

Abstract
Touch sensation is critical for our social and environmental interactions. In mammals, most discriminative light touch sensation is mediated by the Aβ low-threshold mechanoreceptors. Cell bodies of Aβ low-threshold mechanoreceptors are located in the dorsal root ganglia and trigeminal ganglia, which extend a central projection innervating the spinal cord and brain stem and a peripheral projection innervating the specialized mechanosensory end organs. These specialized mechanosensory end organs include Meissner’s corpuscles, Pacinian corpuscles, lanceolate endings, Merkel cells, and Ruffini corpuscles. The morphologies and physiological properties of these mechanosensory end organs and their innervating neurons have been investigated for over a century. In addition, recent advances in mouse genetics have enabled the identification of molecular mechanisms underlying the development of Aβ low-threshold mechanoreceptors, which highlight the crucial roles of neurotrophic factor signaling and transcription factor activity in this process. Here, we will review the anatomy, physiological properties, and development of mammalian low-threshold Aβ mechanoreceptors.
Keywords: mechanoreceptor, Meissner’s corpuscle, Pacinian corpuscle, lanceolate ending, Merkel cell, Ruffini corpuscle, dorsal root ganglion
1. Introduction
Light touch sensation is critical for our social interaction and daily lives. It allows for the detection of diverse stimuli, like a breeze, a kiss, a hug, the texture of fabric, or a shape, and is required for complex tasks, such as using a tool or reading Braille. These stimuli cause vibration, indentation or stretching of the skin, the movement of hair follicles, or some other physical change in the skin, which activate mechanosensory nerve fibers or the specialized mechanosensory end organs in the skin. There are several different types of mammalian low-threshold mechanosensory neurons (mechanoreceptors), including Aβ, Aδ, and C (distinguished according to their transduction velocity), which mediate various form of light touch sensation. For this review, we will focus on Aβ low-threshold mechanoreceptors, which are the main type of primary sensory neurons that mediate discriminative touch and tactile perception in mammals. Cell bodies of Aβ low-threshold mechanoreceptors are located in the trigeminal (TGs) and dorsal root ganglia (DRGs). Each neuron grows a single axon that bifurcates shortly after projecting from the cell body, with the peripheral axon innervating mechanosensory end organs and the central projection innervating the spinal cord and brain stem.
The morphologies and structures of Aβ low-threshold mechanosensory end organs have been extensively examined since their first discovery in the 1800s. Based on morphologies of these end organs, Aβ low-threshold mechanoreceptors are classified into several different subtypes, including Meissner’s corpuscles, Pacinian corpuscles, lanceolate endings, Merkel cells, and Ruffini corpuscles (Iggo and Andres, 1982; Albrecht, 2008). Physiological properties of Aβ low-threshold mechanoreceptors started to be characterized around the middle of 20th century. According to their rates of adaptation to sustained mechanical stimuli, Aβ low-threshold mechanoreceptors are classified as either rapidly adapting (RA) or slowly adapting (SA) (Mountcastle, 1957; Iggo, 1985). Interestingly, their end organ morphologies and physiological properties are very well correlated. Meissner’s corpuscles, lanceolate endings, and Pacinian corpuscles are the RA mechanoreceptors (Iggo and Ogawa, 1977; Iggo, 1985), whereas Merkel cells and Ruffini corpuscles are the SA mechanoreceptors (Iggo and Muir, 1969; Burgess, 1973; Paré et al., 2002) (Fig. 1). In the past two decades, advances in mouse genetic techniques have enabled the dissection of key molecules, mainly neurotrophic factors and transcription factors, involved in controlling the specification and development of different subtypes of mammalian Aβ low-threshold mechanoreceptors.
Figure 1.
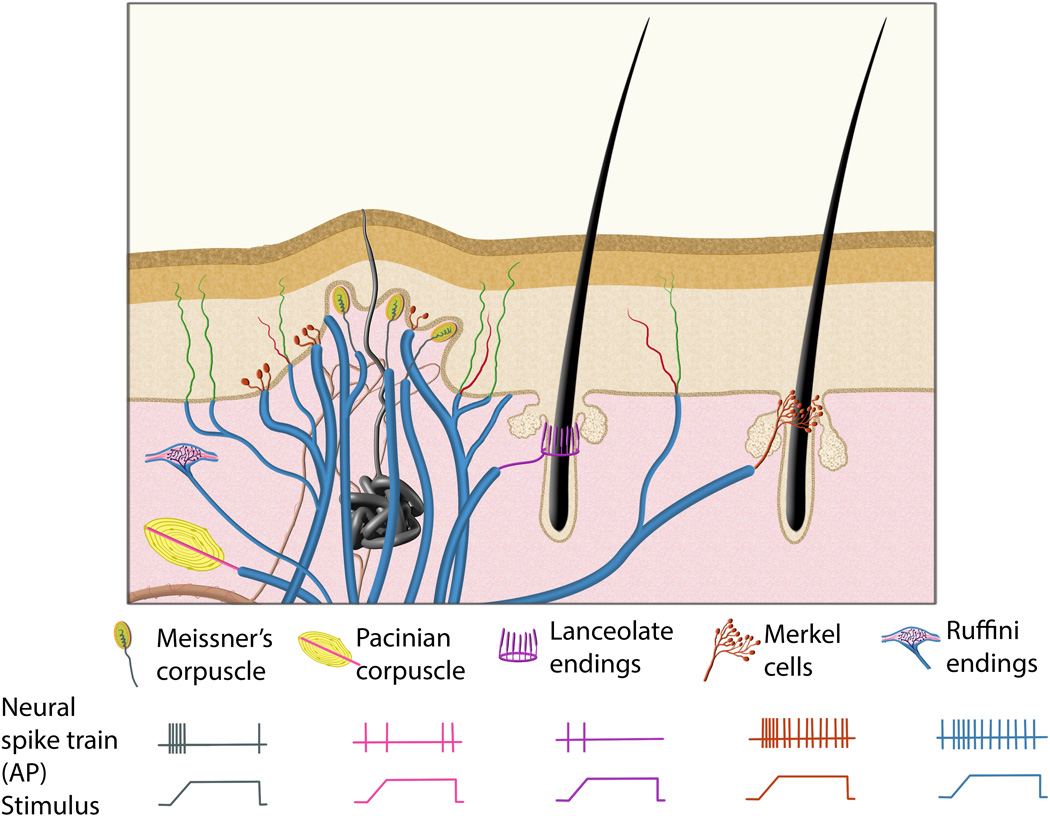
Illustration to demonstrate the morphologies and physiological properties of mammalian Aβ low-threshold mechanoreceptors. In glabrous skin (left side of the illustration), Meissner’s corpuscles are located in the dermal papillae of the dermis, Merkel cells are located in the basal epidermis, Ruffini corpuscles are located in the dermis, and Pacinian corpuscles are located in the dermis, deeper than the other mechanosensory end organs. In hairy skin (right side of the illustration), hair follicles are surrounded by lanceolate endings and Merkel cells. The bottom panel shows the neural activity of different types of Aβ low-threshold mechanoreceptors in response to a sustained stimulus. Meissner’s corpuscle, Pacinian corpuscle and lanceolate ending mechanoreceptors display rapidly adapting mechanosensitive physiological properties, while Merkel cell and Ruffini corpuscle mechanoreceptors display slowly adapting mechanosensitive properties, which fire action potentials (APs) throughout the duration of the stimulus.
2. Meissner’s corpuscles
2.1 Anatomical location and morphology
Meissner’s corpuscles are present in the glabrous (hairless) skin, including palms of the hand, soles of feet, and lips. They are located within the dermal papillae, which are conical protrusions of the dermis into the epidermis, with their long axis perpendicular to the surface of the skin (Munger and Ide, 1988). Meissner’s corpuscles were first described in the fingers and palms of human hands by Georg Meissner and Rudolph Wagner in 1852 (Cauna, 1956), but a high-resolution description of their morphology was not possible until the advent of electron microscopy in the mid-20th century (Pease and Pallie, 1959; Cauna and Ross, 1960). The oval shaped corpuscle structure is composed of disc-like lamellar stacks derived from Schwann cells, which is partially surrounded by a thin fibroblast capsule. In most cases, the corpuscle is innervated by a myelinated Aβ sensory fiber, which loses its myelination after traveling through the bottom third of the corpuscle (Zelena, 1994). The sensory axon takes a circuitous route within the corpuscle, weaving between stacks of lamellar cells. In addition to Aβ innervation, some Meissner’s corpuscles are also innervated by C-fibers which have molecular profiles similar to nociceptors (Paré et al., 2001), but the functional relevance of this innervation is unclear.
A combination of physiological recording and horseradish peroxidase tracing has been employed to reveal morphologies of the central projections of mammalian Aβ low-threshold mechanoreceptors (Brown, 1981). Shortly after entering the spinal cord, their central axon bifurcates and sends projections in the rostral and caudal directions in the superficial layer of the spinal cord. Interstitial branches arise from these projections, and innervate deeper layers of the dorsal horn (Brown, 1981). In cats, Meissner’s corpuscle afferents synapse mostly in the medial aspect of layers III-IV of the dorsal horn, while in rats they mostly innervate the medial aspect of layers III-V of the dorsal horn, with a small number of synapses made in the inner part of layer II (Brown, 1981; Semba et al., 1985; Shortland and Woolf, 1993).
2.2 Physiological properties and function
Meissner’s corpuscles are classified as type I RA (RAI) mechanoreceptors. In single unit recordings from humans and monkeys, Meissner’s corpuscle innervating fibers display a burst of activity upon the onset and offset of a stimulus, but remain silent during the static phase of stimulation (Talbot et al., 1968). They have an extremely low threshold for activation, responding to an indentation of the skin of less than 10µm (Iggo and Ogawa, 1977). In addition, Meissner’s corpuscles have relatively small receptive fields and are most sensitive to low intensity stimuli of ~5–100Hz (Gardner and Palmer, 1990). When stimulated at frequencies within their optimal range, Meissner’s corpuscle afferents produce action potentials in a nearly perfect one-to-one relationship with the stimulus and generate a “fluttering” feeling in human subjects (Talbot et al., 1968). Meissner’s corpuscles may also function as velocity detectors to determine the rate of skin indentation (Willis and Coggeshall, 2004).
