

Abstract
Early arrival at the breeding site positively affects the breeding success of migratory birds. During migration, birds spend most of their time at stopovers. Therefore, determining which factors shape stopover duration is essential to our understanding of avian migration. Because the main purpose of stopover is to accumulate fat as fuel for the next flight bout, fuel reserves at arrival and the accumulation of fuel are both expected to affect stopover departure decisions. Here, we determined whether fuel reserves and fuel accumulation predict a bird's motivation to depart, as quantified by nocturnal migratory restlessness (Zugunruhe), using northern wheatears (Oenanthe oenanthe) that were captured and temporarily contained at spring stopover. We found that fuel reserves at capture were positively correlated with Zugunruhe, and negatively correlated with fuel accumulation. This indicates that fat birds were motivated to depart, whereas lean birds were set on staying and accumulating fuel. Moreover, the change in fuel reserves was positively correlated with the concurrent change in Zugunruhe, providing the first empirical evidence for a direct link between fuel accumulation and Zugunruhe during stopover. Our study indicates that, together with innate rhythms and weather, the size and accumulation of fuel reserves shape stopover duration, and hence overall migration time.
Keywords: fuel reserves, migratory restlessness, Zugunruhe, stopover, migration, bird
1. Introduction
In migratory birds, early arrival at the breeding site positively affects breeding success [1,2]. Most migrants alternate flight bouts with one or more stopovers, during which they replenish the fuel (in the form of fat) used during flight (refuelling). Because the time spent at stopover sites by far exceeds the time spent flying [3,4], determining which factors shape stopover duration is essential to our understanding of avian migration [5]. The size of the fuel reserves (fuel load) remaining when birds land for stopover as well as the speed at which fuel is accumulated may be important determinants of the decision when to depart again. Results from field studies, however, are mixed; some found that fat birds were more likely to depart than lean conspecifics [6–8], whereas in other studies stopover duration was unrelated to fuel load at capture [9,10]. Also, stopover duration is usually unrelated to the speed of fuel accumulation (reviewed in [11], but see [12]). However, the determination of stopover duration is notoriously difficult, because exact arrival time is usually unknown [13], and capture–recapture rates can be biased, for example, when lean birds are more active than fat birds [6,9]. Furthermore, other variables such as weather conditions and food availability can affect departure decisions (reviewed in [11]) and may consequently obscure the relationship between fuel load and stopover duration. To circumvent these issues, the motivation to depart from a stopover site can be quantified in temporarily caged wild birds by measuring their (nocturnal) migratory restlessness, termed Zugunruhe. Studies doing so have revealed biologically meaningful patterns with regard to stopover duration; in birds captured at stopover and subsequently caged, Zugunruhe during the first night after capture was consistently higher in fat birds than in lean birds [6,14,15]. These studies support the idea that stopover departure decisions are affected by fuel load. A main purpose of stopover, however, is to accumulate fuel for the next flight bout. Therefore, to substantiate the claim that fuel load is an important determinant of stopover duration, it should be shown that Zugunruhe increases when birds are accumulating fuel, and that this increase is larger in individuals that accumulate more fuel. A few studies have shown that changes in fuel load, resulting from fasting–re-feeding treatments, affect the expression of Zugunruhe [16–18]. These studies, however, describe changes in fuel load and Zugunruhe averaged over a set of birds and do not reveal whether the amount of fuel accumulated by individual birds predicts their concurrent change in Zugunruhe.
In this study, we captured northern wheatears (Oenanthe oenanthe), a nocturnal long-distance migrant, during spring stopover. Birds were subsequently contained for three days to measure Zugunruhe and to determine the amount of fuel accumulation. We show that Zugunruhe in the night after capture was positively correlated with fuel load at capture. Moreover, we are the first to show that the within-individual change in Zugunruhe was positively correlated with the amount of fuel accumulated. These results not only provide novel evidence for the idea that stopover duration is affected by current fuel load, but also imply that the speed at which fuel reserves are replenished at stopover is an important determinant of the overall time required for migration.
