

Abstract
The molecular chaperone Hsp90 is an essential protein in eukaryotic organisms and is highly conserved throughout all kingdoms of life. It serves as a platform for the folding and maturation of many client proteins including protein kinases and steroid hormone receptors. To fulfill this task Hsp90 performs conformational changes driven by the hydrolysis of ATP. Further, it can resort to a broad set of co-chaperones, which fit the Hsp90 machinery to the needs of specific client proteins. During the last years the number of identified co-chaperones has been consistently rising, implying that the client spectrum of Hsp90 may be much more diverse and larger than currently known. Many cofactors contain a TPR-domain for interactions at the C-terminus of Hsp90 and in many cases their functions and client sets remain to be uncovered. Hsp90 is also a putative target to interfere with cancerous and infectious diseases. Thus the knowledge on more of its cellular functions would provide also more therapeutic options for the future. In this review we compile the current knowledge on the Hsp90 ATPase mechanism, cofactor regulation and prospects of Hsp90 inhibition.
Keywords: Hsp90, chaperone, clients, SHR, kinase, ATPase cyle
Introduction
Heat-shock protein 90 (Hsp90) is a highly conserved molecular chaperone and essential for viability in eukaryotic organisms, like Saccaromyces cerevisiae [1], Caenorhabditis elegans [2] and Drosophila melanogaster [3]. Initially it was discovered as a protein whose abundance increases upon heat stress, but already in unstressed cells the protein amounts to about 1% of the soluble cytosolic protein [4]. In vertebrates two distinct isoforms are encoded - the mostly inducible Hsp90α and the constitutively expressed Hsp90β [5,6]. Two additional homologs of Hsp90 exist in mitochondria and in the endoplasmic reticulum - Trap1 and Grp94. All Hsp90 homologs have the same domain architecture with an N-terminal nucleotide binding domain (NTD), a middle domain (MD) and a C-terminal dimerization site (CTD). They also share the mechanism of ATP hydrolysis connected to domain movements (Figure 1) [7,8]. These are initiated by a short segment of the N-domain, the ATP-lid, flapping over the ATP binding pocket [6]. This alteration induces contact formation at the NTDs and further results in a twisted and closed Hsp90 dimer [7,8] (Figure 1). Crystal structures point to rotation movements of the nucleotide-binding domains against the MD, which may precede the contact formation of the NTDs [9-11]. This closed conformation is essential for ATP hydrolysis, after which the N-domains separate from each other and ADP gets released. Hsp90 client proteins seem to interact with the moving Hsp90 scaffold to reach unfavorable conformations stabilized by the chaperone.
Figure 1.
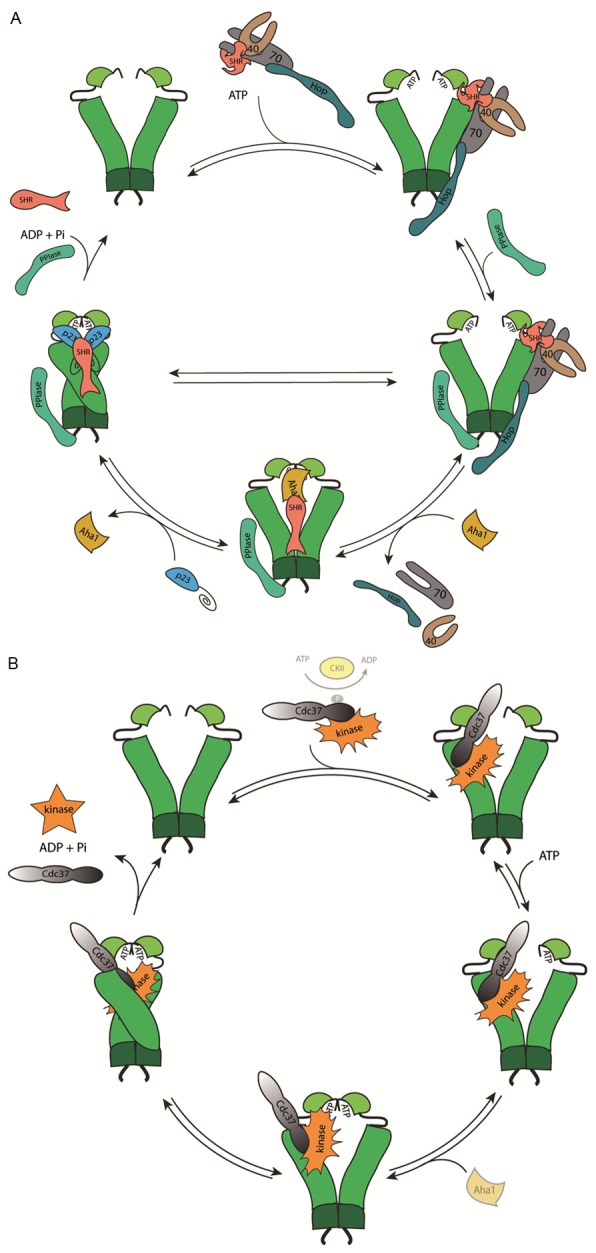
A: Model of the ATPase driven cycle to activate steroid hormone receptors. Different co-chaperones associate during the cycle at different stages of the SHR-activation process. B: A reaction pathway for kinase clients, which are brought into Hsp90 complexes by the kinase-specific co-chaperone Cdc37.
The best characterized of the many client proteins (listed at http://www.picard.ch/) originate from two classes: steroid hormone receptors (SHR) [12] and protein kinases [13]. In fact more than two-thirds of the protein kinases depend on Hsp90, likewise many transcription factors and, as recently reported, also many E3-ligases [14]. Most of these clients are directed to proteasomal degradation if Hsp90 fails its task [15]. It is challenging to elucidate the precise function of Hsp90 in these interactions and still many mechanistic details of client processing are unclear. Do all Hsp90 clients require the same movements? How does Hsp90 recognize and interact with its clients? Does every client need its own set of co-chaperones? In this review we aim to compile the current knowledge on Hsp90 client interactions and describe the approaches to inhibit these processes by using Hsp90 inhibitory compounds.
