C(L) responsible for the deacetylation of lysine residues on the N-terminal part of the core histones (H2A, H2B, H3 and H4).
Figure 4. Demonstrates a common motif in 1800 base pairs upstream to the transcription start site of the upregulated genes.

Strand + and − denotes the sense and antisense DNA sequence. The start denotes the number of base pair upstream to the transcriptional start site. p-value denotes the probability of the presence of motif by chance in the given sequence as a part of group of sequence altered by long-term hypoxic exposure. The bottom of the figure shows a cis-regulatory element for Pbx/knotted 1 homeobox transcription factor, which can bind on the common motif of the genes shown above. Bits refer to the likely hood of the presence of a particular letter of a specific motif, at a specific site, in more number of background sequences of genes altered by long-term hypoxia.
Figure 5. Demonstrates a common motif in 1800 base pairs upstream to the transcription start site of the downregulated genes.

The bottom of the figure shows a cis-regulatory element for TOD6/DOT6 transcription factor, which can bind on the common motif of the genes shown above.
Analysis of the 3′ un-translated region of the altered genes
As shown in Figure 6, the 3′UTR of the upregulated genes demonstrated binding sites for the Sox family of genes. To our surprise, we were unable to identify any transcription factor binding site in the 3′ UTR common to the downregulated genes (data not shown).
Figure 6. Demonstrates a common motif in the 3′ untranslated region of the upregulated genes.

The bottom of the figure shows a cis-regulatory element for Sox family transcription factors, which can bind on the common motif of the genes shown above.
Analysis of the miRNA binding sites in the 3′UTR of the altered genes
Our analysis also demonstrated binding sites for several miRNA complementary to groups of upregulated and downregulated genes (Tables S1 & S2).
Discussion
The present report is a continuation of our studies to understand fundamental mechanisms of LTH acclimatization responses in fetal sheep. In the present study, we identified a number of key genes, cis-regulatory elements, and miRNA that may play an important role in successful acclimatization of the ovine fetal cranial vasculature (and maybe other tissues) in association with exposure to in-utero long-term hypoxia (110 days). As noted, humans, rodents, and many other species show adverse effects in response to moderate prolonged high-altitude hypoxic exposure, whereas sheep appear to be more resilient. The mechanisms by which sheep and some other species successfully acclimatize to high-altitude LTH is not well-understood. Of importance, this may have implications in understanding these (responses, mechanisms) in humans to prevent of modify diseases such as acute/chronic mountain sickness, high altitude associated cerebral edema, or other conditions. Although a number of papers have reported on gene regulation in response to “chronic” hypoxia, almost without exception these exposures have been for a duration of only 48 hours or less [25]–[30]. Also, many of these studies are have been in cancer cell lines and demonstrate an increase in hypoxia inducible factor (HIF) 1α. Of critical importance, it is well documented that with chronic hypoxia HIF1α levels return to normoxic values within several days [31]. Similar to this study, we did not observed significant up-regulation of HIF1α with long-term hypoxia in the present study. Along this line, we have demonstrated that erythropoietin levels increased from 22.8±2.2 to 144±37 mu.ml−1 within 24 h; however, by day 7 it had returned to levels slightly above normal [32].
Also, our group has reported on alterations in fetal gene expression in response to LTH. In perirenal adipose tissue, these include: uncoupling protein 1, 11β hydroxy steroid dehydrogenase type 1, peroxisome proliferator activated receptor gamma coactivator 1, and deiodenase type 1 and 2 receptors [33]. In fetal adrenal zona fasciculata/reticularis these also include: nitric oxide synthase [34] phenylethanolamine-N methyltranferase, tyrosine hydroxylase, and dopamine β-hydroxylase [35]. Thus, LTH exposure during fetal life has a profound effect in modulating the expression of enzymes that mediate adrenomedullary catecholamine synthesis. In addition, these LTH-mediated responses include epigenetic-mediated repression of the estrogen receptor α gene in the uterine arteries [36]. Importantly, many of these genes are unaltered on hypoxia exposure in fetal carotid arteries, demonstrating tissue specific changes in gene expression.