At present, the precise mechanical transduction mechanism for Meissner’s corpuscles is unknown. A recent study has identified the KCNQ4 potassium channel, which is expressed in Meissner’s corpuscle innervating Aβ low-threshold mechanoreceptors, as an important molecule to tune their sensitivity (Heidenreich et al., 2012). KCNQ4 itself is not mechanically gated, but is important for setting the resting potential of neurons. Both mice and humans with KCNQ4 mutations display higher sensitivity to low frequency stimuli, which are mediated by Meissner’s corpuscles (Heidenreich et al., 2012).
2.3 Development
Meissner’s corpuscle innervating axons reach the dermal papilla around birth. Schwann cells associated with the axon begin to differentiate into the lamellar cells, which form the corpuscle structure around one week in rodents. Innervation is required for the development of the corpuscle structure, as corpuscles do not form if the footpad is denervated at birth (Zelená et al., 1990).
Meissner’s corpuscles depend on the neurotrophic receptor tyrosine kinase TrkB for their development. TrkB is expressed in a subset of RA mechanoreceptor neurons starting at early developmental stages (Luo et al., 2009). In TrkB null mice, Meissner’s corpuscles were not found in the dermal papilla of 2–3 week old mice (González-Martínez et al., 2004). Examination of mice lacking the high affinity ligands of TrkB, brain derived neurotrophic factor (BDNF) or neurotrophin-4 (NT4), revealed that BDNF, but not NT4, is required for Meissner’s corpuscle development (González-Martínez et al., 2005). Since the number of DRG neurons is reduced by ~30% in TrkB and BDNF mutant mice in the second postnatal week, it is possible that BDNF-TrkB signaling supports the survival of Meissner’s corpuscle neurons and the loss of Meissner’s corpuscles is due to the death of innervating neurons (Perez-Pinera et al., 2008). However, the mechanism of TrkB signaling in controlling Meissner’s corpuscle formation could be more complicated. Immunolocalization of TrkB in human digit skin shows that the receptor is expressed in the lamellar cells of the corpuscle, but not on the innervating axon (Calavia et al., 2010) and the overexpression of NT4 or BDNF in the skin led to an increase in the size of Meissner’s corpuscles, but no change in the number of sensory neurons in the DRG (LeMaster et al., 1999; Krimm et al., 2006). In addition, some myelinated axons are present in the dermal papillae of neural crest specific TrkB mutants, even though no corpuscle structure is formed, and the central projections of Aβ low-threshold mechanoreceptors do not seem to be affected (Luo et al., 2009). These results suggests that at least some Meissner’s corpuscle innervating neurons do not die in TrkB mutants, and the lack of corpuscles may be due to a pro-survival independent function of TrkB signaling. Future studies using tissue specific knockouts of TrkB and BDNF will be necessary to determine the spatial and temporal requirement of TrkB and its ligands in Meissner’s corpuscle formation.
Meissner’s corpuscles also express another neurotrophic receptor tyrosine kinase, Ret, during early development (early Ret+ RA mechanoreceptors) (Luo et al., 2009). Surprisingly, Ret signaling is not essential for Meissner’s corpuscle formation, as corpuscles are present, although somewhat underdeveloped, in neural crest specific Ret mutants. On the other hand, Ret signaling is absolutely required for the central projections of all RA mechanoreceptors. In Ret mutants, central projections of RA mechanoreceptors reach the dorsal spinal cord, but fail to extend interstitial branches to innervate layers III-V of the dorsal horn (Bourane et al., 2009; Luo et al., 2009; Honma et al., 2010).
In addition to extrinsic neurotrophic signaling, intrinsic transcriptional programs play important roles to specify the neural identity of somatosensory neurons. One transcription factor, Shox2, is critical for the innervation and development of Meissner’s corpuscles. Ablation of Shox2 in mice leads to a lack of TrkB expression in two thirds of the DRG neurons which normally express the receptor during embryonic development (Abdo et al., 2011; Scott et al., 2011). As a result, Shox2 mutant mice lack Meissner’s corpuscles and show reduced innervation of the dermal papillae, while the heterozygous shox2 adult mice display an increased threshold for light touch detection (Abdo et al., 2011). Deletion of Shox2 also caused reduced touch sensory axonal innervation to layers III/IV of the spinal cord (Scott et al., 2011). Additionally, two Maf family transcription factors, MafA and c-Maf, are expressed in the Ret+ RA mechanoreceptors. Ret and MafA reciprocally regulate the expression of each other, whereas c-Maf is upstream of Ret expression (Bourane et al., 2009; Hu et al., 2012; Wende et al., 2012). In c-Maf mutant mice, the number of Meissner’s corpuscles is drastically reduced and the remaining corpuscles have a rudimentary structure, although the innervation of the dermal papillae isn’t affected (Wende et al., 2012). A deficit in the formation of Meissner’s corpuscles was not described in MafA mutant mice since these mutants were not examined any later than postnatal day zero (P0) (Bourane et al., 2009).
3. Pacinian corpuscles
3.1 Anatomical location and morphology
The anatomical location of Pacinian corpuscles varies greatly between species. In primates, Pacinian corpuscles are most prominently located in the subcutaneous fat pads of the fingers, palms, and soles (Zelena, 1994). They are also found in joints, tendons, interosseous membrane, and around some muscles and internal organs, such as the pancreas (Bell et al., 1994). Many early physiological and morphological studies of Pacinian corpuscles were performed on corpuscles isolated from the cat mesentery (Sato, 1961). In rodents, Pacinian corpuscles are not present in the skin but are enriched in the interosseous membrane around the fibula and ulna (Zelená, 1978). The Herbst corpuscle, a structure similar to the Pacinian corpuscle, is found in beak and interosseous membrane of birds (Saxod, 1996; Zelená et al., 1997).
Pacinian corpuscles were first described in the 18th century by Johannes Gottlieb Lehmann, and later rediscovered by Fillipo Pacini in 1841 (Bentivoglio and Pacini, 1995). They are oval shaped end organs and can reach sizes of up to 4 mm in length in adult human (Cauna and Mannan, 1958). Each Pacinian corpuscle is innervated by a single myelinated Aβ somatosensory neuron, which loses its myelination and assumes a relatively straight trajectory through the center of the corpuscle upon entering the corpuscle’s inner core (Quilliam and Sato, 1955). Ultrastructural studies of the corpuscle have revealed that the inner core is composed of two “hemilamellae” on either side of the axon, with two clefts separating them, throughout the length of the inner core. Each hemilamella of the mature corpuscle contains 40–80 layers of lamellar Schwann cells while the outer core is composed of ~30 layers of perinurial epithelial cells (Pease and Quilliam, 1957; Cauna and Mannan, 1958; Zelena, 1994). This layered construction produces an onion-like appearance in cross sections of Pacinian corpuscles.
Central projections of Pacinian afferents form synapses in two distinct regions: a larger dorsal region focused in layer III and outer layer IV with less dense innervation extending dorsally to inner layer II and ventrally to outer layer V, and a smaller ventral region concentrated in layer V but also sparsely innervating layers IV and VI (Brown et al., 1980; Brown, 1981; Semba et al., 1984).
3.2 Physiological properties and function
Pacinian corpuscles are classified as type II RA (RAII) mechanoreceptors, which, like Meissner’s corpuscles, respond to mechanical stimuli at the onset and offset of stimuli. However, the RAI and RAII mechanoreceptors can be distinguished in two ways: 1) RAII mechanoreceptors have larger, less defined receptive fields, suggesting a poor ability of Pacinian corpuscles to localize stimuli (Palmer and Gardner, 1990); and 2) RAII mechanoreceptors respond to a higher frequency of vibration, and are most sensitive to stimuli in the 200–300Hz range (Burgess, 1973; Knibestöl, 1973).
The precise mechanical transduction mechanism for Pacinian corpuscles is also unknown. The RA properties of Pacinian corpuscles are partly due to the corpuscle structure, which acts as a mechanical filter. Very low velocity or static stimuli cause compression of the outer layers of the corpuscle, but this compression does not reach the inner core. Instead, only the dynamic phase of compression results in deformation of the corpuscle to the inner core, evoking a response from the innervating axon (Hubbard, 1958). This model was supported by experiments in which removing the capsule of Pacinian corpuscles resulted in a prolonged generator potential upon a sustained mechanical stimulus (Mendelson and Lowenstein, 1964). However, despite the prolonged generator potential, the innervating Aβ axon still only fired a few action potentials at the onset of stimulation. This suggests that inherent properties of these Aβ axons prevents a steady outward current from producing repetitive impulses, which is another potential mechanism underlying the RA response (Mendelson and Lowenstein, 1964; Loewenstein and Mendelson, 1965).
Pacinian corpuscles are exquisitely sensitive; in physiological preparations ambient vibrations in the building resulted in a response from the innervating axon (Hunt, 1961). These observations suggest that one potential function of Pacinian corpuscles, especially those in the interosseous membrane, may be to sense vibration transmitted through the skeletal system, either due to movement of the animal or due to external environmental vibrations, possibly generated by predators or prey. In humans, Pacinian corpuscles in the hand are tuned to sense the texture of an object or its dimensions indirectly through the use of tools (Brisben et al., 1999). In addition, Pacinian corpuscles are important for detecting the fine texture of objects. Experiments with biomimetic sensors have shown that the normal spacing of fingerprints causes the amplification of vibrations in the ideal detection range of Pacinian corpuscles when scanning across a finely textured surface (Scheibert et al., 2009).
3.3 Development
Most studies regarding the development of Pacinian corpuscles have been performed on the corpuscles in the interosseous membrane of rodents. In rats and mice, an immature inner core and an outer capsule containing only a couple layers is present at birth (Zelena, 1994). The inner core becomes morphologically mature and more outer core layers are added in the first postnatal week. Outer core layers continue to be added during the first few weeks of life, and the corpuscle grows in size over the first few months. Innervation is required for the development of the corpuscle, as Pacinian corpuscles won’t form if the leg is neonatally denervated (Zelená et al., 1990).
