

Abstract
The phagocyte respiratory burst is part of the innate immune response to pathogen infection and involves the production of reactive oxygen species (ROS). ROS are toxic and function to kill phagocytized microorganisms. In vivo quantification of phagocyte-derived ROS provides information regarding an organism's ability to mount a robust innate immune response. Here we describe a protocol to quantify and compare ROS in whole zebrafish embryos upon chemical induction of the phagocyte respiratory burst. This method makes use of a non-fluorescent compound that becomes fluorescent upon oxidation by ROS. Individual zebrafish embryos are pipetted into the wells of a microplate and incubated in this fluorogenic substrate with or without a chemical inducer of the respiratory burst. Fluorescence in each well is quantified at desired time points using a microplate reader. Fluorescence readings are adjusted to eliminate background fluorescence and then compared using an unpaired t-test. This method allows for comparison of the respiratory burst potential of zebrafish embryos at different developmental stages and in response to experimental manipulations such as protein knockdown, overexpression, or treatment with pharmacological agents. This method can also be used to monitor the respiratory burst response in whole dissected kidneys or cell preparations from kidneys of adult zebrafish and some other fish species. We believe that the relative simplicity and adaptability of this protocol will complement existing protocols and will be of interest to researchers who seek to better understand the innate immune response.
Keywords: Immunology, Issue 79, Phagocytes, Immune System, Zebrafish, Reactive Oxygen Species, Immune System Processes, Host-Pathogen Interactions, Respiratory Burst, Immune System Phenomena, innate immunity, bacteria, virus, infection]
Introduction
The immune system is comprised of two branches: innate and adaptive immunity. Innate immunity is evolutionarily more ancient than adaptive immunity. Invertebrates are presently thought to have only innate immunity, whereas vertebrates possess both the innate and adaptive branches. While adaptive immunity confers specific and long-lasting immunity to certain pathogens, innate immunity is an immediate response to invading bacteria, viruses, and fungi. A crucial aspect of the innate immune response involves the release of cytokines and chemokines, which results in inflammation and recruitment of phagocytes (e.g. macrophages, neutrophils) to engulf and destroy foreign invaders.
Successful innate immune responses involve: (1) recognition of invading microorganisms; (2) induction of the appropriate signaling cascades (e.g. release of cytokines and chemokines); (3) proper development/adequate numbers of phagocytic cells; (4) migration of phagocytes to sites of infection; (5) engulfment of pathogens; and (6) destruction of engulfed microorganisms. A deficiency in any one of these steps could lead to the host being overwhelmed by, and succumbing to, the infection. A robust innate immune response is vital to the health of organisms because it is the first line of defense against pathogens in all plants and animals. In vertebrates, it also potentiates the adaptive immune response1. Therefore, it is critical that we are able to evaluate all aspects of the innate immune response in order to better understand it and to optimize its function.
Many model organisms are used to study innate immunity, ranging from Arabadopsis to C. elegans to Drosophila to mice to cultured human cells. An advantage to using the zebrafish (Danio rerio) model system to study innate immunity is that the zebrafish is a vertebrate, with both innate and adaptive immunity, yet the development of innate and adaptive immunity are temporally segregated. Zebrafish rely solely on innate immunity for protection against infection until adaptive immunity becomes fully functional, which occurs around 4-6 weeks post fertilization2. In addition to tools for genetic manipulation, optical clarity and rapid, external development, innate immunity as the principle mode of defense in zebrafish embryos provides a simplified model in which to study the complexities of the innate immune response in vivo.
Multiple protocols have been developed to assess different facets of the innate immune response in zebrafish embryos. Microarrays and RNAseq have validated that the cytokine profiles elicited by the zebrafish innate immune response are similar to that of humans and have also suggested the involvement of unexpected genes in innate immunity3,4. The transparency of the zebrafish embryo and fluorescent, transgenic strains of pathogens and zebrafish allow for visualization of dynamic host-pathogen interactions in vivo in real time. Transgenic zebrafish embryos expressing GFP under control of the neutrophil-specific myeloperoxidase promoter5,6 or the macrophage-specific mpeg1 promoter7 have made it possible to visualize and quantify phagocyte migration to sites of localized infections8 as well as to visualize phagocytosis and destruction of fluorescently labeled pathogens8,9. Zebrafish embryos are also amenable to the generation of high-throughput assays and chemical screens. Accordingly, high-throughput methods of transcriptome analysis upon infection10 and phagocyte migration to sites of chemically induced injury11 have recently been developed.
