

Abstract
Shigellosis, caused by Shigella species, is a major public health problem in Bangladesh. To determine the prevalence and distribution of different Shigella species, we analyzed 10,827 Shigella isolates from patients between 2001 and 2011. S. flexneri was the predominant species isolated throughout the period. However, the prevalence of S. flexneri decreased from 65.7% in 2001 to 47% in 2011, whereas the prevalence of S. sonnei increased from 7.2% in 2001 to 25% in 2011. S. boydii and S. dysenteriae accounted for 17.3% and 7.7% of the isolates respectively throughout the period. Of 200 randomly selected S. sonnei isolates for extensive characterization, biotype g strains were predominant (95%) followed by biotype a (5%). Resistance to commonly used antibiotics including trimethoprim-sulfamethoxazole, nalidixic acid, ciprofloxacin, mecillinam and ampicillin was 89.5%, 86.5%, 17%, 10.5%, and 9.5%, respectively. All isolates were susceptible to ceftriaxone, cefotaxime, ceftazidime and imipenem. Ninety-eight percent of the strains had integrons belonging to class 1, 2 or both. The class 1 integron contained only dfrA5 gene, whereas among class 2 integron, 16% contained dhfrAI-sat1-aadA1-orfX gene cassettes and 84% harbored dhfrA1-sat2 gene cassettes. Plasmids of ∼5, ∼1.8 and ∼1.4 MDa in size were found in 92% of the strains, whereas only 33% of the strains carried the 120 MDa plasmid. PFGE analysis showed that strains having different integron patterns belonged to different clusters. These results show a changing trend in the prevalence of Shigella species with the emergence of multidrug resistant S. sonnei. Although S. flexneri continues to be the predominant species albeit with reduced prevalence, S. sonnei has emerged as the second most prevalent species replacing the earlier dominance by S. boydii and S. dysenteriae in Bangladesh.
Introduction
Bacillary dysentery such as shigellosis is endemic throughout the world, and is one of the major causes of morbidity and mortality, especially among children <5 years of age in many developing countries including Bangladesh [1], [2]. Shigellosis is caused by any one of the four species of Shigella: S. dysenteriae, S. flexneri, S. boydii, and S. sonnei and outbreaks caused by Shigella infection are difficult to control due to their low infectious dose [3], [4]. Globally, every year there are about 165 million cases of Shigella infection and 1.1 million Shigella-related deaths. The majority of these cases occur in developing countries [5]. S. sonnei is the predominant Shigella spp. in developed as well as industrialized countries and is often the second most prevalent Shigella spp. in low income countries [5], [6]. In Bangladesh, shigellosis is endemic. Previously, it accounted for 11% of deaths [7], although a recent report suggests that the mortality rate due to shigellosis has decreased to ∼0.01% overall and ∼0.89% among youngest age group [2]. Previously, S. flexneri was the predominant serogroup (55%) followed by S. dysenteriae (19%), S. boydii (13%) and S. sonnei (7%) [8].
In developing countries, increased resistance to commonly used antibiotics including ampicillin, streptomycin, sulfamethoxazole-trimethoprim, nalidixic acid and tetracycline has been a major concern in the treatment of enteric infections due to various bacterial pathogens [9]. Shigella is transmitted efficiently in low-dose via fecal-oral route in areas of poor hygienic conditions with limited access to clean and potable water [10]. Infections can result from as few as 10 Shigella bacteria [3]. Emergence of multidrug-resistant (MDR) strains is increasing rapidly due to their ability to acquire and disseminate exogenous genes associated with mobile genetic elements such as R-plasmids, transposons, integrons, and genomic islands on the bacterial chromosome [11], [12]. Currently, based on the characteristics of integrase genes, five classes of integrons (classes 1, 2, 3, 4, and 5) have been identified [13]. Several reports indicate that integrons have a role in the dissemination of resistance among gram-negative pathogens and are thus a useful marker of antibiotic resistance [14]. Of the five classes, only class 1 and 2 integrons have been found in Shigella spp. [12], [15], [16]. With the increased movement of people across national and international borders, there is an increased risk of spread of MDR Shigella strains. Following on a recent trend of increased prevalence of S. sonnei in Bangladesh (K. A. Talukder et al., unpublished data), this study was designed to determine the distribution of Shigella species and characterize randomly selected S. sonnei isolated between 2001 and 2011 in Bangladesh.
