

Abstract
Purpose
Pain from vertebral or femoral neck fractures is a particularly important problem in clinical orthopaedics. Transient receptor potential vanilloid 1 (TRPV1) is a ligand-gated nonselective cation channel, and there are recent reports on an association between bone pain and TRPV1. However, an increase in TRPV1 activity has not been reported following femoral fracture.
Materials and Methods
We applied a neurotracer [Fluoro-gold (FG)] onto femur to detect dorsal root ganglia (DRGs) innervating the cortex of the femur in 30 Sprague Dawley rats. Seven days after application, a closed mid-diaphyseal fracture of the femur was performed. FG labeled TRPV1-immunoreactive (ir) DRGs innervating the femur were examined in nonfractured controls, and 3 days, 1 week, 2 weeks, and 4 weeks after fracture. We evaluated bone healing of the femur and compared the ratio of TRPV1-ir DRG neurons innervating the femur at the time points.
Results
Four weeks after fracture, complete bone union was observed. There was no significant difference in the ratio of FG labeled DRG neurons to total DRG neurons at each time point. The percentages of TRPV1-ir neurons in DRGs innervating the femur at 3 days and 1 week after fracture were significantly higher than those in control, 2 weeks, and 4 weeks after fracture (p<0.05).
Conclusion
Fracture induced an increase of TRPV1-ir neurons in DRGs innervating the fractured femur within 3 days, and decreased during bone healing over 4 weeks. These findings show that TRPV1 may play a role in sensory sensation of bone fracture pain.
Keywords: Fracture, pain, transient receptor potential vanilloid 1, femur, rat
INTRODUCTION
Although skeletal pain plays a major role in reducing the quality of life in patients suffering from osteoporosis, osteoarthritis, fracture and bone cancer, little is known about the mechanisms that generate and maintain this pain.
Elderly patients with thoracic and lumbar vertebral fractures caused by osteoporosis often experience back pain, and femoral neck fractures cause severe hip pain. These fractures decrease activities of daily living in patients. Osteoporotic fractures represent a significant public health burden, which will increase in future generations. Lifetime risk is high and lies within the range of 40% to 50% in women and 13% to 22% in men.1 These demographic findings suggest that the number of hip fractures occurring worldwide will increase from 1.66 million in 1990 to 6.26 million in 2050.1 However, the mechanisms for the pain associated with bone fractures are not clear.
Animal models using the femur or tibia have been used to study pain pathways in bone.2-9 Normal pain mechanisms in the femur and tibia, and those associated with aging, inflammation, fracture, and cancer have been explored.2-9 Several authors have studied pain mechanisms using animal fracture models. Pain-related behaviors after a closed femur fracture in mouse showed mechanical hypersensitivity two weeks after fracture, with recovery from pain during bone healing.7,8 A monoclonal antibody raised against nerve growth factor resulted in a reduction of approximately 50% in pain-related behaviors following fracture.7,8 Yasui, et al.9 have reported mechanical hyperalgesia within two weeks using a rat bone injury model with drilling through the tibia, without casting or pin insertion. Anti-nerve growth factor inhibited the hyperalgesia.9 In the normal mouse femur, thinly myelinated and unmyelinated painful peptidergic sensory fibers were observed in mineralized bone, bone marrow, and periosteum of the femur.2 With pain following fracture, nerve growth factor-dependent sensory and sympathetic nerve fibers undergo sprouting and the number of nerve fibers increases.9
Transient receptor potential vanilloid 1 (TRPV1) is a ligand-gated nonselective cation channel, which can be activated by capsaicin and other stimulation such as noxious heat and low pH.10 TRPV1 has attracted attention in acute inflammatory nociception research, and its role in chronic inflammatory pain has begun to attract more interest.11,12 A recent study showed that TRPV1 was involved in bone cancer pain.13 A TRPV1 antagonist significantly reduced the pain and improved weight-bearing in a murine model of bone cancer pain.14 However, there have been no reports that fracture influences TRPV1 in the sensory system innervating bone.
The purpose of the current study was to investigate the existence and the change in TRPV1-immunoreactive (ir) dorsal root ganglia (DRGs), innervating the femur of rats and labeled with neurotracer [(Fluoro-gold (FG)], in nonfractured controls, and 3 days, 1 week, 2 weeks, and 4 weeks after fracture.