Pacinian corpuscle neurons arise from the early Ret+ RA mechanoreceptors and are highly dependent on Ret signaling for their development. In mice mutant for Ret, its co-receptor GFRα2, or its ligand Neurturin, no Pacinian corpuscles are formed (Luo et al., 2009). The cause of the selective loss of Pacinian corpuscles, but not other subtypes of RA mechanoreceptors, in Ret mutant mice is currently unclear.
In addition, several transcription factors play critical roles in the development of Pacinian corpuscles. The ETS transcription factor Er81 is expressed in the inner core Schwann cells of Pacinian corpuscles, and no Pacinian corpuscles are formed in Er81 null mice (Sedý et al., 2006). In c-Maf mutant mice, both the number of Pacinian corpuscles and axons in the interosseous nerve, which innervate Pacinian corpuscles, were greatly reduced. In addition, the remaining corpuscles display abnormal morphology (Hu et al., 2012; Wende et al., 2012). These results suggest that c-Maf is required in Pacinian corpuscle neurons for axonal growth/targeting or corpuscle formation. Interestingly, human patients with c-MAF missense mutations showed a somatosensory deficit specifically related to Pacinian corpuscles. These patients have a decreased sensitivity to high frequency vibration, which is detected by Pacinian corpuscles, while their detection of lower frequencies, which is mediated by Meissner’s corpuscles, is not affected (Wende et al., 2012).
4. Lanceolate endings
4.1 Anatomical location and morphology
The innervation of hair follicles by low-threshold mechanosensory neurons is very complex. Aβ, Aδ, and C low-threshold mechanoreceptors all innervate hair follicles and form a palisade structure surrounding the follicle (Li et al., 2011; Lou et al., 2013). In addition, different types of hair are innervated by different combinations of sensory fibers, and hair types vary by anatomical location and between species. In keeping with the scope of this review, we will only discuss the Aβ low-threshold mechanoreceptors in the hairy skin and whisker pad, which are Aβ lanceolate endings and Merkel cells (see below). Aβ lanceolate endings are associated with the awl and guard hairs of the hairy back skin and the whiskers, but not the zigzag hairs, which are the most numerous hair type in mouse hairy back skin (Mosconi et al., 1993; Li et al., 2011).
Hoggan G and Hoggan F E originally described forked nerve endings surrounding hair follicles in 1893 (Hoggan G and Hoggan F E, 1893). The lanceolate endings form a palisade-like structure which encircles the hair follicle. Individual endings have a flattened, oval shape, with the thin aspect directly abutting the hair follicle. Each ending is composed of a single oblong axonal fiber enclosed by flattened Schwann cells on either side of the axon. Axonal spikes protruding through the sheath where the two Schwann cell faces meet contact both the hair follicle and the surrounding connective tissue (Munger and Ide, 1988). Sparse genetic labeling of hair follicle innervating axons shows that the lanceolate endings which make up each palisade structure are innervated by more than one sensory afferent, and that individual mechanoreceptive neurons can innervate lanceolate endings surrounding multiple hair follicles (Suzuki et al., 2012).
Centrally, Aβ hair follicle afferents form distinct “flame-shaped” collateral arbors, which were first described by Ramon y Cajal and later characterized by Scheibel M E and Scheibel A B in 1968 (Scheibel M E and Scheibel A B, 1968). Upon entering the dorsal horn, the afferent fiber descends to layer IV or V, and then reverses direction and projects dorsally to the outer layer IV and layer III, where the collateral undergoes extensive branching and forms synapses with spinal neurons (Brown, 1981; Woodbury et al., 2001).
4.2 Physiological properties and function
Like Meissner’s and Pacinian corpuscles, Aβ lanceolate endings also display RA properties. In recordings from cat whisker hair, two populations of RA afferents were found. The more numerous population responded only to high velocity stimuli, such as a flick of the hair or vibration from a tuning fork. This population could follow frequencies up to 1000 Hz and was not affected by the direction of the stimulus. A much smaller low velocity population of RA mechanoreceptors responded to stimuli from 5 to 200 Hz and exhibited some direction selectivity. They responded to hair deformation in any direction, but had a much lower activation threshold in the preferred direction (Gottschaldt et al., 1973). The RA hair follicle afferents are also extremely sensitive; in tests for the activation threshold using the skin nerve preparation, the activation threshold of more than half of the units recorded was below the level of resolution (0.07 mN) in mice (Woodbury et al., 2001).
Aβ lanceolate endings most likely function as velocity detectors for the deformation of hair. The mechanical transduction mechanism is unclear, but ultrastructural studies suggest that deformation of the lanceolate ending may occur upon movement of the hair, due to the connections of the Schwann cell structure to the hair follicle and surrounding tissue. This physical deformation may cause a response in the innervating axon (Takahashi-Iwanaga, 2000).
4.3 Development
Due to the complex innervation of hair follicles by different types of sensory neurons, it has been difficult to exclusively study the development of Aβ lanceolate endings. Most studies described below depend on panneural or Schwann cell markers to identify lanceolate endings. Therefore, the observed phenotypes in mutant mice may reflect deficits in Aβ as well as Aδ and C low-threshold mechanoreceptors. Recently, new mouse genetic tools have been developed to specifically label different classes of hair follicle innervating neurons (Li et al., 2011), which will help to identify molecular mechanisms that control development of different types of low-threshold mechanoreceptors in the future.
The trigeminal nerve approaches the site of whisker hair follicles by embryonic day 12 (E12) in mice and the lanceolate endings start to appear at E17 (Maklad et al., 2010). In the back skin, the development of lanceolate endings is slightly delayed; nerve fibers reach the area of hair follicles around E14-E16 and immature lanceolate endings appear around birth (Peters et al., 2002). These anatomical studies correlate well with physiological recordings. Using the ex vivo skin nerve preparation, RA responses from hair follicle innervating neurons could be recorded at P0, and backfilled neurons displayed “flame-like” central innervation morphology typical of hair follicle receptors by P2 (Woodbury et al., 2001).
The role of neurotrophic signaling in controlling the development of Aβ hair follicle innervating neurons is less clear. Like the other low threshold RA mechanoreceptors, Aβ hair follicle innervating neurons express Ret during early development. In P14 neural crest specific Ret null mice, Luo et al. found that the morphology of lanceolate endings is disorganized, but the percentage of hair follicles that receive lanceolate ending innervation was not significantly different using a mixed pan-neuronal marker PGP9.5 and large diameter axon marker NF200 (Luo et al., 2009). In P0 Ret null mice, Bourane et al. found that the total innervated area of NF200+ lanceolate endings around the hair follicle is significantly reduced (Bourane et al., 2009). Although the conclusions from these two studies seem to be slightly different on the surface, which could well be explained by the different staining and quantification method, both studies in fact suggest that the normal morphology of Aβ lanceolate ending is dependent on Ret signaling.
Hair follicle innervation may also be dependent on TrkB signaling. In TrkB mutants, the morphology of lanceolate endings is affected to a similar extent as seen in Ret mutants. In addition, there was a reduction in the number of hair follicles innervated by lanceolate endings in TrkB null mice (Perez-Pinera et al., 2008), but the number of endings was not significantly changed in neural crest specific TrkB mutant mice (Luo et al., 2009). This discrepancy could be due to either the differences between TrkB null and neural crest conditional knockout mice or the technical difficulties of specifically identifying Aβ lanceolate endings and quantifying them. On the other hand, overexpression of TrkB ligands BDNF and NT4 in the skin led to an increased density of hair follicle innervation (LeMaster et al., 1999; Krimm et al., 2006). Lastly, as discussed above, the transcription factor Shox2 promotes the expression of TrkB in DRG neurons. In Shox2 null mice, lanceolate endings are disorganized, a phenotype very similar to the TrkB mutant phenotype (Abdo et al., 2011).
The proper formation of lanceolate endings may also depend on the activity of the innervating neurons. Woo et al. recently found that lanceolate ending innervating neurons express the vesicular glutamate transporter VGLUT2, while the Schwann cells which surround the innervating axons express NMDA receptors. Interestingly, ablating VGLUT2 from somatosensory neurons leads to a reduction in the frequency and organization of lanceolate forming Schwann cells at P0 (the latest stage the mice survived). Moreover, pharmacologically blocking glutamatergic transmission by injecting an NMDA receptor antagonist in the skin led to a reduction in the number and organization of lanceolate Schwann cell processes in adult wild-type mice. Lastly, physiological tests of antagonist treated mice revealed a decrease in the sensitivity and conduction velocity of RA afferents innervating hair follicles (Woo et al., 2012). Collectively, these results suggest that communication between the innervating axon and the surrounding Schwann cell structure are crucial for the development and maintenance of lanceolate endings.
5. Merkel cells
5.1 Anatomical location and morphology
Merkel cells are located in the basal epidermis of both glabrous and hairy skin of mammals. In glabrous skin, clusters of 4–40 Merkel cells are present in the epidermal pegs, which are protrusions of the epidermis into the dermis that surrounds the dermal papillae. In primates and marsupials, smaller clusters of Merkel cells are located at the base of the epidermal ridges which are responsible for the fingerprint pattern of the hands and feet (Halata et al., 2003). These clusters of Merkel cells in glabrous skin are often referred to as “touch spots” (Boulais and Misery, 2007). In hairy skin, Merkel cells are present in “touch domes,” (Iggo and Ogawa, 1969) which can be discerned by a slight elevation in the skin in some species, and can contain up to 150 Merkel cells. Touch domes may or may not be associated with a hair follicle (Zelena, 1994). In rodents, Merkel cells are closely associated with guard hair follicles, located in the epidermis in “collars” surrounding the hair follicle, and whiskers, located in “cuffs” present underneath the glassy membrane of the follicle (Halata and Munger, 1980b; Zelena, 1994).