Materials and Methods
Bacterial Strains
A total of 10,827 Shigella strains were isolated and identified according to standard microbiological and biochemical methods [17] from patients of all ages between 2001 and 2011. The patients either attended the Dhaka treatment centre of the International Centre for Diarrhoeal Disease Research, Bangladesh (icddr,b) or were referred from public and/or private treatment facilities within Dhaka City and/or outside districts. All isolates were confirmed as Shigella spp., by serotyping with commercial Shigella antisera (Denka Seiken, Tokyo, Japan) [18]. Of these, 200 S. sonnei isolates comprising at least one isolate per month and 20 isolates from each year (except 2008, since no strains were available in our lab stock) were randomly selected for further characterization. The strains were grown in trypticase soy broth containing 0.3% yeast extract and stored at −70°C after addition of 15% glycerol.
Biotyping
Biotyping was done using standard methods for fermentation of rhamnose and xylose and hydrolysis of ortho-nitrophenyl-β-D-galactopyranoside (ONPG), and biotypes were designated according to methods described elsewhere [19].
Antimicrobial Susceptibility Test
Antimicrobial susceptibility test was performed by the Kirby-Bauer disc diffusion method on Muller-Hinton agar plates according to the guidelines of the Clinical and Laboratory Standards Institute [20], with commercial antimicrobial discs (Oxoid, Basingstoke, United Kingdom). The antimicrobial discs used in the study were ampicillin (Amp; 10 µg), streptomycin (Str; 10 µg), tetracycline (Te; 30 µg), ciprofloxacin (Cip; 5 µg), nalidixic acid (Na; 30 µg), mecillinam (Mel; 25 µg), sulfamethoxazole-trimethoprim (Sxt; 25 µg), ceftriaxone (Cro, 30 µg), cefotaxime (Ctx; 30 µg), ceftazidime (Caz; 30 µg) and imipenem (Ipm; 10 ug). E. coli ATCC 25922 and Staphylococcus aureus ATCC 25923 were used as control strains for the susceptibility tests.
PCR Assay
Detection of set1 gene (ShET-1), sen gene (ShET-2), ial gene, ipaH gene, stx genes, integrase genes (int1, int2 and int3), class 1 and class 2 cassette regions were performed by polymerase chain reaction (PCR) according to the procedures described previously [1], [21], [22]. The PCR product of representative isolates were sequenced following the procedure described previously [15]. Nucleotide sequences were analysed using BLAST on the website of the National Centre for Biotechnology Information (NCBI). An Escherichia coli (ATCC 25922) strain lacking the140 MDa invasive plasmid and showing susceptibility to all antibiotics was used as a negative control in the PCR assays.
Nucleotide Sequence Accession Numbers
The nucleotide sequences of the gene cassettes in class 1 integron and class 2 integron reported in this paper were submitted to the GenBank using the National Centre for Biotechnology Information (NCBI, Bethesda, MD, USA) under the accession numbers KC964096, KC993867 and KF134652, respectively.
Plasmid Profile Analysis
Plasmid DNA was prepared according to the alkaline lysis method of Kado and Liu (1981) with some modifications [23], [24]. The molecular mass of the plasmid DNA bands was assessed by comparing with the mobility of known molecular mass plasmids in E. coli PDK-9, R1, RP4, Sa and V517 in agarose gels [23].
Pulsed-Field Gel Electrophoresis
PFGE was performed following the PulseNet protocol using the Salmonella Braenderup H9812 strain as molecular weight standard for normalization as recommended previously [25]. Chromosomal DNA was digested with XbaI and fragments were separated using a CHEF DR II apparatus (Bio-Rad Laboratories) under the following conditions: switching time from 2.2 to 54.2 s at 6 V cm−1 for 20 h at 14°C. The resulting profiles were analysed as described earlier [26].
Results
Epidemiologic Studies
Of 10,827 strains of Shigella species isolated from patients of all ages between 2001 and 2011, S. flexneri was the dominant species throughout the study period, but the decreasing tendency of predominance was observed starting from 65.7% (824/1252) in 2001 to 47% (306/650) in 2011. At the same time, the prevalence of S. dysenteriae came down from 10.7% (135/1252) in 2001 to 9% (60/650) in 2011, whereas S. sonnei increased from 7.2% (90/1252) in 2001 to 25% (162/650) in 2011. The frequency of S. boydii increased from 14.6% (185/1252) in 2001 to 16.1% (104/650) in 2011 among all age groups of patients in urban areas of Bangladesh (Figure 1). There were a number of strains isolated each year which had characteristics typical of Shigella but could not be identified to the species level and were therefore designated as Shigella-like organisms (SLOs) which accounted for about 3% of isolates. None of the SLOs agglutinated with any antisera of the established Shigella serovars.
Figure 1. Prevalence of Shigella spp. in Bangladesh from 2001 to 2011.