MATERIALS AND METHODS
Fracture model
Sprague Dawley female rats weighing 200-300 g were housed in the animal unit at 21℃ on a 12-hr light/dark cycle (08:30-20:30 hr) and fed a pellet diet and tap water ad libitum. Experiments were performed with permission from the ethics committee of the Graduate School of Medicine, Chiba University, following the National Institutes of Health guidelines for the care and use of laboratory animals (1996 revision).
Fracture model was made according to previous report.6 Animals were anesthetized with sodium pentobarbital (40 mg/kg, i.p.) and treated aseptically throughout the experiments. An incision of approximately 6 mm was made in the skin, then the proximal patellar ligament of the left femur was severed revealing the synovial space of the knee joint. A 20-gauge needle was used to core between the condyles and into the medullary canal of the left femur. Rats were immediately radiographed to ensure proper medullary coring. A pre-cut 0.8 mm diameter (27 mm long) stainless steel wire pin was inserted into the medullary space. After this procedure, the mid-diaphysis of the femur was fractured, and the skin was closed. Thirty rats were used in this experiment (control, 3 days, 1 week, 2 weeks, and 4 weeks, n=6, each). We confirmed fracture and bone union radiographically.
Retrograde FluoroGold labeling for detection of DRG neurons innervating left femur fractures
Animals were anesthetized with sodium pentobarbital (40 mg/kg, i.p.) and treated aseptically throughout the experiments. For each animal, a longitudinal incision was made and the left femur was exposed under a microscope in control rats and before fracture in the experimental groups. The neurotracer Fluoro-gold (FG; Fluorochrome, Denver, CO, USA) was applied to the mid-diaphysis of the cortex of the left femur in all 30 rats to label DRG neurons innervating the left femur. Small dimple on the surface of cortex was made by 21-gauge needle, and FG was applied. The dimple was immediately sealed with cyanoacrylate adhesive to prevent leakage of FG and the skin was closed.
Immunohistochemistry for TRPV1
In controls and for each time point studied following fracture, rats were anesthetized with sodium pentobarbital (40 mg/kg, i.p.) and transcardially perfused with 0.9% saline, followed by 500 mL of 4% paraformaldehyde in phosphate buffer (0.1 M, pH 7.4). Next, left DRGs from the L1 to L6 levels were resected, and the specimens were immersed in the buffered paraformaldehyde fixative solution overnight at 4℃. After storing in 0.01 M phosphate buffered saline (PBS) containing 20% sucrose for 20 hours at 4℃, each ganglion was sectioned at 10 µm thickness on a cryostat and mounted on poly-L-lysine-coated slides. Endogenous tissue peroxidase activity was quenched by soaking the sections in 0.3% hydrogen peroxide solution in 0.01 M PBS for 30 minutes. The specimens were then treated for 90 minutes at room temperature in blocking solution consisting of 0.01 M PBS containing 0.3% Triton X-100 and 3% skim milk. To stain-DRGs, the sections were processed for TRPV1 immunohistochemistry using a rabbit antibody to TRPV1 (1 : 500; Calbiochem, San Diego, CA, USA) diluted in blocking solution, and incubated for 20 hours at 4℃. Sections were incubated with goat anti-rabbit Alexa 488 fluorescent antibody conjugate for visualization (1 : 400; Molecular Probes, Eugene, OR, USA). After each step, the sections were rinsed three times in 0.01 M PBS and examined using a fluorescence microscope. The total number of DRG neurons, number of FG-labeled neurons, and FG-labeled and TRPV1-ir neurons were counted in a 0.45 mm2 region of interest at 400× magnification using a counting grid.
Statistical analysis
The number of FG-labeled and FG-labeled TRPV1-ir DRG neurons as a proportion of total DRG neurons in each DRG were compared using Welch's unpaired t test. The distribution of the FG-labeled DRG neurons and FG-labeled TRPV1-ir DRG neurons was analyzed using a non-repeated measures ANOVA with Bonferroni post hoc correction. p<0.05 was considered statistically significant. Results are reported as means±SEMs.
RESULTS
Bone fracture and healing
Radiographs showed the course of fracture healing over the four-week experimental period. All rats showed complete bone union four weeks after fracture (Fig. 1).