Merkel cells were first described as “touch cells” by Freidrich Sigmund Merkel in 1875 (Merkel, 1875). They are oval in shape and 10–15 µm in length along the long axis, which is the smallest among all mechanosensory endings discussed in this review. They can be differentiated from the surrounding epidermal cells by their large, multilobated nucleus which is oriented parallel to the dermis-epidermis junction. On the basal side of the cell, numerous dense core vesicles measuring 70–180 nm in diameter are located close to the cell membrane (Iggo and Muir, 1969; Winkelmann and Breathnach, 1973; Tachibana and Nawa, 2002; Halata et al., 2003). Most Merkel cells are associated with an innervating axon, forming a structure referred to as the Merkel cell-neurite complex. A nerve plate, which is formed by a myelinated axon which loses its myelin sheath upon entering the epidermis, directly opposes the vesicle dense basal membrane of the Merkel cell. This plate is separated from the Merkel cell by 15 nm, but in small regions they are separated by only 13 nm, and electron dense material is observed in both the Merkel cell and the nerve plate at these points (Iggo and Muir, 1969; Halata et al., 2003). A small number of Merkel cells do not make contact with an innervating axon. The function of these uninnervated Merkel cells is unknown, but it has been suggested they may have neuroendocrine or immune system functions (Boulais and Misery, 2007).
Central projections of Merkel cell innervating neurons bifurcate upon entering the spinal cord, and send collaterals into the dorsal horn as these branches travel anteriorly and posteriorly. The morphology of the collaterals innervating the dorsal horn are distinct form the RA mechanoreceptors. Individual collaterals dive into the dorsal horn perpendicular to the dorsal surface of the spinal cord. After reaching layer IV or V, the collaterals make a C- or L-shaped turn and then travel medially. During and after this turn, the collaterals gives off terminal arborizations in layers III–V (Brown, 1981).
5.2 Physiological properties and function
Merkel cell-neurite complexes are type I SA (SAI) mechanoreceptors. Unlike the RA mechanoreceptors discussed above, SA mechanoreceptors remain active during the static phase of stimuli. The innervating neuron is usually silent at rest, and responds to the onset of stimulation with a burst of activity, which is proportional to the velocity and displacement of the stimulus. After the initial phasic burst of activity, a tonic firing phase occurs for the duration of the application of the stimulus. The firing pattern during the tonic phase is irregular and can last for over 30 min (Tapper, 1965; Iggo and Muir, 1969; Willis and Coggeshall, 2004).
There has been a long-standing debate with regard to the exact role of Merkel cells in light touch sensation. Many studies suggested that the Merkel cell is critical for transducing the mechanical stimulus into a chemical signal to activate the innervating neurite, while other studies proposed that Merkel cells may play a modulatory role and the neurite is primarily responsible for transducing the mechanical stimulus. Here we will review the physiological and biochemical findings in support of both arguments.
There is abundant biochemical evidence that Merkel cells produce neurotransmitters and the machinery required for synaptic release (Tachibana and Nawa, 2002; Haeberle et al., 2004; Maksimovic et al., 2013). Merkel cells also express voltage gated calcium channels, and calcium induced calcium release from internal stores occurs upon the entry of calcium into the cell, providing a potential mechanism for neurotransmitter release (Yamashita et al., 1992; Senok and Baumann, 1997). In addition, blocking glutamatergic transmission reduced the SA response evoked by activation of Merkel cell-neurite complexes, suggesting that excitatory neurotransmission is required for transducing the mechanical stimulus (Fagan and Cahusac, 2001). The most compelling evidence suggesting a mechanosensory function of Merkel cells comes from Atoh1/Math1 conditional knockout mice, in which Merkel cells are not differentiated but the innervating fibers are still present in touch domes. Strikingly, although the total number of Aβ fibers is not significant changed, SAI responses could not be detected in these animals using the ex vivo skin nerve preparation (Maricich et al., 2009). These results suggest that Merkel cells are essential for mediating the SAI response. However, it is unclear whether these remaining fibers completely lost their mechanosensitivity or display physiological properties similar to RA mechanoreceptors.
On the other hand, using both ultrastructural and electrophysiologicalal evidence, Gottschaldt and Vahle-Hinz argued that the ability of Merkel cell innervating neurites to follow high frequency stimuli up to 1200 Hz with a one-to-one response for up to 500 ms is incompatible with chemical communication, as neurotransmitter could not be released and cleared from the synapse quickly enough to produce such a precise response. In addition, the latency from application of stimulus to response in the innervating fiber was too fast for chemical transmission (Gottschaldt and Vahle-Hinz, 1981), further supporting their model that the innervating neurite acts as the mechanosensitive element.
Diamond’s group also argued that Merkel cells are dispensable for mechanosensation. They found that touch domes were still mechanoresponsive after selective destruction of Merkel cells using quinacrine loading and ultraviolet (UV) light irradiation (Diamond et al., 1988). However, Ikeda et al. found that SAI responses evoked by touch dome stimulation were lost using a different irradiation procedure to eliminate Merkel cells (Ikeda et al., 1994). Further investigation revealed that quinacrine loading/UV irradiation is not selective and incomplete, leaving some Merkel cells relatively intact and damaging other nerve fibers in the skin (Senok et al., 1996). These conflicting results and technical issues make these experiments difficult to interpret.
Others have attempted to reconcile the conflicting findings regarding the role of the Merkel cell in mechanotransduction with a two-receptor-site model, in which both the innervating neurite and the Merkel cell are mechanosensitive. According to this model, the early phasic activity is mediated by the neurite while the late tonic phase is due to chemical communication between the Merkel cell and the neurite (Ogawa, 1996; Maksimovic et al., 2013). A key to resolving this debate is to develop new tools, by which Merkel cell innervating axons can be specifically identified for physiological recording while Merkel cells are acutely and selectively ablated.
As a result of their small sizes, Merkel cell-neurite complexes have the smallest receptive fields among all mechanoreceptors and are best able to distinguish individual closely spaced objects. Due to these characteristics, Merkel cells are proposed to be essential for detecting the fine details of touched objects, such as shape, texture, and curvature (Johnson et al., 2000; Johnson, 2001). A recent study found that mice lacking Merkel cells were unable to detect certain textures with their feet (Maricich et al., 2012).
5.3 Development
In contrast to the RA mechanoreceptor end organs, Merkel cells appear in the skin prior to the arrival of innervating fibers (Saxod, 1996). Immature Merkel cells can be observed in the epidermis of the rat around E16, when innervating axons have only reached the dermis. Axons reach the epidermis and are found in close association with Merkel cells a day later (English et al., 1980).
Since Merkel cells share characteristics with both epidermal and neural cells, there has been argument concerning the embryological origins of Merkel cells in mammals (Lucarz and Brand, 2007). In birds, chick-quail chimera experiments offer strong support to a neural crest origin of Merkel cells (Grim and Halata, 2000). In addition, Merkel cells were labeled by lacZ when all neural crest derived cells were genetically labeled using Wnt1Cre and ROSA26R-β-Galactosidase reporter mice, suggesting that mouse Merkel cells arose from a neural crest origin (Szeder et al., 2003). However, there is also genetic evidence to support an epidermal origin of Merkel cells. The transcription factor Atoh1/Math1 is highly expressed and functionally required for the development of Merkel cells. Surprisingly, Merkel cells still form when Atoh1 is conditionally ablated from neural crest cells. In contrast, when Atoh1 is deleted from the basal layer of the epidermis using a Keratin14Cre line, Merkel cells do not form, suggesting an epidermal origin for Merkel cells in mammals (Morrison et al., 2009). Nevertheless, this study could not exclude a cell non-autonomous effect for Atoh1 in the development of Merkel cells, as Atoh1 expression was also observed in some of the accessory cells surrounding Merkel cells.
Inherent transcriptional programs in the innervating neuron are also required for development of Merkel cell-neurite complex. Similar to Meissner’s corpuscles and lanceolate endings, Merkel cells are dependent on the transcription factor Shox2 for sensory innervation during development. In Shox2 mutants, Merkel cells are present in both glabrous and hairy skin, but there is a dramatic decrease in the percentage of Merkel cells innervated by large diameter sensory fibers (Abdo et al., 2011). In addition, some mechanoreceptive DRG neurons co-express the Runt-related transcription factors Runx1 and Runx3 (Yoshikawa et al., 2013), and the number of Merkel cell-neurite complex surrounding the whiskers is greatly reduced in Runx3 mutant mice (Senzaki et al., 2010). One plausible mechanism by which Runx3 controls Merkel cell-neurite complex development is to regulate TrkC expression (Levanon et al., 2002; Kramer et al., 2006; Nakamura et al., 2008).
In addition to transcriptional programs, Merkel cell-neurite complexes are highly dependent on several types of neurotrophic signaling for their development (Montaño et al., 2010). One population of Merkel cells depends on TrkA/NGF signaling. The number of Merkel cells surrounding hair follicles and the number of innervating axons is reduced in TrkA mutants. However, remaining Merkel cells are maintained into adulthood, suggesting a TrkA independent Merkel cell population. Loss of the TrkA ligand NGF produces a similar, but less severe, phenotype (Fundin et al., 1997).
TrkC/NT3 signaling has a significant and complicated effect on Merkel cell development. TrkC is expressed in both Merkel cells and Merkel cell innervating somatosensory neurons. In mice lacking the kinase domain of TrkC, the number of Merkel cells is reduced at birth. In addition, those that are present at birth are not maintained during the first two postnatal weeks, suggesting that all Merkel cells become dependent on TrkC signaling postnatally (Cronk et al., 2002). Loss of NT3, the TrkC ligand, leads to a more severe deficit, with even fewer Merkel cells present at birth (Airaksinen et al., 1996). The phenotype becomes most severe when all isoforms of TrkC are eliminated in TrkC complete null mice, in which no Merkel cells or innervating fibers are present at birth, suggesting additional kinase-independent roles of TrkC in Merkel cell development (Fundin et al., 1997; Cronk et al., 2002).
TrkB signaling also has an effect on Merkel cell development. In TrkB mutant mice, the number of Merkel cells surrounding hair follicles and in the glabrous skin is greatly reduced (Perez-Pinera et al., 2008). When BDNF is overexpressed in the skin, the number of Merkel cells is increased in the glabrous but not hairy skin (LeMaster et al., 1999). Interestingly, the mechanical threshold of SAI mechanoreceptors increases 8-fold in the BDNF heterozygous and null mice, although the number and morphology of Merkel cells was normal in touch domes of P14 BDNF null animals. This deficit could be rescued by injecting recombinant BDNF into BDNF heterozygous mice (Carroll et al., 1998).