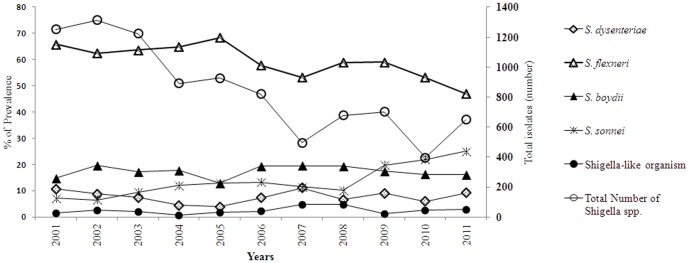
The distribution of each Shigella species is expressed in percentage and the total isolates of Shigella species are expressed in number.
Biotyping
Of 200 strains of S. sonnei, 95% (n = 190) were classified as biotype g (ONPG +, rhamnose−, xylose −) and 5% (n = 10) were belonged to biotype a (ONPG +, rhamnose +, xylose −).
Antibiotic Susceptibility Test
MDR (resistance to ≥ three classes of antimicrobial agents) was detected in 94% (188) of the strains. All the strains were resistant to streptomycin and were susceptible to imipenem, ceftriaxone, cefotaxime, and ceftazidime. Resistance to commonly used antibiotics such as trimethoprim-sulfamethoxazole, nalidixic acid, tetracycline, ciprofloxacin, mecillinam and ampicillin was 89.5%, 86.5%, 84.5%, 17%, 10.5%, and 9.5% respectively. The most prevalent antimicrobial resistance patterns were StrR (100%), StrRSxtR (89.5%), StrRNaR (89.5%), StrRTeR (84.5%), StrRSxtRTeR (83.5%), StrRSxtRNaR (79%), StrRSxtRNaRTeR (74%) and StrRSxtRNaRCipR TeR (12%) (Table 1).
Table 1. Antibiotic resistance patterns of S. sonnei isolated in Bangladesh.
| Antimicrobial Resistance Pattern | No (%) |
| Str | 200 (100%) |
| Str/Sxt | 179 (89.5%) |
| Na/Str | 173 (86.5%) |
| Str/Te | 169 (84.5%) |
| Str/Sxt/Te | 167 (83.5%) |
| Na/Str/Sxt | 158 (79.0%) |
| Na/Str/Sxt/Te | 148 (74.0%) |
| Cip/Str | 34 (17.0%) |
| Cip/Na/Str/Sxt/Te | 24 (12.0%) |
| Mel/Str | 21 (10.5%) |
| Amp/Str | 19 (9.5%) |
| Amp/Str/Sxt/Te | 13 (6.5%) |
| Amp/Na/Str/Sxt/Te | 12 (6.0%) |
Distribution of ipaH, sen, shet and Integrons Genes
The ipaH gene was detected in all tested strains but none of the strains contained stx1 and set1 genes, while only 30% (60/200) strains carried both the ial and sen gene. Of all the strains, 97.5% (195) harbored integrons belonging to either class 1 or 2, or both. Of these strains, two strains carried only class 1 integron (int1), whereas only class 2 integron (int2) was identified in 95% (190) of the strains. None of the strains harbored class 3 integron. Both int1 and int2 integrons were present in three strains (1.5%). Only five strains (2.5%) were lacking both classes of integrons.
The Resistance Gene Cassettes of Integrons
Using the hep58-hep59 primer pairs, specific for the gene cassettes of int1, one type of gene cassette array (562 bp size) was found in five strains and confirmed as dihydrofolate reductase gene type 5 (dfrA5) by DNA sequencing (GenBank accession number KC964096). Two different types of gene cassette arrays were detected using the primer pair hep74-hep51, specific for the gene cassettes of int2. The type-1 1325 bp DNA product of PCR was present in 84% (162/195) of the strains as sequence dhfrA1-sat2 and 16% (31/195) had 2159 bp band as sequence dhfrAI-sat1-aadA1-orfX with the GenBank accession number KC993867 and KF134652, respectively.
Plasmid Profile Analysis
Heterogeneous plasmid patterns ranging in size from approximately 120 to 1.0 MDa was found by analyzing plasmid DNA. A number of small plasmids of ∼5, ∼1.8 and ∼1.4 MDa in size were also found to be present universally in more than 92% (184) of the strains and were considered to be the core plasmids of S. sonnei (Fig.2). Interestingly, 67% (134) of the strains were found to lack the typical 120 MDa invasive plasmid. A middle-ranged plasmid of approximately 35–90 MDa in size was found in 29% (58) of the strains (Fig. 2).
Figure 2. Agarose gel electrophoresis of plasmid DNA showing representative patterns of S. sonnei.

Lane: A, E. coli PDK-9, Lanes: B-M, Representative S. sonnei strains, Lane: N, E. coli V-517, Lane: O, E. coli R-1 and Sa. CHR-indicates the banding position of the chromosome.
