

Abstract
Campylobacter jejuni is the leading cause of bacterial foodborne illness in the world, with symptoms ranging from acute diarrhea to severe neurological disorders. Contaminated poultry meat is a major source of C. jejuni infection, and therefore, strategies to reduce this organism in poultry, are expected to reduce the incidence of Campylobacter-associated diseases. We have investigated whether oral administration of C. jejuni-specific single-domain antibodies would reduce bacterial colonization levels in chickens. Llama single-domain antibodies specific for C. jejuni were isolated from a phage display library generated from the heavy chain IgG variable domain repertoire of a llama immunized with C. jejuni flagella. Two flagella-specific single-domain antibodies were pentamerized to yield high avidity antibodies capable of multivalent binding to the target antigen. When administered orally to C. jejuni-infected two-day old chicks, the pentabodies significantly reduced C. jejuni colonization in the ceca. In vitro, the motility of the bacteria was also reduced in the presence of the flagella-specific pentabodies, suggesting the mechanism of action is through either direct interference with flagellar motility or antibody-mediated aggregation. Fluorescent microscopy and Western blot analyses revealed specific binding of the anti-flagella pentabodies to the C. jejuni flagellin.
Introduction
Campylobacter jejuni, a Gram negative bacterium, is currently one of the most prevalent foodborne pathogens and the leading cause of bacterial gastroenteritis in humans worldwide [1]. In North America, campylobacteriosis outnumbers the reported cases of illnesses caused by Salmonella, Shigella, Listeria and Escherichia coli combined [2]–[4]. Despite relatively mild diarrheal illness, C. jejuni infection has been associated with severe long-term complications, including: Guillain-Barré Syndrome [5]–[7], reactive arthritis and inflammatory bowel disease [8], [9]. It is estimated that between 50–80% of human campylobacteriosis cases can be attributed to consumption of contaminated chicken, and therefore meat from broiler chickens is considered the primary vector for transmitting the pathogen to humans [10]–[12].
Reduction of C. jejuni levels in poultry decreases the incidence of Campylobacter-associated gastroenteritis [12], [13]. Various intervention strategies targeting different stages of poultry meat production are currently under investigation. To date the most accepted strategies work by preventing Campylobacter spp. from entering the flock through installation of hygiene barriers and fly screens, use of high quality water, reduction of slaughter age, and discontinuation of thinning practices [12], [14]–[17]. However, the susceptibility of chickens to infection by C. jejuni and its ubiquity in the environment have negatively impacted the success of biosecurity-based approaches, highlighting the need for alternative approaches by which the bacterial infection can be controlled or eliminated [3], [18], [19]. Antibiotics such as fluoroquinolones and macrolides have been approved for the control of Campylobacter spp. in both poultry and humans. However, their prolonged use in humans and animals has led to a rapid increase of resistant strains in many countries around the world and their use is no longer recommended in animal feed stocks [20]–[22]. Application of Campylobacter-specific lytic bacteriophages has been proposed as an alternative strategy. A reduction in cecal C. jejuni levels of 0.5–5 log10 CFU/g has been reported when bacteriophages were administered to chickens as feed-additives or veterinary drugs [23]–[25]. Development of resistance, however, is considered to be a potential drawback of phage therapy and has been reported following phage treatment in several studies [26], [27]. In addition, finding a phage cocktail that would kill all C. jejuni strains is unlikely. Bacteriocins, which are proteinaceous substances produced by bacteria that inhibit growth, have also been extensively studied. Addition of bacteriocins to poultry drinking water completely eliminated the pathogen in 90% of cases or reduced its levels by 106-fold or more [28]. Other biological reagents such as probiotics [29], [30] and plant bioactive compounds [31], [32] have also been used as food or water additives and have been shown to reduce Campylobacter loads in chickens. The bactericidal effects of probiotic strains such as lactic acid bacteria against C. jejuni have been attributed to the production of organic acids and bacteriocins or bacteriocin-like substances [29], [33]. Medium chain fatty acids such as caprylic acid and monoacylglycerols are alternatives to antibiotics that have been used as feed and water additives to control or eliminate Campylobacter loads in chickens [34]–[36]. However, despite the reported efficacies none of these compounds have been widely adopted in the field due to inconsistency or lack of data on efficacy, safety, toxicity, scale-up production and purification, and the development of resistance [12] (reviewed in [14]). Furthermore, other methods of intervention such as using vaccines [37]–[41], competitive exclusion [12], [14], [37], [42], [43] or producing genetically engineered Campylobacter-resistant chickens [44] have had limited success in preventing C. jejuni colonization in chickens, and therefore, have not been commercialized.
Antibodies were originally recognized as effective antimicrobial reagents by Behring and Kitasato in the early 1890s [45], [46] and since then, serum therapy became an effective strategy to combat many infectious diseases. The presence of specific antibodies in the serum or intestinal secretions has been associated with resistance of rabbits [47]–[49] and mice [49], [50] to colonization by C. jejuni. In young chickens (less than 2–3 weeks of age), the presence of maternal antibodies against Campylobacter delays the onset of colonization and reduces the rate of horizontal spread of C. jejuni in the flock [19], suggesting that passive immunotherapy using anti-Campylobacter antibodies could be an attractive approach for interfering with bacterial colonization in chickens. Indeed, passive immunization with anti-flagella monoclonal antibodies has already been shown to reduce C. jejuni colonization in mice [51]. Similarly, the use of hyperimmunized anti-C. jejuni rabbit serum or anti-C. jejuni antibodies appears to be effective in diminishing the colonization in chickens [52]. Consistent with this, others have shown that poultry abattoir workers who have high titres of Campylobacter-specific IgGs circulating in their blood rarely become ill due to Campylobacter infection [53]. Despite all these facts, antibodies as preventive or therapeutic reagents for Campylobacter treatment and control have not gained market attention largely due to the high cost of manufacturing, sensitivity of conventional antibodies to gastrointestinal (GI) tract proteases, lack of effective GI tract delivery systems, and relatively high antigenic variation among C. jejuni, which requires multiple antibody preparations to target different strains.
For applications such as bacterial toxin neutralization and/or inactivation of infectious agents, antibody fragments (e.g., Fabs, scFvs, single-domain antibodies) are preferable to whole antibodies (e.g., IgGs) due to lower production cost in a bacterial system and ease of genetic manipulation. The smallest, naturally occurring antigen binding fragments are the variable domains of heavy chain antibodies found in camelids [54] and the immunoglobulin new antigen receptors in sharks [55], [56]. The antigen binding sites of these antibodies reside in a single domain. Camelid two chain antibodies, termed heavy chain only antibodies, have been extensively studied and their variable domains, termed VHHs, also known as single domain antibodies (sdAbs) or nanobodies, have been shown to be extremely stable when cloned and expressed as monomers using recombinant expression systems [57], [58]. We have also previously shown that pentavalency can be conferred upon VHHs by fusion of the VHH to a protein domain derived from the verotoxin B homopentamer [59]. The resulting pentabodies are compact, stable antigen-binding molecules and have high functional affinity (avidity). Additionally, the pentavalent antibodies are capable of enhancing agglutination when bound to antigens [59].
In the present study, we describe the isolation of VHH single domain antibodies specific for C. jejuni flagella from a phage-display antibody library. The pentameric forms of VHHs were produced and characterized using various in vitro and functional assays. As well, the efficacies of orally administering these pentabodies in reducing C. jejuni colonization levels in chickens were evaluated.