This is first study to our knowledge to examine the effect of high-altitude (3801 m) long-term (110 days) hypoxia in-utero on global gene expression in the fetal vascular system. Importantly, we conducted this study in sheep that successfully acclimatize to LTH, escaping many of the problems associated with LTH exposure in humans and other species. For instance, in response to long-term (110 days) in-utero hypoxic exposure, sheep does not demonstrate intrauterine growth restriction, reduction in brain weight, alteration in CBF, and/or pulmonary hypertension. Therefore, the present study is of fundamental importance in identifying signaling molecules and pathways that play a protective role in acclimatization to high altitude LTH in the fetus of this species.
In the present study, we observed an increase in the gene expression for Insulin-like growth factor 2 receptor (IGF2R) and IGFBP3. A study in sheep with placental blood flow restriction by carunclectomy with superimposed hypoxia demonstrated IUGR with a failure to increase IGF2R [37]. In contrast, in the present study, we observed an increase in IGF2R without development of IUGR. We speculate that in the sheep fetus, IGF2R may be a vital factor providing protection hypoxia induced dysregulation in cerebral blood flow and brain growth restriction. Similarly, IGFBP3 may also be an important factor in prevention of dysregulated blood flow in the LTH exposed fetal sheep. Also, human studies demonstrate a significant lower IGFBP3 in growth restricted infants [38] as well as a significant increase in IGFBP3 with increased growth on treatment with growth hormone [39]. Thus, IGF2R and IGFBP3 both appear to be associated with protection from growth restriction in the sheep fetuses. Further investigations of these mechanisms are required, however.
We also observed upregulation of three members of the heat shock protein (HSP) 40 family. These were serpin peptidase inhibitor, DnaJ homolog subfamily C member 21, and DnaJ homolog subfamily B member 5 (Table 1). DnaJ/HSP40 proteins are conserved among phyla. They appear to play an important role in protein translation, folding, unfolding, translocation, and degradation, primarily by stimulating the ATPase activity of HSP70 chaperone proteins [40]. HSP47 is a 47 kDa collagen-binding glycoprotein localized in the endoplasmic reticulum. Gene ablation studies have indicated that HSP47 is essential for embryonic development and the maturation of several types of collagen [41]. However, further studies are needed to elucidate the role of these heat shock proteins in hypoxic acclimatization.
Also, in the present study, activating transcription factors (ATF) 3 and 6 were upregulated (Table 1). The mammalian ATF family of transcription factors represents a large group of basic region-leucine zipper (bZip) proteins; these can bind to cAMP-inducible transcription sites at a consensus sequence TGACGT(C/A)(G/A) [42]. These ATFs are important in stress signaling [43]. The present study suggests that these two factors may be important in fetal growth under hypoxic stress. Again, further investigation along this line is needed.
We also observed significant upregulation of solute carrier family 16 member 3 (Table 1), commonly known as monocarboxylic acid transporter 4 (MCT4). MCT4 (SLC16A3) is involved in the transport of metabolically important monocarboxylates such as lactate, pyruvate, acetate, and ketone bodies, and is important for lactate excretion to the extracellular space. It also assumes an important role during hypoxic stress [44]. Previously, upregulation of MCT4 by hypoxia has been demonstrated in the human bladder cancer transcriptome [45].
Other important genes upregulated with long-term hypoxia include AKT1, BCL2, and TGFβ3 (Table 1; Figure 1). AKT1 has been shown to be crucial in cell survival on hypoxic exposure, and failure to up-regulate AKT1 is involved in hypoxia-induced cell death [46]. Of importance, AKT1 knockout mice are growth restricted [47]. Thus, AKT1 may be another important protective molecule playing a role in normal birth weight of newborn lambs, despite their long-term in-utero hypoxic exposure. Additionally, B-cell lymphoma 2 (BCL2) proteins are critical regulators of cell death, whose main function is to regulate the release of cytochrome c from mitochondria in the intrinsic apoptotic pathway [48]. Importantly, BCL2 is known to be reduced in intrauterine growth restriction [49], and upregulation of BCL2 in our studies may play a protective role against LTH for fetal lambs. Moreover, as shown by other studies, BCL2 upregulation is important in cell survival during prolonged hypoxic exposure [50]. Similarly, TGFβ3 has been shown to be upregulated with hypoxia [51]. The role of these proteins in preventing adverse long-term hypoxia effects needs to be addressed.