2. Material and methods
The study was conducted on Helgoland (54°11′ N, 07°55′ E), a small island ca 50 km off the German North Sea coastline. In May 2013, 42 migrating northern wheatears were caught using mealworm-baited spring traps and subsequently caged for three days. Upon capture, CE measured wing length (maximum chord) to the nearest 0.5 mm, and body mass to the nearest 0.1 g. Wing length was used to calculate lean body mass, employing a linear regression based on 220 ‘lean’ northern wheatears caught on Helgoland in previous years: lean body mass (g) = 0.29 g mm−1 × wing length (mm) – 6.85 g (linear regression: n = 220, F1,218 = 95.07, adj-R2 = 0.30, p < 0.0001, after [19]). Fuel load was calculated as: fuel load = (body mass (g) – lean body mass (g))/lean body mass (g). Approximately 1 h after trapping, birds were put in individual cages (40 × 40 × 30 cm) set up in a single indoor room with artificial lighting. Throughout the experiment, the lights in the room went on at 7.00 and off at 21.00 CET, the temperature was held constant at 20°C and birds had ad libitum access to water. Each morning, immediately after the lights went on, each bird was weighed to the nearest 0.1 g, after which it received a food tray with 30 g of mealworms. Food trays were removed at lights out.
Zugunruhe was recorded automatically with motion-sensitive microphones (developed by R. Nagel, Wilhelmshaven, Germany), which were attached to the right wall of each cage. Each time a bird moved, this generated an impulse that was transmitted to a recording device (developed by S. F. Becker, Bremen, Germany). To avoid the recording of occasional non-migratory activity, we set a threshold of three impulses per second before it was recorded as an activity count. Another device (developed by S. F. Becker) created a CSV file summarizing the activity counts over 15-min periods. Zugunruhe was then expressed as the number of 15-min periods in a night during which a bird showed at least five activity counts (following [20]). Statistical analyses were performed using SPSS v. 20.0 (IBM, New York, NY, USA). Variables were normally distributed (Kolmogorov–Smirnov test, all p > 0.13).
3. Results
Fuel load at capture was positively correlated with Zugunruhe in the night following capture (Pearson's r = 0.4, p = 0.009, n = 42; figure 1a), and negatively correlated with the change in fuel load from capture to the third morning in captivity (Pearson's r = −0.51, p = 0.001, n = 42; figure 1b). Hence, fat birds displayed more Zugunruhe and accumulated less fuel than lean birds. The change in fuel load from capture to the third morning in captivity was positively correlated with the change in Zugunruhe from the first to the third night after capture (Pearson's r = 0.47, p = 0.002, n = 42; figure 2). In other words, birds that accumulated more fuel showed a larger increase in Zugunruhe.
Figure 1.
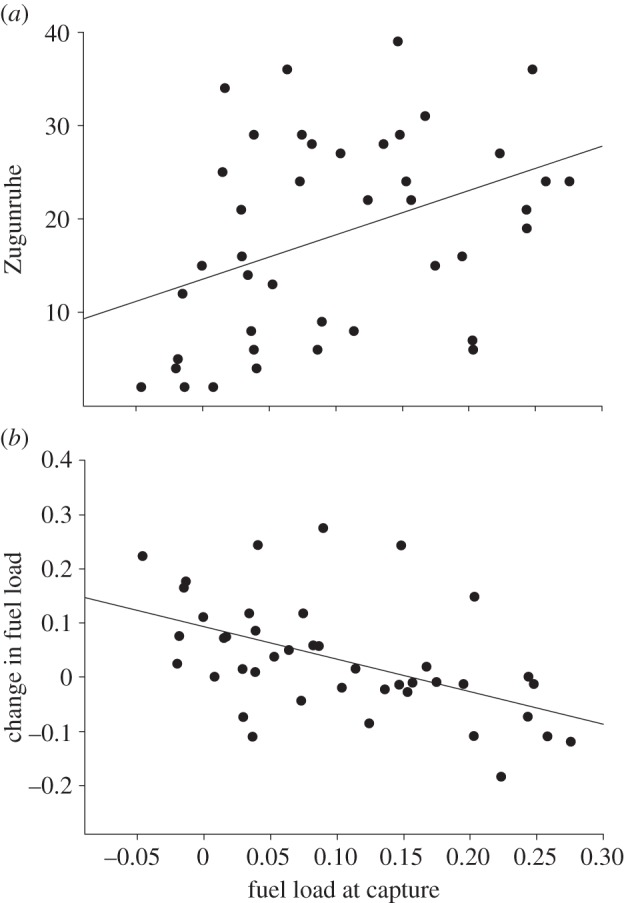
The relationship between fuel load at capture and (a) Zugunruhe in the night following capture, and (b) the change in fuel load from capture to the third morning in captivity (n = 42). Zugunruhe was expressed as the number of 15-min periods in a night during which a bird showed at least five activity counts.