Fig. 1.
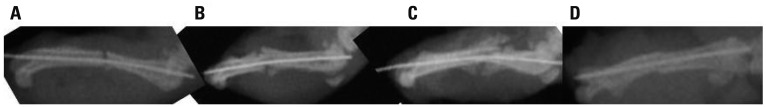
Radiographs showing at 3 days, 1 week, 2 weeks, and 4 weeks after fracture. (A) 3 days, (B) 1 week, (C) 2 weeks, and (D) 4 weeks.
FG-labeled DRG neurons innervating the left femur
FG-labeled DRG neurons, indicating where FG was transported from the femur, were present in the DRG from L1 through L6 (Figs. 2 and 3). There was no significant difference in percentage of FG-labeled neurons of all DRG neurons from L1 to L6 among groups (control, 3 days, 1 week, 2 weeks, and 4 weeks after fracture) (Fig. 3).
Fig. 2.

Innervation of the left femur at 2 weeks. FG-labeled (A) and TRPV1 immunoreactive DRG neurons (B) (A and B are the same sections. Arrows indicate double labeled neurons). FG, Fluoro-gold; TRPV1, transient receptor potential vanilloid 1; DRG, dorsal root ganglia.
Fig. 3.

Distribution and ratio of FG-labeled DRG neurons innervating the left femur in control, 3 days, 1 week, 2 weeks, and 4 weeks groups. FG-labeled DRG neurons are present from L1 through L6, and there was no significant difference in the percentage of FG-labeled neurons of all DRG neurons among control, 3 days, 1 week, 2 weeks, and 4 weeks groups. FG, Fluoro-gold; DRG, dorsal root ganglion.
FG-labeled and TRPV1-ir DRG neurons innervating the left femur
FG-labeled and TRPV1-ir DRG neurons innervating the left femur were present in the DRG from L1 through L6 (Figs. 2 and 4). Of the total DRG neurons innervating the femur, the ratio of FG labeled TRPV1-ir DRG neurons was 1.7±0.3%, 3.0±0.6%, 2.6±0.4%, 1.9±0.3%, and 1.6±0.2% (means±SEMs) in nonfractured controls, and 3 days, 1 week, 2 weeks, and 4 weeks after fracture, respectively. The percentages of TRPV1-ir neurons in the DRGs innervating the femur at 3 days and 1 week after fracture were significantly greater than those in nonfractured controls, and 2 weeks and 4 weeks after fracture (p<0.05) (Fig. 4).
Fig. 4.

Distribution and ratio of FG-labeled and TRPV1-ir DRG neurons innervating the left femur among control, 3 days, 1 week, 2 weeks, and 4 weeks groups. FG-labeled and TRPV1-ir DRG neurons innervating the left femur are present from L1 through L6. The percentages of TRPV1-ir neurons in the DRGs innervating the femur at 3 days and 1 week after fracture were significantly higher than those in nonfractured control, and 2 weeks and 4 weeks after fracture (p<0.05). FG, Fluoro-gold; TRPV1, transient receptor potential vanilloid 1; DRG, dorsal root ganglion; ir, immunoreactive.
DISCUSSION
In this study, of total DRG neurons innervating the femur, the ratio of FG labeled TRPV1-ir DRG neurons was 1.7%, 3.0%, 2.6%, 1.9%, and 1.6% in nonfractured controls, and 3 days, 1 week, 2 weeks, and 4 weeks after fracture, respectively. The percentages of TRPV1-ir neurons in DRGs innervating the femur at 3 days and 1 week after fracture were significantly higher than those in control, 2 weeks, and 4 weeks after fracture. Thus, we speculated that TRPV1 plays an important role in sensory sensation from the fracture of femur.