Processing steroid hormone receptors with the help of cofactors
Complex formation between Hsp90 and its clients has been studied most extensively for SHRs (Figure 1A). After initial reports about Hsp90-SHR complexes in the early 1990s further proteins were identified in these assemblies [16-18] and most of these accessory proteins are helping Hsp90 during assembly of SHR complexes. At least a set of five additional proteins participate to mature SHRs in cell culture systems [19]. In the current model, the SHRs are first recognized by the Hsc70-Hsp40 chaperone complex [20,21]. The adapter protein Hop/STIP1 (Hsp90/Hsp70 organizing protein, previously p60) is then docking this assembly to the open conformation of Hsp90. Hop/STIP1 itself consists of three TPR domains - TPR1, TPR2A and TPR2B. Hsp90 is bound to the TPR2A-domain and Hsc70 to the TPR1 or TPR2B domain enabling the client transfer between the chaperone systems [22,23]. Another TPR-domain containing protein, like the PPIases FKBP4, FKBP5 or PPID*, binds to the second TPR-accessory motif of Hsp90 and forms an asymmetric complex consisting of one PPIase, Hop/STIP1, Hsp90 and the Hsc70-Hsp40 bound client protein [24,25]. ATP-pandered conformational changes in Hsp90 lead to the closing of the N-terminal domains. This reduces the affinity of Hop/STIP1 for the assembly, leading to the exit of the adapter protein and its associated Hsc70-system (Figure 1A). The N-terminal dimerized conformation has high affinity for the cofactor p23, which serves to stabilize this closed complex [26]. In the p23-containing complex the steroid hormone receptor is able to bind steroid, while previous conformations were unable to do so [27]. p23 is not essential to open the hormone binding cleft of SHR but it stabilizes the heterocomplex of Hsp90 and SHR [28]. In particular the closing step can be accelerated by the Hsp90 ATPase activator Aha1 [26]. This active conformation now may exist for a certain time, until the chaperone opens up again for ADP release. Interestingly, for SHR, opening of the steroid binding cleft is the rate-limiting step on the way to a hormone binding state [29]. If steroid hormone is bound during this time, the steroid hormone receptor is released in an active form, is translocated into the nucleus where it can bind to glucocorticoid response elements on the DNA [30,31]. If no hormone binding occurred the steroid hormone receptor is inactive and may re-enter the cycle. Apparently phosphorylation might also be relevant in this cycle, given the complex formation between Hsp90, SHR and the phosphatase PPP5 [32,33].
It is still not clear, how the activation of SHRs by Hsp90 is achieved on a molecular level. Combining the extensive knowledge on the assembly process and on the conformational states of the co-chaperone interaction, it is likely that activation happens during the transfer from the Hop-containing Hsp90-complex to the closed state. It is well known that the p23-bound SHR is active [34] and the exclusive binding of p23 to the closed conformation of Hsp90 supports this activation model. Also, Hsp90 variants which block ATP-hydrolysis at the stage of the closed complex (like E47A-Hsp90) accumulate very stable active receptor complexes, suggesting that the important conformational changes in SHRs happen during the closing reaction of Hsp90 [35,36].
Processing kinase clients
The activation and maturation of kinases differs from the SHR reaction cycle. A kinase-specific cofactor is involved here, the protein Cdc37 (cell division cycle 37). In vitro studies using reticulocyte lysate had initially shown that Raf kinase can exist in complex with Hsp90 and Cdc37 [13] but Cdc37 can also interact with kinases alone. As such it can bind to AKT (protein kinase B) [37], has an influence on Cdc28 and Cak1 [38] and on the catalytic activity of Hck [39]. Taipale and coworkers recently described hundreds of protein kinases that depend on the interaction with Cdc37 and Hsp90 [14].
Also for kinases a reaction cycle needs to be unraveled (Figure 1B). It is known that the N-terminal domain of Cdc37 is necessary to mediate interactions with protein kinases [40-42]. Cdc37 also binds to Hsp90 and inhibits its ATPase activity and conformational flexibility [43]. The Hsp90-Cdc37 interaction seems to involve two sites. C. elegans Cdc37 most strongly binds an M-domain binding site of Hsp90 whereas the human Cdc37 interacts stronger with the NTD [43-47]. It thus is likely that during complex formation with the kinase the binding features are changing and both binding sites are utilized by Cdc37 in the open and closed Hsp90 conformation. In our studies the Cdc37-Hsp90 system can be formed as an open and as a closed complex, suggesting that Cdc37 can participate in both, the open encounter complex and the stable closed complex (Figure 1B). It is unknown, whether the kinase also requires the help of Hsc70-Hsp40 during the transfer to Hsp90. While previous studies imply that Hsp70, Hsp40, Hop/STIP1, Cdc37 and Hsp90 are necessary to chaperone the kinase [48] direct binding studies see strong competition between Hop/STIP1 and Cdc37 for Hsp90-binding, making co-complexes of these proteins unlikely [49]. The kinase specific co-chaperone Cdc37 is putatively regulated by phosphorylation. It can be phosphorylated at Ser13 (Ser14 in yeast) by casein kinase 2α (CK2) [42,50] and gets dephosphorylated by the Hsp90-associated phosphatase PPP5 in the ternary complex of Hsp90, Cdc37 and PPP5 [42,51]. This phosphorylation is assumed to regulate the affinity for the kinase.
Kinases seem to interact with Hsp90 and Cdc37 with structural motifs in the N-terminal lobe of the catalytic domain [52]. It has been shown that a conserved glycine-rich loop (the P-loop), necessary for ATP binding, plays an essential role in this [53,54], but in all cases the catalytic domain of the protein kinase is involved [52]. For kinases, the MD and ND of Hsp90 seem to contain client binding sites [55-57]. Structural evidence from EM reconstitutions of the Hsp90-Cdc37-Cdk4 complex suggests that the kinase is located on the outside of the Hsp90 dimer [55]. Recent studies show that the chaperone system competes with ATP-binding to the kinase, with Cdc37 leaving the complex, as soon as the kinase is stabilized by nucleotide [58].