Of the techniques listed above, none quantitatively assess the final stage of pathogen destruction by phagocytes. This final stage involves a respiratory burst (i.e. production of ROS and other toxic compounds), which kill the engulfed pathogens. The enzyme NADPH oxidase is a major source of ROS in phagocytic cells. Assembly of the subunits of the NADPH oxidase enzyme results in transfer of electrons to oxygen, generating superoxide anions. Through subsequent enzymatic reactions, superoxide can then be converted into hydrogen peroxide and hypochlorous acid (Figure 1A). It is the respiratory burst of phagocytes that kills pathogens and thus, the quantification of the respiratory burst potential of zebrafish embryos is indicative of overall innate immune health. We developed a fluorescence-based assay to quantify the respiratory burst in groups of individual zebrafish embryos12. This assay utilizes the non-fluorescent, reduced form of a commercially available, cell-permeable dye. This dye, 2',7'-dichlorodihydrofluorescein diacetate (H2DCFDA), is converted into the fluorescent compound, 2',7'-dichlorofluorescein (DCF), upon oxidation. The diverse ROS generated by the phagocyte respiratory burst can oxidize H2DCFDA and generate fluorescence24. The appearance of fluorescence can be used to quantify and compare the respiratory burst response between groups of zebrafish. The protein kinase C agonist phorbol myristate acetate (PMA) is used to chemically induce NADPH oxidase to produce ROS and thus increase fluorescence readings (Figure 1B). Herein, we provide a detailed protocol of a modified and optimized version of this zebrafish embryo respiratory burst assay. This assay can be used to compare the respiratory burst between groups of individual zebrafish embryos over time and/or in response to experimental manipulations (e.g. morpholino-mediated protein knockdown). The use of this method, in conjunction with other zebrafish innate immunity assays, will provide a more complete picture of the complex and critical innate immune response.
Protocol
1. Zebrafish Care and Maintenance
Husbandry: Mass spawn adult zebrafish as previously described13. Collect spawned embryos as previously described14.
- Microinjection (if desired): Microinject 1-4 cell stage zebrafish embryos with morpholino oligonucleotides to knockdown gene products or mRNA to overexpress gene products as previously described15.
- Maintain an adequate pool of mock injected controls (at least 48 living, mock injected control fish and 48 living, experimentally manipulated fish are needed to fill a 96 well microplate).
- Maintain embryos: Grow embryos in deep petri dishes at 28 °C in egg water (60 μg/ml Instant Ocean Sea Salt in distilled water-autoclaved) until the desired developmental stage (a respiratory burst response is not detectable using this protocol in zebrafish embryos younger than 2 days post fertilization12). Note: It has been observed that preventing pigmentation in zebrafish embryos through either the use of 1-phenyl 2-thiourea (PTU) or golden/slc24a5 mutant zebrafish does not significantly alter the induction of fluorescence by PMA (unpublished data, embryos tested at 48, 72, and 96 hpf).
- Remove dead embryos daily with a plastic transfer pipette.
- Carefully decant old egg water and replenish with new egg water daily.
Dechorionate embryos: On the day of the experiment, dechorionate embryos (if embryos are still in their chorions) as previously described using two fine forceps14 (this respiratory burst assay can also be performed on dissected kidneys from adult zebrafish12 and detailed protocols for kidney dissection from adult zebrafish have been previously described16,17).
2. Solution Preparation
- Prepare a stock solution of H2DCFDA: Weigh out 1 mg of H2DCFDA.
- Dissolve 1 mg of H2DCFDA in 1 ml of dimethyl sulfoxide (DMSO) to make a 1 mg/ml stock solution.
- Make 22 μl aliquots of this stock solution in 1.7 ml microcentrifuge tubes.
- Wrap the aliquots of H2DCFDA stock solution in aluminum foil and keep in the dark whenever possible because H2DCFDA is light sensitive.
- Store H2DCFDA aliquots at -20 °C for up to 3 months.
- CAUTION - Prepare a stock solution of phorbol myristate acetate (PMA): Weigh out 1 mg of PMA using proper personal protective equipment (i.e. gloves, goggles and mask).
- Dissolve PMA in 1 ml of DMSO to make a 1 mg/ml stock solution.
- Make 11 μl aliquots in 1.7 ml microcentrifuge tubes.
- Store aliquots of PMA stock solution at -80 °C for up to 3 months.
Prepare a working solution of H2DCFDA: On the day of the experiment, make a H2DCFDA working solution by adding 1 part H2DCFDA stock solution to 1 part DMSO (500 μg/ml H2DCFDA final concentration). For example, add 20 μl of H2DCFDA stock solution and 20 μl DMSO to a foil wrapped 1.7 ml microcentrifuge tube.
