

Abstract
Purpose of review
Age-related muscle weakness causes a staggering economic, public, and personal burden. Most research has focused on internal muscular mechanisms as the root cause to strength loss. Here, we briefly discuss age-related impairments in the brain and peripheral nerve structures that may theoretically lead to muscle weakness in old age.
Recent findings
Neuronal atrophy in the brain is accompanied by electrical noise tied to declines in dopaminergic neurotransmission that degrades communication between neurons. Additionally, sensorimotor feedback loops that help regulate corticospinal excitability are impaired. In the periphery, there is evidence for motor unit loss, axonal atrophy, demyelination caused by oxidative damage to proteins and lipids, and modified transmission of the electrical signal through the neuromuscular junction.
Summary
Recent evidence clearly indicates that muscle weakness associated with aging is not entirely explained by classically postulated atrophy of muscle. In this issue, which focuses on ‘Ageing: Biology and Nutrition’ we will highlight new findings on how nervous system changes contribute to the aging muscle phenotype. These findings indicate that the ability to communicate neural activity to skeletal muscle is impaired with advancing age, which raises the question of whether many of these age-related neurological changes are mechanistically linked to impaired performance of human skeletal muscle. Collectively, this work suggests that future research should explore the direct link of these ‘upstream’ neurological adaptions and onset of muscle weakness in elders. In the long term, this new focus might lead to novel strategies to attenuate the age-related loss of muscle strength.
Keywords: dynapenia, sarcopenia, neuromuscular, muscle, brain, physical function
“What we have here is (a) failure to communicate,” said the Captain in the 1967 film Cool Hand Luke. This line rings true today as it relates to the failure of physiologists to communicate the mechanisms of muscle strength to the geriatrics community, where the lack of muscle strength observed in older adults holds high clinical significance. Similarly, there is a relative under recognition in the scientific community for the potential role of the brain’s failure to communicate with skeletal muscle as a central component of muscle weakness in older adults. For the better part of the last quarter century scientific endeavors have primarily focused on the role of muscle wasting (sarcopenia) in explaining strength loss in seniors [1], with relatively little attention paid to understanding the role of the nervous system despite calls for investigations of this nature from preeminent scientists more than 25-years ago [2]. Developing a detailed understanding of the brain, which is commonly referred to as “the final frontier of science”, is still in its relative infancy, but there are already several key observations that clearly attest to the power of the mind as it relates to muscle force production. For example, findings that training with mental imagery of strong muscle contractions increases muscle strength also implicates the brain and its ability to produce a descending command as a key mechanistic determinant of maximal voluntary muscle strength [3]. Collectively, these findings provide general proof-of-concept support for the nervous system, at times at least, being a limiting factor in muscle performance. In this article we will highlight key findings on age-related changes in the nervous system, which theoretically may be linked to impaired performance of human skeletal muscle.
Aging and the Neuromuscular System
It is well established that aging is associated with dramatic reductions in muscle strength (dynapenia) and motor performance [4]. For example, data from the most recent longitudinal aging study suggest that muscle strength decreases at a staggering rate of ~ 3%/year between the ages of 70–79 years [5]. The resultant muscle weakness is independently associated with the development of disability, impairment of functional capacity [6], fall risk [7], and even mortality [8]. While it is clear that senescence of muscle and nervous systems are key targets for understanding declines in voluntary strength, this article will focus its efforts on neural characteristics (See Figure 1 for overview of targeted areas) [4].
Figure 1.
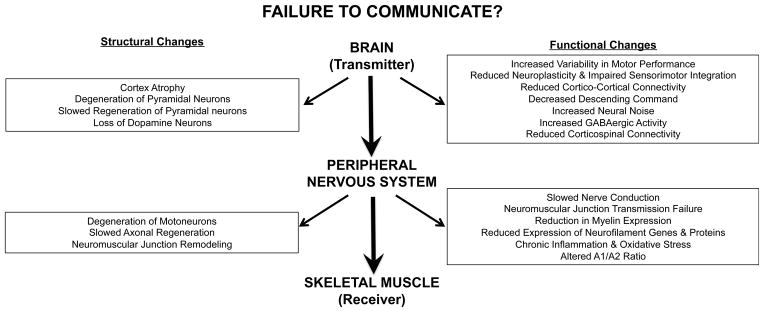
Illustration of target areas in the nervous system that theoretically can limit muscle performance.