In contrast to upregulation, we found a limited number of genes to be downregulated because of antenatal LTH exposure (Table 2). These included brain ribonucleas (BRN-2), which has been shown to be downregulated in perinatal asphyxia in rat brain [52]. We also observed significant reduction in arachndonate 5-lipoxygenase-activating protein (ALOX5AP). ALOX5AP is a non-heme iron containing dioxygenase which incorporates oxygen into arachidonic acid and mediates leukotriene formation [53]. This gene has been shown to be induced by hypoxia and is important in chronic hypoxia-induced vascular remodeling and pulmonary hypertension [53], [54]. Instead of upregulation, as observed in the quoted studies, we observed LTH-induced downregulation of ALOX5AP. In addition, this contrast, with our antenatal maternal LTH exposed newborn lambs, in which pulmonary hypertension did not develop [55]. It appears that sheep can alter multiple molecules, which protect the fetus from reduced brain growth dysregulation of CBF, as well as pulmonary hypertension.
Another important gene downregulated with LTH was cellular retinoic acid binding protein 2 (CRABP2; Table 2). This chaperone protein can bind intracellular retinoic acid with high affinity, and translocate it to the nucleus to induce cellular differentiation [56]. CRABP2 downregulation, may relieve the negative inhibition of retinoic acid signaling on angiogenesis [57]. We also observed a significant downregulation of ribosomal protein L35 (RPL35), a conserved protein in eukaryotes important in ribosome biogenesis [58]. Reduced levels of RPL35 are associated with delay of the G1 phase of cell cycle [58]. Its exact role in long-term hypoxia is not known, however.
Importantly, the question arises, by what mechanisms does LTH alter so many genes in a highly orchestrated manner? Through common cis-regulatory elements and trans-acting factors, prolonged hypoxia appears to lead to the upregulation of several of these genes belonging to different canonical and functional pathways. To address this issue, we examined promoters and 3′ UTRs of the altered genes to find common cis-regulatory elements and sites complementary to miRNAs. We have identified the Pbx/Knotted homeobox binding site in the promoter region of a number of upregulated genes. This agrees with the finding in skeletal muscle, in which chronic hypoxia leads to conversion of fast to slow fibers with increased expression of Pbx/Knotted homeobox [59]. Pbx also has been shown to be involved in cardiac muscle differentiation [60]. Moreover, Pbx1 silencing impairs endothelial cell migration and blocks angiogenesis [61]. Also, in the 3′ UTR of the number of upregulated genes, we observed several binding sites for the Sox family transcription factors important in the development of fetal vasculogenesis [62] and postnatal angiogenesis [63]. Further experiments are needed, however, to establish the role of Pbx1 in the regulation of angiogenesis pathways such as VEGF and PI3/AKT, and their role in LTH acclimatization. The present report provides strong rationale for such studies.
Furthermore, in the promoter region of the downregulated genes we observed binding sites for DOT6 and TOD6, components of the RPD3 histone deacetylase complex RPD3C(L) responsible for the deacetylation of lysine residues on the N-terminal part of the chromatin core histones (H2A, H2B, H3 and H4) [64]. Of importance, histone deacetylation may lead to epigenetic repression and play an important role in transcriptional regulation, cell cycle progression, and developmental events. DOT6 and TOD6 bind to sequences containing the core CGATG, which resembles the PAC (Polymerase A and C) motif [64]. Thus, these studies indicate that chronic hypoxia may lead to DOT6-mediated histone deacetylation to achieve gene downregulation. These findings also require further investigation, however.