Results
High Affinity VHH Antibodies Produced Against Campylobacter Antigens
ELISA analysis of the binding of the ELISA analysis of the binding of immune serum fractions, obtained by protein G and A chromatography of serum from day 95 after the immunization start, to C. jejuni flagella coated on microtitre plates showed that there were strong immune responses in the heavy chain antibody as well as the conventional antibody fractions when compared to the pre-immune serum collected before immunization on day 1 (data not shown).
A VHH library with a size of 5×107 individual transformants was constructed and analyzed for its complexity by sequencing 40 randomly selected colonies. All clones had inserts of expected sizes and were different from each other in their complementarity determining regions (CDRs). After four rounds of panning, individual colonies were screened by phage ELISA against flagella and VHH candidate clones were sequenced. Two unique flagella-specific sequences (FlagV1, FlagV6) were identified (GenBank™ accession numbers KF812523- KF812524), and all were determined to be VHHs based on the presence of the Camelidae hallmark amino acids at positions 42 (F/Y), 49(E/Q/A), 50(R), 52(F/V/G/L) (IMGT numbering) (http://imgt.cines.fr) (Figure 1). The VHH binders were sub-cloned into expression plasmid vectors to allow targeted expression in the periplasm of E. coli TG1. Monomeric and pentameric VHHs were produced at a yield of 10 to 80 mg/L of bacterial culture.
Figure 1. Sequence alignment of flagella-specific VHHs.

Amino acid sequence alignment of flagella–specific VHHs isolated in this study. Framework regions (FRs) and complementarity determining regions (CDRs) (boxed) are aligned according to IMGT numbering system. Hallmark positions 42, 49, 50, and 52, distinguishing VHHs from VHs, are highlighted in gray. The sequence alignment shows 65% protein sequence identity between the two VHHs.
Binding of VHH Antibodies to Flagella
When expressed as monomers (M), two of the flagella binders (Flag V1M and FlagV6M) showed strong binding activity to flagella protein as determined by ELISA. Pentamerization (P) of FlagV1 VHHs resulted in a further increase in their binding capacity as revealed by ELISA and surface plasmon resonance (SPR) (Figures 2 and 3). As shown, 50% maximum binding was achieved at 0.2 µg/ml (15.6 nM) FlagV1M and 0.005 µg/ml (40 pM) FlagV1P indicating an increase of almost 400-fold in functional affinity. The approximate affinity values for the monomeric VHHs closely matches the values which were obtained by surface plasmon resonance (SPR) and shows that FlagV1M has an affinity in the range of 20–30 nM. Similar patterns of binding were observed in ELISA with monomeric and pentameric FlagV6 antibodies (data not shown). The bindings of VHHs to biotinylated antigens captured on streptavidin surfaces were collected and analysed by SPR. The kinetic data for the VHHs is presented in Table 1 and Figure 2.
Figure 2. ELISA analysis of binding of the monomeric and pentameric VHHs to C. jejuni surface antigens.
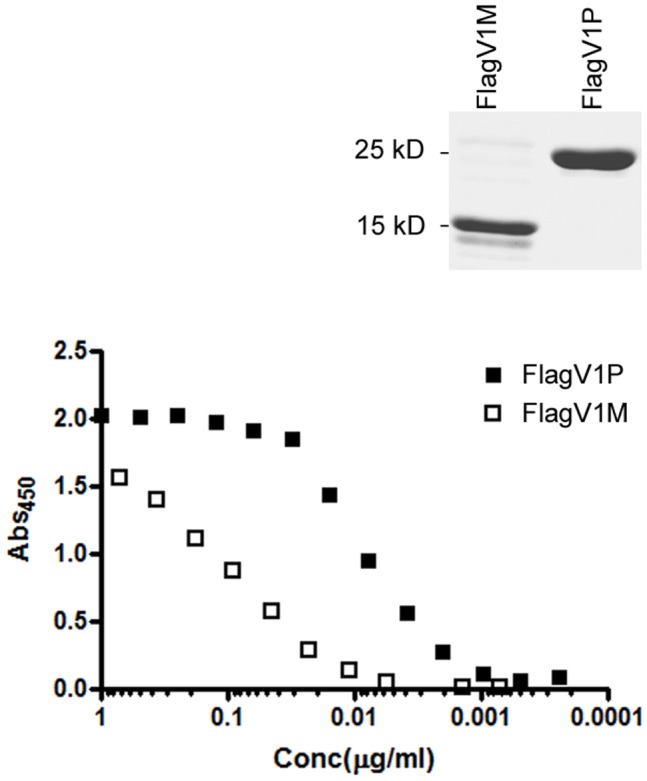
Various concentrations (ranging from 10 to 0.0001 µg/ml) of the monomeric (FlagV1M) and pentameric (FlagV1P) forms of a flagella-specific VHH were used in ELISA. Absorbance data were normalized to represent equal amounts of the monomeric and pentameric VHH formats. Solid squares represent pentabody and open squares monomeric VHH. A representative image of Coomassie stained purified monomer and pentamer VHH is shown above the ELISA graphs.
Figure 3. SPR analysis of the binding of the monomeric and pentameric VHHs to antigen.

Sensorgram overlays (A and B) represent the binding of 28, 42, 56, 70, 140 and 190 nM of FlagV1M and FlagV6M, respectively, to 700 RUs of biotinylated flagellin captured on an SA sensorchip. Black dots are the data points and the red curves are the fitting of the data to a 1∶1 interaction model. (C) The sensograms of FlagV1M (in red) and FlagV1P (in blue) were overlaid to show that a slower off-rate is obtained as a result of pentamerization indicating an increase in the functional affinity (avidity) of FlagV1P.
Table 1. Rate and affinity constants for interaction of monomeric VHHs with their respective antigens.
| VHH | kon (M−1s−1) | koff (s−1) | KD (M) |
| FlagV1M | 1×105 | 2×10−3 | 2×10−8 |
| FlagV6M | 1×106 | 3×10−2 | 2.5×10−8 |
Flagella-Specific VHHs Recognize Non-Overlapping Epitopes
SPR analyses in which FlagV1M and FlagV6M VHH pairs were co-injected on a biotinylated flagella surface showed that FlagV6M is free to bind to flagella when FlagV1M is bound at saturating concentration as evidenced by almost doubling of the R max indicating FlagV1M and FlagV6M recognize different epitopes (Figure 4).
Figure 4. Epitope mapping of anti-flagella antibodies by simultaneous binding of flagella-specific VHHs to the target antigen in SPR co-injection experiments.
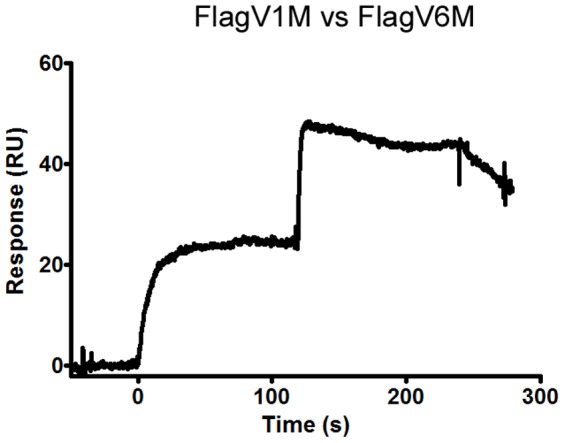
For the antigen, 60–100 µl of each VHH, at a concentration 50× its KD value, were injected over 600–700 RUs of immobilized flagella at 30 µl/min. FlagV1 and FlagV6 appeared to bind distinct, non-overlapping epitopes since the signal approximately doubled with the second injection.