Prepare a working solution of PMA: On the day of the experiment, make a PMA working solution by adding 1 part PMA stock solution to 49 parts nuclease free water (20 μg/ml PMA final concentration). For example, dilute 10 μl PMA stock solution in 490 μl nuclease free water in a 1.7 ml microcentrifuge tube.
- Prepare a dosing solution of H2DCFDA: On the day of the experiment, make a H2DCFDA dosing solution with a final concentration of 1 μg/ml H2DCFDA in egg water. The prepared volumes of the dosing solutions can be modified as desired, but ensure that the final concentrations of the reagents are maintained. For kidneys, instead of egg water, use the same volume of Dulbecco's modified Eagle's medium/F-12 (50% DMEM, 50% F-12, without phenol red). To make 5 ml of H2DCFDA dosing solution, use a 5 ml serological pipette to transfer 5 ml of egg water (or DMEM/F-12) into a 15 ml conical centrifuge tube wrapped in foil and labeled 'H'.
- Remove 10 μl of egg water (or DMEM/F-12) from the 15 ml conical tube labeled 'H' and discard.
- Add 10 μl of H2DCFDA working solution into the 15 ml conical tube labeled 'H' and vortex to mix (this volume is enough for 48 embryo or kidney samples (half of a full 96 well microplate) and these wells will provide measurements of the level of background fluorescence).
- Prepare a dosing solution of H2DCFDA + PMA: On the day of the experiment, make a H2DCFDA + PMA dosing solution with final concentrations of: H2DCFDA - 1 μg/ml and PMA - 400 ng/ml. To make 5 ml of H2DCFDA + PMA dosing solution, use a 5 ml serological pipette to transfer 5 ml of egg water (or DMEM/F-12) into a new 15 ml conical tube labeled 'H + P'.
- Remove 110 μl of egg water (or DMEM/F-12) from the 15 ml conical tube labeled 'H + P' and discard.
- Add 10 μl of H2DCFDA working solution, then add 100 μl of PMA working solution into the 15 ml conical tube labeled 'H + P' and vortex to mix (this volume is enough for 48 samples or the other half of a full 96 well microplate).
Keep the dosing solutions on ice.
3. Microplate Reader Programming
- Prepare instrument: Power on the microplate reader.
- Warm up the light source.
- Set up a program to read fluorescence: e.g. Excitation: 485 nm; Emission: 528 nm; Optics Position: top 510 nm; Sensitivity: 65, with a 5 sec shaking step prior to the read.
4. 96 Well Microplate Set Up (see Figure 2)
Gather supplies: Obtain dishes with dechorionated embryos, black 96 well microplate, p200 pipettor and tips, ice bucket with H2DCFDA and H2DCFDA + PMA dosing solutions, multichannel p200 pipettor, two sterile reservoirs, aluminum foil, and scissors.
- Transfer one embryo into each well of a 96 well microplate: Use scissors to cut a pipette tip such that embryos or kidneys fit through the opening.
- Set a p200 pipettor to 100 μl and transfer one embryo along with egg water (or DMEM/F-12) into as many of the wells of a black 96 well microplate as desired (it is not necessary to change the pipette tip for each different embryo sample within an experimental condition, but it may be necessary to change tips between experimental conditions). Be sure to avoid transferring residual chorions, as these tend to skew the data collected. For the transfer of kidneys, a larger volume pipettor (set to 100 μl) can be used and pipette tips can be cut to obtain a larger bore size, if necessary. It may be necessary to incorporate wells without embryo or kidney samples, but with the dosing solutions to control for some experimental manipulations.
- Add dosing solutions: Pour H2DCFDA dosing solution into a sterile 25 ml reservoir.
- Use a multichannel p200 pipettor and eight tips to simultaneously pipette 100 μl of H2DCFDA dosing solution into one column on the 96 well microplate (500 ng/ml final concentration of H2DCFDA).
- Repeat this (changing tips is not necessary) for as many columns as desired (usually six columns or 48 wells if filling an entire 96 well microplate). Add this solution to half of the control embryo samples and half of the experimentally manipulated embryo samples (an example 96 well microplate set up is shown in Figure 2, these wells (colored orange) will provide background fluorescence data in samples not induced with PMA).
- Pour the H2DCFDA + PMA dosing solution into a new, sterile 25 ml reservoir.
- Use a multichannel p200 pipettor and eight tips to simultaneously pipette 100 μl of H2DCFDA + PMA dosing solution into the remaining columns (colored red in Figure 2) of the 96 well microplate (changing tips is not necessary, final concentrations of H2DCFDA- 500 ng/ml and PMA- 200 ng/ml).
Cover the microplate with aluminum foil.
Shake the microplate for approximately 20 sec at 150 rpm to homogenize the solutions in each well.
Incubate the microplate at 28 °C when it is not being read.