We also analyzed the up- and downregulated genes for putative miRNA binding sites (Table S1 & Table S2). We have identified miRNA with putative binding sites to a number of altered genes. Again, confirmation of these miRNA will require future studies.
Speculation and Perspective
On exposure to antenatal long-term hypoxia, humans and other species demonstrate fetal intrauterine growth restriction, dysregulation of CBF, pulmonary hypertension, and other changes. Sheep does not, however. Also, previously, we have shown that in response to in-utero LTH exposure, fetal lambs develop neither dysregulation of cerebral blood flow nor pulmonary hypertension. The present study provides insight into these mechanisms. However, the genes altered may simply be result of long-term hypoxic exposure and may be detrimental instead of adaptive. The limitation of the present study is that it does not causally examine the effect of the microarray data with functional implications. However, such studies are beyond the scope of the present study and need to be confirmed independently. The present study provides a number of molecules associated with acclimatization to high altitude as possible for such studies
Supporting Information
Putative miRNA complementary to the 3′ UTR of the upregulated genes in high altitude acclimatized fetal carotid arteries compared to normal control.
(DOC)
Putative miRNA complementary to the 3′ UTR of the downregulated genes in high altitude acclimatized fetal carotid arteries compared to normal control.
(DOC)
Acknowledgments
We would like to thank Nina Chu, BS for help with tissue collection.
Funding Statement
The study was supported by United States Public Health Service Grant PO1 HD031226 to LDL. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Reece EA, Hobbins JC (2008) Clinical Obstetrics. Malden, MA: Wiley-Blackwell. 1 pp. [Google Scholar]
- 2. Rueda-Clausen CF, Morton JS, Davidge ST (2008) Effects of hypoxia-induced intrauterine growth restriction on cardiopulmonary structure and function during adulthood. Cardiovasc Res 81: 713–722 10.1093/cvr/cvn341 [DOI] [PubMed] [Google Scholar]
- 3. Kamitomo M, Longo LD, Gilbert RD (1992) Right and left ventricular function in fetal sheep exposed to long-term high-altitude hypoxemia. Am J Physiol 262: H399–H405. [DOI] [PubMed] [Google Scholar]
- 4. Kitanaka T, Alonso JG, Gilbert RD, Siu BL, Clemons GK, et al. (1989) Fetal responses to long-term hypoxemia in sheep. Am J Physiol 256: R1348–R1354. [DOI] [PubMed] [Google Scholar]
- 5. Metcalfe J, Meschia G, HELLEGERS A, PRYSTOWSKY H, HUCKABEE W, et al. (1962) Observations on the growth rates and organ weights of fetal sheep at altitude and sea level. Q J Exp Physiol Cogn Med Sci 47: 305–313. [DOI] [PubMed] [Google Scholar]
- 6. McCullough RE, Reeves JT, Liljegren RL (1977) Fetal growth retardation and increased infant mortility at high altitude. Obstet Gynecol Surv 32: 596–598. [DOI] [PubMed] [Google Scholar]
- 7. Unger C, Weiser JK, McCullough RE, Keefer S, Moore LG (1988) Altitude, low birth weight, and infant mortality in Colorado. Jama 259: 3427–3432 10.1001/jama.1988.03720230037027 [DOI] [PubMed] [Google Scholar]
- 8. Moore LG (2003) Fetal Growth Restriction and Maternal Oxygen Transport during High Altitude Pregnancy. High Alt Med Biol 4: 141–156 10.1089/152702903322022767 [DOI] [PubMed] [Google Scholar]
