

Abstract
Context. Heart failure (HF) is a common condition causing much morbidity and mortality despite major advances in pharmacological and device therapies. Preclinical data suggest a cardioprotective role of Angiotensin-(1-7) in animal models of HF. Objective. Perform a systematic review on the effects of Angiotensin-(1-7) on humans, focusing on HF. Results. 39 studies were included in the review (4 in human HF and (35) in non-HF patients). There is only one intervention study on 8 patients with human HF, using Angiotensin-(1-7), with forearm blood flow (FBF) as the endpoint. Angiotensin-(1-7) caused no significant effect on FBF in this HF study but caused vasodilation in 3 out of 4 non-HF studies. In one other non-HF study, Angiotensin-(1-7) infusion led to a significant increase in blood pressure in normal men; however, effects were <0.03% that of angiotensin II. Cardioprotective effects seen in non-HF studies include for instance beneficial actions against atherosclerosis and myocardial fibrosis. Conclusions. The main finding of our systematic review is that Angiotensin-(1-7) plays an important cardioprotective role in HF in animals and in patients without heart failure. More research is required to test the hypothesis that Angiotensin-(1-7) benefits patients with heart failure.
1. Introduction
Heart failure (HF) is a major health problem for many developed world populations and has a relatively poor prognosis. Within the US population, the incidence approaches 1%, with a lifetime risk of 1 in 5 for both men and women at the age of 40 years [1]. In 2005, there was an estimated near 1.1 million admissions to American hospitals associated with HF, which was up from approximately 400,000 in 1979 [2]. This substantial increase in prevalence and hospitalisation meant an epidemic was declared [3, 4]. Over the coming years, the prevalence is likely to increase due to changing lifestyles and diets of the developing world, advances in heart failure therapeutics, an increase in prevalence of conditions which have cardiovascular consequences such as obesity and diabetes, and better survival from other heart conditions where HF is the end stage especially with the increased use of primary angioplasty for myocardial infarction. Figures from the United States show an estimated bill totalling $39.2 billion in 2010, to cover the direct and indirect cost of HF [5]. One in 8 death certificates in America mentioned HF, and in 20% of cases, it was the primary cause. In 2006, the number of any-mention deaths from HF was 283,000 [1]. Five-year mortality is 45–60% [6], and hospitalisations increased in the months prior to their death [7]; moreover, after the first hospitalisation, the 5-year mortality was greater than 75% [8]. This deadly syndrome, where the renin-angiotensin-aldosterone system (RAAS) is of great importance, is characterised by dyspnoea, venous congestion, and oedema, the consequences of anatomical and functional defects within the heart.
1.1. Angiotensin-(1-7): The Peptide
This bioactive endogenous heptapeptide is taking increasing amounts of interest from investigators because of its potential as a therapeutic agent. Angiotensin-(1-7) (Ang-(1-7)) is becoming recognised as an increasingly important player of the RAAS where accumulating evidence suggests that it may have a key role in the regulation and homeostasis of the RAAS, exerting cardiovascular protection. Ang-(1-7) is different from angiotensin II due to the absence of an amino acid at position 8, described as the most pleiotropic metabolite of angiotensin I (Ang I), and is known to have actions which on most occasions are opposite (but may be identical) of those known for angiotensin II (Ang II) [9, 10]. Many effects of Ang-(1-7) are mediated via the high-affinity G protein-coupled receptor Mas, where Ang-(1-7) is an endogenous ligand [11]. Endopeptidases (such as neutral endopeptidase (NEP)) and angiotensin-converting enzyme-2 (ACE2) contribute to the alternative pathways of Ang-(1-7) generation, using the substrates angiotensin I (Ang I) and angiotensin II (Ang II), respectively [12–14], whereas it has been shown that Ang-(1-7) is degraded to angiotensin- 1-5 (Ang-(1-5)) by angiotensin-converting enzyme (ACE) [15]. In contrast, ACE has also been shown to generate Ang-(1-7) from angiotensin-(1-9) [16]. Ang-(1-7) studies have shown that it is a potential endogenous counterregulator of the RAAS cascade [15, 17]. Physiological concentration of Ang-(1-7) in man is thought to be 10 pmol/L [18, 19]. In 1988, Schiavone et al. suggested that Ang-(1-7) peptide was indeed biologically active [20], raising the possibility of potential benefits of Ang-(1-7).
1.2. Preclinical Studies
There has been an abundance of animal studies investigating the effects of Ang-(1-7) in a variety of environments. Increased activation of the RAAS in heart failure is a compensatory mechanism. However, this increased afterload can have adverse effects on cardiac function. Angiotensin converting enzyme inhibitors are commonly seen in the management plan of HF; one reason is to reduce arterial pressure and to reduce the levels of Ang II. Vasodilatation is a property of Ang-(1-7) [21]. It does so by various postulated mechanisms such as releasing nitric oxide (NO) and prostaglandins [22–25]. Both Brosnihan et al. [26] and Porsti et al. [27], by administering NO synthase inhibitors, suggested that the vasodilatory effect of Ang-(1-7) may be at least partly dependent on the release of NO. Furthermore, Benter et al.'s work would suggest that Ang-(1-7) might oppose the haemodynamic actions of Ang II. Another peptide contributing to the vasodilatory effects of Ang-(1-7) is bradykinin (BK) [29], a molecule Ang-(1-7) is shown to interact with. Paula and colleagues [30] suggested that Ang-(1-7) potentiates the hypotensive effect of bradykinin in vivo and prostaglandins may participate in the mechanism of potentiation by Ang-(1-7).
There are other potentially cardioprotective effects of Ang-(1-7) shown in animal studies.
1.3. Aims and Objectives
The objective of this systematic review will be to discuss human studies on the beneficial effects of angiotensin-(1-7) focusing on patients with heart failure.
2. Methods
2.1. Search Strategy of the Systematic Review
Highly sensitive search strategies were developed using appropriate subject headings and text word terms. The following electronic databases were searched: Pubmed, Embase, and Cochrane. In addition, conference proceedings and reference lists of all included studies were scanned to identify additionally potentially relevant studies. There were no start year restrictions, but the studies examined were restricted to English language reports.
2.2. Data Extraction
Two reviewers screened the titles (and abstracts if available) of all reports identified by the search strategy. Full copies of potentially relevant reports were obtained, studied, and assessed for inclusion. Data was discussed with the senior author, and disagreements were resolved by consensus.
3. Results
Figure 1 helps the reader appreciate the close relationship between Angiotensin-(1-7) and Angiotensin II. Figure 2 summarises the types of studies considered in the review.
Figure 1.
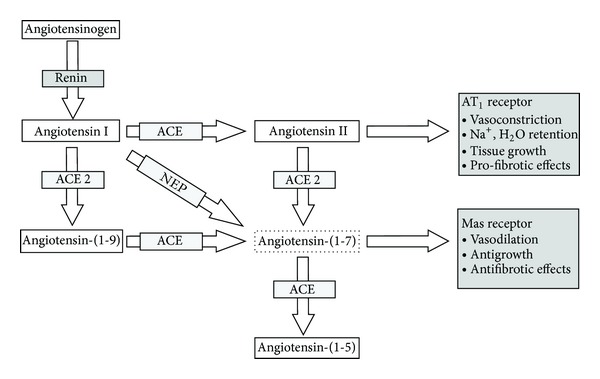
Demonstrating the relationship between Angiotensin I, Angiotensin II, Ang-(1-7), and the converting enzymes (ACE and ACE2).
Figure 2.
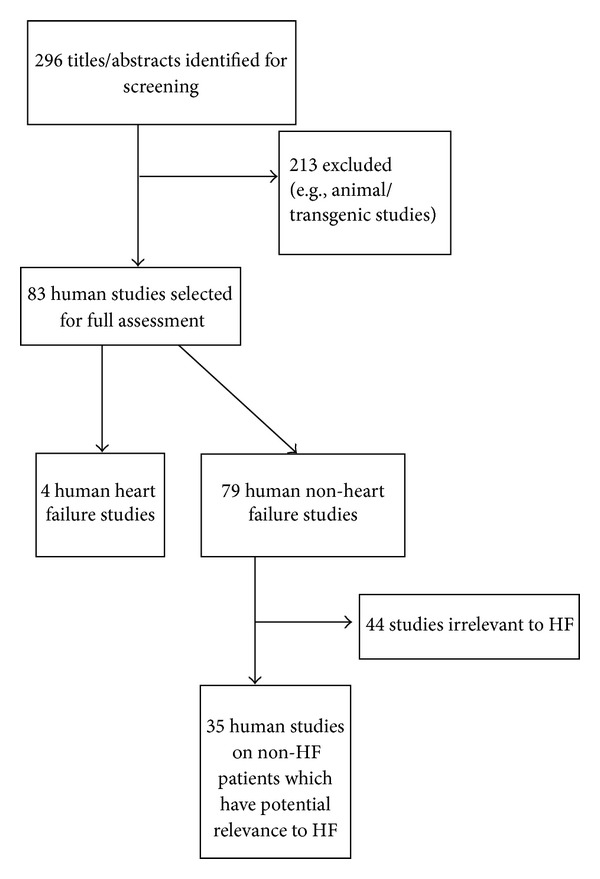
Flow of studies through the review.