Aging and Brain Structure
There are over 100 billion cells in the brain with the cerebral cortex containing between 17 and 26 billion neurons [9, 10]. Neurons in the brain (as well as the spinal cord) essentially come in two flavors, excitatory neurons that transmit and amplify signals, and inhibitory neurons that inhibit and refine those signals. The relative balance of excitatory and inhibitory synaptic inputs determines whether or not a neuronal event occurs (e.g., an action potential). The neurons in the pre-motor and motor cortex form a complex network of glutamatergic interneurons, afferent projections, and pyramidal neurons that project to several areas of the central nervous system that include the striatum and spinal cord. The main output cells of the human motor cortex are pyramidal cells, which use the excitatory amino acid glutamate as their neurotransmitter [11], and terminate directly on motor neurons in the ventral horn of the spinal cord, providing the most direct pathway for movement execution [11]. The non-pyramidal stellate cells, which comprise 25–30% of cortical neurons in the motor cortex, do not project beyond the cortex. Stellate cells are divided into spiny and non-spiny cell types, with spiny stellate cells being primarily located in layer IV and using glutamate as their neurotransmitter and non-spiny stellate cells being located in all layers and using the neurotransmitter γ-aminobutyric acid (GABA) to make inhibitory synapses with pyramidal cells [11]. The GABAergic inhibitory system is largely responsible for the task specificity of pyramidal tract neurons in the motor cortex, plays an essential role in isolating movements, and is also important in neuroplasticity.
Neuronal atrophy, but not the loss of motor cortical neurons per se, primarily occurs with aging [12]. For example, high resolution structural magnetic resonance imaging has been used to demonstrate that prominent atrophy occurs as early as middle age in many regions of the cerebral cortex including the primary motor cortex [13]. In addition to changes in the overall size of the motor cortex, there is also evidence that age-related differences exist in white matter mass and length of myelinated nerve fibers [14]. Specifically, Marner and colleagues examined brain tissue from 36 individuals ranging in age from 18 to 93 years and found that individuals lose ~ 45% of their of myelinated fiber length in the brain white matter, with this reduction being particularly pronounced in the smallest nerve fibers [14]. It has been argued that because senescence does not lead to a widespread loss of cortical neurons, the degeneration of pyramidal neurons can occur without the loss of the bodies of their parent cells [15]. Thus, aging leads, in part, to an inability to regenerate axons following degeneration due to a decline in the rate of transport of the materials necessary for axonal regeneration [15]. From a functional standpoint it seems likely that these age-related changes in the cerebral cortex would affect cortico-cortical and corticospinal connectivity potentially leading to impaired muscle strength.
Aging, Neural Noise, & Human Motor Cortex Plasticity
As the brain ages, its capacity to transmit signals and communicate is decreased. Central to such declines in aging is an increase in neural noise, that is, a greater presence of random background activity in the brain signal. Effectively, the electrical signals transmitted by the aging brain are analogous to a fuzzy television signal received through a satellite receiver or antenna. Much like the poor reception on the television, the precision of the neural information being transmitted becomes inaccurate when contaminated by noise (for review see [16]). In this sense, the activation of neurons and motor units becomes unpredictable as noise results in the strength of the electrical signal sent across the nervous system to randomly fall below the threshold of activation. There is now ample empirical evidence of increased intra-individual variability in both cognitive and motor behavior in aging (for review see reference [17]).
Increased neural noise is often related to declines in dopaminergic neurotransmission, where aging leads to the loss of DA neurons in the striatum[18]. Recent research using PET (positron emission tomography) shows that increased reaction times in the elderly is associated with the loss of dopamine (D1) receptors [19]. Not only does decreased dopamine lead to increased neural noise, it also results in scattered or unfocused patterns of behavior, effectively, reducing the ability to harness available neural resources (for review see reference [20]). A second mechanism of increased noise is the presence of excessively high levels of extracellular glutamate around the neurons. When glutamate uptake is blocked, increased noise in neural signals is observed [21]. Aging is associated with alters the interaction between glutamate, dopamine, and GABA [22]. Overall, the effects of aging lead to reduced dopamine release and glutamate uptake, with both leading to potential effects on the ability to produce muscle force and motor control.