- 9. Pena JP, Tomimatsu T, Hatran DP, McGill LL, Longo LD (2007) Cerebral blood flow and oxygenation in ovine fetus: responses to superimposed hypoxia at both low and high altitude. The Journal of Physiology 578: 359–370 10.1113/jphysiol.2006.119925 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Lee SJ, Hatran DP, Tomimatsu T, Pena JP, McAuley G, et al. (2009) Fetal cerebral blood flow, electrocorticographic activity, and oxygenation: responses to acute hypoxia. The Journal of Physiology 587: 2033–2047 10.1113/jphysiol.2009.166983 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Heistad DD, Marcus ML, Abboud FM (1978) Role of large arteries in regulation of cerebral blood flow in dogs. J Clin Invest 62: 761–768 10.1172/JCI109187 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Dieckhoff D, Kanzow E (1969) On the location of the flow resistance in the cerebral circulation. Pflugers Archiv : European journal of physiology 310: 75–85. [DOI] [PubMed] [Google Scholar]
- 13. Kontos HA, Wei EP, Navari RM, Levasseur JE, Rosenblum WI, et al. (1978) Responses of cerebral arteries and arterioles to acute hypotension and hypertension. Am J Physiol 234: H371–H383. [DOI] [PubMed] [Google Scholar]
- 14. Szymonowicz W, Walker AM, Yu VY, Stewart ML, Cannata J, et al. (1990) Regional cerebral blood flow after hemorrhagic hypotension in the preterm, near-term, and newborn lamb. Pediatr Res 28: 361–366. [DOI] [PubMed] [Google Scholar]
- 15. Kehrer M, Blumenstock G, Ehehalt S, Goelz R, Poets C, et al. (2005) Development of cerebral blood flow volume in preterm neonates during the first two weeks of life. Pediatr Res 58: 927–930 10.1203/01.PDR.0000182579.52820.C3 [DOI] [PubMed] [Google Scholar]
- 16. Goyal R, Longo LD (2012) Gene Expression in Sheep Carotid Arteries: Major Changes with Maturational Development. Pediatr Res 72: 137–146 10.1038/pr.2012.57 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Bailey TL, Boden M, Buske FA, Frith M, Grant CE, et al. (2009) MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res 37: W202–W208 10.1093/nar/gkp335 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Matys V, Kel-Margoulis OV, Fricke E, Liebich I, Land S, et al. (2006) TRANSFAC and its module TRANSCompel: transcriptional gene regulation in eukaryotes. Nucleic Acids Res 34: D108–D110 10.1093/nar/gkj143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Vlachos IS, Kostoulas N, Vergoulis T, Georgakilas G, Reczko M, et al. (2012) DIANA miRPath v.2.0: investigating the combinatorial effect of microRNAs in pathways. Nucleic Acids Res 40: W498–W504 10.1093/nar/gks494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Ramakers C, Ruijter JM, Deprez RHL, Moorman AFM (2003) Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci Lett 339: 62–66. [DOI] [PubMed] [Google Scholar]
- 21. Goyal R, Mittal A, Chu N, Zhang L, Longo LD (2010) alpha(1)-Adrenergic receptor subtype function in fetal and adult cerebral arteries. Am J Physiol Heart Circ Physiol 298: H1797–H1806 10.1152/ajpheart.00112.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Goyal R, Mittal A, Chu N, Arthur RA, Zhang L, et al. (2010) Maturation and long-term hypoxia-induced acclimatization responses in PKC-mediated signaling pathways in ovine cerebral arterial contractility. Am J Physiol Regul Integr Comp Physiol 299: R1377–R1386 10.1152/ajpregu.00344.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Goyal R, Henderson DA, Chu N, Longo LD (2012) Ovine Middle Cerebral Artery Characterization and Quantification of Ultrastructure and Other Features: Changes with Development. AJP: Regulatory, Integrative and Comparative Physiology 302: R433–R445 10.1152/ajpregu.00519.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Zhang X, Lin P, Zhu Z-H, Long H, Wen J, et al. (2009) Expression profiles of early esophageal squamous cell carcinoma by cDNA microarray. Cancer Genetics and Cytogenetics 194: 23–29 10.1016/j.cancergencyto.2009.04.027 [DOI] [PubMed] [Google Scholar]