Fluorescence Microscopy Reveals Binding of FlagV1P and FlagV6P to the Assembled Flagella Filament
Fluorescently labelled FlagV1P bound flagella of C. jejuni strain 81–176, but no binding was observed with a flaA−flaB− mutant strain of 81–176 (Figure 5A). A similar pattern of binding was noted with FlagV6P (Figure 5B). Interestingly FlagV6P, but not FlagV1P, was able to bind to C. jejuni strain 11168. These filaments were confirmed to be flagella with a polyclonal antibody specific to 81–176 flagella (Figure 5C). Western blotting of whole cell lysates of C. jejuni 81–176 and C. jejuni 11168 with FlagV6P and FlagV1P demonstrated a similar pattern of reactivity with denatured flagellin monomers (data not shown). As with binding to flagella filaments on the cell surface, only FlagV6P was able to bind to the flagellin monomers of both C. jejuni strains 81–176 and 11168 by Western blotting. In contrast, FlagV1M was only able to detect flagellin monomers from C. jejuni 81–176.
Figure 5. Binding of anti-flagella pentabodies to intact flagellar filament.
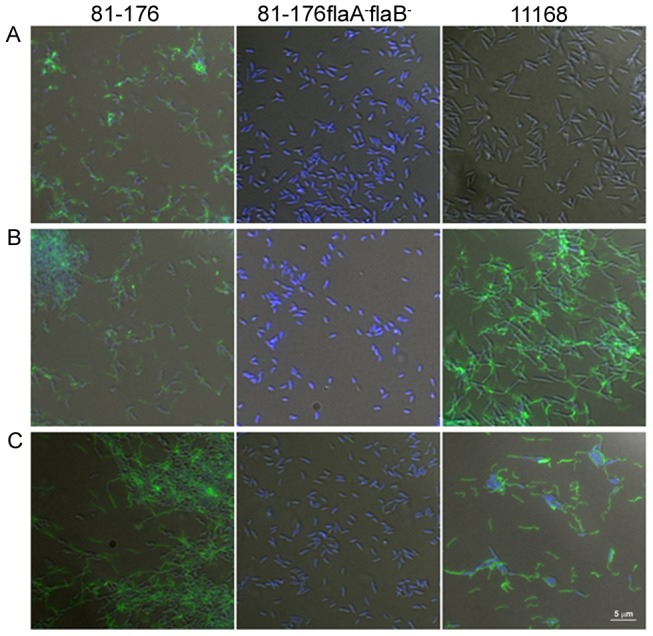
Fluorescence microscopy showing FlagV1P (A) and FlagV6P (B) pentabody binding to C. jejuni flagella. Fluorescently labelled FlagV1P and FlagV6P were hybridized with either C. jejuni strain 81–176, 81–176 flaA−flaB−, or C. jejuni strain 11168. (C) Fluorescence microscopy showing polyclonal anti-81–176 flagella binding to both strains, 81–176 and 11168. Representative fields of view are shown for all images at the same magnification, as indicated by the 5 µm bar.
Cross-Reactivity of Anti-Flagella Pentabodies with Campylobacter Strains
The flagella-specific pentabodies, FlagV1P and FlagV6P, were tested in ELISA with purified flagella prepared from 9 C. jejuni strains. All strains used in the assay were human isolates except strain P2 which was a calf isolate. The bacterial strains tested were 81–176: ATCC BAA-2151, 11168: ATCC 700819, and Penner serotype strains [60]: P1: ATCC 43429, P2: ATCC 43430, P3: ATCC 43431, P4: ATCC 43432, P19: ATCC 43446, P36: ATCC 43456, and P64. ELISA demonstrated a distinct pattern of binding of FlagV1P and FlagV6P to flagella from different strains (Figure 6). FlagV1P interacts strongly with 81–176 (the strain used for immunization) and five other strains, but it does not bind to strains 11168, P2 and P3 under the conditions tested. FlagV6P reacted with all strains tested except strain P4.
Figure 6. ELISA demonstrating binding of Flag1V1P and FlagV6P to flagella isolates from 9 different strains of C. jejuni.

ELISA was performed according to the standard procedure. In brief, wells were coated with 10 µg of C. jejuni flagella proteins and binding was detected using either FlagV1P or FlagV6P pentabodies. Absorbance values indicate an average of two independent experiments.
Anti Flagellin Pentabodies Disrupt Campylobacter Motility
VHH and pentabody-mediated inhibition of Campylobacter motility was studied using a standard soft agar plate assay. Co-incubation of C. jejuni 81–176 with FlagV1M, FlagV1P and FlagV6P monomers or pentabodies (Figure 7, Table 2) showed a marked reduction in bacterial motility. C. jejuni strain 11168 was also examined and demonstrated a similar inhibition in motility on the plate assay with only the FlagV6P antibody (data not shown). The impact of FlagV1M and FlagV1P on disrupting the motility seemed to last for up to 53 h post incubation.
Figure 7. Motility assay of C. jejuni strain 81–176 in the absence or presence of pentabodies.
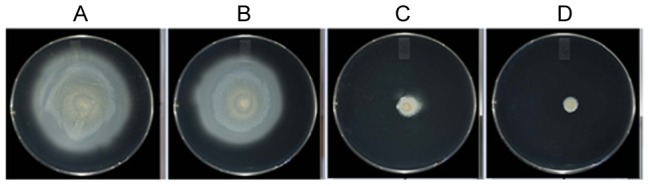
C. jejuni 81–176 was incubated for 30 min with (A) buffer, (B) control unrelated pentabody at 0.5 µg/µl, (C) FlagV1P at 0.25 µg/µl, and (D) FlagV1P at 0.5 µg/µl. Images were taken after 53 h of incubation.
Table 2. C. jejuni 81–176 motility on plates after incubation with FlagV1 and FlagV6 antibodies.
| Treatment | Diameter (mm) ±SD - 24 h | Diameter (mm) ±SD - 48 h |
| PBS | 26.6±2.25 | 82±3.3 |
| Unrelated pentabody | 24±2.7 | 67.3±4.5 |
| FlagV1M | 8.6±1.25* | 19.5±0.5* |
| FlagV1P | 8.8±0.76* | 45.16±5* |
| FlagV6P | 8.75±0.35* | 45.5±14.08 |
| FlagV1P + FlagV6P | 9±1.32* | 28±3.04* |
The diameter of the circles representing the spread of bacteria from the inoculum site was measured.
Asterisk indicates statistical significance of Flag antibody treatments vs the control unrelated pentabody.
The activity of the anti-flagellin pentabodies was also tested on Campylobacter coli strain VC167. Only FlagV6P was effective in reducing motility (Table 3). No significant effect on motility was observed when Salmonella enterica serovar Typhimurium was incubated with the anti-flagellin pentabodies (data not shown).
Table 3. Motility assays show the cross-reactivity of the pentabodies FlagV1P and FlagV6P with C. coli VC167.
| Treatment | diameter (mm) ±SD- 24 h | diameter (mm) ±SD- 48 h | diameter (mm) ±SD- 72 h |
| PBS | 12.5±0.5 | 25.3±1.52 | 43.6±4.5 |
| FlagV1P | 10.6±0.6* | 22±3 | 42.3±4.1 |
| FlagV6P | 5.6±0.66** | 11.5±1.32** | 25.6±3.05** |
Significant reduction in motility of C. coli was noticed with FlagV6P pentabody. The values were subjected to the Student's t-test for statistical analysis.
*p value <0.05.