A number of studies in recent years indicate that while the aging brain retains its plasticity, its neuroplastic and neuromodulatory capabilities are frequently diminished [23–26]. Several transcranial magnetic stimulation (TMS) studies, which can provide insight into the GABAergic inhibitory system [27, 28] have illustrated that older adults have impaired sensorimotor integration of afferent input. For example, older adults have reduced modulatory capacity of short-interval intracortical inhibition (primarily reflecting GABA-mediated inhibition) during response preparation for a motor task [23], and exhibit reductions in the ability to modulate motor cortex excitability in response to electrical stimulation [24, 25]. Because GABAergic neurons inhibit dopamine release, it is plausible that this reduced capacity for inhibitory modulation in aging is associated with increased neural noise.
Mechanisms underlying reduced motor cortex plasticity in elders are poorly understood. It is possible that age-related declines in long-term potentiation [29], neurotransmitters [30], or gene expression [31] important for synaptic plasticity are partly responsible. Unfortunately, the functional relevance of age-related changes in neurotransmitters and neuroplasticity in the cerebral cortex as it relates to muscle weakness is poorly understood. The majority of studies examining the relationships between physiologic and functional parameters have primarily focused on associations with motor control and manual dexterity, but not maximal muscle strength [32, 33]. As such, further work is needed to better understand whether the myriad morphological and functional aging adaptations to the brain have a functional significance as it relates to muscle weakness.
Aging of motor units and peripheral nerves
A motor unit (MU) comprises a single peripheral neuron and its innervated muscle fibers [11]. They serve as the ‘final common pathway’ for all motor commands, with each motor neuron typically having ~ 50,000 synaptic inputs to convey these commands and thus their behavior can be influenced by many factors [11]. Aging is associated with both morphological, physiological, and behavioral changes in motor units [34]. Regarding the former, MU’s are gradually lost over the first six decades of life, but accelerates thereafter [35] – a pattern that closely mimics dynapenia [36]. There are a countless number of physiological and behavioral changes that include reduced amplitude, altered synchronization and more variable discharges of motor units during muscle contractions [37]. Additionally, the conduction velocities of efferent axons demonstrate a marked reduction in late-life [38]. Axonal atrophy and remodeling of the NMJ might provide pathological explanations and are discussed below.
The axons of aged animals and humans exhibit declines in axonal transport, degenerated mitochondria, and accumulations of filaments [39–41]. These changes may culminate in axonal atrophy that is often seen in neurodegenerative diseases, but also seem to occur with senescence [42]. It has been established that myelinated peripheral nerves from aged animals have axon loss and morphological irregularities, as well as a notable reduction in the expression of myelin, neurofilament genes and proteins [43]. Damaged axons undergo a complex degeneration and regeneration process that is impaired and delayed with aging [44]. The process, called “dying back” occurs where an impaired axon progressively regresses towards the cell body [42]. This is the most common pathology when the axon is faced with toxic, metabolic or infectious injuries (e.g. Neuropathy, Alzheimer’s, Parkinson’s, etc.) [44]. The onset of these pathologies is more common in the peripheral nervous system (PNS) because it is not protected by the blood brain barrier [45]. Moreover, aging is associated with elevated levels of chronic inflammation and oxidative stress that could aid in causing an injury to the axon to initiate degeneration. Because Schwann cells — the myelinated glia of the PNS— are rich in fatty acids, they serve as a major substrate for reactive oxygen species and are thought to be particularly vulnerable to the accumulation of oxidative damage seen with aging [46]. Recently, Opalach and colleagues demonstrated that aged animals had a pronounced accumulation of ubiquitinated and oxidatively damaged proteins within myelinated peripheral nerves [47]. This damage subsequently resulted in an immunologic response, which theoretically would result in further damage or impairment of the regeneration process.