5. Fluorescence Quantification
- Read the microplate at time = 0 hr after the addition of PMA using the parameters described in step 3.1.2.
- Continue taking measurements every few minutes for the desired time interval or incubate the foil wrapped microplate at 28 °C until a later time point and then take an endpoint measurement at a desired time (e.g. 4 hr after the addition of PMA).
Use a plastic transfer pipette to retrieve the embryos from the wells.
Euthanize the embryos according to your animal care and usage protocol, e.g. immersion in tricaine MS222.
Dispose of the microplate and other disposable materials in the biohazardous waste container.
6. Data Analysis
Decide on the time point at which you would like to compare fluorescence values (e.g. 4 hr after the addition of PMA, Table 1).
Subtract the average un-induced control group's fluorescence value from the individual PMA-induced control group's fluorescence values.
Repeat this for the experimental group with and without PMA.
Store these normalized fluorescence values in two columns, the control + PMA group and the experimental + PMA group (Table 2).
Calculate the means and standard deviations for the normalized fluorescence values of the control + PMA group and the experimental + PMA group.
Compare the normalized fluorescence values using an unpaired t-test to determine statistical significance (Table 2).
Graph the means of the control + PMA group and the experimental + PMA group with error bars reflecting the appropriate standard deviations.
Record the level of significance on the graph and in the figure legend (Figure 2).
Representative Results
Here, we provide data comparing the respiratory burst response in zebrafish embryos (wild-type, AB background) at 48 and 72 hours post fertilization (hpf). The 48 hpf embryos acted as our control group and the 72 hpf embryos as our experimental group. The sample size used was 24 un-induced embryos and 24 PMA-induced embryos per developmental stage. Raw fluorescence readings (in Relative Fluorescence Units (RFU)) were obtained by reading the microplate 4 hours after the addition of PMA. Raw fluorescence values are provided in Table 1. Raw fluorescence values were consistently higher in the 48 hpf un-induced embryos than the 72 hpf un-induced embryos, with the exception of one 72 hpf embryo (means: 48 hpf = 758 RFU, 72 hpf = 230 RFU). The variability within these two samples was approximately equal (standard deviations: 48 hpf = ±549 RFU, 72 hpf = ±513 RFU), but the 72 hpf group varied more with respect to the mean (standard deviation as a percentage of the mean: 48 hpf = 72%, 72 hpf = 223%). In the PMA-induced groups, raw fluorescence values were mostly higher in the 72 hpf population (means: 48 hpf PMA = 3798 RFU, 72 hpf PMA = 5825 RFU). There was more variability in the 72 hpf PMA-induced group than the 48 hpf PMA-induced group (standard deviations: 48 hpf PMA = 831 RFU, 72 hpf PMA = 1365 RFU), but the variability with respect to the mean was approximately equal (48 hpf PMA = 22%, 72 hpf PMA = 23%).
To account for variability in background levels of fluorescence, normalized fluorescence values were calculated (PMA-induced value minus the mean of the un-induced values of the appropriate developmental stage). These normalized fluorescence values are provided in Table 2, along with the means, standard deviations, and p-value for the 48 and 72 hpf groups. A graph of this data is provided in Figure 2. The normalized fluorescence values are consistently higher in the 72 hpf group than the 48 hpf group (means: 48 hpf = 3040 RFU, 72 hpf = 5596 RFU) and the variability is similar with respect to the mean (48 hpf = 27%, 72 hpf = 24%). This respiratory burst assay revealed that 72 hpf zebrafish embryos are able to produce more ROS and therefore mount a more robust respiratory burst response than 48 hpf embryos after induction with PMA. This finding may be due to an increased number of cells, including an increased number of granulocytes18, in 72 hpf embryos compared to 48 hpf embryos. An unpaired t-test to statistically compare the data confirmed that the respiratory burst response is significantly different in 72 hpf embryos than 48 hpf embryos (*p=2 X 10-9).
In a more general sense, at the time point t = 0 hr after the addition of PMA, the raw fluorescence values should not be statistically different between the PMA-induced and the un-induced groups. The raw fluorescence values will increase over time in both the un-induced and induced samples, with a much larger increase occurring in the PMA-induced embryos12. The increase in the raw fluorescence values in the PMA-induced group, compared to the un-induced group, will become statistically significant at approximately 1-2 hours after the addition of PMA (depending on the developmental stage of the zebrafish embryos, with older embryos mounting a quicker respiratory burst response than younger embryos). Normalized fluorescence values (induced minus un-induced) should increase with the increasing developmental stage of zebrafish embryos, with the largest difference occurring between 48 and 72 hr post fertilization and a smaller difference occurring between 72 and 96 hr post fertilization. Raw fluorescence numbers should be 5 to 60-fold higher in the PMA-induced group relative to the un-induced group, depending on the developmental stage of the zebrafish embryos (between 48 and 96 hr post fertilization). Variation will occur between individual zebrafish embryo samples, thus, it is recommended that approximately 24 embryo samples per treatment be used.