3.1. Studies on Human Heart Failure Patients
See Table 1.
Table 1.
Studies on human heart failure patients.
| Study | Full publication or abstract | N | Endpoint | Key findings |
|---|---|---|---|---|
| Davie and McMurray [31] | Full | 8 | Blood pressure, forearm blood flow (FBF) | Ang-(1-7) has no significant effect on the endpoints. |
|
| ||||
| Zisman et al. [12] | Full | 22 | Ang-(1-7) forming activity | Ang-(1-7) forming activity is increased in failing heart. |
|
| ||||
| Campbell et al. [13] | Full | 9 | Ang-(1-7) levels in arterial and coronary sinus blood | 39- and 22-fold increase of Ang-(1-7) levels in the coronary sinus and arterial blood, respectively (patients were receiving ACE inhibitor therapy). A parallel increase in Ang-(1-7) and Ang I levels was observed in HF patients on ACE inhibitors. A 7.5- and 2.25-fold increase in the Ang-(1-7)/Ang II ratio was associated with ACE inhibition in the coronary sinus and arterial blood, respectively. |
|
| ||||
| Batlle Perales et al. [32] | Abstract | 33 (end-stage HF); 11 (control) |
Increased Mas receptor expression within myocardiums in the remodelling stage (as suggested by MMP3 and collagen I). Ang-(1-7) is therefore suggested to have a role in the remodelling process. | |
3.2. Studies on Non-Heart-Failure Patients
See Table 2.
Table 2.
Studies in non-heart failure patients.
| Study | Full publication or abstract | Disease and control population (if applicable) | Study type, for example, cohort/biopsy from patients/human cells | N | Key endpoint(s) | Key finding(s) |
|---|---|---|---|---|---|---|
| Ueda et al. [18] | Full | Healthy; normotensive | Cohort | 22 | Forearm blood flow (FBF) | Ang-(1-7) attenuates Ang II-induced vasoconstriction. |
|
| ||||||
| Campbell et al. [13] | Full | Coronary artery disease (CAD) | Cohort | 20 | Ang-(1-7) levels in coronary sinus and arterial blood (key endpoint of relevance to Ang-(1-7) ) | No significant change in the level of Ang-(1-7) after acute intravenous administration of an ACE inhibitor. On an assumed haematocrit of 45%, net Ang-(1-7) formation by the heart was unchanged after ACE inhibitor administration. |
|
| ||||||
| Ferrario et al. [33] | Full | Healthy volunteers and essential hypertension (HTN) | Cohort | Healthy volunteers (n = 31), essential HTN (n = 18) | Urinary concentration of Ang-(1-7) | Urinary Ang-(1-7) correlated inversely with arterial pressures reduced levels in untreated essential HTN patients. |
|
| ||||||
| Roks et al. [34] | Full | Patients undergoing coronary artery bypass grafting (CABG) surgery and healthy donors | Biopsies | CABG (n = 25), healthy donor (n = 1) | Plasma/tissue ACE activity and contractile function in human arteries | Ang-(1-7) blocks arterial vasoconstriction and inhibits ACE in plasma, atrial and arterial, tissues. |
|
| ||||||
| Wilsdorf et al. [19] | Full | Healthy subjects | Cohort | 8 | FBF; endothelial TPA release | Ang-(1-7) had no significant effect on the endpoints. |
|
| ||||||
| Pan et al. [35] | Full | NA | Primary cultures of human cardiac (HC) myocytes and HC fibroblasts | NA | Transcription of matrix metalloproteinase (MMP)−1, −2, and −9, and tissue inhibitors of matrix metalloproteinase (TIMP) −1, −2, and −3 | Ang-(1-7) and Ang II have opposing and antagonistic effects. Ratios of MMPs to TIMPs were decreased (suggestive of less myocardial fibrosis) by Ang-(1-7). |
|
| ||||||
| Sampaio et al. [36] | Full | NA | Human aortic endothelial cells (HAECs) | NA | Activation of NAD(P)H oxidase; phosphorylation of extracellular signal regulated kinase (ERK)1/2; c-Src activation; SHP-2 phosphorylation; c-Src and SHP-2 interaction | Ang II signalling is counter-regulated by Ang-(1-7); effects are mediated probably via Mas. |
|
| ||||||
| Rajendran et al. [37] | Full | Normal subjects and patients with acute coronary syndrome (ACS) | Cohort study | Normal (n = 17) ACS (n = 17) | Platelet aggregation and responsiveness | Ang-(1-7) reduces platelet aggregation by potentiating sodium nitroprusside. |
|
| ||||||
| Peltonen et al. [38] | Full | Aortic regurgitation (AR); AR plus fibrosis (AR + F); aortic stenosis (AS); normal valve (control) | Cohort | Control (n = 11); AR (n = 14); AR + F (n = 20); AS (n = 61) | Expression of Mas receptor (key endpoint of relevance to Ang-(1-7)) | Mas receptor mRNA levels in stenotic valves were lower than control, AR, and AR + F valves, further supporting the hypothesis that myocardial fibrosis is attenuated by Ang-(1-7), an endogenous Mas receptor agonist. |
|
| ||||||
| Christofi et al. [39] | Abstract | NA | Human saphenous vein cells (hSVSMC); human coronary artery cells (hCAC) and saphenous vein cells (hVTSM1)-derived immortalised human VSMC | NA | PCR on complimentary DNA expression of the Mas receptor (key endpoint of relevance to Ang-(1-7)) | Mas gene expression was not detected in these cells. |
|
| ||||||
| Sampaio et al. [40] | Full | NA | Human aortic endothelial cells (HAECs) | NA | NO synthase (eNOS) expression/activity; role of Mas; regulation of ser1179 and thr495 phosphorylation sites of NO synthase; role of the phosphatidylinositol 3-kinase (PI3K)/Akt-pathway | HAECs express Mas and via this receptor mediate the activation of eNOS. eNOS activation and NO release by Ang-(1-7) involve the role of the PI3K/Akt pathway. |
|
| ||||||
| Lin et al. [41] | Full | NA | Human cardiac fibroblasts | NA | ACE2 mRNA and protein expression; phosphorylated ERK1/2 (p-ERK1/2) protein expression | Ang-(1-7) upregulates ACE2 expression, possibly independent of the Ang II-Angiotensin type 1 receptor signalling pathway. A positive feedback loop is observed (Ang-(1-7) increases ACE2 expression in vitro; increased ACE2 could promote more Ang-(1-7)). |
|
| ||||||
| Schindler et al. [42] | Full | Healthy young male nonsmokers | Cohort | 8 | Ang-(1-7) peptide level (determined relevant from the results section) | Irbesartan significantly increased Ang-(1-7) peptide levels. Atorvastatin had no effect on the Ang-(1-7) peptide level. |
|
| ||||||
| Rajendran et al. [43] | Full | NA | Whole blood | NA | Ang-(1-7) effect on NO responsiveness of platelets; is this associated with the modulation of O2
− release? Role of a specific Ang-(1-7) receptor |
Effects of Ang-(1-7) occurred only in whole blood (another experiment was done on platelet-rich plasma). The antiaggregatory effects of the NO donor sodium nitroprusside (SNP) were potentiated by Ang-(1-7), probably by a specific Ang-(1-7) receptor. O2 − release suppression by SNP was potentiated by Ang-(1-7). |
|
| ||||||
| Ueda et al. [44] | Full | Healthy normotensive | Cohort | Procedure 1 (n = 8); procedure 2 (n = 8); procedure 3 (n = 6 out of 8 from procedure 2) | FBF | Ang-(1-7) in a dose-dependent manner potentiated the vasodilating effect of BK (doses of 1000 pmol/min and 100 pmol/min). Abolishing effects of an NO synthase inhibitor were not statistically significant. There was no effect of Ang-(1-7) on the vasodilating effects of either acetylcholine or SNP. |
|
| ||||||
| Gironacci et al. [45] | Full | NA | Mas receptor-yellow fluorescent protein (MasR-YFP) transfected human embryonic kidney 293T cells | Na | Relative cellular distribution of MasR-YFP | Ang-(1-7) causes the MasR to undergo endocytosis. |
|
| ||||||
| Luque et al. [46] | Full | Essential HTN | Cohort | 24 | Plasma concentration of Ang-(1-7) in the peripheral venous blood (key endpoint of relevance to Ang-(1-7)) | The last dose of captopril (6 months after) produced significantly greater levels of Ang-(1-7). There was a negative correlation between plasma Ang-(1-7) and diastolic blood pressure in a subset of essential HTN subjects (in whom BP was controlled with captopril only). |
|
| ||||||