Figure 2.
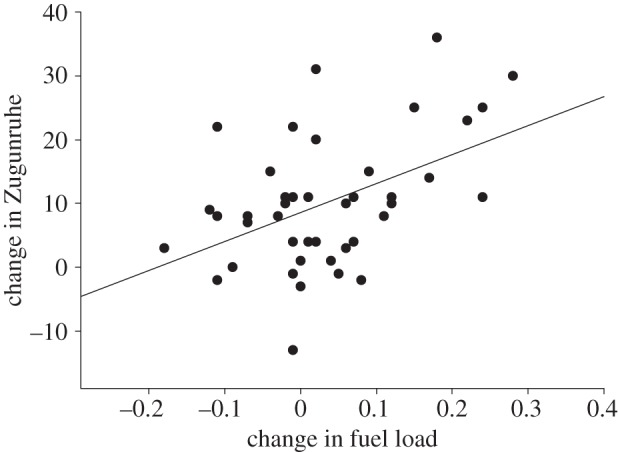
The relationship between the change in fuel load from capture to the third morning in captivity and the change in Zugunruhe from the first to the third night after capture (n = 42). Zugunruhe was expressed as the number of 15-min periods in a night during which a bird showed at least five activity counts.
4. Discussion
We found that birds caught at stopover with large fuel loads showed more Zugunruhe the night after capture, and accumulated less fuel in the subsequent two days in captivity than birds caught with small fuel loads. This indicates that birds with large fuel loads were motivated to continue migration, whereas birds with small fuel loads were more motivated to refuel. We also found that individuals that accumulated large amounts of fuel showed a considerable increase in Zugunruhe, whereas individuals that did not accumulate (or even lost fuel) only marginally increased (or even decreased) their Zugunruhe. Most individuals that did not accumulate fuel already carried large fuel loads at capture, and therefore probably did not need to refuel. Correspondingly, these birds probably already displayed their ‘peak’ amount of Zugunruhe the night after capture, and therefore did not show a clear increase in Zugunruhe in the following nights. Our results not only confirm the earlier finding that Zugunruhe at stopover is higher in fat than in lean birds [6,14,15], but also, and for the first time, show that the amount of fuel accumulated during stopover predicts the within-individual change in Zugunruhe. Our results provide compelling evidence for a direct link between a bird's fuel reserves and its motivation to depart from a stopover site. Moreover, they indicate that the speed at which fuel reserves are replenished can shorten the duration of spring stopovers, which in its turn reduces the overall time required for migration [5].
The physiological mechanism by which migratory birds’ fuel reserves affect the expression of Zugunruhe is currently unclear, but it appears that endocrine signals play a role. Corticosterone may signal migratory readiness as baseline corticosterone level in birds caught at stopover has been shown to be positively correlated with fuel reserves [21,22], as well as with nocturnal activity during the first hour after sunset [23]. Hormones produced by the adipose tissue itself, such as adiponectin and visfatin, may also act to signal body condition of birds in migratory condition and thereby affect departure decisions [24].