In the current study, the proportion of TRPV1-ir DRG neurons innervating the fractured femur in rat increased compared to the normal femur. Several authors have reported a relationship between TRPV1 and pain transmission from bone disease. In the vertebral body harvested from patients with back pain, many sensory nerves show immunoreactivity for neuropeptides involved in pain transmission, such as substance P (SP) and calcitonin gene related peptide (CGRP); SP-ir and CGRP-ir nociceptors have also been observed.15 In the mouse femur, CGRP-ir fibers associated with pain due to inflammation are present throughout the bone marrow, mineralized bone and periosteum, and predominantly in bone marrow.2 Orita, et al.16 have reported the existence of TRPV1 and CGRP-ir sensory nerve fibers in rat vertebral body. TRPV1 and CGRP expressions are elevated in osteoporotic vertebrae and induce pain in rat. TRPV1 has been reported to modulate the synthesis and release of CGRP in sensory nerves by its activation.17 Neonatal capsaicin treatment results in a significant depletion in the density of CGRP positive sensory nerve fibers in the periosteum of the femur,18 and there was a 50% reduction in the severity of pain from the fractured femur, when treated with capsaicin,18 suggesting that capsaicin-sensitive CGRP positive sensory nerve fibers participate in driving skeletal fracture pain.18 These reports together suggest that neuropeptides such as SP and CGRP, and TRPV1 modulate bone pain originating from fracture.
A relationship between TRPV1 and bone remodeling, independent of pain, has been reported. In an animal heterotopic ossification model, bone morphogenetic proteins induced the release of neuroinflammatory molecules wuch as SP and CGRP from peripheral sensory nerve fibers.19 In animals lacking TRPV1, bone morphogenetic proteins mediating increases of SP and CGRP were suppressed and ossification was dramatically inhibited, suggesting that TRPV1 plays a functional role in bone formation.19 Furthermore, TRPV1 regulates human osteoclast and osteoblast formation;20 TRPV1 antagonist inhibits ovariectomy-induced bone loss in mice and histomorphometric analysis shows inhibitory effects on indices of bone resorption and bone formation. In the current study, we focused on the relationship between pain and bone fracture, but there is a possibility that TRPV1 plays an important role in remodeling after bone fracture. On the other hand, however, the percentages of TRPV1-ir neurons in DRGs innervating the femur at 3 days and 1 week after fracture were significantly higher than those in control, 2 weeks, and 4 weeks after fracture. Therefore, it is highly possible that TRPV1 plays an important role in sensory sensation in acute phase rather than remodeling after bone fracture.
What kinds of therapies are currently available to treat bone fracture pain? Non-steroidal anti-inflammatory drugs (NSAIDs) and opioids are currently available for pain treatment after fracture. However, some authors have reported an inhibitory effect of NSAIDs on bone union in both animals and humans. Murnaghan, et al.21 reported that NSAIDs have a significantly negative effect on blood flow across the fracture gap as well as an inhibitory effect on fracture repair in mice, and concluded that NSAIDs should be used with caution in all patients following osseous trauma. In human fracture of the femoral diaphysis, there was a marked association between nonunion and the use of NSAIDs after injury and delayed healing was noted in patients who took NSAIDs.22 Opioids cause increased somnolence, agitation, constipation, dizziness and cognitive impairment which can reduce mobility. Community-dwelling older women taking opioids have an increased risk for any nonspine fracture including hip fracture,23 the ability to participate in physical rehabilitation is decreased, resulting in loss of bone and/or muscle mass, which further results in delayed bone healing.23 King, et al.24 reported the inhibitory effect of opioids for bone union in an animal cancer model; morphine did not alter tumor growth or tumor burden but accelerated sarcoma-induced bone destruction and doubled the incidence of spontaneous fractures in a dose-dependent manner. Furthermore, morphine increased osteoclast activity, suggesting enhancement of sarcoma-induced osteolysis.24 In the future, therefore, antagonists of TRPV1 may become a therapeutic target for pain after bone fracture.
There are some limitations in the current study. We demonstrated that TRPV1-ir DRG neurons innervated the femur. However, we did not directly examine the sensory innervation of femur or the function of TRPV1. Furthermore, we did not evaluate pain originating from fracture, because this has already been shown previously in the same model as we used.6-8 We rather focused on the relation between pain and bone fracture, and did not examine the role of TRPV1 in bone remodeling. Further study is needed to clarify the pain mechanism of TRPV1 and pain transmission from fractured bone.
In conclusion, the present study demonstrated that the ratio of TRPV1-ir DRG neurons innervating the fractured rat femur was higher than those in normal femur and in healed femur, suggesting that TRPV1 may modulate sensory transmission from the fractured bone.
Footnotes
The authors have no financial conflicts of interest.