**p value <0.005.
Anti-Flagella Pentabody Reduce Campylobacter Colonization of the Chicken Gut
The efficacy of the pentabodies in reducing C. jejuni colonization was evaluated in 2-day-old SPF chickens (Figure 8). Oral administration of FlagV1P at 1 h, 24 h and 48 h after inoculation of chickens with 108 CFU of C. jejuni significantly reduced bacterial loads in the ceca (Figure 8, p<0.001). A negative control group was also included in each experiment which showed baseline levels of C. jejuni in the ceca of uninfected chickens.
Figure 8. Effect of oral administration of FlagV1P on the levels of C. jejuni colonization of chickens inoculated with 108 C. jejuni cells.
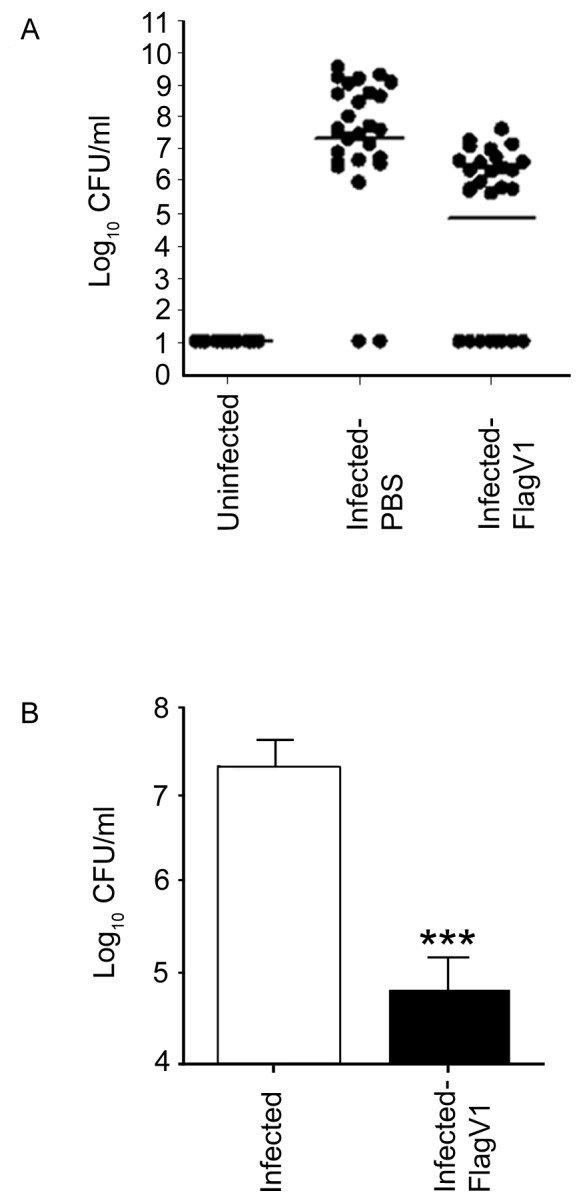
At 1C. jejuni, chickens (n = 28/group, 14 chickens in each of 2 containment units) received 300 µl FlagV1P. An uninfected, negative control group was also included (n = 15). (A) Bacterial burdens (mean ±SEM) in the individual ceca are indicated by scatterplot and the mean cecal bacterial burden is denoted by a horizontal line. (B) Bacterial colonization levels (mean ±SEM) in the ceca of chickens treated with PBS (open bars), FlagV1P (closed bars). *** p<0.001, one-way ANOVA, followed by Bonferroni multiple comparison test. The detection limit of C. jejuni in the cecum sample was determined to be 10 CFU/ml.
Administration of Anti-Flagella Pentabody Does not Impact Chicken Weight Gain
The effect of gavaged FlagV1 pentabody on chicken body weight was investigated by measuring body weights at one day and four days after challenging with Campylobacter alone or when the pentabody was administered orally. As a control, saline buffer was used and the average body weights were measured (Figure 9). No significant differences were found between the groups.
Figure 9. The effect of administration of flagella-specific antibodies on chicken body weight.
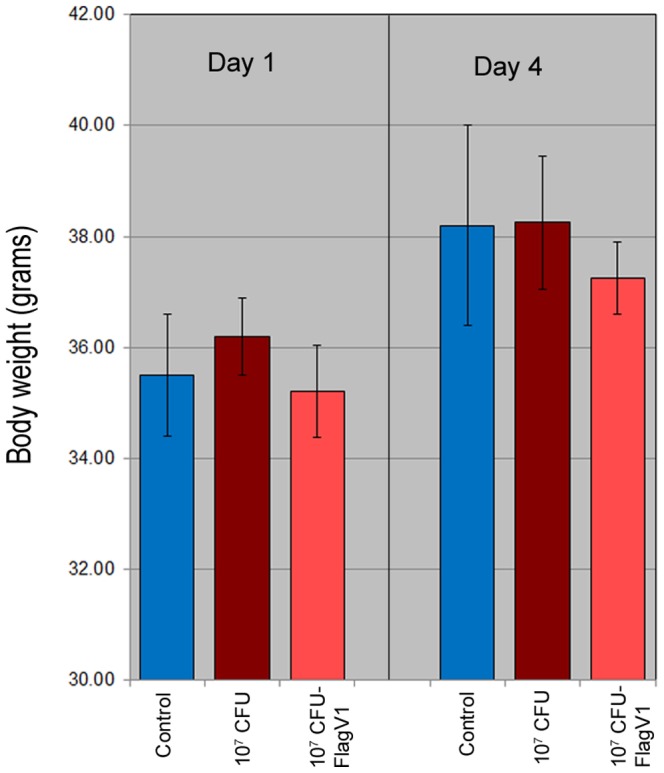
Chicken were weighed one day and four days after challenging with C. jejuni alone or C. jejuni followed by pentabody administration. PBS was used as a control and body weights (in grams) were measured at day one and day four. The average body weight ± standard deviation of the values obtained from 28 replicates is shown for each group. No significant differences were found between the groups.
Orally-Administered Flagella-Specific Pentabody Localize to the Chicken Ileum and Cecum
The chicken intestinal tract was dissected to examine for the presence of gavaged pentabodies (FlagV1P). Intestinal fluids were collected from the cecum, ileum, jejunum and duodenum and used in 2-fold serial dilutions in a sandwich ELISA where anti-verotoxin antibody was used as the capturing antibody and anti-His-HRP antibody for detection (Figure 10). On average, higher concentrations of FlagV1P pentabody were found in the Ileum and cecum fluid extracts compared to the other regions of the intestine. Intestinal fluids from control chicken showed no significant binding in the same ELISA.
Figure 10. Detection of anti-C. jejuni pentabodies (FlagV1P) in different sections of the chicken intestinal tract.

Separate chickens were gavaged with the flagella-specific pentabody according to the schedule that was followed in our chicken colonization studies. Intestinal fluids were collected from the cecum, ileum, jejunum and duodenum and used in 2-fold serial dilutions {1/2 (no. 2)-1/2048 (no. 12)} in ELISA using anti-verotoxin antibody to capture the pentabodies and anti-His-HRP antibody conjugates for their detection. Chickens were gavaged with 1 mg of FlagV1P.
Discussion
Single-domain VHH antibodies are emerging as novel biological reagents against bacterial and viral infections [61], [62]. Their small size facilitates binding to epitopes that are inaccessible to conventional antibodies and their unique physical properties such as resistance to proteolysis, denaturation and aggregation allows potential applications in oral delivery of therapeutics in humans or livestock. In addition, they can be economically produced in large amounts in bacteria and yeast for various applications.