| Hayashi et al. [47] | Full | NA | Smooth muscle cells and endothelial cells | NA | ERK1/2 phosphorylation; smooth muscle cell proliferation; adhesion of monocytes to endothelial cells. | D-Ala (Mas antagonist) pretreatment decreased the inhibitory effect of olmesartan (in response to Ang II stimulation). Ang II increased the endpoints which olmesartan inhibited. |
|
| ||||||
| Zisman et al. [48] | Full | Orthotopic heart transplantation recipients (normal coronary anatomy and left ventricular function) | Cohort | 4 | 123I-Ang-(1-7) was quantified in the myocardial circulation. | Ang-(1-7) is produced in the myocardial circulation. Reduced levels of Ang II (via enalaprilat) caused levels of Ang-(1-7) to decrease. |
|
| ||||||
| Gallagher and Tallant [49] | Full | NA | Lung adenocarcinoma cells; adenocarcinoma cells; squamous cell; carcinoma cells |
NA | Cell number; DNA synthesis; inhibition time course of DNA synthesis by Ang-(1-7); Mas mRNA; DNa replication; ERK1/2 activities | Lung cancer cell growth is inhibited by Ang-(1-7), possibly via the activation of an Angiotensin peptide receptor, which may involve the ERK signal transduction pathway. |
|
| ||||||
| Pignone et al. [50] | Full | Systemic sclerosis (SSc); control subjects | Cohort | SSc (n = 32); control (n = 55) | Ang-(1-7), neutral endopeptidase (NEP), NO and prostaglandin I2 (PGI2) levels | Lower Ang-(1-7) levels were detected in patients with SSc. The Ang II/Ang-(1-7) ratio in SSc patients showed greater Ang II levels to Ang-(1-7). In the controls, Ang-(1-7) was prevalent. NEP, NO, and PGI2 levels are reduced in the SSc group. |
|
| ||||||
| Silva et al. [51] | Full | HTN (12 renovascular; 15 essential HTN); normotensive subjects; paediatric population |
Cohort | Renovascular HTN (n = 12); essential HTN (n = 15); normotension (n = 32) | Ang-(1-7) levels in the blood (key endpoint of relevance to Ang-(1-7)) | Ang-(1-7) levels are significantly higher in HTN (renovascular) patients compared to normal children. In the essential HTN subjects, Ang-(1-7) levels were significantly increased compared with normotensive subjects. Calcium channel blockers had no effect on the RAAS measurements. |
|
| ||||||
| Simões E Silva et al. [52] | Full | Normotensive healthy subjects; normotensive chronic renal failure (CRF); hypertensive CRF; end-stage renal disease (ESRD) | Cohort | Normotensive healthy subjects (n = 32); normotensive CRF (n = 23); hypertensive CRF (n = 34); ESRD (n = 21) | Radioimmunoassays for Ang-(1-7) levels (key endpoint of relevance to Ang-(1-7)) | In the hypertensive CRF subjects, Ang-(1-7) levels were higher compared with normotensive CRF and healthy subjects. There was no difference between normotensive CRF and healthy subjects. |
|
| ||||||
| Nickenig et al. [53] | Full | NA | Human skin fibroblasts (from a skin biopsy; purchased) | NA | Ang-(1-7) competed for the Ang II binding, causing 80% loss of binding activity (approximately). Ang-(1-7) may be involved in DNA synthesis. Effects of Ang-(1-7) may be mediated via the Mas receptor. |
|
|
| ||||||
| Anton et al. [54] | Full | NA | Human umbilical vein endothelial cells (HUVECs) | NA | Tube formation of HUVECs | Ang-(1-7) exerted inhibitory effects on HUVEC tube formation. The effect of Ang-(1-7) was reversed by A779. Losartan also reversed the Ang-(1-7) mediated inhibition (similarly as to A779). |
|
| ||||||
| Villalobos et al. [55] |
Abstract | NA | Cultured human vascular smooth muscle cells (HASMC) | NA | Levels of inducible NO synthase (iNOS) and NO release | Ang-(1-7) acts via Mas receptor and partially prevented vascular smooth muscle inflammation |
|
| ||||||
| Montezano et al. [56] | Abstract | NA | Cultured human microvascular endothelial cells (HMEC) | NA | Expression of a proinflammatory mediator, cell growth marker, and ETBreceptor. ETA receptor gene expression | Proinflammatory and mitogenic actions of ET − 1/ETA receptor were negatively modulated by Ang-(1-7). |
|
| ||||||
| Peltonen et al. [57] | Abstract | Normal valves (control); AR; AR + F; AS | Cohort | Control (n = 11); AR (n = 11); AR + F (n = 17); AS (n = 57) | Expression of the Mas receptor in aortic valves (key endpoint of relevance to Ang-(1-7)) | The Mas receptor is downregulated in stenotic aortic valves. Mas gene expression is not affected by statin treatment in the AS group. |
|
| ||||||
| Zhiming et al. [58] | Abstract | NA | THP-1 derived macrophages (human) | NA | mRNA and protein expression of ATP-binding cassette transporter A1 (ABCA1); macrophage cholesterol efflux | Decreased THP-1 induced macrophage cholesterol efflux, and ABCA1 expression by Ang II was reversed by Ang-(1-7) in a dose-response relationship. A-779 (Ang-(1-7) Mas receptor antagonist) had no effect on the endpoints. |
|
| ||||||
| Vzquez-Bella et al. [59] | Abstract | NA | Cultured human umbilical endothelial veins (HUVECs) | NA | Endothelial nitric oxide synthase (eNOS) and NO levels; L-arginine (eNOS substrate) asymmetric dimethylarginine (ADMA) (eNOS inhibitor); expression of ICAM-1 and VCAM-1 (cell adhesion molecules) | Ang-(1-7) pretreatment increased the NO release mediated by BK, an effect inhibited by A779 pre-treatment. Pretreatment with A779 increases levels of the inactive phosphorylated (Thr 495) form of eNOS and reduces the L-arginine/ADMA ratio. There was no effect of Ang-(1-7) on VCAM-1 and ICAM-1 expressions in nonstimulated (non-Ang II) HUVECs. There was reduced induction of ICAM-1 and VAM-1 in stimulated cells. |
|
| ||||||
| Santos et al. [60] |
Abstract | NA | Confluent cultured human aortic and umbilical vein endothelial cells | NA | Formation of Ang-(1-7) | Generation of 125I-Angiotensin-(1-7) was time dependent when incubated with 125I-Angiotensin I. |
|
| ||||||
| Hermenegildo et al. [61] | Abstract | NA | Human umbilical vein endothelial cells (HUVECs) | NA | Expression of mRNA and protein of the enzymes associated with Ang-(1-7) production and NO synthesis | Oestradiol (E2) increased the expression of enzymes implicated in the production of NO and NO receptor expressions. A779 abolished E2's effect on NO synthase and NO receptor expression. A779 inhibited NO levels (NO levels increased by E2). |
|
| ||||||
| Kono et al. [62] | Full | Normal men and Bartter syndrome (BS) | Cohort | Normal (n = 5); BS (n = 3) | BP; aldosterone and plasma renin (key endpoint of relevance to Ang-(1-7)) | A significant increase in BP was observed in normal men after Ang-(1-7) infusion. After the infusion of Ang-(1-7), it took 20 min (for systolic) and 30 min (for diastolic) for the pressor actions to cease. In patients with BS, Ang-(1-7) had no effect. Ang-(1-7) had no effect on aldosterone; however, it did lower plasma renin activity in both normal and BS groups. Ang-(1-7) has pressor actions, however, having effects <0.03% of Ang II. |
|
| ||||||
| Sasaki et al. [63] | Full | Essential HTN and normotensive controls | Cohort | Normotension (n = 8); essential HTN (n = 8) | FBF | Ang-(1-7) increases FBF through NO independent manner. |
|
| ||||||
| Calò et al. [64] | Full | Bartter syndrome/Gitelman syndrome* (BS/GS); normotensive healthy subjects; essential HTN | Cohort | BS/GS (n = 10); normotensive healthy subjects (n = 10); untreated essential HTN (n = 10) |
Levels of ACE2 and Ang-(1-7) | Ang-(1-7) levels are elevated in BS/GS patients compared with the other two groups. In BS/GS patients, there was a direct correlation between ACE2 and Ang-(1-7). This study provides further support of the hypothesis that Ang-(1-7) regulates vascular tone. |
*In Bartter syndrome/Gitelman syndrome, patients have gene defects in specific kidney transporters and ion channels, resulting in raised plasma Ang II and aldosterone, but intriguingly, they have normal or even low blood pressure. Their peripheral resistance is reduced, and they exhibit hyporesponsiveness to pressors.