Alternatively, genetic factors, which can also influence age-related neurological conditions, might also regulate axonal regeneration. For example, the gene that codes for apolipoprotein E isoform 4 (ApoE4) was studied in relationship to peripheral nerve regeneration [48]. The ApoE4 allele is present in one-third of the population and when coupled with aging is most commonly associated with risk of neurodegeneration in the CNS. However, ApoE4 is expressed throughout the PNS and could have functional significance [48]. ApoE4 expression significantly disrupts nerve regeneration and subsequent neuromuscular junction re-innervation following nerve crush [48]. Collectively, age-related peripheral nerve degeneration and impaired regeneration seem to follow similar patterns found in neurodegenerative diseases [45]. The altered cellular milieu due to aging places axons in a vulnerable environment for degeneration and genetic predisposition to impaired regenerative capacity has the potential to impair motor coordination and muscle strength.
Aging is associated with NMJ remodeling that is characterized by retraction of nerve terminal endings resulting in unoccupied post-synaptic receptors [49]. As a result, aged NMJ’s expand with a larger motor endplate. It is logical to surmise that NMJ remodeling is a consequence of muscle atrophy. However, these modifications can precede muscle fiber atrophy and thus could be involved with initiating muscle loss [49]. Interestingly, axons separated from their NMJ showed no evidence of atrophy, suggesting that a local (e.g. spinal cord), as opposed to an upstream, mechanism might be responsible [50]. Li and colleagues have recently challenged the notion that NMJ remodeling occurs prior to muscle fiber atrophy. They found that NMJs mostly remain stable throughout the lifespan, but become fragmented during regeneration that occurs following an injury to the muscle fiber [51]. The specific mechanisms aren’t completely understood, but these new data suggest that post-synaptic necrosis and impaired regeneration of muscle fibers might initiate NMJ remodeling seen with aging.
Transmission at the NMJ is also altered with aging [52]. Aged nerve terminal endings release larger amounts of neurotransmitter upon stimulation, which result in a higher endplate potential at the muscle fiber [53]. While this seems like an effective compensatory strategy to improve NMJ transmission, doing so leads to a faster rate of NMJ failure often seen in aged animals. Additionally, adenosine receptors, which act to control evoked endplate potential at the NMJ are modified with aging [51]. Specifically, aging changes the ratio of inhibitory A1 receptors and excitatory adenosine A2A receptors that are key components of regulating the endplate response to stimulation [11]. The magnitude of excitatory effects of the A1/A2A receptors recedes while the inhibitory regulation stays constant with increasing age [51]. Therefore, for an unknown reason, the NMJ preferentially selects to activate inhibitory over excitatory receptors. The selection of inhibitory over excitatory receptors at the NMJ with aging should theoretically decrease the ability to produce muscle force.
Conclusions and Perspectives
Although age-related strength loss originates from multiple sources, the literature reviewed here suggests that a significant component is the breakdown in communication between brain and muscle. With aging, the changes in the central and peripheral nervous system may reduce an individual’s ability to activate available musculature. While there is a strong theoretical rationale for connecting brain and muscle, there is a general lack of evidence that shows brain aging is associated with muscle strength impairment in older adults. Based on the literature reviewed here, clinical approaches to maintaining strength levels in seniors has to be extended beyond targeting skeletal muscle size and intramuscular factors and be combined with novel approaches that sustain nervous system function.
Key points.
Age-related changes in the nervous system, at times at least, can be a limiting factor in muscle performance.
Individuals over 65 years of age exhibit a large volumetric reduction in areas of the brain that are responsible for generating voluntary muscle contraction.
The loss in dopamine neurons with aging might contribute to neural noise, which can lead to a reduced ability to harness available resources for muscle performance
Aging is associated with a reduced peripheral nerve regeneration and neuromuscular junction remodeling that are theoretically linked with poor muscle performance.
Acknowledgments
This work was partially supported by the following organizations: National Institute on Aging (R21AG031974 to T.M. Manini) and Eunice Kennedy Shriver National Institute of Child Health and Human Development (R15HD065552 to B. C. Clark). T.M. Manini was also support by the University of Florida Claude D. Pepper Center awarded by the National Institute on Aging (P30AG028740).
Footnotes
Conflicts of interest
B.C. Clark has received consulting fees from Regeneron Pharmaceuticals, Inc. and Abbott Laboratories. No other conflicts were reported.