In the present study we used phage display technology to construct and screen an immune library of VHH antibodies against C. jejuni flagella. We demonstrated that VHHs and their pentameric versions were able to bind to flagella and were effective in significantly lowering C. jejuni colonization levels in chickens when administered orally. The rationale for our experimental design is based on a number of previous studies that demonstrated a direct correlation between resistance to or delay in colonization by C. jejuni and the presence of the organism or flagella-specific antibodies in the serum/intestinal secretions in both animal models and in humans [19], [47]–[51], [65]. We believe that the unique features of Camelidae single domain antibodies, in particular, their robust neutralization power and low production cost, may provide excellent opportunities for antibody-based reduction of Campylobacter colonization.
The isolated VHHs were well expressed in E. coli, up to 10–80 mg/litre in shaker flask culture, and size exclusion chromatography demonstrated that all exist as non-aggregating monomers (results not shown). The antibodies also reacted strongly with the protein target with affinities in the range of 20–30 nM. Deduced amino acid sequences of the antibodies showed no significant sequence homology between the CDRs of FlagV1 and FlagV6. This was additionally supported by our SPR data indicating binding of the two VHHs to different epitopes. Analyses of the VHH sequences suggests that the FlagV1M VHH belong to subfamily I, whereas the FlagV6M VHH contains most of the subfamily II-specific residues based on the VHH classification system described by Harmsen et al., 2000 [63]. The anti-flagella VHHs isolated here are among the highest affinity single-domain antibodies characterized to date.
In vitro motility studies showed that FlagV1P and FlagV6P recognized native flagella and upon binding prevented cell motility of C. jejuni 81–176. The interference with bacterial motility was even more pronounced with FlagV1M compared to FlagV1P 53 h after incubation. This might be explained by potentially higher stability of the FlagV1M VHH on the assay plate. The antibodies were also able to recognize other strains of C. jejuni; and FlagV6P was even able to reduce the motility of C. coli strain VC167. Variability in the pattern of binding to different strains of C. jejuni was also identified when flagella from 9 different strains were tested by ELISA, suggesting that FlagV1 and FlagV6 CDRs target different epitopes on the flagella. This was further confirmed by SPR analyses which indicated that distinct epitopes were targeted by FlagV1M and FlagV6M. We are currently investigating the actual peptide and/or carbohydrate epitopes responsible for pentabody binding.
As shown by fluorescence microscopy, FlagV1P and FlagV6P bind to the flagella at the poles of C. jejuni. As with the ELISA, FlagV1P was able to bind 81–176, but not 11168, whereas FlagV6P was able to bind both strains. As this binding was lost with a non-flagellated mutant (flaA−, flaB−), it confirmed the binding was specific to flagellin proteins.
To determine the efficacy of the anti-flagella pentabody in reducing C. jejuni colonization, chicken studies were performed using FlagV1P due to the slower off-rate of FlagV1 compared to FlagV6 (Figure 3). The antibodies were used in the chicken studies with the assumption that the bacterial agglutination or motility inhibition caused by the VHH domains of the pentabodies would impair the ability of C. jejuni to colonize the chicken gastrointestinal tract. Flagellin is considered a major C. jejuni virulence factor and motility is important for maneuvering through the viscous intestinal mucus [64], [65]. Our experiments demonstrated an approximately 3 log reduction in the rate of colonization in the group treated with FlagV1P. This further substantiates the important role of flagella in colonization of C. jejuni. Our antibody localization studies using ELISA suggested that at least a percentage of the pentabodies remain functional after the passage through the GI tract and reach the cecum, which is the principal site of C. jejuni colonization [24], [66]. The demonstration that the flagellar pentabodies also inhibit motility could further contribute to their increased efficacy in reducing C. jejuni colonization in chickens.
While this is, to our knowledge, the first demonstration of the efficacy of VHH-based molecules in the reduction of C. jejuni levels in poultry, there are other reports of their successful application in reducing pathogen levels at mucosal surfaces. Harmsen et al. (2005) [67] observed that VHHs specific for the F4 fimbriae of enterotoxigenic E. coli (ETEC) were highly effective in reducing pathogen binding to piglet intestinal brush borders ex vivo and somewhat effective in reducing ETEC-induced fluid loss when perfused into a piglet jejunal segment. The research team later reported an improvement of the in vivo function of the VHHs by isolating protease-resistant versions of VHHs specific to F4 fimbriae of ETEC [68]. Preliminary studies have indicated that the pentabodies used in the current work are susceptible to degradation by trypsin, chymotrypsin and pepsin (data not shown) yet still demonstrated efficacy in in vivo studies. We expect a second-generation protease resistant version of the VHHs and their pentabodies used in this study will be even more effective in reducing the colonization of C. jejuni in the chicken cecum, and, therefore, oral application of VHHs and pentabodies might be considered as novel strategies for reducing C. jejuni contamination of poultry products. Future experiments will be designed to address the feasibility of obtaining protease-resistant pentabodies and evaluating their effectiveness in the prevention of C. jejuni colonization over the full growth cycle of the chicken, as well as in treatment of chickens that have already been colonized with the bacteria.
Materials and Methods
Ethics Statement
The animals were maintained and used in accordance with the recommendations of the Canadian Council on Animal Care Guide to the Care and Use of Experimental Animals. The experimental procedures were approved by the institutional animal care committee.
Antigen Preparation and Immunization
C. jejuni 81–176 flagella were isolated as described previously [69]. Briefly, bacteria were cultured 16 h in Mueller-Hinton broth (Sigma-Aldrich, St. Louis, MO, USA) under microaerobic conditions. Approximately 5×1012 cells were then harvested by centrifugation (11,000×g, 30 min) and resuspended in 100 ml of Tris-buffered saline solution. Flagella were sheared using a Waring blender on ice. Cell debris was pelleted by two low speed centrifugations (10,000×g, 20 min) and the supernatant was transferred to an ultracentrifugation tube. Flagella were pelleted by centrifugation for 1 h at 100,000×g. Further purification was done by resuspending the pellet in 1% (vol/vol) SDS in distilled water and re-pelleting the flagella by centrifugation. Pellets were finally resuspended in 200–500 µl of dH2O.
A male llama (Lama glama) was immunized subcutaneously with C. jejuni flagella. Seven injections were performed consisting of 100 µg of the antigen in a total volume of 0.5 ml mixed with an equal volume of either complete (day 1) or incomplete (days: 21, 35, 49, and 63) Freund's adjuvant (Sigma-Aldrich). The last two injections (days: 76 and 90) were performed without adjuvant. Pre-immune blood (15–20 ml) was collected before the first injection with subsequent collections on days 21, 56, 70 and 95. The specific immune response to flagella was analyzed by ELISA using total pre-immune and immune sera as described below. Serum from day 95 was fractionated as described before [54]. Protein G and A columns (GE Healthcare, Pittsburgh, PA, USA) were used for serum fractionation according to the manufacturer's instructions and separated fractions were adjusted to pH 6 with 1 M Tris/HCl, pH 8.8, and dialyzed against PBS at 4°C overnight. Individual heavy fractions (G1, A1 and A2) and G2 (conventional IgG) were analyzed for specific binding to flagella antigen by ELISA. Briefly, 96-well Maxisorp™ plates (Nalgene Nunc International, Rochester, NY) were coated overnight at 4°C with 5 µg/ml of flagella antigen in PBS. Wells were rinsed and blocked with 200 µl of 1% casein. Different dilutions of purified IgG fractions (G1, G2, A1 and A2) were added and incubated at room temperature for 1.5 h. Wells were washed with PBS with 0.05% Tween-20, and incubated with goat anti-llama IgG (H+L) (1∶1,000 in PBS) (Bethyl Laboratories, Montgomery, TX) followed by swine-anti-goat-HRP (1∶3,000 in PBS) (Cedarlane, Burlington, ON, Canada). Binding was detected by adding 100 µl Tetramethylbenzidine (TMB) peroxidase substrate per well (Kirkegaard and Perry Laboratories, Gaithersburg, MD, USA). Reactions were stopped by adding 100 µl 1 M phosphoric acid and A450 was measured using a Bio-Rad ELISA plate reader.