4. Discussion
The main finding of this systematic review is that Ang-(1-7) plays an important cardioprotective role in heart failure in animals and in patients without heart failure. However, to date, the evidence for the role of Ang-(1-7) in human heart failure is limited (Table 1), whilst there are many more studies in non-heart-failure patients (Table 2).
The only study which evaluated the effects of Ang-(1-7) on patients with heart failure examined only 8 patients. These patients were all treated with an ACE inhibitor. The interaction of Ang-(1-7) with bradykinin was elegantly studied [31]. Ang-(1-7) did not have any significant effect on the pulse rate, blood pressure, or forearm blood flow in the noninfused arm during infusion of the peptide. Slight vasoconstriction was observed during Ang-(1-7) infusion at 500 pmol/min and 5000 pmol/min; however, this was not statistically significant, and the absolute magnitude of the effect was very small. Importantly, no effect was observed with lower or higher doses. When Ang-(1-7) was coinfused with BK, the results were very similar to the initial infusion of BK alone. Interestingly, it showed a slight reduction in response to BK, though this was not statistically significant. Thus, intriguingly, the results from this study [31] are contradictory to the wealth of evidence gained from animal studies. Nevertheless, the findings were similar to another small study conducted on 8 non-heart-failure subjects [19]. Another study also found that if Ang-(1-7) was administered above a certain concentration (dose), its actions were abolished [37].
It is important to bear in mind the species gap between humans and animals and also the differences in methodologies used in the studies (e.g., some animal studies studied the effect of Ang-(1-7) on coronary arteries and others looked at mesenteric arteries). It seems that Ang-(1-7) is biologically inactive, in keeping with a study by Kono et al. [62] who also reported biological inactivity. If anything, its effects appear to have opposite actions (albeit nonsignificantly) to what is generally believed, a vasodilating, antiproliferative, and counterregulatory peptide. This lack of effect of Ang-(1-7) on haemodynamics is intriguing. It should be noted that the study had a relatively small sample size: 8 patients with chronic heart failure secondary to left ventricular dysfunction were recruited. The study could be underpowered statistically. All the patients in the study were already taking ACE inhibitors; therefore, the effect of Ang-(1-7) potentiating BK may have been obscured.
On the other hand, Ang-(1-7) attenuates vasoconstriction and increases blood flow in some studies, involving a larger number of patients, across different vascular beds, including the forearm and internal mammary artery [18, 32, 34, 44]. Thus, there are human studies demonstrating that the vasodilating property Ang-(1-7) is believed to hold. Ueda et al. [18] provided evidence of Ang-(1-7)'s action of attenuating vasoconstriction by Ang II in a dose-dependent manner. Sasaki et al. [63] supported this observation of vasodilation. The findings from Roks et al. [34] and Ueda et al. [44] were also consistent with the detection of a vasodilatory effect of Ang-(1-7). Reference [34] had a study population of 25 patients undergoing CABG (one did not have CABG), and here, Ang-(1-7) was found to behave like an ACE inhibitor. The blockade of vasoconstriction in this study could be due to many mechanisms which include the counterregulation of Ang II signalling [36] and the inhibition of ACE. Reference [44] involved a double-blind crossover design, allowing less room for bias. Coinfusion of Ang-(1-7) at 1000 pmol/min with BK significantly shifted the dose-response curve to the left; this was not the case at 100 and 10 pmol/min (additional analysis was performed to take into account the possible tachyphylaxis of BK in the study, which then found Ang-(1-7) at 100 pmol/min to show effect). Ang-(1-7) seems to operate within a specified range according to a few human studies so far [31, 37, 44, 63]. Wilsdorf et al. [19] result of Ang-(1-7) having no significant effect on BK may have been due to fact that Wilsdorf et al. did not take into account tachyphylaxis in the data; therefore, the effect of Ang-(1-7) may have been masked by this possible phenomenon. References [44, 65] both found tachyphylactic responses to BK. Moreover, Wilsdorf and colleagues [19] did not analyse and measure greater doses of Ang-(1-7), which [44] did and found a significant effect.
Other studies about Ang-(1-7) in human heart failure have provided important insight into its role and suggested mechanisms, whereby it might be further exploited as a peptide with cardioprotective therapeutic benefits. Zisman et al. [12] validated a potential role for Ang-(1-7) in the human failing heart in an elegant study involving explanted hearts. This study involved 22 patients with heart failure due to either end-stage idiopathic cardiomyopathy (IDC) or primary pulmonary hypertension (PPH), compared with 13 patients with normal left ventricles (LV) on echocardiography prior to organ donation, which eventually sadly could not take place due to ABO blood type or donor/recipient size mismatch. In the transplanted hearts, Ang-(1-7) forming activity was significantly increased in the IDC left ventricle; the forming activity was greater for Ang II as a substrate than that for Ang I. Furthermore, the forming activity was greater than 4-fold (Ang II as substrate) compared to the nonfailing left ventricle. Ang II as a substrate for Ang-(1-7) appears to have activity in more areas of the heart than for Ang I, increasing the levels of Ang-(1-7) in both IDC ventricles and the PPH right ventricle. It should be remembered that the PPH left ventricle was not dysfunctional, which may explain the lack of increased Ang-(1-7) forming activity in the left ventricles of patients suffering from primary pulmonary hypertension. ACE inhibitor therapy did not significantly affect angiotensinase activity. Angiotensinase activity was higher in the failing hearts than the non-failing hearts. Zisman et al. [12] indicated a role for neutral endopeptidase and ACE2 in the formation of Ang-(1-7). The Ang-(1-7) forming activity was greater when Ang II was the substrate; moreover, the forming activity was identified in more areas of the heart, suggesting a major role for ACE2 in the failing heart.
Furthermore, few animal studies have shown that AT2 mediates antiproliferative and apoptotic signaling [66]. This has potentially beneficial consequences: it may positively influence the remodelling process known to be detrimental to cardiac function. However, it is unlikely that a direct relation between Ang-(1-7) and the AT2 receptor exists because it is not a natural ligand for it. It may be possible for there to be cross-talk between the Mas receptor and AT2 receptor. There is a strong correlation between Ang-(1-7) and Ang II, being efficiently converted, and the given evidence from previous studies in favour of Ang-(1-7) as a promising peptide to oppose Ang II effects suggests a potential counterregulatory role of Ang-(1-7) in the RAAS, thus being cardioprotective. This study has a few drawbacks; because of the method used to homogenise tissue, influential enzymes may have been lost, therefore unable to exert their effects on either Ang I or Ang II. Not only that, but also, as the authors were focusing on membrane-bound angiotensinases, they may have not identified the potential effects of the soluble ACE2. ACE2 gene expression was shown to be upregulated in the human failing heart in a study by Goulter et al. [67]. Putting the evidence from Zisman et al. [12], Goulter et al. [67], and Pan et al. [35] who declared positive influences by Ang-(1-7) on metalloproteinases (as a measure of myocardial fibrosis in the failing human heart) together, they showed Ang-(1-7) to be a promising physiological peptide in the failing human heart. In the postmyocardial infarction period, an increased matrix metalloproteinase (MMP)/tissue inhibitors of matrix metalloproteinases (TIMPs) ratio contributes to the remodelling stage [68]. Spinale et al. [69] and Schwartzkopff et al. [70] also reported changes in the MMP/TIMP ratio in the human failing heart.