Also for kinases evidence suggests that in particular the closed conformation is the high affinity client binding state. Two-hybrid screens to detect Hsp90 clients in yeast only resulted in kinase hits, if the ATPase deficient E33A-variant was used as bait [59], implying that like with steroid hormone receptors, here also the enrichment of the closed state during the cycle results in stable complex formation.
Processing of other clients by Hsp90
Despite SHRs and kinases also the interaction site of Hsp90 with the tumor suppressor protein p53 has been studied in detail. This interaction relies mainly on electrostatic charges in the CTD and MD of Hsp90 [60]. The strong hydrophilic character of the binding may suggest that Hsp90 client complexes do not go much beyond the initial encounter complex, which is usually characterized by very hydrophilic interactions [61]. This electrostatic interplay, in contrast to the more hydrophobic interaction with Hsc70, likewise has been observed in very early studies on SHR-Hsp90 complexes [34], complexes between the artificial client protein Δ131Δ of staphylococcus nuclease with Hsp90 [56] and between Hsp90 and citrate synthase [62].
Given the highly diverse client set of Hsp90, it may be rewarding to speculate about the nature of the client-Hsp90 interaction in the context of the potential development of this folding helper. How did these chaperone machines evolve and how did the chaperone client proteins become dependent on the chaperones? One explanation might be that any energy-driven cellular machine, which performs directed movements utilizing freely available energy sources like ATP, potentially attracts the interest of proteins in need of conformational rearrangements. In cases where this interaction is beneficial and sufficient evolutionary pressure is present on the future client, the chaperone machinery and the client protein may co-evolve to improve their interaction, manifesting the dependency of the client protein on Hsp90. Other clients may also utilize the same machinery, maybe even in a different way, and evolve to make maximal use of the folding helper [63,64]. Hsp90 thus would become a scaffold, which utilizes energy to perform a more or less predictable set of movements and adaptive client proteins learned to interact with these in a beneficial way.
Like the number of client proteins, also the number of Hsp90-associated cofactors increased strongly in higher eukaryotes. This is evident from the high number of TPR-containing proteins in mammals [65]. Some of these proteins, originally devoid of a TPR-motif obtained the ability to interact with Hsp90 by adopting a TPR-domain. One such example is UNC-45, which plays an essential role - together with Hsp90 - during myosin fiber formation in muscles [49,66,67]. Like UNC-45, several other TPR-domain containing cofactors emerged in higher eukaryotes (Figure 2). For some of these proteins only limited information on their function is available, but it is to be expected that new TPR-domain containing Hsp90 cofactors enable new cellular functions (Figure 2). These cofactors include for example the protein Xap2/AIP, which was found in complex with the dioxin-receptor [68,69] or the mitochondrial TOM70 import receptor. While the core system is remarkably conserved in all eukaryotes, several other proteins, which may constitute Hsp90 partner proteins, were identified by bioinformatics approaches in higher eukaryotes, including TTC28, TTC12, WDTC1 or RPAP3 [65]. This increasing amount of co-chaperones provides opportunities for new classes of clients. It therefore is not surprising that entirely new unrelated clients emerge in higher eukaryotes like the reverse transcriptase from Hepatits B virus or the RNA-interacting domain of the Argonaut proteins [70,71].
Figure 2.
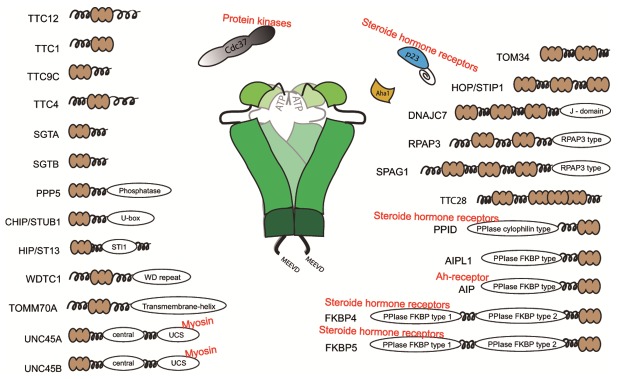
Overview of TPR-domain containing co-chaperones and the non-TPR containing Cdc37, p23 and Aha1. The different TPR-proteins are presumed to interact with Hsp90. The TPR domain is shown in beige.
Hsp90 inhibitors
Some of the established Hsp90 clients play a central role in human diseases like cancer, neurodegenerative disorders and viral infections. Mutated or overexpressed kinases like Akt [72] Bcr-Abl [73] and Raf1 [74] control cancerogenic growth [75] and it was shown that especially Hsp90 is overexpressed in cancer cells [76]. In mamma carcinomas Hsp90 helps cancer cells to obtain resistance to certain forms of chemotherapy [77,78]. Thus it is of interest to use the chaperone as a therapeutic target.
The first identified inhibitors were the natural benzoquinone ansamycin geldanamycin (GM) and the macrolactone radicicol (RD). They show antiproliferative and antitumor effects in vitro and in vivo [74,79,80]. Both inhibitors bind to the N-terminal ATP binding pocket of Hsp90 with a much higher affinity than the nucleotide [79-82]. As a consequence nucleotide binding is blocked, the chaperone cycle arrested and the stable complex with closed N-terminal domains becomes unachievable, leading to degradation of the Hsp90 client [83].
GM and RD have some pharmacological disadvantages like limited solubility and off-target toxicities [84]. Therefore semisynthetic GM analogues were developed: 17-allylamino-17-demethoxygeldanamycin (17-AAG), a carbon-17 substituted derivate, is less toxic and shows similar anti-proliferative effects [85]. Also fully synthetic inhibitors have been developed employing purine scaffolds, fragment-based screening or virtual-screening methods leading to new chemical inhibitors of the Hsp90 ATPase [84,86]. Besides N-terminal inhibitors of Hsp90, novobiocin, an amino coumarin, is a compound targeting the dimerization domain of Hsp90 [87,88]. Several of these Hsp90 inhibitors are in clinical trials [89].