Lastly, the low affinity neurotrophin receptor p75 also plays a role in Merkel cell development. p75 can bind NGF, BDNF, NT3, and NT4, and interact with the Trk receptors (Skaper, 2012). In p75 mutant mice, Merkel cells develop normally during the first two postnatal weeks but then then slowly decrease in number over the following months until very few remain (Fundin et al., 1997; Kinkelin et al., 1999).
6. Ruffini corpuscles
6.1 Anatomical location and morphology
The Ruffini corpuscle is an elongated structure with tapered ends. Morphologically, it is quite similar to the Golgi tendon organs which are innervated by proprioceptors (Halata and Munger, 1980a). The corpuscle is usually encased in a capsule of 4–5 layers of perineural cells and contains an inner core of Schwann cells and collagen, which is innervated by a single large diameter myelinated axon that loses its myelination upon entry into the inner core. The axon gives off numerous terminal branches within the inner core (Chambers et al., 1972; Willis and Coggeshall, 2004). Collagen fibers associated with the inner core exit the poles of the Ruffini corpuscle and interact with collagen in the surrounding tissue, providing a potential mechanism for mechanically linking the inner core with the surrounding tissue (Halata, 1977).
The central projections of SA type II (SAII) mechanoreceptors, which are presumed to innervate Ruffini corpuscles, are distinct form other mechanoreceptors. Collaterals innervating the dorsal horn project to layer III, and then branch into at least two processes. These processes travel deeper into the dorsal horn and branch extensively, forming terminal arborizations from layer III-VI (Brown, 1981).
The first extensive morphological characterization of Ruffini corpuscles was performed on hairy skin of the cat (Chambers et al., 1972). However, the anatomical location and existence of Ruffini corpuscles between tissues and species is currently under debate. In many cases, numerous units with SAII responses can be recorded in nerve fibers innervating a tissue, but the Ruffini corpuscles cannot be found in the tissue following careful histological examination. For example, physiological recordings of nerves innervating the glabrous skin of raccoons and humans have shown a relatively high proportion of units exhibiting SAII responses (Johansson and Vallbo, 1979; Rasmusson and Turnbull, 1986). However, when glabrous skin from monkeys and raccoons was examined, no Ruffini corpuscles were found (Rice and Rasmusson, 2000; Paré et al., 2002). In humans, a single Ruffini corpuscle was found in the skin of the index finger, which is much less than what would be expected based on the physiological recordings (Paré et al., 2003). Notably, Paré, et al., observed innervation of blood vessels which looked morphologically similar to previous descriptions of Ruffini corpuscles. The authors suggest that previous studies may have misidentified these structures as Ruffini corpuscles, which could explain the discrepancy in previous findings (Paré et al., 2002). In mouse hairy skin, SAII fibers are also identified by physiological recordings but no definite Ruffini corpuscle structure has been reported (Wellnitz et al., 2010).
In many species, sensory endings which are morphologically similar to Ruffini corpuscles have been identified. In monkeys and raccoons, unencapsulated Ruffini-like endings were found at the base of the fingernail/claw (Rice and Rasmusson, 2000; Paré et al., 2002). Ruffini corpuscles have also been found in association with hair follicles, where they are sometimes referred to as pilo-Ruffini complexes (Biemesderfer et al., 1978). Additionally, unencapsulated periodontal Ruffini-like corpuscles have been identified surrounding the teeth of rodents (Byers, 1985).
6.2 Physiology and function
Although not well defined morphologically, the physiology of SAII Aβ low-threshold mechanoreceptors have been extensively characterized in both humans and model organisms (Johansson and Vallbo, 1979; Wellnitz et al., 2010). Like the SAI response, the SAII responses is characterized by an early dynamic phase which is sensitive to both the velocity and displacement of the stimulus, followed by a static response phase that last throughout the application of stimulus. However, SAII responses can be differentiated from SAI responses because they usually display some background firing activity when no stimulus is applied, they fire at a much more regular rate during the static phase, and their maximum frequency of the response is less than that of the SAI response (Chambers et al., 1972).
SAII Aβ low-threshold mechanoreceptors are proposed to act primarily as stretch receptors. In psychophysical recordings SAII units were less sensitive than SAI units to skin indentation, but were much more sensitive to stretching of the skin (Johnson et al., 2000). The stretch receptors in the skin may have two functions. They may work in combination with RA mechanoreceptors to sense movement of grasped objects. In addition, they may work in concert with proprioceptors to sense the position of the fingers and hand, as skin stretch will vary based on grasp (Johnson, 2001).
6.3 Development
Due to the difficulty in clearly identifying Ruffini corpuscles by morphology, relatively little work has been done to study their development compared to the other mechanoreceptors discussed in this review. Both periodontal Ruffini-like endings and those associated with whisker hair are dependent on TrkB neurotrophic signaling in mice. Periodontal Ruffini-like endings are absent in TrkB mutant mice, and mice lacking either BDNF or NT4 show Ruffini-like endings with immature morphology (Matsuo et al., 2002; Hoshino et al., 2003; Maruyama et al., 2005). The Ruffini-like endings of whisker hairs are also absent in TrkB mutants. In addition, the number of whisker Ruffini-like endings is greatly reduced in BDNF mutants but is unaffected in NT4 mutants. Furthermore, the number of Ruffini-like endings associated with whiskers is increased in NT3 mutant mice (Fundin et al., 1997).
Footnotes
Compliance with ethics guidelines
The authors declare no conflict of interest.
References
- Abdo H, Li L, Lallemend F, Bachy I, Xu XJ, Rice FL, Ernfors P. Dependence on the transcription factor Shox2 for specification of sensory neurons conveying discriminative touch. Eur J Neurosci. 2011;34(10):1529–1541. doi: 10.1111/j.1460-9568.2011.07883.x. PMID:22103411. [DOI] [PubMed] [Google Scholar]
- Airaksinen MS, Koltzenburg M, Lewin GR, Masu Y, Helbig C, Wolf E, Brem G, Toyka KV, Thoenen H, Meyer M. Specific subtypes of cutaneous mechanoreceptors require neurotrophin-3 following peripheral target innervation. Neuron. 1996;16(2):287–295. doi: 10.1016/s0896-6273(00)80047-1. PMID:8789944. [DOI] [PubMed] [Google Scholar]
- Albrecht FLRPJ. The Senses: A Comprehensive Reference. Vol. 6. San Diego: Academic Press; 2008. Cutaneous Mechanisms of Tactile Perception: Morphological and Chemical Organization of the Innervation to the Skin; pp. 1–32. [Google Scholar]
- Bell J, Bolanowski S, Holmes MH. The structure and function of Pacinian corpuscles: a review. Prog Neurobiol. 1994;42(1):79–128. doi: 10.1016/0301-0082(94)90022-1. PMID:7480788. [DOI] [PubMed] [Google Scholar]
- Bentivoglio M, Pacini P. Filippo Pacini: a determined observer. Brain Res Bull. 1995;38(2):161–165. doi: 10.1016/0361-9230(95)00083-q. PMID:7583342. [DOI] [PubMed] [Google Scholar]
- Biemesderfer D, Munger BL, Binck J, Dubner R. The pilo-Ruffini complex: a non-sinus hair and associated slowly-adapting mechanoreceptor in primate facial skin. Brain Res. 1978;142(2):197–222. doi: 10.1016/0006-8993(78)90631-5. PMID:415796. [DOI] [PubMed] [Google Scholar]
- Boulais N, Misery L. Merkel cells. J Am Acad Dermatol. 2007;57(1):147–165. doi: 10.1016/j.jaad.2007.02.009. PMID:17412453. [DOI] [PubMed] [Google Scholar]
- Bourane S, Garces A, Venteo S, Pattyn A, Hubert T, Fichard A, Puech S, Boukhaddaoui H, Baudet C, Takahashi S, Valmier J, Carroll P. Low-threshold mechanoreceptor subtypes selectively express MafA and are specified by Ret signaling. Neuron. 2009;64(6):857–870. doi: 10.1016/j.neuron.2009.12.004. PMID:20064392. [DOI] [PubMed] [Google Scholar]
- Brisben AJ, Hsiao SS, Johnson KO. Detection of vibration transmitted through an object grasped in the hand. J Neurophysiol. 1999;81(4):1548–1558. doi: 10.1152/jn.1999.81.4.1548. PMID:10200190. [DOI] [PubMed] [Google Scholar]
- Brown AG. Organization in the spinal cord: the anatomy and physiology of identified neurones. Berlin, New York: Springer-Verlag; 1981. [Google Scholar]