PFGE
PFGE analysis of XbaI-digested chromosomal DNA of S. sonnei strains yielded 15–24 reproducible DNA fragments, ranging in size from approximately ∼20 to 485 kb. These strains yielded four major clusters designated: pulsotype A (n = 88), B (n = 17), C (n = 2) and D (n = 3). The similarity indexes for each cluster were 97.5%, 98.9%, 81.7% and 88.3% respectively. The major pulsotype A was further subdivided into three subtypes (A1–A3) and pulsotype B into two subtypes (B1–B2) (Fig. 3).
Phylogenetic dendrogram showing the genetic distances among S. sonnei using XbaI macrorestriction analysis.

Correlation among Biotype, Integron Cascades and Pulsotypes
All strains belonging to pulsotype A harbored ∼1325 bp int2 and pulsotype B carried ∼2159 bp int2. Strains lacking integrons were grouped into pulsotype C. Pulsotype D contained two strains which harbored both integrons and one strain which harbored only int1. All strains of pulsotypes A and B were biotype g, whereas the rest were biotype a (Table 2).
Table 2. Correlation among Biotype, Pulsotypes, Integron Type and Antibiotic Resistance Pattern of S. sonnei.
| Properties | Class 1 Integron (n = 5) | Class 2 Integron (n = 193) | |
| Gene cascade size | 562 bp (100%, n = 5) | 2159 bp (16%, n = 31) | 1325 bp (84%, n = 162) |
| Gene sequence | dfrA5 gene | dhfrAI-sat1-aadA1-orfX | dhfrA1-sat2 |
| Biotype | a | g | g |
| Pulsotype | C | B | A |
| Resistance pattern | Amp, Str, Sxt | Str, Sxt, Te | Mixed patterns |
Discussion
This study was done for a baseline assessment of the epidemiological features of Shigella species, with special emphasis on characterization of the recent endemic S. sonnei in Bangladesh. A changing trend in the distribution of Shigella species was found in this study. The rising trend of shigellosis due to S. sonnei (7.2% to 25%) and S. boydii (14.8% in 23to 16.1%), and decreasing trend due to S. flexneri (65.8% to 47%) and S. dysenteriae (10.8% to 9.2%) was observed in urban areas of Bangladesh from 2001 to 2011. In the past, from 1999 to 2003 about 8% of shigellosis cases per year was caused by S. sonnei [21]. Since then the increasing trend was seen from ∼12% in 2004 to ∼25% in 2011 in urban area of Bangladesh. The increasing trend of S. sonnei from 35% (50/144) in 2010 to 41% (63/154) in 2012 was also observed in another study conducted in rural areas of Bangladesh (Talukder et al., unpublished data). Similar trend was noticed in many developed and industrialized countries [27] including India [28] and Pakistan [29]. The reasons for this shifting trend have been suggested to be an improvement of overall nutritional status [8], [30], socioeconomic status [31], sanitation condition [32], and a reduction of the chances of cross immunity imparted by Plesiomonas shigelloides [33].
In the present study, we found that biotype g (rhamnose non-farmenter) was the predominant (95%) biotype followed by biotype a (rhamnose farmenter). It is interesting to note that all strains were found to be rhamnose farmenter in an earlier study [21]. Recent emergence of biotype g strains has also been reported from the developing and industrialized countries including Ireland [4], Italy [34], Australia [35], Malaysia [36] and South Korea [37]. Therefore, biotype g appears to have emerged and spread globally in recent decades contributing to the prevalence of this biotype throughout the world [34], [38]–[40].
Antimicrobial resistance has emerged as a serious global public health concern. In low and middle income countries including Bangladesh, the declining susceptibility to commonly used antibiotics and emergence of MDR bacteria have been linked to the indiscriminate or inappropriate use of antibiotics. The other contributing factors include bacterial evolution, climate changes, cheap and ready availability of antibiotics, lack of medical practitioner with proper training, poor-quality of available drugs and unhygienic sanitary conditions [32], [41], [42]. The majority of the strains in this study were resistant to commonly used antibiotics including Na, Str, Sxt and Te. We detected that resistance of S. sonnei to Cip was about 10% in 2007 and increased dramatically by sevenfold (70%) in 2011. In Bangladesh, resistance of S. dysenteriae 1 to Cip was first detected as early as in 2003 but in case of S. flexneri and S. boydii it was detected in 2007 and 2008, respectively [8], [43]. In the present study, all the tested strains were susceptible to 3rd generation cephalosporins (Cro, Ctx and Caz) and Ipm probably due to their very infrequent use as alternative therapeutic regimens in this geographic area. Notably, Ipm resistance was found in other enteric pathogens except Shigella spp. [44].