Hsp90 is also relevant for neurodegenerative and infectious diseases. It stabilizes p35, a CDK5 activating protein leading to aberrant tau phosphorylation and also directly interacts with a mutated form of tau protein [90]. It thereby influences the accumulation of toxic tau aggregates in tauopathy like Alzheimer’s disease (AD) and frontotemporal dementia (FTD) [90]. In a mouse model of tauopathy, a purine-scaffold Hsp90 inhibitor leads to a decrease in relevant tau species [91]. In viral infections Hsp90 plays a central role in the folding of essential viral proteins [92]. Here it is even possible to inhibit the replication of poliovirus and paramyxovirus by Hsp90-inhibition [92,93]. Thus the importance to understand the Hsp90 machinery in detail is also underscored by the growing list of diseases associated with this machinery and future developments to modulate its activity.
Acknowledgements
K.R. is grateful to the Deutsche Forschungsgemeinschaft for funding (RI1873/1-3). J.M.E. also acknowledges the support of the TUM Graduate School’s Faculty Graduate Center of Chemistry at the Technische Universität München. K.R. and J.M.E. thank Veronika Haslbeck for critical reading the manuscript.
References
- 1.Borkovich KA, Farrelly FW, Finkelstein DB, Taulien J, Lindquist S. hsp82 is an essential protein that is required in higher concentrations for growth of cells at higher temperatures. Mol Cell Biol. 1989;9:3919–30. doi: 10.1128/mcb.9.9.3919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Birnby DA, Link EM, Vowels JJ, Tian H, Colacurcio PL, Thomas JH. A transmembrane guanylyl cyclase (DAF-11) and Hsp90 (DAF-21) regulate a common set of chemosensory behaviors in caenorhabditis elegans. Genetics. 2000;155:85–104. doi: 10.1093/genetics/155.1.85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.van der Straten A, Rommel C, Dickson B, Hafen E. The heat shock protein 83 (Hsp83) is required for Raf-mediated signalling in Drosophila. EMBO J. 1997;16:1961–9. doi: 10.1093/emboj/16.8.1961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lai BT, Chin NW, Stanek AE, Keh W, Lanks KW. Quantitation and intracellular localization of the 85K heat shock protein by using monoclonal and polyclonal antibodies. Mol Cell Biol. 1984;4:2802–10. doi: 10.1128/mcb.4.12.2802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sreedhar AS, Kalmar E, Csermely P, Shen YF. Hsp90 isoforms: functions, expression and clinical importance. FEBS Lett. 2004;562:11–5. doi: 10.1016/s0014-5793(04)00229-7. [DOI] [PubMed] [Google Scholar]
- 6.Wandinger SK, Richter K, Buchner J. The Hsp90 chaperone machinery. J Biol Chem. 2008;283:18473–7. doi: 10.1074/jbc.R800007200. [DOI] [PubMed] [Google Scholar]
- 7.Richter K, Muschler P, Hainzl O, Buchner J. Coordinated ATP hydrolysis by the Hsp90 dimer. J Biol Chem. 2001;276:33689–96. doi: 10.1074/jbc.M103832200. [DOI] [PubMed] [Google Scholar]
- 8.Ali MM, Roe SM, Vaughan CK, Meyer P, Panaretou B, Piper PW, Prodromou C, Pearl LH. Crystal structure of an Hsp90-nucleotide-p23/Sba1 closed chaperone complex. Nature. 2006;440:1013–7. doi: 10.1038/nature04716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Richter K, Reinstein J, Buchner J. A Grp on the Hsp90 mechanism. Mol Cell. 2007;28:177–9. doi: 10.1016/j.molcel.2007.10.007. [DOI] [PubMed] [Google Scholar]
- 10.Soldano KL, Jivan A, Nicchitta CV, Gewirth DT. Structure of the N-terminal domain of GRP94. Basis for ligand specificity and regulation. J Biol Chem. 2003;278:48330–8. doi: 10.1074/jbc.M308661200. [DOI] [PubMed] [Google Scholar]
- 11.Shiau AK, Harris SF, Southworth DR, Agard DA. Structural Analysis of E. coli hsp90 reveals dramatic nucleotide-dependent conformational rearrangements. Cell. 2006;127:329–40. doi: 10.1016/j.cell.2006.09.027. [DOI] [PubMed] [Google Scholar]
- 12.Picard D, Khursheed B, Garabedian MJ, Fortin MG, Lindquist S, Yamamoto KR. Reduced levels of hsp90 compromise steroid receptor action in vivo. Nature. 1990;348:166–8. doi: 10.1038/348166a0. [DOI] [PubMed] [Google Scholar]
- 13.Stancato LF, Chow YH, Hutchison KA, Perdew GH, Jove R, Pratt WB. Raf exists in a native heterocomplex with hsp90 and p50 that can be reconstituted in a cell-free system. J Biol Chem. 1993;268:21711–6. [PubMed] [Google Scholar]
- 14.Taipale M, Krykbaeva I, Koeva M, Kayatekin C, Westover KD, Karras GI, Lindquist S. Quantitative analysis of HSP90-client interactions reveals principles of substrate recognition. Cell. 2012;150:987–1001. doi: 10.1016/j.cell.2012.06.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Arndt V, Rogon C, Hohfeld J. To be, or not to be--molecular chaperones in protein degradation. Cell Mol Life Sci. 2007;64:2525–41. doi: 10.1007/s00018-007-7188-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Johnson JL, Toft DO. A novel chaperone complex for steroid receptors involving heat shock proteins, immunophilins, and p23. J Biol Chem. 1994;269:24989–93. [PubMed] [Google Scholar]