- 25. Kourembanas S, Morita T, Liu Y, Christou H (1997) Mechanisms by which oxygen regulates gene expression and cell-cell interaction in the vasculature. Kidney Int 51: 438–443. [DOI] [PubMed] [Google Scholar]
- 26. van Malenstein H, Verslype C, Windmolders P, van Eijsden R, Nevens F, et al. (2012) Characterization of a cell culture model for clinically aggressive hepatocellular carcinoma induced by chronic hypoxia. Cancer Lett 315: 178–188 10.1016/j.canlet.2011.09.039 [DOI] [PubMed] [Google Scholar]
- 27. Geiger K, Leiherer A, Muendlein A, Stark N, Geller-Rhomberg S, et al. (2011) Identification of hypoxia-induced genes in human SGBS adipocytes by microarray analysis. PLoS ONE 6: e26465 10.1371/journal.pone.0026465.t002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Dukhande VV, Sharma GC, Lai JCK, Farahani R (2011) Chronic hypoxia-induced alterations of key enzymes of glucose oxidative metabolism in developing mouse liver are mTOR dependent. Mol Cell Biochem 357: 189–197 10.1007/s11010-011-0889-z [DOI] [PubMed] [Google Scholar]
- 29. Guadall A, Orriols M, Rodríguez-Calvo R, Calvayrac O, Crespo J, et al. (2011) Fibulin-5 is up-regulated by hypoxia in endothelial cells through a hypoxia-inducible factor-1 (HIF-1α)-dependent mechanism. 286: 7093–7103 10.1074/jbc.M110.162917 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Brown ST, Reyes EP, Nurse CA (2011) Chronic hypoxia upregulates adenosine 2a receptor expression in chromaffin cells via hypoxia inducible factor-2α: role in modulating secretion. Biochem Biophys Res Commun 412: 466–472 10.1016/j.bbrc.2011.07.122 [DOI] [PubMed] [Google Scholar]
- 31. Ginouvès A, Ilc K, Macías N, Pouysségur J, Berra E (2008) PHDs overactivation during chronic hypoxia “desensitizes” HIFalpha and protects cells from necrosis. PNAS 105: 4745–4750 10.1073/pnas.0705680105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Kitanaka T, Gilbert RD, Longo LD (1989) Maternal responses to long-term hypoxemia in sheep. Am J Physiol 256: R1340–R1347. [DOI] [PubMed] [Google Scholar]
- 33. Myers DA, Hanson K, Mlynarczyk M, Kaushal KM, Ducsay CA (2008) Long-term hypoxia modulates expression of key genes regulating adipose function in the late-gestation ovine fetus. Am J Physiol Regul Integr Comp Physiol 294: R1312–R1318 10.1152/ajpregu.00004.2008 [DOI] [PubMed] [Google Scholar]
- 34. Monau TR, Vargas VE, King N, Yellon SM, Myers DA, et al. (2009) Long-term hypoxia increases endothelial nitric oxide synthase expression in the ovine fetal adrenal. Reprod Sci 16: 865–874 10.1177/1933719109336678 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Ducsay CA, Hyatt K, Mlynarczyk M, Root BK, Kaushal KM, et al. (2007) Long-term hypoxia modulates expression of key genes regulating adrenomedullary function in the late gestation ovine fetus. Am J Physiol Regul Integr Comp Physiol 293: R1997–R2005 10.1152/ajpregu.00313.2007 [DOI] [PubMed] [Google Scholar]
- 36. Dasgupta C, Chen M, Zhang H, Yang S, Zhang L (2012) Chronic hypoxia during gestation causes epigenetic repression of the estrogen receptor-α gene in ovine uterine arteries via heightened promoter methylation. Hypertension 60: 697–704 10.1161/HYPERTENSIONAHA.112.198242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Gentili S, Morrison JL, McMillen IC (2009) Intrauterine growth restriction and differential patterns of hepatic growth and expression of IGF1, PCK2, and HSDL1 mRNA in the sheep fetus in late gestation. Biology of Reproduction 80: 1121–1127 10.1095/biolreprod.108.073569 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Giudice LC, de Zegher F, Gargosky SE, Dsupin BA, las Fuentes de L, et al. (1995) Insulin-like growth factors and their binding proteins in the term and preterm human fetus and neonate with normal and extremes of intrauterine growth. J Clin Endocrinol Metab 80: 1548–1555. [DOI] [PubMed] [Google Scholar]