Plasmid profile analysis is a well documented and important tool for epidemiological studies of enteric pathogens [3]. It has been reported earlier that S. sonnei strains possessed multiple numbers of plasmids with a heterogeneous combination but three plasmids of ∼5, ∼1.8 and ∼1.4 MDa (core plasmids) were commonly present (90%) in most S. sonnei [21]. Of 200 strains analyzed in this study, 92% contained these core plasmids supporting our previous report [21] and 33% of the strains contained both the sen and ial gene and the typical 120-MDa invasive plasmid. The possible explanations for the absence of the 120 MDa plasmid might be a loss of the plasmid due to (i) long-term storage at −70°C and (ii) repeated subculturing of the strains [45]. Self-transmissible middle-range plasmids (∼30–90 MDa) in Shigella spp. are associated with antibiotic resistance [46]. In this study only 17% of the MDR strains had middle-range plasmids. Most Shigella strains harbored one or two classes of integrons. The presence of integrons in Shigella species varied in numbers in diverse geographical regions [1], [4], [26], [47], [48]. The apparent correlation between multidrug resistance and presence of integron in the strains was not due to the presence of integrons on conjugative plasmids. Integrons found in most strains were chromosomal, and thus, it can be concluded that antibiotic resistance was not only due to the presence of middle-range plasmids but also for the presence of chromosome mediated genes [1].
The low prevalence (2.5%) of int1 found in this study was also detected in other countries [16], [48]. In this study we found that only dfrA5 gene in int1 cassette conferred trimethoprim resistance, an observation rarely reported in S. sonnei [49]. Several investigators reported the presence of this gene in S. flexneri and other members of Enterobacteriaceae [48], [50]. The presence of int2 was significantly higher in MDR strains (p <0.001). Although other mechanisms are possible, resistance to streptomycin, spectinomycin and trimethoprim seems to be attributable to expression of genes contained in int2 of the members of Tn7 family. Generally, int2 carries the open reading frames dhfrI, sat1 and aadA. However, due to the presence of a defective integrase, some different structures have been very infrequently found in endemic S. sonnei strains [4], [48]. This study showed a higher prevalence (16%) of int2 of ∼2159 bp in size harboring dhfrI, sat1 and aadA gene compared to previous studies. Strains harboring ∼2159 bp integron showed uniform resistance to Str, Sxt and Te, whereas no specific association with antibiotic resistance pattern was found in strains containing ∼1325 bp class 2 integron. The same array of gene cassettes of int2 was found in other members of Enterobacteriaceae [38], [51]. The variation of int2 cassette content among the MDR S. sonnei strains from different geographical locations might be due to the independent acquisitions of the integron over time and further diversification by clonal expansion and subsequent global spreading [40].
To date, a number of different genotyping methods including ribotyping, inter-IS1 spacer typing (IST) method, enterobacterial repetitive intergenic consensus sequence based PCR, multilocus variable-number tandem-repeat analysis (MLVA) and PFGE have been used for the epidemiological investigation and phylogenetic study of S. sonnei [26], [52], [53]. Although MLVA has good discriminatory power for distinguishing epidemiologic relationship, it is not a universal method and not 100% reproducible, and has no properly validated protocol for use in surveillance networks [54]. On the other hand, PFGE is the gold standard, has well-established global surveillance network and has been used successfully for the investigations of outbreaks as well as epidemiological studies of Shigella species [21]. PFGE analysis of these strains showed that S. sonnei biotype a strains were genetically more diverse than biotype g strains, and revealed that strains having different integron patterns belonged to different clusters. This finding is congruent with a previous study [38].
Overall, our findings suggest a decrease in prevalence of S. flexneri and at the same time a drastic increase in S. sonnei to become the second predominant species by replacing S. boydii and S. dysenteriae in Bangladesh. S. sonnei biotype g carrying int2 was prevalent. The high prevalence of integrons in Shigella may have clinical significance, as multiple gene cassettes could be integrated into integrons leading to multidrug resistance, even to broad-spectrum antibiotics such as cephalosporins and quinolones among Shigella species. Development of a suitable vaccine against S. sonnei should be a priority in order to reduce the disease burden due to Shigella infection in Bangladesh, as all S. sonnei share a common O antigen that has proven to be a potential vaccine candidate.