- 17.Johnson JL, Toft DO. Binding of p23 and hsp90 during assembly with the progesterone receptor. Mol Endocrinol. 1995;9:670–8. doi: 10.1210/mend.9.6.8592513. [DOI] [PubMed] [Google Scholar]
- 18.Nelson GM, Prapapanich V, Carrigan PE, Roberts PJ, Riggs DL, Smith DF. The heat shock protein 70 cochaperone hip enhances functional maturation of glucocorticoid receptor. Mol Endocrinol. 2004;18:1620–30. doi: 10.1210/me.2004-0054. [DOI] [PubMed] [Google Scholar]
- 19.Pratt WB, Toft DO. Regulation of signaling protein function and trafficking by the hsp90/hsp70-based chaperone machinery. Exp Biol Med (Maywood) 2003;228:111–33. doi: 10.1177/153537020322800201. [DOI] [PubMed] [Google Scholar]
- 20.Hernandez MP, Chadli A, Toft DO. HSP40 binding is the first step in the HSP90 chaperoning pathway for the progesterone receptor. J Biol Chem. 2002;277:11873–81. doi: 10.1074/jbc.M111445200. [DOI] [PubMed] [Google Scholar]
- 21.Hernandez MP, Sullivan WP, Toft DO. The assembly and intermolecular properties of the hsp70-Hop-hsp90 molecular chaperone complex. J Biol Chem. 2002;277:38294–304. doi: 10.1074/jbc.M206566200. [DOI] [PubMed] [Google Scholar]
- 22.Gaiser AM, Brandt F, Richter K. The non-canonical Hop protein from Caenorhabditis elegans exerts essential functions and forms binary complexes with either Hsc70 or Hsp90. J Mol Biol. 2009;391:621–34. doi: 10.1016/j.jmb.2009.06.051. [DOI] [PubMed] [Google Scholar]
- 23.Schmid AB, Lagleder S, Grawert MA, Rohl A, Hagn F, Wandinger SK, Cox MB, Demmer O, Richter K, Groll M, Kessler H, Buchner J. The architecture of functional modules in the Hsp90 co-chaperone Sti1/Hop. EMBO J. 2012;31:1506–17. doi: 10.1038/emboj.2011.472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li J, Richter K, Buchner J. Mixed Hsp90-cochaperone complexes are important for the progression of the reaction cycle. Nat Struct Mol Biol. 2011;18:61–6. doi: 10.1038/nsmb.1965. [DOI] [PubMed] [Google Scholar]
- 25.Ebong IO, Morgner N, Zhou M, Saraiva MA, Daturpalli S, Jackson SE, Robinson CV. Heterogeneity and dynamics in the assembly of the heat shock protein 90 chaperone complexes. Proc Natl Acad Sci U S A. 2011;108:17939–44. doi: 10.1073/pnas.1106261108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Harst A, Lin H, Obermann WM. Aha1 competes with Hop, p50 and p23 for binding to the molecular chaperone Hsp90 and contributes to kinase and hormone receptor activation. Biochem J. 2005;387:789–96. doi: 10.1042/BJ20041283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Pratt WB, Toft DO. Steroid receptor interactions with heat shock protein and immunophilin chaperones. Endocr Rev. 1997;18:306–60. doi: 10.1210/edrv.18.3.0303. [DOI] [PubMed] [Google Scholar]
- 28.Morishima Y, Kanelakis KC, Murphy PJ, Lowe ER, Jenkins GJ, Osawa Y, Sunahara RK, Pratt WB. The hsp90 cochaperone p23 is the limiting component of the multiprotein hsp90/hsp70-based chaperone system in vivo where it acts to stabilize the client protein: hsp90 complex. J Biol Chem. 2003;278:48754–63. doi: 10.1074/jbc.M309814200. [DOI] [PubMed] [Google Scholar]
- 29.Pratt WB, Morishima Y, Osawa Y. The Hsp90 chaperone machinery regulates signaling by modulating ligand binding clefts. J Biol Chem. 2008;283:22885–9. doi: 10.1074/jbc.R800023200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bledsoe RK, Montana VG, Stanley TB, Delves CJ, Apolito CJ, McKee DD, Consler TG, Parks DJ, Stewart EL, Willson TM, Lambert MH, Moore JT, Pearce KH, Xu HE. Crystal structure of the glucocorticoid receptor ligand binding domain reveals a novel mode of receptor dimerization and coactivator recognition. Cell. 2002;110:93–105. doi: 10.1016/s0092-8674(02)00817-6. [DOI] [PubMed] [Google Scholar]
- 31.Simons SS Jr, Sistare FD, Chakraborti PK. Steroid binding activity is retained in a 16-kDa fragment of the steroid binding domain of rat glucocorticoid receptors. J Biol Chem. 1989;264:14493–7. [PubMed] [Google Scholar]
- 32.Silverstein AM, Galigniana MD, Chen MS, Owens-Grillo JK, Chinkers M, Pratt WB. Protein phosphatase 5 is a major component of glucocorticoid receptor. hsp90 complexes with properties of an FK506-binding immunophilin. J Biol Chem. 1997;272:16224–30. doi: 10.1074/jbc.272.26.16224. [DOI] [PubMed] [Google Scholar]
- 33.Chen MS, Silverstein AM, Pratt WB, Chinkers M. The tetratricopeptide repeat domain of protein phosphatase 5 mediates binding to glucocorticoid receptor heterocomplexes and acts as a dominant negative mutant. J Biol Chem. 1996;271:32315–20. doi: 10.1074/jbc.271.50.32315. [DOI] [PubMed] [Google Scholar]
- 34.Smith DF. Dynamics of heat shock protein 90-progesterone receptor binding and the disactivation loop model for steroid receptor complexes. Mol Endocrinol. 1993;7:1418–29. doi: 10.1210/mend.7.11.7906860. [DOI] [PubMed] [Google Scholar]
- 35.Ellis RJ, Dobson C, Hartl U. Sequence does specify protein conformation. Trends Biochem Sci. 1998;23:468. doi: 10.1016/s0968-0004(98)01324-3. [DOI] [PubMed] [Google Scholar]