- Brown AG, Fyffe RE, Noble R. Projections from Pacinian corpuscles and rapidly adapting mechanoreceptors of glabrous skin to the cat’s spinal cord. J Physiol. 1980;307:385–400. doi: 10.1113/jphysiol.1980.sp013441. PMID:7205669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgess PRPER. Cutaneious mechanoreceptors and nociceptors. In: Iggo A, editor. Handbook of Sensory Physiology. Vol. 11. Berlin: Springer; 1973. pp. 29–78. [Google Scholar]
- Byers MR. Sensory innervation of periodontal ligament of rat molars consists of unencapsulated Ruffini-like mechanoreceptors and free nerve endings. J Comp Neurol. 1985;231(4):500–518. doi: 10.1002/cne.902310408. PMID:3968252. [DOI] [PubMed] [Google Scholar]
- Calavia MG, Feito J, López-Iglesias L, de Carlos F, García-Suarez O, Pérez-Piñera P, Cobo J, Vega JA. The lamellar cells in human Meissner corpuscles express TrkB. Neurosci Lett. 2010;468(2):106–109. doi: 10.1016/j.neulet.2009.10.076. PMID:19879330. [DOI] [PubMed] [Google Scholar]
- Carroll P, Lewin GR, Koltzenburg M, Toyka KV, Thoenen H. A role for BDNF in mechanosensation. Nat Neurosci. 1998;1(1):42–46. doi: 10.1038/242. PMID:10195107. [DOI] [PubMed] [Google Scholar]
- Cauna N. Nerve supply and nerve endings in Meissner’s corpuscles. Am J Anat. 1956;99(2):315–350. doi: 10.1002/aja.1000990206. PMID:13372495. [DOI] [PubMed] [Google Scholar]
- Cauna N, Mannan G. The structure of human digital pacinian corpuscles (corpus cula lamellosa) and its functional significance. J Anat. 1958;92(1):1–20. PMID:13513492. [PMC free article] [PubMed] [Google Scholar]
- Cauna N, Ross LL. The fine structure of Meissner’s touch corpuscles of human fingers. J Biophys Biochem Cytol. 1960;8(2):467–482. doi: 10.1083/jcb.8.2.467. PMID:13691669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chambers MR, Andres KH, von Duering M, Iggo A. The structure and function of the slowly adapting type II mechanoreceptor in hairy skin. Q J Exp Physiol Cogn Med Sci. 1972;57(4):417–445. doi: 10.1113/expphysiol.1972.sp002177. PMID:4484588. [DOI] [PubMed] [Google Scholar]
- Cronk KM, Wilkinson GA, Grimes R, Wheeler EF, Jhaveri S, Fundin BT, Silos-Santiago I, Tessarollo L, Reichardt LF, Rice FL. Diverse dependencies of developing Merkel innervation on the trkA and both full-length and truncated isoforms of trkC. Development. 2002;129(15):3739–3750. doi: 10.1242/dev.129.15.3739. PMID:12117822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diamond J, Mills LR, Mearow KM. Evidence that the Merkel cell is not the transducer in the mechanosensory Merkel cell-neurite complex. Prog Brain Res. 1988;74:51–56. doi: 10.1016/s0079-6123(08)62997-0. PMID:3055053. [DOI] [PubMed] [Google Scholar]
- English KB, Burgess PR, Kavka-Van Norman D. Development of rat Merkel cells. J Comp Neurol. 1980;194(2):475–496. doi: 10.1002/cne.901940212. PMID:7440811. [DOI] [PubMed] [Google Scholar]
- Fagan BM, Cahusac PM. Evidence for glutamate receptor mediated transmission at mechanoreceptors in the skin. Neuroreport. 2001;12(2):341–347. doi: 10.1097/00001756-200102120-00032. PMID:11209947. [DOI] [PubMed] [Google Scholar]
- Fundin BT, Silos-Santiago I, Ernfors P, Fagan AM, Aldskogius H, DeChiara TM, Phillips HS, Barbacid M, Yancopoulos GD, Rice FL. Differential dependency of cutaneous mechanoreceptors on neurotrophins, trk receptors, and P75 LNGFR. Dev Biol. 1997;190(1):94–116. doi: 10.1006/dbio.1997.8658. PMID:9331334. [DOI] [PubMed] [Google Scholar]
- Gardner EP, Palmer CI. Simulation of motion on the skin. III. Mechanisms used by rapidly adapting cutaneous mechanoreceptors in the primate hand for spatiotemporal resolution and two-point discrimination. J Neurophysiol. 1990;63(4):841–859. doi: 10.1152/jn.1990.63.4.841. PMID:2341881. [DOI] [PubMed] [Google Scholar]
- González-Martínez T, Fariñas I, Del Valle ME, Feito J, Germanà G, Cobo J, Vega JA. BDNF, but not NT-4, is necessary for normal development of Meissner corpuscles. Neurosci Lett. 2005;377(1):12–15. doi: 10.1016/j.neulet.2004.11.078. PMID:15722178. [DOI] [PubMed] [Google Scholar]
- González-Martínez T, Germanà GP, Monjil DF, Silos-Santiago I, de Carlos F, Germanà G, Cobo J, Vega JA. Absence of Meissner corpuscles in the digital pads of mice lacking functional TrkB. Brain Res. 2004;1002(1–2):120–128. doi: 10.1016/j.brainres.2004.01.003. PMID:14988041. [DOI] [PubMed] [Google Scholar]
- Gottschaldt KM, Iggo A, Young DW. Functional characteristics of mechanoreceptors in sinus hair follicles of the cat. J Physiol. 1973;235(2):287–315. doi: 10.1113/jphysiol.1973.sp010388. PMID:4763992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gottschaldt KM, Vahle-Hinz C. Merkel cell receptors: structure and transducer function. Science. 1981;214(4517):183–186. doi: 10.1126/science.7280690. PMID:7280690. [DOI] [PubMed] [Google Scholar]
- Grim M, Halata Z. Developmental origin of avian Merkel cells. Anat Embryol (Berl) 2000;202(5):401–410. doi: 10.1007/s004290000121. PMID:11089931. [DOI] [PubMed] [Google Scholar]
- Haeberle H, Fujiwara M, Chuang J, Medina MM, Panditrao MV, Bechstedt S, Howard J, Lumpkin EA. Molecular profiling reveals synaptic release machinery in Merkel cells. Proc Natl Acad Sci USA. 2004;101(40):14503–14508. doi: 10.1073/pnas.0406308101. PMID:15448211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halata Z. The ultrastructure of the sensory nerve endings in the articular capsule of the knee joint of the domestic cat (Ruffini corpuscles and Pacinian corpuscles) J Anat. 1977;124(Pt 3):717–729. PMID: 604339. [PMC free article] [PubMed] [Google Scholar]
- Halata Z, Grim M, Bauman KI. Friedrich Sigmund Merkel and his “Merkel cell”, morphology, development, and physiology: review and new results. Anat Rec A Discov Mol Cell Evol Biol. 2003;271(1):225–239. doi: 10.1002/ar.a.10029. PMID:12552639. [DOI] [PubMed] [Google Scholar]
- Halata Z, Munger BL. The sensory innervation of primate eyelid. Anat Rec. 1980a;198(4):657–670. doi: 10.1002/ar.1091980410. PMID:7212315. [DOI] [PubMed] [Google Scholar]
- Halata Z, Munger BL. Sensory nerve endings in rhesus monkey sinus hairs. J Comp Neurol. 1980b;192(4):645–663. doi: 10.1002/cne.901920403. PMID:7419748. [DOI] [PubMed] [Google Scholar]
- Heidenreich M, Lechner SG, Vardanyan V, Wetzel C, Cremers CW, De Leenheer EM, Aránguez G, Moreno-Pelayo MÁ, Jentsch TJ, Lewin GR. KCNQ4 K(+) channels tune mechanoreceptors for normal touch sensation in mouse and man. Nat Neurosci. 2012;15(1):138–145. doi: 10.1038/nn.2985. PMID:22101641. [DOI] [PubMed] [Google Scholar]
- Hoggan G, Hoggan FE. Forked Nerve Endings on Hairs. J Anat Physiol. 1893;27(Pt 2):224–231. [PMC free article] [PubMed] [Google Scholar]
- Honma Y, Kawano M, Kohsaka S, Ogawa M. Axonal projections of mechanoreceptive dorsal root ganglion neurons depend on Ret. Development. 2010;137(14):2319–2328. doi: 10.1242/dev.046995. PMID:20534675. [DOI] [PubMed] [Google Scholar]
- Hoshino N, Harada F, Alkhamrah BA, Aita M, Kawano Y, Hanada K, Maeda T. Involvement of brain-derived neurotrophic factor (BDNF) in the development of periodontal Ruffini endings. Anat Rec A Discov Mol Cell Evol Biol. 2003;274(1):807–816. doi: 10.1002/ar.a.10094. PMID:12923891. [DOI] [PubMed] [Google Scholar]
- Hu J, Huang T, Li T, Guo Z, Cheng L. c-Maf is required for the development of dorsal horn laminae III/IV neurons and mechanoreceptive DRG axon projections. J Neurosci. 2012;32(16):5362–5373. doi: 10.1523/JNEUROSCI.6239-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hubbard SJ. A study of rapid mechanical events in a mechanoreceptor. J Physiol. 1958;141(2):198–218. doi: 10.1113/jphysiol.1958.sp005968. PMID:13539833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunt CC. On the nature of vibration receptors in the hind limb of the cat. J Physiol. 1961;155:175–186. doi: 10.1113/jphysiol.1961.sp006621. PMID:13716841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iggo A. Sensory receptors in the skin of mammals and their sensory functions. Rev Neurol (Paris) 1985;141(10):599–613. PMID:4089395. [PubMed] [Google Scholar]
- Iggo A, Andres KH. Morphology of cutaneous receptors. Annu Rev Neurosci. 1982;5(1):1–31. doi: 10.1146/annurev.ne.05.030182.000245. PMID:6280572. [DOI] [PubMed] [Google Scholar]
- Iggo A, Muir AR. The structure and function of a slowly adapting touch corpuscle in hairy skin. J Physiol. 1969;200(3):763–796. doi: 10.1113/jphysiol.1969.sp008721. PMID:4974746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iggo A, Ogawa H. Correlative physiological and morphological studies of rapidly adapting mechanoreceptors in cat’s glabrous skin. J Physiol. 1977;266(2):275–296. doi: 10.1113/jphysiol.1977.sp011768. PMID:853451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikeda I, Yamashita Y, Ono T, Ogawa H. Selective phototoxic destruction of rat Merkel cells abolishes responses of slowly adapting type I mechanoreceptor units. J Physiol. 1994;479(Pt 2):247–256. doi: 10.1113/jphysiol.1994.sp020292. PMID:7799224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johansson RS, Vallbo AB. Tactile sensibility in the human hand: relative and absolute densities of four types of mechanoreceptive units in glabrous skin. J Physiol. 1979;286:283–300. doi: 10.1113/jphysiol.1979.sp012619. PMID:439026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson KO. The roles and functions of cutaneous mechanoreceptors. Curr Opin Neurobiol. 2001;11(4):455–461. doi: 10.1016/s0959-4388(00)00234-8. PMID:11502392. [DOI] [PubMed] [Google Scholar]