Funding Statement
This study was funded by icddr,b and its donors which provide unrestricted support to icddr,b for its operations and research. Current donors providing unrestricted support include: Government of the People's Republic of Bangladesh, Australian Agency for International Development (AusAID), Canadian International Development Agency (CIDA), Swedish International Development Cooperation Agency (Sida), and the Department for International Development, UK (DFID). The authors gratefully acknowledge these donors for their support and commitment to icddr,b′s research efforts. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Wen X, Wu Y, Bian F, Sun Y, Zheng X, et al. (2012) High prevalence of atypical class 1 integrons and class 2 integrons in multi-drug resistance Shigella flexneri isolated from China. Afr J Microbiol Res 6: 6987–6993. [Google Scholar]
- 2. Bardhan P, Faruque ASG, Naheed A, Sack DA (2010) Decreasing shigellosis-related deaths without Shigella spp.–specific interventions, Asia. Emerg Infect Dis 16: 1718–1723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Talukder KA, Azmi IJ (2012) Population genetics and molecular epidemiology of Shigella species. In: Faruque SM, editor. Foodborne and waterborne bacterial pathogens epidemiology, evolution and molecular biology: Caister Academic Press pp. 63–76.
- 4. DeLappe N, O'Halloran F, Fanning S, Corbett-Feeney G, Cheasty T, et al. (2003) Antimicrobial resistance and genetic diversity of Shigella sonnei isolates from western Ireland, an area of low incidence of infection. J Clin Microbiol 41: 1919–1924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Kotloff KL, Winickoff JP, Ivanoff B, Clemens JD, Swerdlow DL, et al. (1999) Global burden of Shigella infections: implications for vaccine development and implementation of control strategies. Bull World Health Organ 77: 651–666. [PMC free article] [PubMed] [Google Scholar]
- 6. Niyogi SK (2005) Shigellosis. J Microbiol 43: 133–143. [PubMed] [Google Scholar]
- 7. Bennish ML, Salam MA, Haider R, Barza M (1990) Therapy for shigellosis. II. Randomized, double-blind comparison of ciprofloxacin and ampicillin. J Infect Dis 162: 711–716. [DOI] [PubMed] [Google Scholar]
- 8. Khatun F, Faruque A, Koeck J, Olliaro P, Millet P, et al. (2011) Changing species distribution and antimicrobial susceptibility pattern of Shigella over a 29-year period (1980–2008). Epidemiol Infect 139: 446–452. [DOI] [PubMed] [Google Scholar]
- 9. Ke X, Gu B, Pan S, Tong M (2011) Epidemiology and molecular mechanism of integron-mediated antibiotic resistance in Shigella . Arch Microbiol 193: 767–774. [DOI] [PubMed] [Google Scholar]
- 10. Weissman JB, Gangorosa EJ, Schmerler A, Marier RL, Lewis JN (1975) Shigellosis in day-care centres. Lancet 1: 88–90. [DOI] [PubMed] [Google Scholar]
- 11. Rowe-Magnus DA, Mazel D (2002) The role of integrons in antibiotic resistance gene capture. Int J Med Microbiol 292: 115–125. [DOI] [PubMed] [Google Scholar]
- 12. Goh K, Chua D, Beck B, McKee ML, Bhagwat AA (2010) Arginine-dependent acid-resistance pathway in Shigella boydii . Arch Microbiol 193: 179–185. [DOI] [PubMed] [Google Scholar]
- 13. Cambray G, Guerout AM, Mazel D (2010) Integrons. Annu Rev Genet 44: 141–166. [DOI] [PubMed] [Google Scholar]
- 14. Fluit AC, Schmitz FJ (2004) Resistance integrons and super integrons. Clin Microbiol Infect 10: 272–288. [DOI] [PubMed] [Google Scholar]
- 15. White PA, McIver CJ, Rawlinson WD (2001) Integrons and gene cassettes in the enterobacteriaceae . Antimicrob Agents Chemother 45: 2658–2661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Madiyarov RS, Bektemirov AM, Ibadova GA, Abdukhalilova GK, Khodiev AV, et al. (2010) Antimicrobial resistance patterns and prevalence of class 1 and 2 integrons in Shigella flexneri and Shigella sonnei isolated in Uzbekistan. Gut Pathog 2: 18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.WHO (1987) Programme for control of diarrhoeal disease (CDD/93.3 Rev. 1). In Manual for laboratory investigation of acute enteric infections. Geneva, Switzerland.: World Health Organization. pp. 9–20.
- 18. Talukder KA, Khajanchi BK, Islam MA, Dutta DK, Islam Z, et al. (2006) The emerging strains of Shigella dysenteriae type 2 in Bangladesh are clonal. Epidemiol Infect 134: 1249–1256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Nastasi A, Pignato S, Mammina C, Giammanco G (1993) rRNA gene restriction patterns and biotypes of Shigella sonnei . Epidemiol Infect 110: 23–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.CLSI (2010) Performance standards for antimicrobial susceptibility testing: 20th informational supplement (June 2010 update), M100-S20-U, Wayne, PA. Clinical and Laboratory Standards Institute.