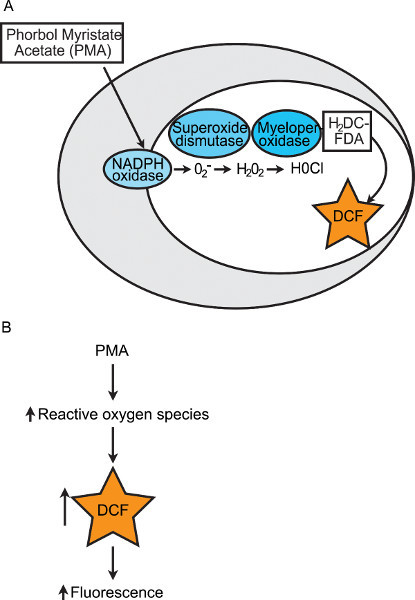 Figure 1. Diagram of chemical induction of the respiratory burst within a phagocyte. (A) In this respiratory burst assay, PMA is used to induce production of superoxide anions by NADPH oxidase. Superoxide is converted into oxygen and hydrogen peroxide by superoxide dismutase. Hydrogen peroxide and chloride anions are converted into hypochlorous acid by myeloperoxidase. The non-fluorescent dye, H2DCFDA, is converted into the fluorescent compound, DCF, when oxidized by many different ROS. (B) Diagram of how quantification of fluorescence is a read out of the respiratory burst response.
Figure 1. Diagram of chemical induction of the respiratory burst within a phagocyte. (A) In this respiratory burst assay, PMA is used to induce production of superoxide anions by NADPH oxidase. Superoxide is converted into oxygen and hydrogen peroxide by superoxide dismutase. Hydrogen peroxide and chloride anions are converted into hypochlorous acid by myeloperoxidase. The non-fluorescent dye, H2DCFDA, is converted into the fluorescent compound, DCF, when oxidized by many different ROS. (B) Diagram of how quantification of fluorescence is a read out of the respiratory burst response.
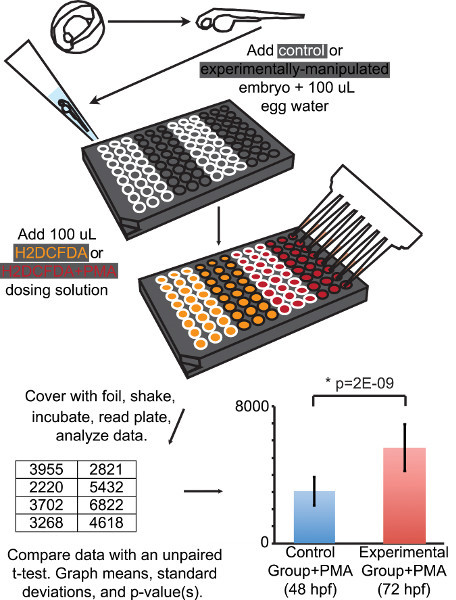 Figure 2. Schematic of the experimental design for the respiratory burst assay. Individual dechorionated zebrafish embryos are transferred into the wells of a 96 well microplate. The wells outlined in white contain control embryos and the wells outlined in black contain embryos from the experimental group. H2DCFDA or H2DCFDA+PMA dosing solutions are added to the appropriate wells (colored orange or red, respectively). Fluorescence is quantified using a microplate reader. Respiratory burst assay data is then analyzed, compared, and presented. The table in this figure is a subset of the normalized fluorescence values displayed in Table 2. The graph in this figure includes the means, standard deviations and p-value for the entire data set presented in this manuscript, which is also displayed in Table 2. * p=2 X 10--9.
Figure 2. Schematic of the experimental design for the respiratory burst assay. Individual dechorionated zebrafish embryos are transferred into the wells of a 96 well microplate. The wells outlined in white contain control embryos and the wells outlined in black contain embryos from the experimental group. H2DCFDA or H2DCFDA+PMA dosing solutions are added to the appropriate wells (colored orange or red, respectively). Fluorescence is quantified using a microplate reader. Respiratory burst assay data is then analyzed, compared, and presented. The table in this figure is a subset of the normalized fluorescence values displayed in Table 2. The graph in this figure includes the means, standard deviations and p-value for the entire data set presented in this manuscript, which is also displayed in Table 2. * p=2 X 10--9.