References
Papers of particular interest, published with the annual period of review, have been highlighted as:
* of special interest
** of outstanding interest
- 1**.Manini TM, Clark BC. Dynapenia and Aging: An Update. The journals of gerontology. Series A, Biological sciences and medical sciences. 2011 doi: 10.1093/gerona/glr010. This is an literature update to the original sarcopenia and dynapenia article published in 2008. The article reviews existing data in the literature to compares muscle strength and mass for predicting age-related health conditions in older adults. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Enoka RM. Muscle strength and its development. Sports Med. 1988;6:146–168. doi: 10.2165/00007256-198806030-00003. [DOI] [PubMed] [Google Scholar]
- 3.Ranganathan VK, Siemionow V, Liu JZ, Sahgal V, Yue GH. From mental power to muscle power--gaining strength by using the mind. Neuropsychologia. 2004;42(7):944–56. doi: 10.1016/j.neuropsychologia.2003.11.018. [DOI] [PubMed] [Google Scholar]
- 4.Clark B, Issac LC, Lane JL, Damron LA, Hoffman RL. Neuromuscular plasticity during and following 3-weeks of human forearm cast immobilization. J Appl Physiol. 2008;105:868–878. doi: 10.1152/japplphysiol.90530.2008. [DOI] [PubMed] [Google Scholar]
- 5.Delmonico MJ, Harris TB, Visser M, Park SW, Conroy MB, Velasquez-Mieyer P, Boudreau R, Manini TM, Nevitt M, Newman AB, Goodpaster BH. Longitudinal study of muscle strength, quality, and adipose tissue infiltration. Am J Clin Nutr. 2009;90(6):1579–85. doi: 10.3945/ajcn.2009.28047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Manini TM, Visser M, Won-Park S, Patel KV, Strotmeyer ES, Chen H, Goodpaster B, De Rekeneire N, Newman AB, Simonsick EM, Kritchevsky SB, Ryder K, Schwartz AV, Harris TB. Knee extension strength cutpoints for maintaining mobility. Journal of the American Geriatrics Society. 2007;55(3):451–7. doi: 10.1111/j.1532-5415.2007.01087.x. [DOI] [PubMed] [Google Scholar]
- 7.de Rekeneire N, Visser M, Peila R, Nevitt MC, Cauley JA, Tylavsky FA, Simonsick EM, Harris TB. Is a fall just a fall: correlates of falling in healthy older persons. The Health, Aging and Body Composition Study. J Am Geriatr Soc. 2003;51(6):841–6. doi: 10.1046/j.1365-2389.2003.51267.x. [DOI] [PubMed] [Google Scholar]
- 8.Newman AB, Kupelian V, Visser M, Simonsick EM, Goodpaster BH, Kritchevsky SB, Tylavsky FA, Rubin SM, Harris TB. Strength, but not muscle mass, is associated with mortality in the health, aging and body composition study cohort. J Gerontol A Biol Sci Med Sci. 2006;61(1):72–7. doi: 10.1093/gerona/61.1.72. [DOI] [PubMed] [Google Scholar]
- 9.Azevedo FA, Carvalho LR, Grinberg LT, Farfel JM, Ferretti RE, Leite RE, Jacob Filho W, Lent R, Herculano-Houzel S. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. The Journal of comparative neurology. 2009;513(5):532–41. doi: 10.1002/cne.21974. [DOI] [PubMed] [Google Scholar]
- 10.Pelvig DP, Pakkenberg H, Stark AK, Pakkenberg B. Neocortical glial cell numbers in human brains. Neurobiology of aging. 2008;29(11):1754–62. doi: 10.1016/j.neurobiolaging.2007.04.013. [DOI] [PubMed] [Google Scholar]
- 11.Kandel ER, Schwartz JH, Jessell TM, Siegelbaum SA, Hudspeth AJ. Principles of Neural Science. 5. McGraw-Hill Companies, Inc; 2012. [Google Scholar]
- 12.Ward NS. Compensatory mechanisms in the aging motor system. Ageing Res Rev. 2006;5(3):239–54. doi: 10.1016/j.arr.2006.04.003. [DOI] [PubMed] [Google Scholar]
- 13.Salat DH, Buckner RL, Snyder AZ, Greve DN, Desikan RS, Busa E, Morris JC, Dale AM, Fischl B. Thinning of the cerebral cortex in aging. Cereb Cortex. 2004;14(7):721–30. doi: 10.1093/cercor/bhh032. [DOI] [PubMed] [Google Scholar]