- 21. Talukder KA, Islam Z, Dutta DK, Islam MA, Khajanchi BK, et al. (2006) Antibiotic resistance and genetic diversity of Shigella sonnei isolated from patients with diarrhoea between 1999 and 2003 in Bangladesh. J Med Microbiol 55: 1257–1263. [DOI] [PubMed] [Google Scholar]
- 22. Vrints M, Mairiaux E, Van ME, Collard JM, Bertrand S (2009) Surveillance of antibiotic susceptibility patterns among Shigella sonnei strains isolated in Belgium during the 18-year period 1990 to 2007. J Clin Microbiol 47: 1379–1385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Talukder KA, Islam MA, Dutta DK, Hassan F, Safa A, et al. (2002) Phenotypic and genotypic characterization of serologically atypical strains of Shigella flexneri type 4 isolated in Dhaka, Bangladesh. J Clin Microbiol 40: 2490–2497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Kado C, amp, Liu S (1981) Rapid procedure for detection and isolation of large and small plasmids. J Bacteriol 145: 1365–1373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Ribot EM, Fair MA, Gautom R, Cameron DN, Hunter SB, et al. (2006) Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157: H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog Dis 3: 59–67. [DOI] [PubMed] [Google Scholar]
- 26. Koh XP, Chiou CS, Ajam N, Watanabe H, Ahmad N, et al. (2012) Characterization of Shigella sonnei in Malaysia, an increasingly prevalent etiologic agent of local shigellosis cases. BMC Infect Dis 12: 122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Shiferaw B, Solghan S, Palmer A, Joyce K, Barzilay EJ, et al. (2012) Antimicrobial susceptibility patterns of Shigella isolates in foodborne diseases active surveillance network (foodnet) sites, 2000–2010. Clin Infect Dis 54: S458–S463. [DOI] [PubMed] [Google Scholar]
- 28. Nandy S, Mitra U, Rajendran K, Dutta P, Dutta S (2010) Subtype prevalence, plasmid profiles and growing fluoroquinolone resistance in Shigella from Kolkata, India (2001–2007): a hospital-based study. Trop Med Int Health 15: 1499–1507. [DOI] [PubMed] [Google Scholar]
- 29. Zafar A, Hasan R, Nizami SQ, von Seidlein L, Soofi S, et al. (2009) Frequency of isolation of various subtypes and antimicrobial resistance of Shigella from urban slums of Karachi, Pakistan. Int J Infect Dis 13: 668–672. [DOI] [PubMed] [Google Scholar]
- 30. Faruque ASG, Ahmed AS, Ahmed T, Islam MM, Hossain MI, et al. (2008) Nutrition: basis for healthy children and mothers in Bangladesh. J Health Popul Nutr 26: 325–339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Hogan MC, Foreman KJ, Naghavi M, Ahn SY, Wang M, et al. (2010) Maternal mortality for 181 countries, 1980–2008: a systematic analysis of progress towards Millennium Development Goal 5. Lancet 375: 1609–1623. [DOI] [PubMed] [Google Scholar]
- 32. Fewtrell L, Kaufmann RB, Kay D, Enanoria W, Haller L, et al. (2005) Water, sanitation, and hygiene interventions to reduce diarrhoea in less developed countries: a systematic review and meta-analysis. Lancet Infect Dis 5: 42–52. [DOI] [PubMed] [Google Scholar]
- 33. Sayeed S, Sack D, Qadri F (1992) Protection from Shigella sonnei infection by immunisation of rabbits with Plesiomonas shigelloides (SVC O1). J Med Microbiol 37: 382–384. [DOI] [PubMed] [Google Scholar]
- 34. Mammina C, Aleo A, Romani C, Nastasi A (2006) Shigella sonnei biotype g carrying class 2 integrons in southern Italy: a retrospective typing study by pulsed field gel electrophoresis. BMC Infect Dis 6: 117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Mclver CJ, White PA, Jones LA, Karagiannis T, Harkness J, et al. (2002) Epidemic strains of Shigella sonnei biotype g carrying integrons. J Clin Microbiol 40: 1538–1540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Hoe CH, Yasin RM, Koh YT, Thong KL (2005) Antimicrobial susceptibility and pulsed-field gel electrophoresis of Shigella sonnei strains in Malaysia (1997-2000). J Appl Microbiol 99: 133–140. [DOI] [PubMed] [Google Scholar]