| 48 hpf | 72 hpf | |
| 3955 | 2821 | |
| 2220 | 5432 | |
| 3702 | 6822 | |
| 3268 | 4618 | |
| 2411 | 5553 | |
| 2984 | 6535 | |
| 2970 | 5448 | |
| 1767 | 3042 | |
| 2110 | 4638 | |
| 4010 | 5914 | |
| 2975 | 4199 | |
| 3220 | 8259 | |
| 4266 | 5391 | |
| 2212 | 5786 | |
| 2953 | 7408 | |
| 3474 | 5893 | |
| 4386 | 4650 | |
| 1240 | 4405 | |
| 2226 | 6442 | |
| 2553 | 5924 | |
| 2785 | 5274 | |
| 3870 | 5728 | |
| 3879 | 5601 | |
| 3530 | 8505 | |
| Mean | 3040 | 5596 |
| Stdev | 831 | 1365 |
| P-value | 2E-09 |
Table 1. Raw fluorescence values from un-induced (orange background) and induced (red background) 48 or 72 hr post fertilization (hpf) zebrafish embryos (white or black numbers, respectively) at 4 hr after the addition of PMA. The mean is calculated for the un-induced samples.
| 48 hpf embryos | 72 hpf embryos | ||||
| 408 | 326 | 358 | 92 | 83 | 208 |
| 276 | 381 | 1124 | 473 | 106 | 86 |
| 518 | 180 | 1085 | 110 | 93 | 109 |
| 1196 | 232 | 380 | 143 | 152 | 81 |
| 1416 | 339 | 735 | 123 | 85 | 81 |
| 489 | 390 | 347 | 110 | 98 | 118 |
| 2139 | 1183 | 1015 | 95 | 97 | 2609 |
| 1660 | 385 | 1630 | 111 | 115 | 136 |
| Mean | 758 | 230 | |||
| 48 hpf embroys + PMA | 72 hpf embryos + PMA | ||||
| 4713 | 2868 | 5144 | 3051 | 4868 | 4880 |
| 2978 | 4768 | 1998 | 5662 | 6144 | 4635 |
| 4460 | 3733 | 2984 | 7052 | 4429 | 6672 |
| 4026 | 3978 | 3311 | 4848 | 8489 | 6154 |
| 3169 | 5024 | 3543 | 5783 | 5621 | 5504 |
| 3742 | 2970 | 4628 | 6765 | 6016 | 5958 |
| 3728 | 3711 | 4637 | 5678 | 7638 | 5831 |
| 2525 | 4232 | 4288 | 3272 | 6123 | 8735 |
Table 2. Normalization and statistical comparison of the data. To obtain normalized fluorescence values, calculate the difference between each PMA-induced fluorescence value and the appropriate un-induced mean value. Statistically compare these two sets of data using an unpaired t-test. Calculate the means and standard deviations. Display the means, standard deviations, and p-value(s) in graphical form (as in Figure 2).
Discussion
The primary function of phagocytes is to detect, engulf, and destroy pathogens. The ability of phagocytes to produce an adequate respiratory burst is critical for this function. Thus, quantification of the respiratory burst response is one method to allow comparison of general innate immune health and function between groups of individuals and/or in response to experimental manipulations. Here, we describe a protocol to induce, quantify, and compare the respiratory burst response between groups of individual zebrafish embryos. In short, PMA results in the production of ROS by the enzyme NADPH oxidase. These ROS act on a non-fluorescent dye and oxidize it to form a fluorescent compound. A microplate reader is used to detect the relative fluorescence in each well. Fluorescence generated upon PMA induction is indicative of the respiratory burst potential and general innate immune health of zebrafish embryos. Comparison of the normalized level of fluorescence between PMA induced groups using an unpaired t-test can be used to determine whether there is a significant difference in the respiratory burst potential between groups of zebrafish. Experimental manipulations resulting in significant differences in the respiratory burst response will provide insights into mechanisms of innate immunity, e.g. gene products necessary for the phagocyte respiratory burst, drugs that potentiate or antagonize the respiratory burst response.
Protocols to assay different facets of the innate immune response in the zebrafish model have been developed (see Introduction). Relatively few of these techniques measure the production of ROS. The technique detailed in this article quantifies production of ROS in zebrafish embryos upon stimulation of an innate immune response with PMA. This technique is similar to respiratory burst assays performed on isolated whole blood samples, in which PMA is also used to induce a respiratory burst response, and then the ROS generated are measured using a fluorogenic substrate (e.g. Phagoburst, Orpegen Pharma). The method described here can be used with whole zebrafish embryos, as well as dissected kidneys from adult zebrafish13, as the zebrafish anterior kidney is analogous to human bone marrow, which is the site of adult hematopoiesis19. The method described here is also versatile enough to be used with other fish species and cell systems, such as cell preparations from fathead minnow kidneys20.