- Johnson KO, Yoshioka T, Vega-Bermudez F. Tactile functions of mechanoreceptive afferents innervating the hand. J Clin Neurophysiol. 2000;17(6):539–558. doi: 10.1097/00004691-200011000-00002. PMID:11151974. [DOI] [PubMed] [Google Scholar]
- Kinkelin I, Stucky CL, Koltzenburg M. Postnatal loss of Merkel cells, but not of slowly adapting mechanoreceptors in mice lacking the neurotrophin receptor p75. Eur J Neurosci. 1999;11(11):3963–3969. doi: 10.1046/j.1460-9568.1999.00822.x. PMID:10583485. [DOI] [PubMed] [Google Scholar]
- Knibestöl M. Stimulus-response functions of rapidly adapting mechanoreceptors in human glabrous skin area. J Physiol. 1973;232(3):427–452. doi: 10.1113/jphysiol.1973.sp010279. PMID:4759677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kramer I, Sigrist M, de Nooij JC, Taniuchi I, Jessell TM, Arber S. A role for Runx transcription factor signaling in dorsal root ganglion sensory neuron diversification. Neuron. 2006;49(3):379–393. doi: 10.1016/j.neuron.2006.01.008. PMID:16446142. [DOI] [PubMed] [Google Scholar]
- Krimm RF, Davis BM, Noel T, Albers KM. Overexpression of neurotrophin 4 in skin enhances myelinated sensory endings but does not influence sensory neuron number. J Comp Neurol. 2006;498(4):455–465. doi: 10.1002/cne.21074. PMID:16937395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LeMaster AM, Krimm RF, Davis BM, Noel T, Forbes ME, Johnson JE, Albers KM. Overexpression of brain-derived neurotrophic factor enhances sensory innervation and selectively increases neuron number. J Neurosci. 1999;19(14):5919–5931. doi: 10.1523/JNEUROSCI.19-14-05919.1999. PMID:10407031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levanon D, Bettoun D, Harris-Cerruti C, Woolf E, Negreanu V, Eilam R, Bernstein Y, Goldenberg D, Xiao C, Fliegauf M, Kremer E, Otto F, Brenner O, Lev-Tov A, Groner Y. The Runx3 transcription factor regulates development and survival of TrkC dorsal root ganglia neurons. EMBO J. 2002;21(13):3454–3463. doi: 10.1093/emboj/cdf370. PMID:12093746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Rutlin M, Abraira VE, Cassidy C, Kus L, Gong S, Jankowski MP, Luo W, Heintz N, Koerber HR, Woodbury CJ, Ginty DD. The functional organization of cutaneous low-threshold mechanosensory neurons. Cell. 2011;147(7):1615–1627. doi: 10.1016/j.cell.2011.11.027. PMID:22196735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loewenstein WR, Mendelson M. Components of Receptor adaptation in a Pacinian corpuscle. J Physiol. 1965;177:377–397. doi: 10.1113/jphysiol.1965.sp007598. PMID:14321486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lou S, Duan B, Vong L, Lovell BB, Ma Q. Runx1 controls terminal morphology and mechanosensitivity of VGLUT3-expressing C-mechanoreceptors. J Neurosci. 2013;33(3):870–882. doi: 10.1523/JNEUROSCI.3942-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lucarz A, Brand G. Current considerations about Merkel cells. Eur J Cell Biol. 2007;86(5):243–251. doi: 10.1016/j.ejcb.2007.02.001. PMID:17337089. [DOI] [PubMed] [Google Scholar]
- Luo W, Enomoto H, Rice FL, Milbrandt J, Ginty DD. Molecular identification of rapidly adapting mechanoreceptors and their developmental dependence on ret signaling. Neuron. 2009;64(6):841–856. doi: 10.1016/j.neuron.2009.11.003. PMID:20064391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maklad A, Conway M, Hodges C, Hansen LA. Development of innervation to maxillary whiskers in mice. Anat Rec (Hoboken) 2010;293(9):1553–1567. doi: 10.1002/ar.21194. PMID:20648571. [DOI] [PubMed] [Google Scholar]
- Maksimovic S, Baba Y, Lumpkin EA. Neurotransmitters and synaptic components in the Merkel cell-neurite complex, a gentle-touch receptor. Ann N Y Acad Sci. 2013;1279(1):13–21. doi: 10.1111/nyas.12057. PMID:23530998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maricich SM, Morrison KM, Mathes EL, Brewer BM. Rodents rely on Merkel cells for texture discrimination tasks. J Neurosci. 2012;32(10):3296–3300. doi: 10.1523/JNEUROSCI.5307-11.2012. PMID:22399751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maricich SM, Wellnitz SA, Nelson AM, Lesniak DR, Gerling GJ, Lumpkin EA, Zoghbi HY. Merkel cells are essential for light-touch responses. Science. 2009;324(5934):1580–1582. doi: 10.1126/science.1172890. PMID:19541997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maruyama Y, Harada F, Jabbar S, Saito I, Aita M, Kawano Y, Suzuki A, Nozawa-Inoue K, Maeda T. Neurotrophin-4/5-depletion induces a delay in maturation of the periodontal Ruffini endings in mice. Arch Histol Cytol. 2005;68(4):267–288. doi: 10.1679/aohc.68.267. PMID:16477147. [DOI] [PubMed] [Google Scholar]
- Matsuo S, Ichikawa H, Silos-Santiago I, Kiyomiya K, Kurebe M, Arends JJ, Jacquin MF. Ruffini endings are absent from the periodontal ligament of trkB knockout mice. Somatosens Mot Res. 2002;19(3):213–217. doi: 10.1080/0899022021000009134. PMID:12396578. [DOI] [PubMed] [Google Scholar]
- Mendelson M, Lowenstein WR. Mechanisms of Receptor Adaptation. Science. 1964;144(3618):554–555. doi: 10.1126/science.144.3618.554. PMID:14194104. [DOI] [PubMed] [Google Scholar]
- Merkel F. Tastzellen and Tastkoerperchen bei den Hausthieren und beim Menschen. Arch Mikrosc Anat. 1875;11(S1):636–652. [Google Scholar]
- Montaño JA, Pérez-Piñera P, García-Suárez O, Cobo J, Vega JA. Development and neuronal dependence of cutaneous sensory nerve formations: Lessons from neurotrophins. Microsc Res Tech. 2010;73(5):513–529. doi: 10.1002/jemt.20790. PMID:19839059. [DOI] [PubMed] [Google Scholar]
- Morrison KM, Miesegaes GR, Lumpkin EA, Maricich SM. Mammalian Merkel cells are descended from the epidermal lineage. Dev Biol. 2009;336(1):76–83. doi: 10.1016/j.ydbio.2009.09.032. PMID:19782676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mosconi TM, Rice FL, Song MJ. Sensory innervation in the inner conical body of the vibrissal folliclesinus complex of the rat. J Comp Neurol. 1993;328(2):232–251. doi: 10.1002/cne.903280206. PMID:8423242. [DOI] [PubMed] [Google Scholar]
- Mountcastle VB. Modality and topographic properties of single neurons of cat’s somatic sensory cortex. J Neurophysiol. 1957;20(4):408–434. doi: 10.1152/jn.1957.20.4.408. PMID:13439410. [DOI] [PubMed] [Google Scholar]
- Munger BL, Ide C. The structure and function of cutaneous sensory receptors. Arch Histol Cytol. 1988;51(1):1–34. doi: 10.1679/aohc.51.1. PMID:3137944. [DOI] [PubMed] [Google Scholar]
- Nakamura S, Senzaki K, Yoshikawa M, Nishimura M, Inoue K, Ito Y, Ozaki S, Shiga T. Dynamic regulation of the expression of neurotrophin receptors by Runx3. Development. 2008;135(9):1703–1711. doi: 10.1242/dev.015248. PMID:18385258. [DOI] [PubMed] [Google Scholar]
- Ogawa H. The Merkel cell as a possible mechanoreceptor cell. Prog Neurobiol. 1996;49(4):317–334. doi: 10.1016/0301-0082(96)00018-4. PMID:8888113. [DOI] [PubMed] [Google Scholar]
- Palmer CI, Gardner EP. Simulation of motion on the skin. IV. Responses of Pacinian corpuscle afferents innervating the primate hand to stripe patterns on the OPTACON. J Neurophysiol. 1990;64(1):236–247. doi: 10.1152/jn.1990.64.1.236. PMID:2388068. [DOI] [PubMed] [Google Scholar]
- Paré M, Behets C, Cornu O. Paucity of presumptive ruffini corpuscles in the index finger pad of humans. J Comp Neurol. 2003;456(3):260–266. doi: 10.1002/cne.10519. PMID:12528190. [DOI] [PubMed] [Google Scholar]
- Paré M, Elde R, Mazurkiewicz JE, Smith AM, Rice FL. The Meissner corpuscle revised: a multiafferented mechanoreceptor with nociceptor immunochemical properties. J Neurosci. 2001;21(18):7236–7246. doi: 10.1523/JNEUROSCI.21-18-07236.2001. PMID:11549734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paré M, Smith AM, Rice FL. Distribution and terminal arborizations of cutaneous mechanoreceptors in the glabrous finger pads of the monkey. J Comp Neurol. 2002;445(4):347–359. doi: 10.1002/cne.10196. PMID:11920712. [DOI] [PubMed] [Google Scholar]
- Pease DC, Pallie W. Electron microscopy of digital tactile corpuscles and small cutaneous nerves. J Ultrastruct Res. 1959;2(3):352–365. doi: 10.1016/s0022-5320(59)80007-1. PMID:13655354. [DOI] [PubMed] [Google Scholar]