Phage Library Construction and Panning
A phage display library was constructed as previously described [70]. In brief, total RNA was isolated from approximately 2×107 lymphocytes collected on day 95 post-immunization start using a QIAamp RNA blood mini kit (Qiagen, Mississauga, Ontario, Canada). First-strand cDNA was synthesized with oligo(dT) primer using 5 µg total RNA as template according to the manufacturer's recommendations (GE Healthcare). Immunoglobulin variable domains and part of the constant region (VH/VHH-CH2) DNA were amplified by PCR using oligonucleotides MJ1-3 (sense) and two CH2 domain antisense primers CH2 and CH2b3 as previously described [70]. Briefly, the PCR reaction mixture was set up in a total volume of 50 µl with the following components: 1–3 µl cDNA, 10 pmol of MJ1-3 primer mixture, 10 pmol of either CH2 or CH2b3 primers, 5 µl of 10× reaction buffer, 1 µl of 10 mM dNTP, 2.5 units of Taq DNA polymerase (Hoffmann-La Roche Limited, Mississauga, ON, Canada). The PCR protocol consisted of an (i) initial step at 94°C for 3 min, followed by (ii) 30 cycles of 94°C for 1 min, 55°C for 30 s, 72°C for 30 s and (iii) a final extension step at 72°C for 7 min. The heavy chain fragments (550–650 bp in length) were gel-purified using a QIA quick gel extraction kit (Qiagen). The variable regions of the heavy chain antibodies (IgG2 and IgG3) were re-amplified in a second PCR reaction using MJ7and MJ8 oligonucleotides and the same conditions as described above [70]. The amplified PCR products were purified with a QIAquick PCR purification kit (Qiagene), digested with SfiI (New England Biolabs, Pickering, Ontario, Canada) and re-purified using the same kit. Twelve micrograms of digested VHH fragments were ligated with 40 µg (3∶1 molar ratio, respectively) SfiI-digested pMED1 phagemid vector [70] using LigaFast Rapid DNA ligation system (Promega, Madison, WI). Electrocompetent TG1 E. coli cells (Stratagene, La Jolla, CA) were transformed with using the ligated products as described previously and a library of approximately 5×107 transformants was obtained. The VHH fragments from 40 colonies were PCR-amplified and sequenced to analyze the complexity of the library. The library was expanded by culturing for 3–4 h in 2× YT (yeast extract tryptone) [71] containing ampicillin (100 µg/ml)/glucose (2% w/v) medium at 37°C. The bacterial cells were pelleted, resuspended in the same medium with 20% glycerol and stored at −80°C.
Panning was performed for a total of four rounds against the flagella as essentially described by Arbabi Ghahroudi et al. 2009 [70]. In summary, 1 ml of the library stock (5×1010 bacterial cells) was grown for 1–2 h at 37°C, with shaking at 250 rpm in 2× YT/Amp-Glucose (2% w/v) medium (A600 = 0.4–0.5) and infected with M13KO7 helper phage (20∶1 phage to cells ratio) (New England Biolabs) for 1 h at 37°C. After centrifugation of the culture at 4°C, the infected cell pellets were resuspended in 200 ml of 2× YT/Amp with 50 µg/ml kanamycin and incubated for 16 h at 37°C with shaking at 250 rpm. The phage particles in the culture supernatant were precipitated with polyethylene glycol and the phage pellets were resuspended in 2 ml of sterile PBS and the phage titre was determined as previously described [70]. For panning, 96-well Maxisorp™ plates were coated with flagella antigen and the wells were rinsed with PBS and blocked with PBS/1% (w/v) casein for 2 h at 37°C. Approximately 1012 rescued phage particles were added to the blocked wells and incubated for 2 h at 37°C. The wells were washed 5× with PBS with 0.1% Tween-20 and 5× with PBS. The bound phages were eluted with 0.1 M triethylamine for 10 min, neutralized with 1 M Tris-HCl, pH 7.4 and used to infect exponentially growing E. coli TG1 cells. After 30 min incubation at 37°C, the cells were superinfected with M13KO7 for an additional 15 min and grown in 2× YT containing ampicillin (100 µg/ml) and kanamycin (50 µg/ml) overnight at 37°C. Panning was continued for three more rounds following the same procedures except that the antigen concentration was reduced to 20, 15, and 10 µg/well and the washing cycle was increased to 7, 10 and 12× with PBS-T and PBS for the second, third and fourth rounds of panning, respectively. After four rounds of panning, 48 colonies were randomly picked and phage were prepared and tested for binding to flagella using a published phage ELISA protocol [70]. All positive clones were sequenced and unique sequences for each antigen were selected for sub-cloning and large-scale expression and purification. Two anti-flagella VHHs were isolated and further characterized. The sequencing data were deposited in GenBank™, accession numbers KF812523 (FlagV1M) and KF812524 (FlagV6M).
Expression and Purification of Soluble and Pentameric VHHs
VHH DNA was PCR amplified from the pMED1 phagemid vector using oligonucletides BbsI-VHH (5′-TATGAAGACACCAGGCCCAGGTAAAGCTGGAGGAGTCT- 3′) and BamHI-VHH (5′-TTGTTCGGATCCTGAGGAGACGGTGACCTG-3′) (for the monomer) or ApaI-VHH (5′-ATTATTATGGGCCCTGAGGAGACGGTGACCTGGGTC-3′) as the reverse primer with BbsI (for the pentamer). These PCR fragments were digested with BbsI-BamHI or BbsI-ApaI restriction enzyme pairs (New England Biolabs) and ligated separately with either a pUC derivative pSJF2H (monomer) or pVT2 (pentamer) expression vectors [70]. Upon ligation, all plasmids were used to transform electrocompetent TG1 E. coli followed by selection on LB ampicillin agar plates. Colonies were screened by PCR for inserts and the DNA was sequenced.
VHH antibodies were expressed in bacteria and purified as previously described [70]. In brief, protein expression was induced by addition of IPTG to the media, and the periplasmic contents were extracted from the cell pellet. Briefly, the cell pellets of monomeric VHH cultures were resuspended in 20 ml of ice cold TES (0.2 M Tris-HCl pH 8.0, 20% (w/v) sucrose, 0.5 mM EDTA) and incubated on ice for 30 min. Next, 30 ml of ice-cold 1/8 TES (diluted in dH2O) was added followed by incubation for 30 min on ice, and then centrifugation at 14,000×g for 30 min at 4°C. The resulting supernatant containing VHHs was dialysed overnight against the metal-affinity chromatography (IMAC) buffer A (10 mM HEPES pH 7.0, 500 mM NaCl) prior to loading on the HiTrap™ Chelating HP columns (GE Healthcare) for VHH purification as described previously [70]. Eluted protein fractions were analyzed by SDS-PAGE and Western blotting before being pooled and dialysed against PBS. Briefly, protein samples were run on a 12.5% acrylamide gel in duplicate and one stained with Coomassie blue dye. The protein on the second gel was transferred onto a nitrocellulose membrane using a Trans-BlotTM SD for semi-dry Western blotting (Bio-Rad Laboratories). The membrane was incubated with mouse anti-6× His monoclonal antibody followed by goat-anti-mouse alkaline phosphatase (AP) conjugate and developed by addition of AP substrate buffer as previously described [70]. For pentabody isolation, the cells were lysed using a lysozyme lysis method and the cell lysates were centrifuged and filtered through 0.22 µm membrane filters prior to antibody purification by IMAC as described previously [70].