- 14.Marner L, Nyengaard JR, Tang Y, Pakkenberg B. Marked loss of myelinated nerve fibers in the human brain with age. J Comp Neurol. 2003;462(2):144–52. doi: 10.1002/cne.10714. [DOI] [PubMed] [Google Scholar]
- 15*.Pannese E. Morphological changes in nerve cells during normal aging. Brain structure & function. 2011;216(2):85–9. doi: 10.1007/s00429-011-0308-y. This is a review article that discusses changes in nerve cell structure associated with advancing age. [DOI] [PubMed] [Google Scholar]
- 16.Faisal AA, Selen LP, Wolpert DM. Noise in the nervous system. Nature reviews Neuroscience. 2008;9(4):292–303. doi: 10.1038/nrn2258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nesselroade JR, Salthouse TA. Methodological and theoretical implications of intraindividual variability in perceptual-motor performance. The journals of gerontology Series B, Psychological sciences and social sciences. 2004;59(2):49–55. doi: 10.1093/geronb/59.2.p49. [DOI] [PubMed] [Google Scholar]
- 18.Darbin O. The aging striatal dopamine function. Parkinsonism & related disorders. 2012;18(5):426–32. doi: 10.1016/j.parkreldis.2011.11.025. [DOI] [PubMed] [Google Scholar]
- 19**.MacDonald SW, Karlsson S, Rieckmann A, Nyberg L, Backman L. Aging-related increases in behavioral variability: relations to losses of dopamine D1 receptors. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2012;32(24):8186–91. doi: 10.1523/JNEUROSCI.5474-11.2012. This article uses neuroimaging to link increased variability in cognitive responses to loss of dopamine receptors in key brain regions, specifically anterior cingulate gyrus, dorsolateral prefrontal cortex, and parietal cortex. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hills TT. Animal foraging and the evolution of goal-directed cognition. Cognitive science. 2006;30(1):3–41. doi: 10.1207/s15516709cog0000_50. [DOI] [PubMed] [Google Scholar]
- 21.Arnth-Jensen N, Jabaudon D, Scanziani M. Cooperation between independent hippocampal synapses is controlled by glutamate uptake. Nature neuroscience. 2002;5(4):325–31. doi: 10.1038/nn825. [DOI] [PubMed] [Google Scholar]
- 22.Mora F, Segovia G, Del Arco A. Glutamate-dopamine-GABA interactions in the aging basal ganglia. Brain Res Rev. 2008;58(2):340–53. doi: 10.1016/j.brainresrev.2007.10.006. [DOI] [PubMed] [Google Scholar]
- 23.Fujiyama H, Hinder MR, Schmidt MW, Tandonnet C, Garry MI, Summers JJ. Age-related differences in corticomotor excitability and inhibitory processes during a visuomotor RT task. Journal of cognitive neuroscience. 2012;24(5):1253–63. doi: 10.1162/jocn_a_00201. [DOI] [PubMed] [Google Scholar]
- 24***.Smith AE, Ridding MC, Higgins RD, Wittert GA, Pitcher JB. Cutaneous afferent input does not modulate motor intracortical inhibition in ageing men. The European journal of neuroscience. 2011;34(9):1461–9. doi: 10.1111/j.1460-9568.2011.07869.x. This article provides evidence to suggest that a contributing factor in the decline of motor function with ageing is loss of short-intracortical inhibiiton modulation that is likely attributed to altered cortical sensorimotor integration of afferent input. [DOI] [PubMed] [Google Scholar]
- 25.Degardin A, Devos D, Cassim F, Bourriez JL, Defebvre L, Derambure P, Devanne H. Deficit of sensorimotor integration in normal aging. Neuroscience letters. 2011;498(3):208–12. doi: 10.1016/j.neulet.2011.05.010. [DOI] [PubMed] [Google Scholar]