An alternative method to detect ROS in whole zebrafish embryos makes use of an in vivo hydrogen peroxide sensor. Messenger RNA for a redox-sensitive fluorescent protein is injected into zebrafish embryos and the fluorescence ratio is monitored over time following tail fin wounding21. This genetically encoded redox sensor permits visualization of the temporal and spatial dynamics of hydrogen peroxide production in vivo. Application of this method revealed the surprising result that the hydrogen peroxide produced initially preceded arrival of the first innate immune cells, suggesting that hydrogen peroxide released by wounded epithelial cells acts as a signal to recruit phagocytes21. This technique, while powerful and informative, involves time consuming and technically difficult embryological and microscopy procedures. This redox sensor is also specific for hydrogen peroxide, which is only one of the many ROS functioning during the innate immune response.
In comparison to the approach discussed above, our method also measures in vivo ROS, and yet is technically less demanding and uses a chemical to induce an innate immune response rather than a physical wound. In contrast to the above method, which specifically measures hydrogen peroxide production, our procedure measures the total level of ROS. An advantage to detecting a single ROS is that roles for that compound alone in innate immunity can be elucidated. Similar to the method discussed above, our approach also shares the inherent advantages and disadvantages of being performed in the whole animal. The interactions between phagocytes and their environment, which are eliminated when respiratory burst assays are performed on isolated blood samples, are preserved in these whole animal assays. However, use of the whole animal likely results in detection of ROS from non-phagocyte sources (e.g. mitochondrially-derived ROS, nonphagocyte NADPH oxidase). In an attempt to address the specificity of our method for detecting phagocyte-derived ROS, we referred to a study in which this respiratory burst assay was performed in zebrafish embryos lacking phagocyte-specific NADPH oxidase9. Phagocyte-specific NADPH oxidase was inhibited via morpholino-mediated knock down of p47phox or p91phox protein. In zebrafish embryos, at the developmental stage assayed, the expression of these NADPH oxidase subunits is restricted to a subset of blood cells, likely macrophages22,23. The respiratory burst potential of embryos lacking phagocyte-specific NADPH oxidase was approximately one third that of controls9 (personal communication, Dr. Robert Wheeler). This result suggests that while our respiratory burst assay does detect ROS from sources other than phagocytes, the majority of the fluorescence detected can be attributed to the respiratory burst via phagocyte NADPH oxidase. Further specificity of this assay was demonstrated using bis-indolylmaleimide I (BisI), a pharmacological inhibitor of protein kinase C, to prevent the action of PMA, a protein kinase C agonist, on NADPH oxidase. Pre-treatment of PMA-induced zebrafish kidneys or embryos with BisI resulted in fluorescence levels similar to un-induced controls12. An ideal respiratory burst assay would be an in vivo high-throughput assay where phagocyte-specific ROS could be both quantified and visualized over time. Until this is possible, the rapid nature and technical ease of the method described here provides a useful alternative.
Disclosures
The authors have nothing to disclose.
Acknowledgments
The authors would like to acknowledge past and present members of the Kim laboratory, Mark Nilan for zebrafish care and maintenance, Dr. Robert Wheeler for helpful discussions and data sharing, and NIH grants 3RO1GM087308-02S1 and 1P20RR024475-01A2 and the Maine Agricultural and Forest Experiment Station (Publication Number 3303) for funding.