Campbell et al. [13] have importantly provided evidence of Ang-(1-7)'s presence within the coronary sinus and arterial (radial) circulation of patients with heart failure taking ACE inhibitors. Here, an increased level of Ang-(1-7) was shown, by 39- and 22-fold in the coronary sinus and radial arterial blood, respectively, compared to non-heart-failure patients who were also not on ACE inhibitors. Ang-(1-7) appeared to rise in a parallel manner with Ang I levels. These are significant findings as ACE inhibitors now form the cornerstone of the management plan for heart failure. In patients treated with ACE inhibitors, angiotensin I goes up, leading to a rise in Ang-(1-7) via the NEP-like pathway. Thus, the NEP pathway is recognised as an alternate route for the production of Ang-(1-7) [12, 71]. When associated with ACE inhibition, the Ang-(1-7)/Ang II ratio has been found to be increased by 7.5- and 2.25-fold in the coronary sinus and arterial blood respectively. This provides further strong evidence in support of the role of the NEP-like pathway.
As discussed, Ang-(1-7) attenuates vasoconstriction and increases FBF in some studies [18, 34, 44, 63]. Further, Ang-(1-7) has anti-aggregatory effects [37, 43], opposes Ang II signalling in endothelial cells [36], and has beneficial effects against human atherosclerosis [58]. Therefore, finding it within the coronary sinus is highly regarded. However, Davie and McMurray [31] and Wilsdorf et al. [19] did not find any effect on the FBF in a small heart failure cohort and another small study of non-heart-failure patients. Campbell and colleagues [13] have presented data displaying a shift in the balance within the RAAS towards Ang-(1-7), increasing the physiological armoury against the deleterious effects of the ACE-Ang II-angiotensin subtype 1 receptor (AT1R) axis. Lin et al. [41] suggested a positive feedback loop where increased levels of Ang-(1-7) could increase the levels of ACE2 which would further increase Ang-(1-7). This was only shown in vitro. The common theme from these two studies along with Zisman et al. [12] is an increased level of Ang-(1-7). The study population of [13] was relatively small, involving 9 patients; however, this study has provided results proposing a beneficial cardioprotective role for Ang-(1-7) in heart failure. The study population was not homogeneous; moreover, there were a number of differences between the two groups, for instance, drug therapy. One possible explanation for the results in the study is that ACE inhibitor therapy can increase Ang-(1-7) levels [46].
Additionally, in 33 patients with end-stage HF undergoing heart transplant, as well as 11 controls, MAS17, MMP3, and collagen I mRNA expression were analysed in myocardial biopsies [32]. The patients were relatively young (mean age 54) but suffered severe left ventricular impairment (mean EF 21%). Their left ventricles were significantly dilated (mean 7.2 cm). Just over half had ischaemic heart failure, and 16 had nonischaemic aetiology. In this study, Perales et al. [32] demonstrated that the level of MMP3 and collagen I expressions was suggestive of the tissue being in the remodelling stage in a proportion of myocardium studied. Importantly, within the same subset of myocardium, there was an increased expression of the Mas receptor. As this receptor has been shown to mediate many actions of Ang II, it suggests a role for Ang-(1-7) in the remodelling process. Interestingly, in the small sample of hearts studied, there was no significant difference between patients with MMP3 expression regarding aetiology, severity of symptoms as measured by NYHA class, medication, or left ventricular dilation, although the abstract (which is as yet unpublished in full) did not specify whether there was any significant effect of ejection fraction on MMP3 expression.
5. Clinical Implications and Future Research
Heart failure (HF) is a common condition associated with significant morbidity and mortality despite major advances in medical, revascularization, and modern cardiac resynchronisation therapy (CRT)/implantable defibrillator (ICD) devices [72]. In 2006, more than 400,000 hospitalisations were recognised as due to or complicated by HF, accounting for about 4 million NHS hospital bed-days annually. The UK national HF audit for 2008/2009 showed that 26% of patients aged <75 years and 56% of those aged >75 years would be dead within a year.
Stem cell therapy is the subject of many substantial research programmes. An alternative approach is to stimulate progenitor cells and vascular proliferation. Angiotensin (Ang)-(1-7) is an endogenous ligand for the G protein-coupled receptor Mas [11], which has a pivotal role in preserving normal endothelium-dependent relaxation [73]. Ang-(1-7) infusion stimulated proliferation of endothelial progenitor cells isolated from rodents. The theoretical beneficial effects of Ang-(1-7) in animal research on endothelial progenitor cells need to be further tested in humans. Human studies already suggest that Ang-(1-7) attenuates vasoconstriction, increases FBF in non-heart-failure patients, and opposes Ang II signalling in endothelial cells.
Importantly, Professor Walther's team has also found that infusion of Ang-(1-7) after MI in an animal model increased the number of c-Kit- and vascular endothelial growth factor-positive cells in infarcted hearts, inhibited cardiac hypertrophy, and improved cardiac function [74]. Furthermore, the nonpeptidic Ang-(1-7) receptor agonist AVE0991 also improved cardiac contractility in diabetic rats [75]. In addition, Ang (1-7) stimulates haematopoietic progenitor cells in vitro and in vivo [76]. Ang-(1-7) has already been studied in phase I/II trials in patients with solid tumours [77, 78]. Daily subcutaneous doses of 2.5–100 micrograms/kg/day were safe and well tolerated. Thus, Ang-(1-7) is a candidate molecule that has already been administered to patients with breast cancer undergoing chemotherapy to correct anaemia (a common comorbidity in HF). In a small study of 8 patients with chronic HF, Ang (1-7) had no significant effect on blood pressure (BP) and caused no adverse effects [31]. Absence of a vasodilator response in patients who are already on ACE inhibitors might be explained at least in part by the fact that the peptide Ang-(1-7) is metabolised by ACE. There is growing evidence to suggest that the beneficial effects of ACE inhibitors and angiotensin receptor blockers (ARBs) are at least in part mediated via Ang-(1-7).
In addition, chronic angiotensin-(1-7) selectively prevents cardiac fibrosis in the DOCA-salt model of hypertension, without any effect on blood pressure or cardiac hypertrophy [79]. Further support of the role of Ang-(1-7) in helping myocardial fibrosis came from observation by Raizada's team that the antifibrotic effect of an ACE2 activator correlated with increased cardiac Ang-(1-7) immunostaining [80]. Metalloproteinases and TIMPs appear to have an important role in myocardial fibrosis and cardiac dysfunction in HF. Pan et al. [35] demonstrated how Ang-(1-7) decreased ratios of MMPs to TIMPs in human cardiac cells. A study performed on patients with heart failure examining the effects of Ang-(1-7) on the expression of MMPs and TIMPs will prove beneficial. The ratios of these enzymes are altered to some degree in HF [69, 70]. This may show direct effects of Ang-(1-7) on the remodelling process known to be detrimental to the failing myocardium.
To follow on from Davie and McMurray [31], it would be worthwhile to recruit a much larger sample size to test the hypothesis that Ang-(1-7) has beneficial effects on haemodynamics in human heart failure.
Other cardioprotective effects that were demonstrated in animal studies might be translated into humans. We are planning to conduct a randomised controlled trial to test the hypotheses that Ang-(1-7) might improve cardiac hypertrophy and function.
Investigating the expression of Mas-gene mRNA or Mas-receptor distribution throughout the human failing heart will provide good insight into activity of Ang-(1-7). Furthermore, the study can also check if there is a correlation between expression and stage of HF and the remodelling process, ventricular size, and ejection fraction.
The nonpeptidic Ang (1-7) receptor agonist AVE0991 has already been shown to improve cardiac contractility in diabetic rats. Development of an oral nonpeptidic Ang (1-7) receptor agonist will enable us to perform a well-designed randomised controlled trial to study the cardioprotective effects of Ang-(1-7) in patients on ACE inhibitors or ARBs [81].
6. Conclusions
The main finding of this systematic review is that Ang-(1-7) plays an important cardioprotective role in heart failure in animals and in patients without heart failure. However, to date, the evidence for the role of Ang-(1-7) in human heart failure is limited. Nevertheless, accumulating evidence from a few studies demonstrated the importance of Ang-(1-7) as a potential therapeutic agent in heart failure.