- 39. Cabrol S, Perin L, Colle M, Coutant R, Jésuran-Perelroizen M, et al. (2011) Evolution of IGF-1 in children born small for gestational age and with growth retardation, treated by growth hormone adapted to IGF-1 levels after 1 Year. Horm Res Paediatr 76: 419–427 10.1159/000334651 [DOI] [PubMed] [Google Scholar]
- 40. Qiu X-B, Shao Y-M, Miao S, Wang L (2006) The diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70 chaperones. Cell Mol Life Sci 63: 2560–2570 10.1007/s00018-006-6192-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Ishida Y, Kubota H, Yamamoto A, Kitamura A, Bächinger HP, et al. (2006) Type I collagen in Hsp47-null cells is aggregated in endoplasmic reticulum and deficient in N-propeptide processing and fibrillogenesis. Mol Biol Cell 17: 2346–2355 10.1091/mbc.E05-11-1065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Lin YS, Green MR (1988) Interaction of a common cellular transcription factor, ATF, with regulatory elements in both E1a- and cyclic AMP-inducible promoters. PNAS 85: 3396–3400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Hai T, Hartman MG (2001) The molecular biology and nomenclature of the activating transcription factor/cAMP responsive element binding family of transcription factors: activating transcription factor proteins and homeostasis. Gene 273: 1–11. [DOI] [PubMed] [Google Scholar]
- 44. Ullah MS, Davies AJ, Halestrap AP (2006) The plasma membrane lactate transporter MCT4, but not MCT1, is up-regulated by hypoxia through a HIF-1alpha-dependent mechanism. J Biol Chem 281: 9030–9037 10.1074/jbc.M511397200 [DOI] [PubMed] [Google Scholar]
- 45. Ord JJ, Streeter EH, Roberts ISD, Cranston D, Harris AL (2005) Comparison of hypoxia transcriptome in vitro with in vivo gene expression in human bladder cancer. Br J Cancer 93: 346–354 Available:http://www.nature.com/doifinder/10.1038/sj.bjc.6602666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Box AH, Kim S-M, Demetrick DJ (2010) AKT loss in human epithelial cells treated with severe hypoxia. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 1803: 951–959 10.1016/j.bbamcr.2010.03.011 [DOI] [PubMed] [Google Scholar]
- 47. Yang Z-Z, Tschopp O, Hemmings-Mieszczak M, Feng J, Brodbeck D, et al. (2003) Protein kinase B alpha/Akt1 regulates placental development and fetal growth. J Biol Chem 278: 32124–32131 10.1074/jbc.M302847200 [DOI] [PubMed] [Google Scholar]
- 48. Luo X, Budihardjo I, Zou H, Slaughter C, Wang X (1998) Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell 94: 481–490 10.1016/S0092-8674(00)81589-5 [DOI] [PubMed] [Google Scholar]
- 49. Ninomiya M, Numakawa T, Adachi N, Furuta M, Chiba S, et al. (2010) Cortical neurons from intrauterine growth retardation rats exhibit lower response to neurotrophin BDNF. Neurosci Lett 476: 104–109 10.1016/j.neulet.2010.03.082 [DOI] [PubMed] [Google Scholar]
- 50. Lee S-D, Kuo W-W, Wu C-H, Lin Y-M, Lin JA, et al. (2006) Effects of short- and long-term hypobaric hypoxia on Bcl2 family in rat heart. International Journal of Cardiology 108: 376–384 10.1016/j.ijcard.2005.05.046 [DOI] [PubMed] [Google Scholar]