- 26.Rogasch NC, Dartnall TJ, Cirillo J, Nordstrom MA, Semmler JG. Corticomotor plasticity and learning of a ballistic thumb training task are diminished in older adults. Journal of applied physiology. 2009;107(6):1874–83. doi: 10.1152/japplphysiol.00443.2009. [DOI] [PubMed] [Google Scholar]
- 27.Kobayashi M, Pascual-Leone A. Transcranial magnetic stimulation in neurology. Lancet Neurol. 2003;2(3):145–56. doi: 10.1016/s1474-4422(03)00321-1. [DOI] [PubMed] [Google Scholar]
- 28.Reis J, Swayne OB, Vandermeeren Y, Camus M, Dimyan MA, Harris-Love M, Perez MA, Ragert P, Rothwell JC, Cohen LG. Contribution of transcranial magnetic stimulation to the understanding of cortical mechanisms involved in motor control. J Physiol. 2008;586(2):325–51. doi: 10.1113/jphysiol.2007.144824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Burke SN, Barnes CA. Neural plasticity in the ageing brain. Nature reviews Neuroscience. 2006;7(1):30–40. doi: 10.1038/nrn1809. [DOI] [PubMed] [Google Scholar]
- 30.Clayton DA, Grosshans DR, Browning MD. Aging and surface expression of hippocampal NMDA receptors. The Journal of biological chemistry. 2002;277(17):14367–9. doi: 10.1074/jbc.C200074200. [DOI] [PubMed] [Google Scholar]
- 31.Pang PT, Teng HK, Zaitsev E, Woo NT, Sakata K, Zhen S, Teng KK, Yung WH, Hempstead BL, Lu B. Cleavage of proBDNF by tPA/plasmin is essential for long-term hippocampal plasticity. Science. 2004;306(5695):487–91. doi: 10.1126/science.1100135. [DOI] [PubMed] [Google Scholar]
- 32.Clark BC, Taylor JL. Age-Related Changes in Motor Cortical Properties and Voluntary Activation of Skeletal Muscle. Current aging science. 2011 doi: 10.2174/1874609811104030192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Marneweck M, Loftus A, Hammond G. Short-interval intracortical inhibition and manual dexterity in healthy aging. Neuroscience research. 2011;70(4):408–14. doi: 10.1016/j.neures.2011.04.004. [DOI] [PubMed] [Google Scholar]
- 34.Deschenes MR. Motor unit and neuromuscular junction remodeling with aging. Current aging science. 2011;4(3):209–20. doi: 10.2174/1874609811104030209. [DOI] [PubMed] [Google Scholar]
- 35.Gordon T, Hegedus J, Tam SL. Adaptive and maladaptive motor axonal sprouting in aging and motoneuron disease. Neurological research. 2004;26(2):174–85. doi: 10.1179/016164104225013806. [DOI] [PubMed] [Google Scholar]
- 36.Lauretani F, Russo CR, Bandinelli S, Bartali B, Cavazzini C, Di Iorio A, Corsi AM, Rantanen T, Guralnik JM, Ferrucci L. Age-associated changes in skeletal muscles and their effect on mobility: an operational diagnosis of sarcopenia. J Appl Physiol. 2003;95(5):1851–60. doi: 10.1152/japplphysiol.00246.2003. [DOI] [PubMed] [Google Scholar]
- 37.Shinohara M. Adaptations in motor unit behavior in elderly adults. Curr Aging Sci. 2011;4(3):200–208. doi: 10.2174/1874609811104030200. [DOI] [PubMed] [Google Scholar]
- 38.Di Iorio A, Cherubini A, Volpato S, Sparvieri E, Lauretani F, Franceschi C, Senin U, Abate G, Paganelli R, Martin A, Andres-Lacueva C, Ferrucci L. Markers of inflammation, vitamin E and peripheral nervous system function: the InCHIANTI study. Neurobiol Aging. 2006;27(9):1280–8. doi: 10.1016/j.neurobiolaging.2005.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Coleman M. Molecular signaling how do axons die? Advances in genetics. 2011;73:185–217. doi: 10.1016/B978-0-12-380860-8.00005-7. [DOI] [PubMed] [Google Scholar]
- 40.Cheng A, Hou Y, Mattson MP. Mitochondria and neuroplasticity. ASN neuro. 2010;2(5):e00045. doi: 10.1042/AN20100019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bahr BA, Bendiske J. The neuropathogenic contributions of lysosomal dysfunction. Journal of neurochemistry. 2002;83(3):481–9. doi: 10.1046/j.1471-4159.2002.01192.x. [DOI] [PubMed] [Google Scholar]