References
- 1.Lloyd-Jones D, Adams RJ, Brown TM, et al. Executive summary: heart disease and stroke statistics-2010 update: a report from the american heart association. Circulation. 2010;121(7):e46–e215. doi: 10.1161/CIRCULATIONAHA.109.192667. [DOI] [PubMed] [Google Scholar]
- 2.Dallas TX, American Heart Association . Heart Disease and Stroke Statistics 2008 Update. Chicago, Ill, USA: American Heart Association; 2008. [Google Scholar]
- 3.McCullough PA, Philbin EF, Spertus JA, Kaatz S, Sandberg KR, Weaver WD. Confirmation of a heart failure epidemic: findings from the resource utilization among congestive heart failure (REACH) study. Journal of the American College of Cardiology. 2002;39(1):60–69. doi: 10.1016/s0735-1097(01)01700-4. [DOI] [PubMed] [Google Scholar]
- 4.Croft JB, Giles WH, Pollard RA, Keenan NL, Casper ML, Anda RF. Heart failure survival among older adults in the United States: a poor prognosis for an emerging epidemic in the Medicare population. Archives of Internal Medicine. 1999;159(5):505–510. doi: 10.1001/archinte.159.5.505. [DOI] [PubMed] [Google Scholar]
- 5.Centers for Medicare. Health Care Financing Review: Medicare and Medicaid Statistical Supplement. Baltimore, Md, USA: Centers for Medicare and Medicaid Services; 2005. (Table 5. 5: Discharges, Total Days of Care, and Program Payments for Medicare Beneficiaries Discharged from Short-Stay Hospitals, by Principal Diagnoses Within Major Diagnostic Classifications (MDCs): Calendar Year 2006). http://www.cms.hhs.gov/MedicareMedicaid StatSupp/ [Google Scholar]
- 6.Levy D, Kenchaiah S, Glarson M, et al. Long-term trends in the incidence of and survival with heart failure. New England Journal of Medicine. 2002;347(18):1397–1402. doi: 10.1056/NEJMoa020265. [DOI] [PubMed] [Google Scholar]
- 7.Dunlay SM, Redfield MM, Weston SA, et al. Hospitalizations after heart failure diagnosis. A community perspective. Journal of the American College of Cardiology. 2009;54(18):1695–1702. doi: 10.1016/j.jacc.2009.08.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Goldberg RJ, Ciampa J, Lessard D, Meyer TE, Spencer FA. Long-term survival after heart failure: a contemporary population-based perspective. Archives of Internal Medicine. 2007;167(5):490–496. doi: 10.1001/archinte.167.5.490. [DOI] [PubMed] [Google Scholar]
- 9.Ferrario CM, Iyer SN. Angiotensin-(1-7): a bioactive fragment of the renin-angiotensin system. Regulatory Peptides. 1998;78(1–3):13–18. doi: 10.1016/s0167-0115(98)00134-7. [DOI] [PubMed] [Google Scholar]
- 10.Ferrario CM. Angiotensin-(1-7) and antihypertensive mechanisms. Journal of Nephrology. 1998;11(6):278–283. [PubMed] [Google Scholar]
- 11.Santos RAS, Simoes e Silva AC, Maric C, et al. Angiotensin-(1-7) is an endogenous ligand for the G protein-coupled receptor Mas. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(14):8258–8263. doi: 10.1073/pnas.1432869100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zisman LS, Keller RS, Weaver B, et al. Increased angiotensin-(1-7)-forming activity in failing human heart ventricles: evidence for upregulation of the angiotensin-converting enzyme homologue ACE2. Circulation. 2003;108(14):1707–1712. doi: 10.1161/01.CIR.0000094734.67990.99. [DOI] [PubMed] [Google Scholar]
- 13.Campbell DJ, Zeitz CJ, Esler MD, Horowitz JD. Evidence against a major role for angiotensin converting enzyme-related carboxypeptidase (ACE2) in angiotensin peptide metabolism in the human coronary circulation. Journal of Hypertension. 2004;22(10):1971–1976. doi: 10.1097/00004872-200410000-00020. [DOI] [PubMed] [Google Scholar]
- 14.Keidar S, Kaplan M, Gamliel-Lazarovich A. ACE2 of the heart: from angiotensin I to angiotensin (1-7) Cardiovascular Research. 2007;73(3):463–469. doi: 10.1016/j.cardiores.2006.09.006. [DOI] [PubMed] [Google Scholar]
- 15.Ferrario CM, Chappell MC, Tallant EA, Brosnihan KB, Diz DI. Counterregulatory actions of angiotensin-(1-7) Hypertension. 1997;30(3):535–541. doi: 10.1161/01.hyp.30.3.535. [DOI] [PubMed] [Google Scholar]
- 16.Calò LA, Schiavo S, Davis PA, et al. ACE2 and angiotensin 1-7 are increased in a human model of cardiovascular hyporeactivity: pathophysiological implications. Journal of Nephrology. 2010;23(4):472–477. [PubMed] [Google Scholar]
- 17.Mercure C, Yogi A, Callera GE, et al. Angiotensin(1-7) blunts hypertensive cardiac remodeling by a direct effect on the heart. Circulation Research. 2008;103(11):1319–1326. doi: 10.1161/CIRCRESAHA.108.184911. [DOI] [PubMed] [Google Scholar]
- 18.Ueda S, Masumori-Maemoto S, Ashino K, et al. Angiotensin-(1-7) attenuates vasoconstriction evoked by angiotensin II but not by noradrenaline in man. Hypertension. 2000;35(4):998–1001. doi: 10.1161/01.hyp.35.4.998. [DOI] [PubMed] [Google Scholar]
- 19.Wilsdorf T, Gainer JV, Murphey LJ, Vaughan DE, Brown NJ. Angiotensin-(1-7) does not affect vasodilator or TPA responses to bradykinin in human forearm. Hypertension. 2001;37(4):1136–1140. doi: 10.1161/01.hyp.37.4.1136. [DOI] [PubMed] [Google Scholar]
- 20.Schiavone MT, Santos RAS, Brosnihan KB, Khosla MC, Ferrario CM. Release of vasopressin from the rat hypothalamo-neurohypophysial system by angiotensin-(1-7) heptapeptide. Proceedings of the National Academy of Sciences of the United States of America. 1988;85(11):4095–4098. doi: 10.1073/pnas.85.11.4095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Santos RAS, Campagnole-Santos MJ, Andrade SP. Angiotensin-(1-7): an update. Regulatory Peptides. 2000;91(1–3):45–62. doi: 10.1016/s0167-0115(00)00138-5. [DOI] [PubMed] [Google Scholar]
- 22.Heitsch H, Brovkovych S, Malinski T, Wiemer G. Angiotensin-(1-7)-stimulated nitric oxide and superoxide release from endothelial cells. Hypertension. 2001;37(1):72–76. doi: 10.1161/01.hyp.37.1.72. [DOI] [PubMed] [Google Scholar]
- 23.Li P, Chappell MC, Ferrario CM, Brosnihan KB. Angiotensin-(1-7) augments bradykinin-induced vasodilation by competing with ACE and releasing nitric oxide. Hypertension. 1997;29(1):394–400. doi: 10.1161/01.hyp.29.1.394. [DOI] [PubMed] [Google Scholar]
- 24.Trachte GJ, Meixner K, Ferrario CM, Khosla MC. Prostaglandin production in response to angiotensin-(1-7) in rabbit isolated vasa deferentia. Prostaglandins. 1990;39(4):385–394. doi: 10.1016/0090-6980(90)90120-k. [DOI] [PubMed] [Google Scholar]
- 25.Jaiswal N, Tallant EA, Diz DI, Khosla MC, Ferrario CM. Subtype 2 angiotensin receptors mediate prostaglandin synthesis in human astrocytes. Hypertension. 1991;17(6):1115–1120. doi: 10.1161/01.hyp.17.6.1115. [DOI] [PubMed] [Google Scholar]
- 26.Brosnihan KB, Li P, Ferrario CM. Angiotensin-(1-7) dilates canine coronary arteries through kinins and nitric oxide. Hypertension. 1996;27(3):523–528. doi: 10.1161/01.hyp.27.3.523. [DOI] [PubMed] [Google Scholar]
- 27.Porsti I, Bara AT, Busse R, Hecker M. Release of nitric oxide by angiotensin-(1-7) from porcine coronary endothelium: implications for a novel angiotensin receptor. British Journal of Pharmacology. 1994;111(3):652–654. doi: 10.1111/j.1476-5381.1994.tb14787.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Benter IF, Ferrario CM, Morris M, Diz DI. Antihypertensive actions of angiotensin-(1-7) in spontaneously hypertensive rats. American Journal of Physiology. 1995;269(1, part 2):H313–H319. doi: 10.1152/ajpheart.1995.269.1.H313. [DOI] [PubMed] [Google Scholar]