In conclusion, our study provides clear evidence in support of the hypothesis that in migratory birds, the likelihood of departure from stopover is affected by their fuel reserves. Furthermore, our study indicates that the speed at which fuel reserves are replenished at stopover is an important determinant of the overall time required for migration.
Acknowledgements
We thank Jochen Dierschke and Klaus Müller for help on Helgoland. Oscar Vedder, Karen Bouwman and two anonymous referees provided useful comments.
Data accessibility
Data are available from the Dryad Digital Repository: http://dx.doi.org/10.5061/dryad.703s1 [25].
Funding statement
All procedures were approved by the Ministry for Agriculture, the Environment and rural Areas, Schleswig-Holstein, Germany.
References
- 1.Currie D, Thompson DBA, Burke T. 2000. Patterns of territory settlement and consequences for breeding success in the northern wheatear Oenanthe oenanthe. Ibis 142, 389–398 (doi:10.1111/j.1474-919X.2000.tb04435.x) [Google Scholar]
- 2.Smith RJ, Moore FR. 2005. Arrival timing and seasonal reproductive performance in a long-distance migratory landbird. Behav. Ecol. Sociobiol. 57, 231–239 (doi:10.1007/s00265-004-0855-9) [Google Scholar]
- 3.Green M, Alerstam T, Clausen P, Drent R, Ebbinge BS. 2002. Dark-bellied brent geese Branta bernicla bernicla, as recorded by satellite telemetry, do not minimize flight distance during spring migration. Ibis 144, 106–121 (doi:10.1046/j.00191019.2001.00017.x) [Google Scholar]
- 4.Schmaljohann H, Fox JW, Bairlein F. 2012. Phenotypic response to environmental cues, orientation and migration costs in songbirds flying halfway around the world. Anim. Behav. 84, 623–640 (doi:10.1016/j.anbehav.2012.06.018) [Google Scholar]
- 5.Alerstam T, Lindström A. 1990. Optimal bird migration: the relative importance of time, energy and safety. In Bird migration (ed. Gwinner E.), pp. 31–351 Berlin, Germany: Springer [Google Scholar]
- 6.Bairlein F. 1985. Body weight and fat deposition of Palaeartic passerine migrants in the central Sahara. Oecologia 66, 141–146 (doi:10.1007/BF00378566) [DOI] [PubMed] [Google Scholar]
- 7.Biebach H, Friedrich W, Heine G. 1986. Interaction of bodymass, fat, foraging and stopover period in trans-Sahara migrating passerine birds. Oecologia 69, 370–379 (doi:10.1007%2FBF00377059) [DOI] [PubMed] [Google Scholar]
- 8.Goymann W, Spina F, Ferri A, Fusani L. 2010. Body fat influences departure from stopover sites in migratory birds: evidence from whole-island telemetry. Biol. Lett. 6, 478–481 (doi:10.1098/rsbl.2009.1028) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Salewski V, Schaub M. 2007. Stopover duration of Paleartic passerine migrants in the western Sahara—independent of fat stores? Ibis 149, 223–236 (doi:10.1111/j.1474-919X.2006.00608.x) [Google Scholar]
- 10.Tsvey A, Bulyuk VN, Kosarev V. 2007. Influence of body condition and weather on departures of first-year European robins, Erithacus rubecula, from an autumn migratory stopover site. Behav. Ecol. Sociobiol. 61, 1665–1674 (doi:10.1007/s00265-007-0397-z) [Google Scholar]
- 11.Jenni L, Schaub M. 2003. Behavioural and physiological reactions to environmental variation in bird migration: a review. In Avian migration (eds Berthold P, Gwinner E, Sonnenschein E.), pp. 155–171 Berlin, Germany: Springer [Google Scholar]