References
- 1.Dennison E, Mohamed MA, Cooper C. Epidemiology of osteoporosis. Rheum Dis Clin North Am. 2006;32:617–629. doi: 10.1016/j.rdc.2006.08.003. [DOI] [PubMed] [Google Scholar]
- 2.Mach DB, Rogers SD, Sabino MC, Luger NM, Schwei MJ, Pomonis JD, et al. Origins of skeletal pain: sensory and sympathetic innervation of the mouse femur. Neuroscience. 2002;113:155–166. doi: 10.1016/s0306-4522(02)00165-3. [DOI] [PubMed] [Google Scholar]
- 3.Mantyh WG, Jimenez-Andrade JM, Stake JI, Bloom AP, Kaczmarska MJ, Taylor RN, et al. Blockade of nerve sprouting and neuroma formation markedly attenuates the development of late stage cancer pain. Neuroscience. 2010;171:588–598. doi: 10.1016/j.neuroscience.2010.08.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Yang CJ, Wang XW, Li X, Wu GC, Wang YQ, Mao-Ying QL. A rat model of bone inflammation-induced pain by intra-tibial complete Freund's adjuvant injection. Neurosci Lett. 2011;490:175–179. doi: 10.1016/j.neulet.2010.12.027. [DOI] [PubMed] [Google Scholar]
- 5.Jimenez-Andrade JM, Mantyh WG, Bloom AP, Freeman KT, Ghilardi JR, Kuskowski MA, et al. The effect of aging on the density of the sensory nerve fiber innervation of bone and acute skeletal pain. Neurobiol Aging. 2012;33:921–932. doi: 10.1016/j.neurobiolaging.2010.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Freeman KT, Koewler NJ, Jimenez-Andrade JM, Buus RJ, Herrera MB, Martin CD, et al. A fracture pain model in the rat: adaptation of a closed femur fracture model to study skeletal pain. Anesthesiology. 2008;108:473–483. doi: 10.1097/ALN.0b013e3181649351. [DOI] [PubMed] [Google Scholar]
- 7.Koewler NJ, Freeman KT, Buus RJ, Herrera MB, Jimenez-Andrade JM, Ghilardi JR, et al. Effects of a monoclonal antibody raised against nerve growth factor on skeletal pain and bone healing after fracture of the C57BL/6J mouse femur. J Bone Miner Res. 2007;22:1732–1742. doi: 10.1359/jbmr.070711. [DOI] [PubMed] [Google Scholar]
- 8.Jimenez-Andrade JM, Martin CD, Koewler NJ, Freeman KT, Sullivan LJ, Halvorson KG, et al. Nerve growth factor sequestering therapy attenuates non-malignant skeletal pain following fracture. Pain. 2007;133:183–196. doi: 10.1016/j.pain.2007.06.016. [DOI] [PubMed] [Google Scholar]
- 9.Yasui M, Shiraishi Y, Ozaki N, Hayashi K, Hori K, Ichiyanagi M, et al. Nerve growth factor and associated nerve sprouting contribute to local mechanical hyperalgesia in a rat model of bone injury. Eur J Pain. 2012;16:953–965. doi: 10.1002/j.1532-2149.2011.00094.x. [DOI] [PubMed] [Google Scholar]
- 10.Tominaga M, Caterina MJ, Malmberg AB, Rosen TA, Gilbert H, Skinner K, et al. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron. 1998;21:531–543. doi: 10.1016/s0896-6273(00)80564-4. [DOI] [PubMed] [Google Scholar]
- 11.Guo A, Vulchanova L, Wang J, Li X, Elde R. Immunocytochemical localization of the vanilloid receptor 1 (VR1): relationship to neuropeptides, the P2X3 purinoceptor and IB4 binding sites. Eur J Neurosci. 1999;11:946–958. doi: 10.1046/j.1460-9568.1999.00503.x. [DOI] [PubMed] [Google Scholar]
- 12.Yu L, Yang F, Luo H, Liu FY, Han JS, Xing GG, et al. The role of TRPV1 in different subtypes of dorsal root ganglion neurons in rat chronic inflammatory nociception induced by complete Freund's adjuvant. Mol Pain. 2008;4:61. doi: 10.1186/1744-8069-4-61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Niiyama Y, Kawamata T, Yamamoto J, Omote K, Namiki A. Bone cancer increases transient receptor potential vanilloid subfamily 1 expression within distinct subpopulations of dorsal root ganglion neurons. Neuroscience. 2007;148:560–572. doi: 10.1016/j.neuroscience.2007.05.049. [DOI] [PubMed] [Google Scholar]