- 36.Gano JJ, Simon JA. A proteomic investigation of ligand-dependent HSP90 complexes reveals CHORDC1 as a novel ADP-dependent HSP90-interacting protein. Mol Cell Proteomics. 2010;9:255–70. doi: 10.1074/mcp.M900261-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Basso AD, Solit DB, Chiosis G, Giri B, Tsichlis P, Rosen N. Akt forms an intracellular complex with heat shock protein 90 (Hsp90) and Cdc37 and is destabilized by inhibitors of Hsp90 function. J Biol Chem. 2002;277:39858–66. doi: 10.1074/jbc.M206322200. [DOI] [PubMed] [Google Scholar]
- 38.Farrell A, Morgan DO. Cdc37 promotes the stability of protein kinases Cdc28 and Cak1. Mol Cell Biol. 2000;20:749–54. doi: 10.1128/mcb.20.3.749-754.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Scholz G, Hartson SD, Cartledge K, Hall N, Shao J, Dunn AR, Matts RL. p50(Cdc37) can buffer the temperature-sensitive properties of a mutant of Hck. Mol Cell Biol. 2000;20:6984–95. doi: 10.1128/mcb.20.18.6984-6995.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Grammatikakis N, Lin JH, Grammatikakis A, Tsichlis PN, Cochran BH. p50(cdc37) acting in concert with Hsp90 is required for Raf-1 function. Mol Cell Biol. 1999;19:1661–72. doi: 10.1128/mcb.19.3.1661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Shao J, Irwin A, Hartson SD, Matts RL. Functional dissection of cdc37: characterization of domain structure and amino acid residues critical for protein kinase binding. Biochemistry. 2003;42:12577–88. doi: 10.1021/bi035138j. [DOI] [PubMed] [Google Scholar]
- 42.Shao J, Prince T, Hartson SD, Matts RL. Phosphorylation of serine 13 is required for the proper function of the Hsp90 co-chaperone, Cdc37. J Biol Chem. 2003;278:38117–20. doi: 10.1074/jbc.C300330200. [DOI] [PubMed] [Google Scholar]
- 43.Eckl JM, Rutz DA, Haslbeck V, Zierer BK, Reinstein J, Richter K. Cdc37 (cell division cycle 37) restricts Hsp90 (heat shock protein 90) motility by interaction with N-terminal and middle domain binding sites. J Biol Chem. 2013;288:16032–42. doi: 10.1074/jbc.M112.439257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Roe SM, Ali MM, Meyer P, Vaughan CK, Panaretou B, Piper PW, Prodromou C, Pearl LH. The Mechanism of Hsp90 regulation by the protein kinase-specific cochaperone p50(cdc37) Cell. 2004;116:87–98. doi: 10.1016/s0092-8674(03)01027-4. [DOI] [PubMed] [Google Scholar]
- 45.Siligardi G, Panaretou B, Meyer P, Singh S, Woolfson DN, Piper PW, Pearl LH, Prodromou C. Regulation of Hsp90 ATPase activity by the co-chaperone Cdc37p/p50cdc37. J Biol Chem. 2002;277:20151–9. doi: 10.1074/jbc.M201287200. [DOI] [PubMed] [Google Scholar]
- 46.Zhang W, Hirshberg M, McLaughlin SH, Lazar GA, Grossmann JG, Nielsen PR, Sobott F, Robinson CV, Jackson SE, Laue ED. Biochemical and structural studies of the interaction of Cdc37 with Hsp90. J Mol Biol. 2004;340:891–907. doi: 10.1016/j.jmb.2004.05.007. [DOI] [PubMed] [Google Scholar]
- 47.Shao J, Grammatikakis N, Scroggins BT, Uma S, Huang W, Chen JJ, Hartson SD, Matts RL. Hsp90 regulates p50(cdc37) function during the biogenesis of the activeconformation of the heme-regulated eIF2 alpha kinase. J Biol Chem. 2001;276:206–14. doi: 10.1074/jbc.M007583200. [DOI] [PubMed] [Google Scholar]
- 48.Caplan AJ, Mandal AK, Theodoraki MA. Molecular chaperones and protein kinase quality control. Trends Cell Biol. 2007;17:87–92. doi: 10.1016/j.tcb.2006.12.002. [DOI] [PubMed] [Google Scholar]
- 49.Gaiser AM, Kaiser CJ, Haslbeck V, Richter K. Downregulation of the Hsp90 system causes defects in muscle cells of Caenorhabditis elegans. PLoS One. 2011;6:e25485. doi: 10.1371/journal.pone.0025485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Litchfield DW. Protein kinase CK2: structure, regulation and role in cellular decisions of life and death. Biochem J. 2003;369:1–15. doi: 10.1042/BJ20021469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Vaughan CK, Mollapour M, Smith JR, Truman A, Hu B, Good VM, Panaretou B, Neckers L, Clarke PA, Workman P, Piper PW, Prodromou C, Pearl LH. Hsp90-dependent activation of protein kinases is regulated by chaperone-targeted dephosphorylation of Cdc37. Mol Cell. 2008;31:886–95. doi: 10.1016/j.molcel.2008.07.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Prince T, Matts RL. Definition of protein kinase sequence motifs that trigger high affinity binding of Hsp90 and Cdc37. J Biol Chem. 2004;279:39975–81. doi: 10.1074/jbc.M406882200. [DOI] [PubMed] [Google Scholar]
- 53.Zhao Q, Boschelli F, Caplan AJ, Arndt KT. Identification of a conserved sequence motif that promotes Cdc37 and cyclin D1 binding to Cdk4. J Biol Chem. 2004;279:12560–4. doi: 10.1074/jbc.M308242200. [DOI] [PubMed] [Google Scholar]