- 12.Schaub M, Jenni L, Bairlein F. 2008. Fuel stores, fuel accumulation, and the decision to depart from a migration stopover site. Behav. Ecol. 19, 657–666 (doi:10.1093/beheco/arn023) [Google Scholar]
- 13.Schaub M, Pradel R, Jenni L, Lebreton JD. 2001. Migrating birds stop over longer than usually thought: an improved capture–recapture analysis. Ecology 82, 852–859 (doi:10.2307/2680203) [Google Scholar]
- 14.Yong W, Moore FR. 1993. Relation between migratory activity and energetic condition among thrushes (Turdinae) following passage across the Gulf of Mexico. Condor 95, 934–943 (doi:10.2307/1369429) [Google Scholar]
- 15.Fusani L, Cardinale M, Carere C, Goymann W. 2009. Stopover decision during migration: physiological conditions predict nocturnal restlessness in wild passerines. Biol. Lett. 5, 302–305 (doi:10.1098/rsbl.2008.0755) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Berthold P. 1976. Über den Einfluß der Fettdeposition auf die Zugunruhe bei der Gartengrasmücke Sylvia borin. Vogelwarte 28, 263–266 [Google Scholar]
- 17.Biebach H. 1985. Sahara stopover in migratory flycatchers: fat and food affect the time program. Experientia 41, 695–697 (doi:10.1007%2FBF02007727) [Google Scholar]
- 18.Fusani L, Gwinner E. 2004. Simulation of migratory flight and stopover affects night levels of melatonin in a nocturnal migrant. Proc. R. Soc. Lond. B 271, 205–211 (doi:10.1098%2Frspb.2003.2561) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Schmaljohann H, Naef-Daenzer B. 2011. Body condition and wind support initiate the shift of migratory direction and timing of nocturnal departure in a songbird. J. Anim. Ecol. 80, 1115–1122 (doi:10.1111/j.1365-2656.2011.01867.x) [DOI] [PubMed] [Google Scholar]
- 20.Maggini I, Bairlein F. 2010. Endogenous rhythms of seasonal migratory body mass changes in different populations of northern wheatears Oenanthe oenanthe. J. Biol. Rhythms 25, 268–276 (doi:10.1177/0748730410373442) [DOI] [PubMed] [Google Scholar]
- 21.Landys-Cianelli MM, Ramenofsky M, Piersma T, Jukema J, Castricum Ringing Group, Wingfield JC 2002. Baseline and stress-induced plasma corticosterone during long-distance migration in the bar-tailed godwit, Limosa lapponica. Physiol. Biochem. Zool. 75, 101–110 (doi:10.1086/338285) [DOI] [PubMed] [Google Scholar]
- 22.Eikenaar C, Fritzsch A, Bairlein F. 2013. Corticosterone and migratory fueling in Northern wheatears facing different barrier crossings. Gen. Comp. Endocrinol. 186, 181–186 (doi:10.1016/j.ygcen.2013.02.042) [DOI] [PubMed] [Google Scholar]
- 23.Lõhmus M, Sandberg R, Holberton RL, Moore FR. 2003. Corticosterone levels in relation to migratory readiness in red-eyed vireos (Vireo olivaceus). Behav. Ecol. Sociobiol. 54, 233–239 (doi:10.1007/s00265-003-0618-z) [Google Scholar]
- 24.Stuber EF, Verpeut J, Horvat-Gordon M, Ramachandran R, Bartell PA. 2013. Differential regulation of adipokines may influence migratory behavior in the white-throated sparrow (Zonotrichia albicollis). PLoS ONE 8, e59097 (doi:10.1371/journal.pone.0059097) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cas E, Jan LS. 2013. Data from: size and accumulation of fuel reserves at stopover predict nocturnal restlessness in a migratory bird. Dryad Digital Repository: http://dx.doi.org/10.5061/dryad.703s1 [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data are available from the Dryad Digital Repository: http://dx.doi.org/10.5061/dryad.703s1 [25].