- 51. Yinon Y, Nevo O, Xu J, Many A, Rolfo A, et al. (2008) Severe intrauterine growth restriction pregnancies have increased placental endoglin levels: hypoxic regulation via transforming growth factor-beta 3. Am J Pathol 172: 77–85 10.2353/ajpath.2008.070640 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Lubec B, Labudova O, Hoeger H, Kirchner L, Lubec G (2002) Expression of transcription factors in the brain of rats with perinatal asphyxia. Biol Neonate 81: 266–278. [DOI] [PubMed] [Google Scholar]
- 53. Gonsalves CS, Kalra VK (2010) Hypoxia-mediated expression of 5-lipoxygenase-activating protein involves HIF-1alpha and NF-kappaB and microRNAs 135a and 199a-5p. J Immunol 184: 3878–3888 10.4049/jimmunol.0902594 [DOI] [PubMed] [Google Scholar]
- 54. Voelkel NF, Tuder RM, Wade K, Höper M, Lepley RA, et al. (1996) Inhibition of 5-lipoxygenase-activating protein (FLAP) reduces pulmonary vascular reactivity and pulmonary hypertension in hypoxic rats. J Clin Invest 97: 2491–2498 10.1172/JCI118696 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Wilson SM, Papamatheakis DG, Terry MR, Merritt T, Merrill-Henry J, et al. (2010) In utero hypoxia augments hypoxic pulmonary vasoconstriction in newborn lamb. The FASEB Journal 24: 1023.19940258 [Google Scholar]
- 56. Campos B, Warta R, Chaisaingmongkol J, Geiselhart L, Popanda O, et al. (2012) Epigenetically mediated downregulation of the differentiation-promoting chaperon protein CRABP2 in astrocytic gliomas. Int J Cancer 131: 1963–1968 10.1002/ijc.27446 [DOI] [PubMed] [Google Scholar]
- 57. Kim M-S, Kim YK, Eun HC, Cho KH, Chung JH (2006) All-Trans Retinoic Acid Antagonizes UV-Induced VEGF Production and Angiogenesis via the Inhibition of ERK Activation in Human Skin Keratinocytes. J Investig Dermatol 126: 2697–2706 10.1038/sj.jid.5700463 [DOI] [PubMed] [Google Scholar]
- 58. Reyes Babiano J de LC (2010) Ribosomal protein L35 is required for 27SB pre-rRNA processing in Saccharomyces cerevisiae. Nucleic Acids Res 38: 5177–5192 10.1093/nar/gkq260 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. LaFramboise WA, Jayaraman RC, Bombach KL, Ankrapp DP, Krill-Burger JM, et al. (2009) Acute molecular response of mouse hindlimb muscles to chronic stimulation. Am J Physiol Cell Physiol 297: C556–C570 10.1152/ajpcell.00046.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Maves L, Tyler A, Moens CB, Tapscott SJ (2009) Pbx acts with Hand2 in early myocardial differentiation. Developmental biology 333: 409–418 10.1016/j.ydbio.2009.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Charboneau A, East L, Mulholland N, Rohde M, Boudreau N (2005) Pbx1 is required for Hox D3-mediated angiogenesis. Angiogenesis 8: 289–296 10.1007/s10456-005-9016-7 [DOI] [PubMed] [Google Scholar]
- 62. Sarah De Val BLB (2009) Transcriptional control of endothelial cell development. Developmental cell 16: 180–195 10.1016/j.devcel.2009.01.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Matsui T, Kanai-Azuma M, Hara K, Matoba S, Hiramatsu R, et al. (2006) Redundant roles of Sox17 and Sox18 in postnatal angiogenesis in mice. J Cell Sci 119: 3513–3526 10.1242/jcs.03081 [DOI] [PubMed] [Google Scholar]
- 64. Huber A, French SL, Tekotte H, Yerlikaya S, Stahl M, et al. (2011) Sch9 regulates ribosome biogenesis via Stb3, Dot6 and Tod6 and the histone deacetylase complex RPD3L. EMBO J 30: 3052–3064 10.1038/emboj.2011.221 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Putative miRNA complementary to the 3′ UTR of the upregulated genes in high altitude acclimatized fetal carotid arteries compared to normal control.
(DOC)
Putative miRNA complementary to the 3′ UTR of the downregulated genes in high altitude acclimatized fetal carotid arteries compared to normal control.
(DOC)