- Pease DC, Quilliam TA. Electron microscopy of the pacinian corpuscle. J Biophys Biochem Cytol. 1957;3(3):331–342. doi: 10.1083/jcb.3.3.331. PMID:13438918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez-Pinera P, García-Suarez O, Germanà A, Díaz-Esnal B, de Carlos F, Silos-Santiago I, del Valle ME, Cobo J, Vega JA. Characterization of sensory deficits in TrkB knockout mice. Neurosci Lett. 2008;433(1):43–47. doi: 10.1016/j.neulet.2007.12.035. PMID:18248898. [DOI] [PubMed] [Google Scholar]
- Peters EM, Botchkarev VA, Müller-Röver S, Moll I, Rice FL, Paus R. Developmental timing of hair follicle and dorsal skin innervation in mice. J Comp Neurol. 2002;448(1):28–52. doi: 10.1002/cne.10212. PMID:12012374. [DOI] [PubMed] [Google Scholar]
- Quilliam TA, Sato M. The distribution of myelin on nerve fibres from Pacinian corpuscles. J Physiol. 1955;129(1):167–176. doi: 10.1113/jphysiol.1955.sp005345. PMID:13252591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rasmusson DD, Turnbull BG. Sensory innervation of the raccoon forepaw: 2. Response properties and classification of slowly adapting fibers. Somatosens Res. 1986;4(1):63–75. doi: 10.3109/07367228609144598. PMID:3797915. [DOI] [PubMed] [Google Scholar]
- Rice FL, Rasmusson DD. Innervation of the digit on the forepaw of the raccoon. J Comp Neurol. 2000;417(4):467–490. doi: 10.1002/(sici)1096-9861(20000221)417:4<467::aid-cne6>3.0.co;2-q. PMID:10701867. [DOI] [PubMed] [Google Scholar]
- Sato M. Response of Pacinian corpuscles to sinusoidal vibration. J Physiol. 1961;159:391–409. doi: 10.1113/jphysiol.1961.sp006817. PMID:14497425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saxod R. Ontogeny of the cutaneous sensory organs. Microsc Res Tech. 1996;34(4):313–333. doi: 10.1002/(SICI)1097-0029(19960701)34:4<313::AID-JEMT4>3.0.CO;2-P. PMID:8807616. [DOI] [PubMed] [Google Scholar]
- Scheibel ME, Scheibel AB. Terminal axonal patterns in cat spinal cord. II. The dorsal horn. Brain Res. 1968;9(1):32–58. doi: 10.1016/0006-8993(68)90256-4. PMID:5699822. [DOI] [PubMed] [Google Scholar]
- Scheibert J, Leurent S, Prevost A, Debrégeas G. The role of fingerprints in the coding of tactile information probed with a biomimetic sensor. Science. 2009;323(5920):1503–1506. doi: 10.1126/science.1166467. PMID:19179493. [DOI] [PubMed] [Google Scholar]
- Scott A, Hasegawa H, Sakural K, Yaron A, Cobb J, Wang F. Transcription factor short stature homeobox 2 is required for proper development of tropomyosin-related kinase B-expressing mechanosensory neurons. J Neurosci. 2011;31(18):6741–6749. doi: 10.1523/JNEUROSCI.5883-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sedý J, Tseng S, Walro JM, Grim M, Kucera J. ETS transcription factor ER81 is required for the Pacinian corpuscle development. Dev Dyn. 2006;235(4):1081–1089. doi: 10.1002/dvdy.20710. PMID:16493690. [DOI] [PubMed] [Google Scholar]
- Semba K, Masarachia P, Malamed S, Jacquin M, Harris S, Egger MD. Ultrastructure of pacinian corpuscle primary afferent terminals in the cat spinal cord. Brain Res. 1984;302(1):135–150. doi: 10.1016/0006-8993(84)91293-9. PMID:6203612. [DOI] [PubMed] [Google Scholar]
- Semba K, Masarachia P, Malamed S, Jacquin M, Harris S, Yang G, Egger MD. An electron microscopic study of terminals of rapidly adapting mechanoreceptive afferent fibers in the cat spinal cord. J Comp Neurol. 1985;232(2):229–240. doi: 10.1002/cne.902320208. PMID:3973092. [DOI] [PubMed] [Google Scholar]
- Senok SS, Baumann KI. Functional evidence for calcium-induced calcium release in isolated rat vibrissal Merkel cell mechanoreceptors. J Physiol. 1997;500(Pt 1):29–37. doi: 10.1113/jphysiol.1997.sp021996. PMID:9097930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Senok SS, Baumann KI, Halata Z. Selective phototoxic destruction of quinacrine-loaded Merkel cells is neither selective nor complete. Exp Brain Res. 1996;110(3):325–334. doi: 10.1007/BF00229133. PMID:8871092. [DOI] [PubMed] [Google Scholar]
- Senzaki K, Ozaki S, Yoshikawa M, Ito Y, Shiga T. Runx3 is required for the specification of TrkC-expressing mechanoreceptive trigeminal ganglion neurons. Mol Cell Neurosci. 2010;43(3):296–307. doi: 10.1016/j.mcn.2009.12.003. PMID:20034568. [DOI] [PubMed] [Google Scholar]
- Shortland P, Woolf CJ. Morphology and somatotopy of the central arborizations of rapidly adapting glabrous skin afferents in the rat lumbar spinal cord. J Comp Neurol. 1993;329(4):491–511. doi: 10.1002/cne.903290406. PMID:8454737. [DOI] [PubMed] [Google Scholar]
- Skaper SD. The neurotrophin family of neurotrophic factors: an overview. Methods Mol Biol. 2012;846:1–12. doi: 10.1007/978-1-61779-536-7_1. PMID:22367796. [DOI] [PubMed] [Google Scholar]
- Suzuki M, Ebara S, Koike T, Tonomura S, Kumamoto K. How many hair follicles are innervated by one afferent axon? A confocal microscopic analysis of palisade endings in the auricular skin of thy1-YFP transgenic mouse. Proc Jpn Acad, Ser B, Phys Biol Sci. 2012;88(10):583–595. doi: 10.2183/pjab.88.583. PMID:23229751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szeder V, Grim M, Halata Z, Sieber-Blum M. Neural crest origin of mammalian Merkel cells. Dev Biol. 2003;253(2):258–263. doi: 10.1016/s0012-1606(02)00015-5. PMID:12645929. [DOI] [PubMed] [Google Scholar]
- Tachibana T, Nawa T. Recent progress in studies on Merkel cell biology. Anat Sci Int. 2002;77(1):26–33. doi: 10.1046/j.0022-7722.2002.00008.x. PMID:12418081. [DOI] [PubMed] [Google Scholar]
- Takahashi-Iwanaga H. Three-dimensional microanatomy of longitudinal lanceolate endings in rat vibrissae. J Comp Neurol. 2000;426(2):259–269. doi: 10.1002/1096-9861(20001016)426:2<259::aid-cne7>3.0.co;2-n. PMID:10982467. [DOI] [PubMed] [Google Scholar]
- Talbot WH, Darian-Smith I, Kornhuber HH, Mountcastle VB. The sense of flutter-vibration: comparison of the human capacity with response patterns of mechanoreceptive afferents from the monkey hand. J Neurophysiol. 1968;31(2):301–334. doi: 10.1152/jn.1968.31.2.301. PMID:4972033. [DOI] [PubMed] [Google Scholar]
- Tapper DN. Stimulus-response relationships in the cutaneous slowly-adapting mechanoreceptor in hairy skin of the cat. Exp Neurol. 1965;13(4):364–385. doi: 10.1016/0014-4886(65)90125-1. PMID:5847283. [DOI] [PubMed] [Google Scholar]
- Wellnitz SA, Lesniak DR, Gerling GJ, Lumpkin EA. The regularity of sustained firing reveals two populations of slowly adapting touch receptors in mouse hairy skin. J Neurophysiol. 2010;103(6):3378–3388. doi: 10.1152/jn.00810.2009. PMID:20393068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wende H, Lechner SG, Cheret C, Bourane S, Kolanczyk ME, Pattyn A, Reuter K, Munier FL, Carroll P, Lewin GR, Birchmeier C. The transcription factor c-Maf controls touch receptor development and function. Science. 2012;335(6074):1373–1376. doi: 10.1126/science.1214314. PMID:22345400. [DOI] [PubMed] [Google Scholar]
- Willis WD, Coggeshall RE. Sensory Mechanisms of the Spinal Cord. New York: Kluwer Academic/Plenum Publishers; 2004. [Google Scholar]
- Winkelmann RK, Breathnach AS. The Merkel cell. J Invest Dermatol. 1973;60(1):2–15. doi: 10.1111/1523-1747.ep13069480. PMID:4346159. [DOI] [PubMed] [Google Scholar]
- Woo SH, Baba Y, Franco AM, Lumpkin EA, Owens DM. Excitatory glutamate is essential for development and maintenance of the piloneural mechanoreceptor. Development. 2012;139(4):740–748. doi: 10.1242/dev.070847. PMID:22241839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woodbury CJ, Ritter AM, Koerber HR. Central anatomy of individual rapidly adapting low-threshold mechanoreceptors innervating the “hairy” skin of newborn mice: early maturation of hair follicle afferents. J Comp Neurol. 2001;436(3):304–323. PMID:11438932. [PubMed] [Google Scholar]
- Yamashita Y, Akaike N, Wakamori M, Ikeda I, Ogawa H. Voltage-dependent currents in isolated single Merkel cells of rats. J Physiol. 1992;450:143–162. doi: 10.1113/jphysiol.1992.sp019120. PMID:1331421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshikawa M, Murakami Y, Senzaki K, Masuda T, Ozaki S, Ito Y, Shiga T. Coexpression of Runx1 and Runx3 in mechanoreceptive dorsal root ganglion neurons. Dev Neurobiol. 2013;73(6):469–479. doi: 10.1002/dneu.22073. PMID:23378040. [DOI] [PubMed] [Google Scholar]
- Zelená J. The development of Pacinian corpuscles. J Neurocytol. 1978;7(1):71–91. doi: 10.1007/BF01213461. PMID:632855. [DOI] [PubMed] [Google Scholar]
- Zelena J. Nerves and Mechanoreceptors. London: Chapman & Hall; 1994. [Google Scholar]
- Zelená J, Halata Z, Szeder V, Grim M. Crural Herbst corpuscles in chicken and quail: numbers and structure. Anat Embryol (Berl) 1997;196(4):323–333. doi: 10.1007/s004290050101. PMID:9363854. [DOI] [PubMed] [Google Scholar]
- Zelená J, Jirmanová I, Nitatori T, Ide C. Effacement and regeneration of tactile lamellar corpuscles of rat after postnatal nerve crush. Neuroscience. 1990;39(2):513–522. doi: 10.1016/0306-4522(90)90287-e. PMID:2087271. [DOI] [PubMed] [Google Scholar]