ELISA
ELISA was performed to determine specific binding of the monomer and pentamer VHHs to the protein target as described above except that after washing the plate with PBST and blocking with PBS-casein (1%), a 5 µg/ml solution of either anti-flagella monomer (FlagV1M) or the corresponding pentamer (FlagV1P) was added to the respective wells and incubated for 1 h at 37°C. Wells were washed with PBST (0.05% v/v Tween-20) and rabbit anti-His6 antibody conjugated to HRP (1∶5000 in PBS) (Bethyl Laboratories) was added followed by incubation for 1 h at room temperature. Binding was detected with the TMB substrate (Kirkegaard and Perry Laboratories), the reaction was stopped with 1 M H3PO4, and A450 was measured using an ELISA plate reader as described above.
To determine the cross-reactivity of the purified anti-flagella pentamers (FlagV1P or FlagV6P) with different strains of C. jejuni, flagella were prepared from different strains and used for coating of the microtitre plates. ELISA was performed as described above. The Campylobacter strains were originally obtained from ATCC and kindly provided to us by Dr. Michel Gilbert, NRC, Canada: 81–176: ATCC BAA-2151, 11168: ATCC 700819, and Penner serotype strains [60]: P1: ATCC 43429, P2: ATCC 43430, P3: ATCC 43431, P4: ATCC 43432, P19: ATCC 43446, P36: ATCC 43456, and P64: originally from Erasmus University, also a gift from Dr. Michel Gilbert.
Surface Plasmon Resonance
For surface plasmon resonance, monomeric and pentameric VHHs were passed through size exclusion columns, Superdex 75 and 200 (GE Healthcare), respectively, in 10 mM HEPES, pH 7.4, containing 150 mM NaCl and 3 mM EDTA to remove aggregates. Monomeric and pentameric VHH peak fractions were collected and protein concentrations determined from A280 measurements. Purified flagella protein was biotinylated by reaction with Pierce EZ-Link Sulfo-NHS-LC-LC-biotin (GE Healthcare) followed by dialysis against PBS to remove excess unincorporated biotin. SPR analyses were performed using a Biacore 3000 instrument (GE Healthcare). All measurements were conducted at 25°C in 10 mM HEPES, pH 7.4, containing 150 mM NaCl, 3 mM EDTA and 0.005% surfactant P20 (GE Healthcare). Approximately 700–900 RUs of biotinylated flagella was captured on an SA sensor chip (GE Healthcare) at a flow rate of 5 µl/min. Various concentrations of the antibodies were injected over flagella-SA surface at a flow rate of 40 µl/min. Surfaces were regenerated by washing with running buffer and data were analyzed with BIAevaluation 4.1 software. Epitope mapping of flagella-specific antibodies was performed using Biacore co-injection experiments. Briefly, 60 µl of the anti-flagella VHHs diluted in HBS-EP buffer to a concentration of 50× their respective KD values were injected over 700 RUs of immobilized biotinylated flagella at 30 µl/min. A FlagV1 injection was followed by a FlagV6 injection.
Western Blotting
Whole cell lysates of overnight cultures of C. jejuni strains 81–176 and 11168, C. coli strain VC167 and the flagellar mutant strain 81–176 flaA−/flaB− were separated on 12.5% SDS polyacrylamide gels under reducing conditions and proteins transferred to nitrocellulose membranes. Membranes were blocked with 3% (w/v) bovine serum albumin (BSA) in phosphate buffered saline (PBS) and hybridized with FlagV1P and FlagV6P pentabodies for 1 h at room temperature. After washing five times with PBST, membranes were incubated with anti-His AP conjugates (diluted 1∶5,000 in blocking buffer) (Abcam, Cambridge, MA). Finally, the membranes were washed four times and incubated with AP substrate (Bio-Rad Laboratories, Mississauga, Canada) for 10 min. The AP reaction was stopped by rinsing the membranes with distilled H2O and air dried.
FITC Labelling of Pentabodies and Fluorescence Microscopy
FlagV1P and FlagV6P were labelled at a concentration of 2 mg/ml with FITC using the FITC-labeling kit from Invitrogen according the manufacturer's instructions. The labeled pentabodies were dialysed against PBS several times to remove unincorporated FITC. C. jejuni 81–176 wild type; 81–176 flaA−flaB− and C. jejuni 11168 were cultured in Mueller-Hinton broth (Sigma-Aldrich) until reaching exponential growth phase, pelleted and fixed in 3% formalin overnight. The cells were washed with PBS to remove formalin, and then 10 µl of cells at a final concentration of ∼1×108/ml were air dried on glass coverslips. Non-specific binding was blocked with 50 µl 5% Bacto skim milk (Difco)-PBS for 1 h at room temperature. The cells were incubated for 1 h at room temperature in 50 µl FITC labelled FlagV1P or FlagV6 pentabody diluted in PBS to 80 µg/ml. The cells were washed with PBS/0.1% Tween and then mounted onto glass slides with addition of Vectashield-DAPI (Vector Laboratories, Burlington, Canada) mounting medium. The slides were examined with a Zeiss Axiovert 200 M microscope (Zeiss, Toronto, Canada). The experiment was done in duplicate, on at least three independent occasions, with at least three fields of view on each coverslip imaged. To confirm the pattern of fluorescence observed were flagella filaments; a control of a rabbit polyclonal anti-81-176 flagellin was used at 1∶1,000 dilution in PBS, in place of the FlagV1 and FlagV6P pentabodies. The cells were washed with PBS/0.1% Tween, then incubated for 45 min at room temperature with 50 µl Alexafluor 488/FITC goat anti-rabbit IgG (Invitrogen) diluted 1∶1,000 in PBS. The slides were washed with PBS/0.1% Tween then mounted as stated above.
Motility Assay
A motility assay was performed as described previously [72]. Antibodies, at a final concentration of 1 µg/µl, were incubated with C. jejuni or C. coli (5×107 CFUs) at RT for 30 min. The mixtures were plated in the center of a petri dish containing Mueller-Hinton agar (0.4%) and incubated at 37°C under microaerobic conditions (5% O2, 10% CO2, and 85% N2). Bacterial motility was determined by measuring the diameter of the growth at 24 h, 48 h, and 72 h after inoculating the bacteria.
C. jejuni Colonization of Specific Pathogen-Free Leghorn Chicks
Cultures for chick colonization experiments were prepared by harvesting C. jejuni 81–176 grown for 18 h in Mueller Hinton broth (Sigma). Bacterial cells were diluted in PBS and maintained on ice until immediately before use. One day old SPF leghorns (mixed sex) were obtained from the hatchery at the Canadian Food Inspection Agency (CFIA, Ottawa, ON, Canada). The chicks were randomly assigned into negative control, positive control, and treatment groups, weighed, ID tagged, housed in animal containment units and provided with feed and water ad libitum. The units were housed in an environmentally controlled level 2 bio-containment room. On arrival, 10% of the birds were randomly tested for colonization with C. jejuni. On day two, positive control and treatment groups were orally challenged with 300 µl C. jejuni 81–176 (108 cfu/ml). Positive control groups received 300 µl PBS and treatment groups received 300 µl (1 mg) of the anti-flagella pentabody at 1 h, 24 h, and 48 h after the challenge. Birds were euthanized 48 h after pentabody treatments by cervical dislocation according to the approved guidelines of the Canadian Council for Animal Care. Ceca were aseptically collected for quantitative assessment of colonization. Cecal contents were serially plated onto Karmali agar (Oxoid) and C. jejuni counts were done after incubation for 2 days at 37°C under microaerobic conditions. The chicken body weights (in grams) were also measured on day 1 and 4 after challenging with C. jejuni alone or following pentabody administration, to determine the impact of the experimental treatments.