- 29.Fernandes L, Fortes ZB, Nigro D, Tostes RCA, Santos RAS, Catelli de Carvalho MH. Potentiation of bradykinin by angiotensin-(1-7) on arterioles of spontaneously hypertensive rats studied in vivo. Hypertension. 2001;37(2, part 2):703–709. doi: 10.1161/01.hyp.37.2.703. [DOI] [PubMed] [Google Scholar]
- 30.Paula RD, Lima CV, Khosla MC, Santos RAS. Angiotensin-(1-7) potentiates the hypotensive effect of bradykinin in conscious rats. Hypertension. 1995;26(6, part 2):1154–1159. doi: 10.1161/01.hyp.26.6.1154. [DOI] [PubMed] [Google Scholar]
- 31.Davie AP, McMurray JJV. Effect of angiotensin-(1-7) and bradykinin in patients with heart failure treated with an ACE inhibitor. Hypertension. 1999;34(3):457–460. doi: 10.1161/01.hyp.34.3.457. [DOI] [PubMed] [Google Scholar]
- 32.Batlle Perales M, Perez-Villa F, Lazaro A, et al. The Ang(1-7) mas receptor expression is increased in myocardial tissue from heart failure patients that are in a highly active remodelling stage. European Journal of Heart Failure. 2009;8:p. ii813. [Google Scholar]
- 33.Ferrario CM, Martell N, Yunis C, et al. Characterization of angeotensin-(1-7) in the urine of normal and essential hypertensive subjects. American Journal of Hypertension. 1998;11(2):137–146. doi: 10.1016/s0895-7061(97)00400-7. [DOI] [PubMed] [Google Scholar]
- 34.Roks AJM, Van Geel PP, Pinto YM, et al. Angiotensin-(1-7) is a modulator of the human renin-angiotensin system. Hypertension. 1999;34(2):296–301. doi: 10.1161/01.hyp.34.2.296. [DOI] [PubMed] [Google Scholar]
- 35.Pan C, Wen C, Lin C. Interplay of angiotensin II and angiotensin(1-7) in the regulation of matrix metalloproteinases of human cardiocytes. Experimental Physiology. 2008;93(5):599–612. doi: 10.1113/expphysiol.2007.041830. [DOI] [PubMed] [Google Scholar]
- 36.Sampaio WO, De Castro CH, Santos RAS, Schiffrin EL, Touyz RM. Angiotensin-(1-7) counterregulates angiotensin II signaling in human endothelial cells. Hypertension. 2007;50(6):1093–1098. doi: 10.1161/HYPERTENSIONAHA.106.084848. [DOI] [PubMed] [Google Scholar]
- 37.Rajendran S, Chirkov YY, Campbell DJ, Horowitz JD. Angiotensin-(1-7) enhances anti-aggregatory effects of the nitric oxide donor sodium nitroprusside. Journal of Cardiovascular Pharmacology. 2005;46(4):459–463. doi: 10.1097/01.fjc.0000176729.51819.a6. [DOI] [PubMed] [Google Scholar]
- 38.Peltonen T, Näpänkangas J, Ohtonen P, et al. (Pro)renin receptors and angiotensin converting enzyme 2/angiotensin-(1-7)/Mas receptor axis in human aortic valve stenosis. Atherosclerosis. 2011;216(1):35–43. doi: 10.1016/j.atherosclerosis.2011.01.018. [DOI] [PubMed] [Google Scholar]
- 39.Christofi F, Wijetunge S, Sever PS, Hughes AD. Expression and function of the tissue-renin-angiotensin system in human vascular smooth muscle cells in culture. Cardiovascular Research. 2010;87:p. S60. [Google Scholar]
- 40.Sampaio WO, Dos Santos RAS, Faria-Silva R, Da Mata Machado LT, Schiffrin EL, Touyz RM. Angiotensin-(1-7) through receptor Mas mediates endothelial nitric oxide synthase activation via Akt-dependent pathways. Hypertension. 2007;49(1):185–192. doi: 10.1161/01.HYP.0000251865.35728.2f. [DOI] [PubMed] [Google Scholar]
- 41.Lin C, Pan C, Wen C, Yang T, Kuan T. Regulation of angiotensin converting enzyme II by angiotensin peptides in human cardiofibroblasts. Peptides. 2010;31(7):1334–1340. doi: 10.1016/j.peptides.2010.03.026. [DOI] [PubMed] [Google Scholar]
- 42.Schindler C, Brosnihan KB, Ferrario CM, et al. Comparison of inhibitory effects of irbesartan and atorvastatin treatment on the renin angiotensin system (RAS) in veins: a randomized double-blind crossover trial in healthy subjects. Journal of Clinical Pharmacology. 2007;47(1):112–120. doi: 10.1177/0091270006294280. [DOI] [PubMed] [Google Scholar]
- 43.Rajendran S, Chirkov YY, Horowitz JD. Potentiation of platelet responsiveness to nitric oxide by angiotensin-(1-7) is associated with suppression of superoxide release. Platelets. 2007;18(2):158–164. doi: 10.1080/09537100600936158. [DOI] [PubMed] [Google Scholar]
- 44.Ueda S, Masumori-Maemoto S, Wada A, Ishii M, Brosnihan KB, Umemura S. Angiotensin(1-7) potentiates bradykinin-induced vasodilatation in man. Journal of Hypertension. 2001;19(11):2001–2009. doi: 10.1097/00004872-200111000-00010. [DOI] [PubMed] [Google Scholar]
- 45.Gironacci MM, Adamo HP, Corradi G, Santos RA, Ortiz P, Carretero OA. Angiotensin (1-7) induces mas receptor internalization. Hypertension. 2011;58(2):176–181. doi: 10.1161/HYPERTENSIONAHA.111.173344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Luque M, Martin P, Martell N, Fernandez C, Brosnihan KB, Ferrario CM. Effects of captopril related to increased levels of prostacyclin and angiotensin-(1-7) in essential hypertension. Journal of Hypertension. 1996;14(6):799–805. doi: 10.1097/00004872-199606000-00017. [DOI] [PubMed] [Google Scholar]
- 47.Hayashi N, Yamamoto K, Ohishi M, et al. The counterregulating role of ACE2 and ACE2-mediated angiotensin 1-7 signaling against angiotensin II stimulation in vascular cells. Hypertension Research. 2010;33(11):1182–1185. doi: 10.1038/hr.2010.147. [DOI] [PubMed] [Google Scholar]
- 48.Zisman LS, Meixell GE, Bristow MR, Canver CC. Angiotensin-(1-7) formation in the intact human heart: in vivo dependence on angiotensin II as substrate. Circulation. 2003;108(14):1679–1681. doi: 10.1161/01.CIR.0000094733.61689.D4. [DOI] [PubMed] [Google Scholar]
- 49.Gallagher PE, Tallant EA. Inhibition of human lung cancer cell growth by angiotensin-(1-7) Carcinogenesis. 2004;25(11):2045–2052. doi: 10.1093/carcin/bgh236. [DOI] [PubMed] [Google Scholar]
- 50.Pignone A, Del Rosso A, Brosnihan KB, et al. Reduced circulating levels of angiotensin-(1-7) in systemic sclerosis: a new pathway in the dysregulation of endothelial-dependent vascular tone control. Annals of the Rheumatic Diseases. 2007;66(10):1305–1310. doi: 10.1136/ard.2006.064493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Silva ACSE, Diniz JSS, Regueira Filho A, Santos RAS. The renin angiotensin system in childhood hypertension: selective increase of angiotensin-(1-7) in essential hypertension. Journal of Pediatrics. 2004;145(1):93–98. doi: 10.1016/j.jpeds.2004.03.055. [DOI] [PubMed] [Google Scholar]
- 52.Simões E Silva AC, Diniz JSS, Pereira RM, Pinheiro SVB, Santos RAS. Circulating renin angiotensin system in childhood chronic renal failure: marked increase of angiotensin-(1-7) in end-stage renal disease. Pediatric Research. 2006;60(6):734–739. doi: 10.1203/01.pdr.0000246100.14061.bc. [DOI] [PubMed] [Google Scholar]
- 53.Nickenig G, Geisen G, Vetter H, Sachinidis A. Characterization of angiotensin receptors on human skin fibroblasts. Journal of Molecular Medicine. 1997;75(3):217–222. doi: 10.1007/s001090050106. [DOI] [PubMed] [Google Scholar]
- 54.Anton L, Merrill DC, Neves LAA, Brosnihan KB. Angiotensin-(1-7) inhibits in vitro endothelial cell tube formation in human umbilical vein endothelial cells through the AT1-7 receptor. Endocrine. 2007;32(2):212–218. doi: 10.1007/s12020-007-9022-1. [DOI] [PubMed] [Google Scholar]