References
- Medzhitov R, Janeway CA., Jr Innate Immunity: Impact on the Adaptive Immune Response. Current Opinion in Immunology. 1997;9:4–9. doi: 10.1016/s0952-7915(97)80152-5. [DOI] [PubMed] [Google Scholar]
- Lam SH, Chua HL, et al. Development and Maturation of the Immune System in Zebrafish, Danio rerio: A Gene expression Profiling. In Situ Hybridization and Immunological. 2004;28:9–28. doi: 10.1016/s0145-305x(03)00103-4. [DOI] [PubMed] [Google Scholar]
- Stockhammer OW, Zakrzewska A, et al. Transcriptome Profiling and Functional Analyses of the Zebrafish Embryonic Innate Immune Response to Salmonella Infection. J Immunol. 9. 2009;9:5641–5653. doi: 10.4049/jimmunol.0900082. [DOI] [PubMed] [Google Scholar]
- Ordas A, Hegedus Z, et al. Deep Sequencing of the Innate Immune Transcriptomic Response of Zebrafish Embryos to Salmonella Infection. Fish & Shellfish Immunology. 2011;31:716–724. doi: 10.1016/j.fsi.2010.08.022. [DOI] [PubMed] [Google Scholar]
- Renshaw SA, Loynes CA, et al. A Transgenic Zebrafish Model of Neutrophilic Inflammation. Blood. 2006;13:3976–3978. doi: 10.1182/blood-2006-05-024075. [DOI] [PubMed] [Google Scholar]
- Mathias JR, Perrin BJ, et al. Resolution of Inflammation by Retrograde Chemotaxis of Neutrophils in Transgenic Zebrafish. J. Leukoc. Biol. 2006;6:1281–1288. doi: 10.1189/jlb.0506346. [DOI] [PubMed] [Google Scholar]
- Ellett F, Pase L, et al. mpeg1 Promoter Transgenes Direct Macrophage-Lineage Expression in Zebrafish. Blood. 2011;4:56–56. doi: 10.1182/blood-2010-10-314120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phennicie RT, Sullivan MJ, et al. Specific Resistance to Pseudomonas aeruginosa Infection in Zebrafish is Mediated by the Cystic Fibrosis Transmembrane Conductance Regulator. Infect. Immun. 2010;11:4542. doi: 10.1128/IAI.00302-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brothers KM, Newman ZR, et al. Live Imaging of Disseminated Candidiasis in Zebrafish Reveals Role of Phagocyte Oxidase in Limiting Filamentous Growth. Eukaryotic Cell. 2011;7:932–944. doi: 10.1128/EC.05005-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rotman J, van Gils W, et al. Rapid Screening of Innate Immune Gene Expression in Zebrafish using Reverse Transcription - Multiplex Ligation-Dependent Probe Amplification. BMC Research Notes. 2011;4 doi: 10.1186/1756-0500-4-196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- d'Alencon CA, Pena OA, et al. A High-Throughput Chemically Induced Inflammation Assay in Zebrafish. BMC Biology. 2010;8:151. doi: 10.1186/1741-7007-8-151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hermann AC, Millard PJ, et al. Development of a Respiratory Burst Assay using Zebrafish Kidneys and Embryos. Journal of Immunological Methods. 2004;292:119–129. doi: 10.1016/j.jim.2004.06.016. [DOI] [PubMed] [Google Scholar]
- Avdesh A, Chen M, et al. Regular Care and Maintenance of a Zebrafish (Danio rerio) Laboratory: An Introduction. J. Vis. Exp. 2012. p. e4196. [DOI] [PMC free article] [PubMed]
- Brothers KM, Wheeler RT. Non-invasive Imaging of Disseminated Candidiasis in Zebrafish Larvae. J. Vis. Exp. 2012. p. e4051. [DOI] [PMC free article] [PubMed]
- Yuan S, Sun Z. Microinjection of mRNA and Morpholino Antisense Oligonucleotides in Zebrafish Embryos. J. Vis. Exp. 2009. p. e1113. [DOI] [PMC free article] [PubMed]
- Gerlach GF, Schrader LN, et al. Dissection of the Adult Zebrafish Kidney. J. Vis. Exp. 2011. p. e2839. [DOI] [PMC free article] [PubMed]
- Gupta T, Mullins MC. Dissection of Organs from the Adult Zebrafish. J. Vis. Exp. 2010. p. e1717. [DOI] [PMC free article] [PubMed]
- Le Guyader D, Redd MJ, et al. Origins and Unconventional Behavior of Neutrophils in Developing Zebrafish. Blood. 2008;111:132–141. doi: 10.1182/blood-2007-06-095398. [DOI] [PubMed] [Google Scholar]
- Davidson AJ, Zon LI. The 'Definitive' (and 'Primitive') Guide to Zebrafish Hematopoiesis. Oncogene. 2004;23:7233–7246. doi: 10.1038/sj.onc.1207943. [DOI] [PubMed] [Google Scholar]
- Jovanovic B, Goetz FW, et al. Immunological Stimuli Change Expression of Genes and Neutrophil Function in Fathead Minnow Pimephales promelas Rafinesque. Journal of Fish Biology. 2011;78:1054–1072. doi: 10.1111/j.1095-8649.2011.02919.x. [DOI] [PubMed] [Google Scholar]
- Niethammer P, Grabher C, et al. A Tissue-Scale Gradient of Hydrogen Peroxide Mediates Rapid Wound Detection in Zebrafish. Nature. 2009;459:996–1000. doi: 10.1038/nature08119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thisse B, Pflumio S, et al. Expression of the zebrafish genome during embryogenesis. (NIH R01 RR15402) ZFIN Direct Data Submission. 2001.
- Thisse B, Thisse C. Fast Release Clones: A High Throughput Expression Analysis. ZFIN Direct Data Submission. 2004.
- Table 18.4. The Molecular Probes Handbook. 11th