- 54.Terasawa K, Yoshimatsu K, Iemura S, Natsume T, Tanaka K, Minami Y. Cdc37 interacts with the glycine-rich loop of Hsp90 client kinases. Mol Cell Biol. 2006;26:3378–89. doi: 10.1128/MCB.26.9.3378-3389.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Vaughan CK, Gohlke U, Sobott F, Good VM, Ali MM, Prodromou C, Robinson CV, Saibil HR, Pearl LH. Structure of an Hsp90-Cdc37-Cdk4 complex. Mol Cell. 2006;23:697–707. doi: 10.1016/j.molcel.2006.07.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Street TO, Lavery LA, Agard DA. Substrate binding drives large-scale conformational changes in the Hsp90 molecular chaperone. Mol Cell. 2011;42:96–105. doi: 10.1016/j.molcel.2011.01.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Street TO, Lavery LA, Verba KA, Lee CT, Mayer MP, Agard DA. Cross-monomer substrate contacts reposition the Hsp90 N-terminal domain and prime the chaperone activity. J Mol Biol. 2012;415:3–15. doi: 10.1016/j.jmb.2011.10.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Polier S, Samant RS, Clarke PA, Workman P, Prodromou C, Pearl LH. ATP-competitive inhibitors block protein kinase recruitment to the Hsp90-Cdc37 system. Nat Chem Biol. 2013;9:307–12. doi: 10.1038/nchembio.1212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Millson SH, Truman AW, King V, Prodromou C, Pearl LH, Piper PW. A two-hybrid screen of the yeast proteome for Hsp90 interactors uncovers a novel Hsp90 chaperone requirement in the activity of a stress-activated mitogen-activated protein kinase, Slt2p (Mpk1p) Eukaryot Cell. 2005;4:849–60. doi: 10.1128/EC.4.5.849-860.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Hagn F, Lagleder S, Retzlaff M, Rohrberg J, Demmer O, Richter K, Buchner J, Kessler H. Structural analysis of the interaction between Hsp90 and the tumor suppressor protein p53. Nat Struct Mol Biol. 2011;18:1086–93. doi: 10.1038/nsmb.2114. [DOI] [PubMed] [Google Scholar]
- 61.Schreiber G, Keating AE. Protein binding specificity versus promiscuity. Curr Opin Struct Biol. 2011;21:50–61. doi: 10.1016/j.sbi.2010.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Jakob U, Lilie H, Meyer I, Buchner J. Transient interaction of Hsp90 with early unfolding intermediates of citrate synthase. Implications for heat shock in vivo. J Biol Chem. 1995;270:7288–94. doi: 10.1074/jbc.270.13.7288. [DOI] [PubMed] [Google Scholar]
- 63.Wang C, Chen J. Phosphorylation and hsp90 binding mediate heat shock stabilization of p53. J Biol Chem. 2003;278:2066–71. doi: 10.1074/jbc.M206697200. [DOI] [PubMed] [Google Scholar]
- 64.Whitesell L, Lindquist SL. HSP90 and the chaperoning of cancer. Nat Rev Cancer. 2005;5:761–72. doi: 10.1038/nrc1716. [DOI] [PubMed] [Google Scholar]
- 65.Haslbeck V, Eckl JM, Kaiser CJ, Papsdorf K, Hessling M, Richter K. Chaperone-interacting TPR proteins in Caenorhabditis elegans. J Mol Biol. 2013;425:2922–39. doi: 10.1016/j.jmb.2013.05.019. [DOI] [PubMed] [Google Scholar]
- 66.Etard C, Roostalu U, Strahle U. Shuttling of the chaperones Unc45b and Hsp90a between the A band and the Z line of the myofibril. J Cell Biol. 2008;180:1163–75. doi: 10.1083/jcb.200709128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Hawkins TA, Haramis AP, Etard C, Prodromou C, Vaughan CK, Ashworth R, Ray S, Behra M, Holder N, Talbot WS, Pearl LH, Strähle U, Wilson SW. The ATPase-dependent chaperoning activity of Hsp90a regulates thick filament formation and integration during skeletal muscle myofibrillogenesis. Development. 2008;135:1147–56. doi: 10.1242/dev.018150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Meyer BK, Petrulis JR, Perdew GH. Aryl hydrocarbon (Ah) receptor levels are selectively modulated by hsp90-associated immunophilin homolog XAP2. Cell Stress Chaperones. 2000;5:243–54. doi: 10.1379/1466-1268(2000)005<0243:aharla>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Petrulis JR, Perdew GH. The role of chaperone proteins in the aryl hydrocarbon receptor core complex. Chem Biol Interact. 2002;141:25–40. doi: 10.1016/s0009-2797(02)00064-9. [DOI] [PubMed] [Google Scholar]
- 70.Hu J, Toft DO, Seeger C. Hepadnavirus assembly and reverse transcription require a multi-component chaperone complex which is incorporated into nucleocapsids. EMBO J. 1997;16:59–68. doi: 10.1093/emboj/16.1.59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Hu J, Flores D, Toft D, Wang X, Nguyen D. Requirement of heat shock protein 90 for human hepatitis B virus reverse transcriptase function. J Virol. 2004;78:13122–31. doi: 10.1128/JVI.78.23.13122-13131.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sato S, Fujita N, Tsuruo T. Modulation of Akt kinase activity by binding to Hsp90. Proc Natl Acad Sci U S A. 2000;97:10832–7. doi: 10.1073/pnas.170276797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.An WG, Schulte TW, Neckers LM. The heat shock protein 90 antagonist geldanamycin alters chaperone association with p210bcr-abl and v-src proteins before their degradation by the proteasome. Cell Growth Differ. 2000;11:355–60. [PubMed] [Google Scholar]
- 74.Schulte TW, Blagosklonny MV, Romanova L, Mushinski JF, Monia BP, Johnston JF, Nguyen P, Trepel J, Neckers LM. Destabilization of Raf-1 by geldanamycin leads to disruption of the Raf-1-MEK-mitogen-activated protein kinase signalling pathway. Mol Cell Biol. 1996;16:5839–45. doi: 10.1128/mcb.16.10.5839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100:57–70. doi: 10.1016/s0092-8674(00)81683-9. [DOI] [PubMed] [Google Scholar]