- 55.Villalobos L, Romacho T, Cercas E, Palacios E, Sanchez Ferrer C, Peir C. The angiotensin-(1-7)/Mas receptor axis exhibits anti-inflammatory properties in human vascular smooth muscle cells. European Journal of Clinical Pharmacology. 2010;66:p. S40. [Google Scholar]
- 56.Montezano AC, Yusuf H, Santos RA, Castro CH, Touyz RM. Angiotensin 1-7 attenuates growth and inflammation induced by ET-1 in human endothelial cells-Crosstalk between mas and ETB receptors. Hypertension. 2010;56(5):p. e144. [Google Scholar]
- 57.Peltonen T, Napankangas J, Ohtonen P, et al. Defining local renin-angiotensin system in human aortic valve stenosis: gene-expression of ACE2 and mas receptor in stenotic valves. Cardiology. 2010;115(4):281–282. [Google Scholar]
- 58.Zhiming Y. A study on effects of Ang II and Ang-(107) on cholesterol efflux in THP-1 human macrophages. Circulation. 2010;122(2):p. e139. [Google Scholar]
- 59.Vzquez-Bella M, Valleja S, Azcutia V, et al. The role of the angiotensin-(1-7)/receptor Mas axis in endothelial dysfunction. Methods and Findings in Experimental and Clinical Pharmacology A. 2009;31(supplement 6):p. 140. [Google Scholar]
- 60.Santos RAS, Brosnihan KB, Jacobsen DW, DiCorleto PE, Ferrario CM. Production of angiotensin-(1-7) by human vascular endothelium. Hypertension. 1992;19(supplement 2):II56–II61. doi: 10.1161/01.hyp.19.2_suppl.ii56. [DOI] [PubMed] [Google Scholar]
- 61.Hermenegildo C, Sobrino A, Monsalve E, et al. Angiotensin 1-7 mas receptor mediates nitric oxide production induced by estradiol in endothelial cells. European Journal of Clinical Pharmacology. 2010;66:p. S40. [Google Scholar]
- 62.Kono T, Taniguchi A, Imura H. Biological activities of angiotensin II-(1-6)-hexapeptide and angiotensin II-(1-7)-heptapeptide in man. Life Sciences. 1986;38(16):1515–1519. doi: 10.1016/0024-3205(86)90565-5. [DOI] [PubMed] [Google Scholar]
- 63.Sasaki S, Higashi Y, Nakagawa K, Matsuura H, Kajiyama G, Oshima T. Effects of angiotensin-(1-7) on forearm circulation in normotensive subjects and patients with essential hypertension. Hypertension. 2001;38(1):90–94. doi: 10.1161/01.hyp.38.1.90. [DOI] [PubMed] [Google Scholar]
- 64.Calò LA, Schiavo S, Davis PA, et al. ACE2 and angiotensin 1-7 are increased in a human model of cardiovascular hyporeactivity: pathophysiological implications. Journal of Nephrology. 2010;23(4):472–477. [PubMed] [Google Scholar]
- 65.Benjamin N, Cockcroft JR, Collier JG, Dollery CT, Ritter JM, Webb DJ. Local inhibition of converting enzyme and vascular responses to angiotensin and bradykinin in the human forearm. Journal of Physiology. 1989;412:543–555. doi: 10.1113/jphysiol.1989.sp017630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Von Bohlen und Halbach O, Walther T, Bader M, Albrecht D. Genetic deletion of angiotensin AT2 receptor leads to increased cell numbers in different brain structures of mice. Regulatory Peptides. 2001;99(2-3):209–216. doi: 10.1016/s0167-0115(01)00258-0. [DOI] [PubMed] [Google Scholar]
- 67.Goulter AB, Goddard MJ, Allen JC, Clark KL. ACE2 gene expression is up-regulated in the human failing heart. BMC Medicine. 2004;2:p. 19. doi: 10.1186/1741-7015-2-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Li YY, Feldman AM, Sun Y, McTiernan CF. Differential expression of tissue inhibitors of metalloproteinases in the failing human heart. Circulation. 1998;98(17):1728–1734. doi: 10.1161/01.cir.98.17.1728. [DOI] [PubMed] [Google Scholar]
- 69.Spinale FG, Coker ML, Heung LJ, et al. A matrix metalloproteinase induction/activation system exists in the human left ventricular myocardium and is upregulated in heart failure. Circulation. 2000;102(16):1944–1949. doi: 10.1161/01.cir.102.16.1944. [DOI] [PubMed] [Google Scholar]
- 70.Schwartzkopff B, Fassbach M, Pelzer B, Brehm M, Strauer BE. Elevated serum markers of collagen degradation in patients with mild to moderate dilated cardiomyopathy. European Journal of Heart Failure. 2002;4(4):439–444. doi: 10.1016/s1388-9842(02)00092-2. [DOI] [PubMed] [Google Scholar]
- 71.Yamamoto K, Chappell MC, Brosnihan KB, Ferrario CM. In vivo metabolism of angiotensin I by neutral endopeptidase (EC 3.4.24.11) in spontaneously hypertensive rats. Hypertension. 1992;19(supplement 6):692–696. doi: 10.1161/01.hyp.19.6.692. [DOI] [PubMed] [Google Scholar]
- 72.Cleland JGF, Daubert J, Erdmann E, et al. The effect of cardiac resynchronization on morbidity and mortality in heart failure. New England Journal of Medicine. 2005;352(15):1539–1549. doi: 10.1056/NEJMoa050496. [DOI] [PubMed] [Google Scholar]
- 73.Peiró C, Vallejo S, Gembardt F, et al. Endothelial dysfunction through genetic deletion or inhibition of the G protein-coupled receptor Mas: a new target to improve endothelial function. Journal of Hypertension. 2007;25(12):2421–2425. doi: 10.1097/HJH.0b013e3282f0143c. [DOI] [PubMed] [Google Scholar]
- 74.Wang Y, Qian C, Roks AJM, et al. Circulating rather than cardiac angiotensin-(1-7) stimulates cardioprotection after myocardial infarction. Circulation. 2010;3(2):286–293. doi: 10.1161/CIRCHEARTFAILURE.109.905968. [DOI] [PubMed] [Google Scholar]
- 75.Ebermann L, Spillmann F, Sidiropoulos M, et al. The angiotensin-(1-7) receptor agonist AVE0991 is cardioprotective in diabetic rats. European Journal of Pharmacology. 2008;590(1–3):276–280. doi: 10.1016/j.ejphar.2008.05.024. [DOI] [PubMed] [Google Scholar]
- 76.Heringer-Walther S, Eckert K, Schumacher S, et al. Angiotensin-(1-7) stimulates hematopoietic progenitor cells in vitro and in vivo. Haematologica. 2009;94(6):857–860. doi: 10.3324/haematol.2008.000034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Petty WJ, Miller AA, McCoy TP, Gallagher PE, Tallant EA, Torti FM. Phase I and pharmacokinetic study of angiotensin-(1-7), an endogenous antiangiogenic hormone. Clinical Cancer Research. 2009;15(23):7398–7404. doi: 10.1158/1078-0432.CCR-09-1957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Rodgers KE, Oliver J, DiZerega GS. Phase I/II dose escalation study of angiotensin 1-7 [A(1-7)] administered before and after chemotherapy in patients with newly diagnosed breast cancer. Cancer Chemotherapy and Pharmacology. 2006;57(5):559–568. doi: 10.1007/s00280-005-0078-4. [DOI] [PubMed] [Google Scholar]
- 79.Grobe JL, Mecca AP, Mao H, Katovich MJ. Chronic angiotensin-(1-7) prevents cardiac fibrosis in DOCA-salt model of hypertension. American Journal of Physiology. 2006;290(6):H2417–H2423. doi: 10.1152/ajpheart.01170.2005. [DOI] [PubMed] [Google Scholar]
- 80.Ferreira AJ, Shenoy V, Qi Y, et al. Angiotensin-converting enzyme 2 activation protects against hypertension-induced cardiac fibrosis involving extracellular signal-regulated kinases. Experimental Physiology. 2011;96(3):287–294. doi: 10.1113/expphysiol.2010.055277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Schindler C, Bramlage P, Kirch W, Ferrario CM. Role of the vasodilator peptide angiotensin-(1-7) in cardiovascular drug therapy. Vascular Health and Risk Management. 2007;3(1):125–137. [PMC free article] [PubMed] [Google Scholar]