- 42.Misgeld T. Lost in elimination: mechanisms of axonal loss. e-Neuroforum. 2011;2(2):21–34. [Google Scholar]
- 43.Melcangi RC, Magnaghi V, Cavarretta I, Martini L, Piva F. Age-induced decrease of glycoprotein Po and myelin basic protein gene expression in the rat sciatic nerve. Repair by steroid derivatives. Neuroscience. 1998;85(2):569–78. doi: 10.1016/s0306-4522(97)00628-3. [DOI] [PubMed] [Google Scholar]
- 44.Kovacic U, Sketelj J, Bajrovic FF. Chapter 26: Age-related differences in the reinnervation after peripheral nerve injury. International review of neurobiology. 2009;87:465–82. doi: 10.1016/S0074-7742(09)87026-8. [DOI] [PubMed] [Google Scholar]
- 45.Mattson MP, Magnus T. Ageing and neuronal vulnerability. Nature reviews Neuroscience. 2006;7(4):278–94. doi: 10.1038/nrn1886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Selman C, Blount JD, Nussey DH, Speakman JR. Oxidative damage, ageing, and life-history evolution: where now? Trends in ecology & evolution. 2012 doi: 10.1016/j.tree.2012.06.006. [DOI] [PubMed] [Google Scholar]
- 47.Opalach K, Rangaraju S, Madorsky I, Leeuwenburgh C, Notterpek L. Lifelong calorie restriction alleviates age-related oxidative damage in peripheral nerves. Rejuvenation Res. 2010;13(1):65–74. doi: 10.1089/rej.2009.0892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48**.Comley LH, Fuller HR, Wishart TM, Mutsaers CA, Thomson D, Wright AK, Ribchester RR, Morris GE, Parson SH, Horsburgh K, Gillingwater TH. ApoE isoform-specific regulation of regeneration in the peripheral nervous system. Human molecular genetics. 2011;20(12):2406–21. doi: 10.1093/hmg/ddr147. The article illustrates that the importance of Apolipoprotein E4 as a candidate gene that regulates regeneration of peripheral nerves. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49*.Deschenes MR. Motor Unit and Neuromuscular Junction Remodeling with Aging. Current aging science. 2011 doi: 10.2174/1874609811104030209. This is a recent review article that gives excellent details regarding the age-related remodeling of the neuromuscular junction. [DOI] [PubMed] [Google Scholar]
- 50.Prakash YS, Sieck GC. Age-related remodeling of neuromuscular junctions on type-identified diaphragm fibers. Muscle & Nerve. 1998;21(7):887–95. doi: 10.1002/(sici)1097-4598(199807)21:7<887::aid-mus6>3.0.co;2-2. [DOI] [PubMed] [Google Scholar]
- 51**.Li Y, Lee Y, Thompson WJ. Changes in aging mouse neuromuscular junctions are explained by degeneration and regeneration of muscle fiber segments at the synapse. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2011;31(42):14910–9. doi: 10.1523/JNEUROSCI.3590-11.2011. This is a key article that suggests remodeling of the neuromuscular junction is dependent on necrotic events that occur in the muscle fiber. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Jang YC, Van Remmen H. Age-associated alterations of the neuromuscular junction. Experimental gerontology. 2011;46(2–3):193–8. doi: 10.1016/j.exger.2010.08.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53*.Deschenes MR, Roby MA, Glass EK. Aging influences adaptations of the neuromuscular junction to endurance training. Neuroscience. 2011 doi: 10.1016/j.neuroscience.2011.05.070. This article demonstrates that the neuromuscular junction is less sensitive to remodeling due to exercise training in old compared to young animals. It also reveals that exercise adaptations and effects of aging are fiber type dependent. [DOI] [PMC free article] [PubMed] [Google Scholar]