- 76.Yufu Y, Nishimura J, Nawata H. High constitutive expression of heat shock protein 90 alpha in human acute leukemia cells. Leuk Res. 1992;16:597–605. doi: 10.1016/0145-2126(92)90008-u. [DOI] [PubMed] [Google Scholar]
- 77.Ciocca DR, Fuqua SA, Lock-Lim S, Toft DO, Welch WJ, McGuire WL. Response of human breast cancer cells to heat shock and chemotherapeutic drugs. Cancer Res. 1992;52:3648–54. [PubMed] [Google Scholar]
- 78.Jameel A, Skilton RA, Campbell TA, Chander SK, Coombes RC, Luqmani YA. Clinical and biological significance of HSP89 alpha in human breast cancer. Int J Cancer. 1992;50:409–15. doi: 10.1002/ijc.2910500315. [DOI] [PubMed] [Google Scholar]
- 79.Stebbins CE, Russo AA, Schneider C, Rosen N, Hartl FU, Pavletich NP. Crystal structure of an Hsp90-geldanamycin complex: targeting of a protein chaperone by an antitumor agent. Cell. 1997;89:239–50. doi: 10.1016/s0092-8674(00)80203-2. [DOI] [PubMed] [Google Scholar]
- 80.Grenert JP, Sullivan WP, Fadden P, Haystead TA, Clark J, Mimnaugh E, Krutzsch H, Ochel HJ, Schulte TW, Sausville E, Neckers LM, Toft DO. The amino-terminal domain of heat shock protein 90 (hsp90) that binds geldanamycin is an ATP/ADP switch domain that regulates hsp90 conformation. J Biol Chem. 1997;272:23843–50. doi: 10.1074/jbc.272.38.23843. [DOI] [PubMed] [Google Scholar]
- 81.Prodromou C, Roe SM, O’Brien R, Ladbury JE, Piper PW, Pearl LH. Identification and structural characterization of the ATP/ADP-binding site in the Hsp90 molecular chaperone. Cell. 1997;90:65–75. doi: 10.1016/s0092-8674(00)80314-1. [DOI] [PubMed] [Google Scholar]
- 82.Schulte TW, Akinaga S, Murakata T, Agatsuma T, Sugimoto S, Nakano H, Lee YS, Simen BB, Argon Y, Felts S, Toft DO, Neckers LM, Sharma SV. Interaction of radicicol with members of the heat shock protein 90 family of molecular chaperones. Mol Endocrinol. 1999;13:1435–48. doi: 10.1210/mend.13.9.0339. [DOI] [PubMed] [Google Scholar]
- 83.Xu W, Marcu M, Yuan X, Mimnaugh E, Patterson C, Neckers L. Chaperone-dependent E3 ubiquitin ligase CHIP mediates a degradative pathway for c-ErbB2/Neu. Proc Natl Acad Sci U S A. 2002;99:12847–52. doi: 10.1073/pnas.202365899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Chiosis G, Neckers L. Tumor selectivity of Hsp90 inhibitors: the explanation remains elusive. ACS Chem Biol. 2006;1:279–84. doi: 10.1021/cb600224w. [DOI] [PubMed] [Google Scholar]
- 85.Schulte TW, Neckers LM. The benzoquinone ansamycin 17-allylamino-17-demethoxygeldanamycin binds to HSP90 and shares important biologic activities with geldanamycin. Cancer Chemother Pharmacol. 1998;42:273–9. doi: 10.1007/s002800050817. [DOI] [PubMed] [Google Scholar]
- 86.Roughley SD, Hubbard RE. How well can fragments explore accessed chemical space? A case study from heat shock protein 90. J Med Chem. 2011;54:3989–4005. doi: 10.1021/jm200350g. [DOI] [PubMed] [Google Scholar]
- 87.Yu XM, Shen G, Neckers L, Blake H, Holzbeierlein J, Cronk B, Blagg BS. Hsp90 inhibitors identified from a library of novobiocin analogues. J Am Chem Soc. 2005;127:12778–9. doi: 10.1021/ja0535864. [DOI] [PubMed] [Google Scholar]
- 88.Donnelly A, Blagg BS. Novobiocin and additional inhibitors of the Hsp90 C-terminal nucleotide-binding pocket. Curr Med Chem. 2008;15:2702–17. doi: 10.2174/092986708786242895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Jhaveri K, Modi S. HSP90 inhibitors for cancer therapy and overcoming drug resistance. Adv Pharmacol. 2012;65:471–517. doi: 10.1016/B978-0-12-397927-8.00015-4. [DOI] [PubMed] [Google Scholar]
- 90.Luo W, Dou F, Rodina A, Chip S, Kim J, Zhao Q, Moulick K, Aguirre J, Wu N, Greengard P, Chiosis G. Roles of heat-shock protein 90 in maintaining and facilitating the neurodegenerative phenotype in tauopathies. Proc Natl Acad Sci U S A. 2007;104:9511–6. doi: 10.1073/pnas.0701055104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Dickey CA, Kamal A, Lundgren K, Klosak N, Bailey RM, Dunmore J, Ash P, Shoraka S, Zlatkovic J, Eckman CB, Patterson C, Dickson DW, Nahman NS Jr, Hutton M, Burrows F, Petrucelli L. The high-affinity HSP90-CHIP complex recognizes and selectively degrades phosphorylated tau client proteins. J Clin Invest. 2007;117:648–58. doi: 10.1172/JCI29715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Geller R, Vignuzzi M, Andino R, Frydman J. Evolutionary constraints on chaperone-mediated folding provide an antiviral approach refractory to development of drug resistance. Genes Dev. 2007;21:195–205. doi: 10.1101/gad.1505307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Geller R, Andino R, Frydman J. Hsp90 inhibitors exhibit resistance-free antiviral activity against respiratory syncytial virus. PLoS One. 2013;8:e56762. doi: 10.1371/journal.pone.0056762. [DOI] [PMC free article] [PubMed] [Google Scholar]