- 37. Oh JY, Yu HS, Kim SK, Seol SY, Cho DT, et al. (2003) Changes in patterns of antimicrobial susceptibility and integron carriage among Shigella sonnei isolates from southwestern Korea during epidemic periods. J Clin Microbiol 41: 421–423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Ranjbar R, Aleo A, Giammanco GM, Dionisi AM, Sadeghifard N, et al. (2007) Genetic relatedness among isolates of Shigella sonnei carrying class 2 integrons in Tehran, Iran, 2002–2003. BMC Infect Dis 7: 62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Izumiya H, Tada Y, Ito K, Morita-Ishihara T, Ohnishi M, et al. (2009) Characterization of Shigella sonnei isolates from travel-associated cases in Japan. J Med Microbiol 58: 1486–1491. [DOI] [PubMed] [Google Scholar]
- 40. Holt KE, Baker S, Weill F-X, Holmes EC, Kitchen A, et al. (2012) Shigella sonnei genome sequencing and phylogenetic analysis indicate recent global dissemination from Europe. Nat Genet 44: 1056–1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Putnam SD, Riddle MS, Wierzba TF, Pittner BT, Elyazeed RA, et al. (2004) Antimicrobial susceptibility trends among Escherichia coli and Shigella spp. isolated from rural Egyptian paediatric populations with diarrhoea between 1995 and 2000. Clin Microbiol Infect 10: 804–810. [DOI] [PubMed] [Google Scholar]
- 42. Church DL (2004) Major factors affecting the emergence and re-emergence of infectious diseases. Clin Lab Med 24: 559–586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Talukder KA, Khajanchi BK, Islam MA, Islam Z, Dutta DK, et al. (2006) Fluoroquinolone resistance linked to both gyrA and parC mutations in the quinolone resistance-determining region of Shigella dysenteriae type 1. Curr Microbiol 52: 108–111. [DOI] [PubMed] [Google Scholar]
- 44. Islam M, Talukdar P, Hoque A, Huq M, Nabi A, et al. (2012) Emergence of multidrug-resistant NDM-1-producing Gram-negative bacteria in Bangladesh. Eur J Clin Microbiol Infect Dis 31: 2593–2600. [DOI] [PubMed] [Google Scholar]
- 45. Vargas M, Gascon J, De Anta MTJ, Vila J (1999) Prevalence of Shigella enterotoxins 1 and 2 among Shigella strains isolated from patients with traveler's diarrhea. J Clin Microbiol 37: 3608–3611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Haider K, Huq MI, Talukder KA, Ahmad QS (1989) Electropherotyping of plasmid DNA of different serotypes of Shigella flexneri isolated in Bangladesh. Epidemiol Infect 102: 421–428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Jin YH, Oh YH, Jung JH, Kim SJ, Kim JA, et al. (2010) Antimicrobial resistance patterns and characterization of integrons of Shigella sonnei isolates in Seoul, 1999–2008. J Microbiol 48: 236–242. [DOI] [PubMed] [Google Scholar]
- 48. Sow AG, Aïdara-Kane A, Barraud O, Gatet M, Denis F, et al. (2010) High prevalence of trimethoprim-resistance cassettes in class 1 and 2 integrons in Senegalese Shigella spp. isolates. J Infect Dev Ctries 4: 207–212. [DOI] [PubMed] [Google Scholar]
- 49. Seol SY, Kim YT, Jeong YS, Oh JY, Kang HY, et al. (2006) Molecular characterization of antimicrobial resistance in Shigella sonnei isolates in Korea. J Med Microbiol 55: 871–877. [DOI] [PubMed] [Google Scholar]
- 50. Dawes FE, Kuzevski A, Bettelheim KA, Hornitzky MA, Djordjevic SP, et al. (2010) Distribution of class 1 integrons with IS26-mediated deletions in their 3′-conserved segments in Escherichia coli of human and animal origin. PloS One 5: e12754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. van Essen-Zandbergen A, Smith H, Veldman K, Mevius D (2007) Occurrence and characteristics of class 1, 2 and 3 integrons in Escherichia coli, Salmonella and Campylobacter spp. in the Netherlands. J Antimicrob Chemother 59: 746–750. [DOI] [PubMed] [Google Scholar]
- 52. Chiou CS, Watanabe H, Wang YW, Wang WL, Terajima J, et al. (2009) Utility of multilocus variable-number tandem-repeat analysis as a molecular tool for phylogenetic analysis of Shigella sonnei . J Clin Microbiol 47: 1149–1154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Liu P, Lau YJ, Hu BS, Shyr JM, Shi ZY, et al. (1995) Analysis of clonal relationships among isolates of Shigella sonnei by different molecular typing methods. J Clin Microbiol 33: 1779–1783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Sabat A, Budimir A, Nashev D, Sa-Leao R, van Dijl J, et al. (2013) Overview of molecular typing methods for outbreak detection and epidemiological surveillance. Euro Surveill 18.. [DOI] [PubMed] [Google Scholar]