- 14.Niiyama Y, Kawamata T, Yamamoto J, Furuse S, Namiki A. SB366791, a TRPV1 antagonist, potentiates analgesic effects of systemic morphine in a murine model of bone cancer pain. Br J Anaesth. 2009;102:251–258. doi: 10.1093/bja/aen347. [DOI] [PubMed] [Google Scholar]
- 15.Brown MF, Hukkanen MV, McCarthy ID, Redfern DR, Batten JJ, Crock HV, et al. Sensory and sympathetic innervation of the vertebral endplate in patients with degenerative disc disease. J Bone Joint Surg Br. 1997;79:147–153. doi: 10.1302/0301-620x.79b1.6814. [DOI] [PubMed] [Google Scholar]
- 16.Orita S, Ohtori S, Koshi T, Yamashita M, Yamauchi K, Inoue G, et al. The effects of risedronate and exercise on osteoporotic lumbar rat vertebrae and their sensory innervation. Spine (Phila Pa 1976) 2010;35:1974–1982. doi: 10.1097/BRS.0b013e3181d5959e. [DOI] [PubMed] [Google Scholar]
- 17.Luo D, Zhang YW, Peng WJ, Peng J, Chen QQ, Li D, et al. Transient receptor potential vanilloid 1-mediated expression and secretion of endothelial cell-derived calcitonin gene-related peptide. Regul Pept. 2008;150:66–72. doi: 10.1016/j.regpep.2008.05.007. [DOI] [PubMed] [Google Scholar]
- 18.Jimenez-Andrade JM, Bloom AP, Mantyh WG, Koewler NJ, Freeman KT, Delong D, et al. Capsaicin-sensitive sensory nerve fibers contribute to the generation and maintenance of skeletal fracture pain. Neuroscience. 2009;162:1244–1254. doi: 10.1016/j.neuroscience.2009.05.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Salisbury E, Rodenberg E, Sonnet C, Hipp J, Gannon FH, Vadakkan TJ, et al. Sensory nerve induced inflammation contributes to heterotopic ossification. J Cell Biochem. 2011;112:2748–2758. doi: 10.1002/jcb.23225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Idris AI, Landao-Bassonga E, Ralston SH. The TRPV1 ion channel antagonist capsazepine inhibits osteoclast and osteoblast differentiation in vitro and ovariectomy induced bone loss in vivo. Bone. 2010;46:1089–1099. doi: 10.1016/j.bone.2010.01.368. [DOI] [PubMed] [Google Scholar]
- 21.Murnaghan M, Li G, Marsh DR. Nonsteroidal anti-inflammatory drug-induced fracture nonunion: an inhibition of angiogenesis? J Bone Joint Surg Am. 2006;88(Suppl 3):140–147. doi: 10.2106/JBJS.F.00454. [DOI] [PubMed] [Google Scholar]
- 22.Giannoudis PV, MacDonald DA, Matthews SJ, Smith RM, Furlong AJ, De Boer P. Nonunion of the femoral diaphysis. The influence of reaming and non-steroidal anti-inflammatory drugs. J Bone Joint Surg Br. 2000;82:655–658. doi: 10.1302/0301-620x.82b5.9899. [DOI] [PubMed] [Google Scholar]
- 23.Ensrud KE, Blackwell T, Mangione CM, Bowman PJ, Bauer DC, Schwartz A, et al. Central nervous system active medications and risk for fractures in older women. Arch Intern Med. 2003;163:949–957. doi: 10.1001/archinte.163.8.949. [DOI] [PubMed] [Google Scholar]
- 24.King T, Vardanyan A, Majuta L, Melemedjian O, Nagle R, Cress AE, et al. Morphine treatment accelerates sarcoma-induced bone pain, bone loss, and spontaneous fracture in a murine model of bone cancer. Pain. 2007;132:154–168. doi: 10.1016/j.pain.2007.06.026. [DOI] [PMC free article] [PubMed] [Google Scholar]