Localization of Pentabodies in the Chicken GI Tact by Sandwich ELISA
The presence and approximate concentration of the pentabodies in different regions of the chicken GI tract were evaluated by ELISA assay. Wells of Maxisorp™ ELISA plates were coated with 1 µg of mouse monoclonal anti-verotoxin antibody recognizing the verotoxin component of FlagV1P and FlagV6P, overnight at 4°C. After blocking with PBS-casein (1%), intestinal fluids collected from the cecum, ileum, jejunum and duodenum regions were added to the ELISA plate wells in 2-fold serial dilutions (1/2-1/2048) and incubated at 37°C for 1 h. Subsequently, the pentabodies were detected using rabbit anti-His6 IgG conjugated to HRP (1∶5,000 in PBS) (Bethyl Laboratories) and TMB substrate. The reactions were stopped and absorbance measurements were made at 450 nm.
Statistical Analyses
Data are presented as means ±SEM for each chicken group, unless otherwise specified. Differences in tissue bacterial burdens were assessed by the Student's t-test or one-way analysis of variance (ANOVA) followed by Bonferroni's post-hoc multiple comparison tests, when appropriate. Differences were considered significant when p<0.05.
Acknowledgments
The authors wish to thank Mary Foss and Annie Aubry for assistance with flagella preparations and motility assays, Nathalie Gaudette and Hong-Tong Sevinc for help with antibody library construction and antibody purification, and Shannon Ryan for technical assistance with the chicken colonization studies. We thank Patricia Guerry for providing the 81-176 flaA−flaB− strain and the anti-flagellin rabbit polyclonal antisera. This is National Research Council of Canada Publication # 50023.
Funding Statement
The authors are grateful to Dow AgroSciences and AbCelex Technologies for partially funding this project. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Man SM (2011) The clinical importance of emerging Campylobacter species. Nat Rev Gastroenterol Hepatol 8: 669–685. [DOI] [PubMed] [Google Scholar]
- 2. Stern NJ, Robach MC (2003) Enumeration of Campylobacter spp. in broiler feces and in corresponding processed carcasses. J Food Prot 66: 1557–1563. [DOI] [PubMed] [Google Scholar]
- 3. Newell DG, Fearnley C (2003) Sources of Campylobacter colonization in broiler chickens. Appl Environ Microbiol 69: 4343–4351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Blaser MJ (1997) Epidemiologic and clinical features of Campylobacter jejuni infections. J Infect Dis 176 Suppl 2 S103–S105. [DOI] [PubMed] [Google Scholar]
- 5. Humphrey T, O'Brien S, Madsen M (2007) Campylobacters as zoonotic pathogens: a food production perspective. Int J Food Microbiol 117: 237–257. [DOI] [PubMed] [Google Scholar]
- 6. Hariharan H, Murphy GA, Kempf I (2004) Campylobacter jejuni: Public health hazards and potential control methods in poultry: a review. Vet Med - Czech 49: 441–446. [Google Scholar]
- 7. Blaser MJ, Perez GP, Smith PF, Patton C, Tenover FC, et al. (1986) Extraintestinal Campylobacter jejuni and Campylobacter coli infections: host factors and strain characteristics. J Infect Dis 153: 552–559. [DOI] [PubMed] [Google Scholar]
- 8. Gellynck X, Messens W, Halet D, Grijspeerdt K, Hartnett E, et al. (2008) Economics of reducing Campylobacter at different levels within the Belgian poultry meat chain. J Food Prot 71: 479–485. [DOI] [PubMed] [Google Scholar]
- 9. Rautelin H, Koota K, von Essen R, Jahkola M, Siitonen A, et al. (1990) Waterborne Campylobacter jejuni epidemic in a Finnish hospital for rheumatic diseases. Scand J Infect Dis 22: 321–326. [DOI] [PubMed] [Google Scholar]
- 10. Hermans D, Pasmans F, Messens W, Martel A, Van Immerseel F, et al. (2012) Poultry as a host for the zoonotic pathogen Campylobacter jejuni. Vector Borne Zoonotic Dis 12: 89–98. [DOI] [PubMed] [Google Scholar]
- 11. Suzuki H, Yamamoto S (2009) Campylobacter contamination in retail poultry meats and by-products in the world: a literature survey. J Vet Med Sci 71: 255–261. [DOI] [PubMed] [Google Scholar]
- 12. European Food Safety Authority (EFSA) (2011) Scientific Opinion on Campylobacter in broiler meat production: controloptions and performance objectives and/or targets at different stages of the food chain. EFSA Journal 9: 1–141. [Google Scholar]
- 13. Nyachuba DG (2010) Foodborne illness: is it on the rise? Nutr Rev 68: 257–269. [DOI] [PubMed] [Google Scholar]
- 14. Lin J (2009) Novel approaches for Campylobacter control in poultry. Foodborne Pathog Dis 6: 755–765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Newell DG, Wagenaar JA (2000) Poultry infections and their control at the farm level. In: Campylobacter, Second edition Nachamkin I & Blaser MJ, ASM Press, Washington, DC,USA, 497–509.
- 16.Wagenaar JA, Jacobs-Reitsma W, Hofshagen M, Newell DG (2008) Poultry colonization with campylobacter and its control at the primary productionlevel. In: Nachamkin I, editor Szymanski CM, editor; Blaser MJ, Campylobacter. 3rd edition, Washington, DC, USA, 667–678.
- 17. Wagenaar JA, Mevius DJ, Havelaar AH (2006) Campylobacter in primary animal production and control strategies to reduce the burden of human campylobacteriosis. Rev Sci Tech 25: 581–594. [PubMed] [Google Scholar]
- 18. Mead GC (2002) Factors affecting intestinal colonization of poultry by campylobacter and role of microflora in control. World Poult Sci J 58: 169–178. [Google Scholar]
- 19. Sahin O, Luo N, Huang S, Zhang Q (2003) Effect of Campylobacter-specific maternal antibodies on Campylobacter jejuni colonization in young chickens. Appl Environ Microbiol 69: 5372–5379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Alfredson DA, Korolik V (2007) Antibiotic resistance and resistance mechanisms in Campylobacter jejuni and Campylobacter coli. FEMS Microbiol Lett 277: 123–132. [DOI] [PubMed] [Google Scholar]
- 21. Silva J, Leite D, Fernandes M, Mena C, Gibbs PA, et al. (2011) Campylobacter spp. as a Foodborne Pathogen: A Review. Front Microbiol 2: 200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Smith JL, Fratamico PM (2010) Fluoroquinolone resistance in campylobacter. J Food Prot 73: 1141–1152. [DOI] [PubMed] [Google Scholar]
- 23. el Shibiny A, Scott A, Timms A, Metawea Y, Connerton P, et al. (2009) Application of a group II Campylobacter bacteriophage to reduce strains of Campylobacter jejuni and Campylobacter coli colonizing broiler chickens. J Food Prot 72: 733–740. [DOI] [PubMed] [Google Scholar]
- 24. Loc CC, Atterbury RJ, el Shibiny A, Connerton PL, Dillon E, et al. (2005) Bacteriophage therapy to reduce Campylobacter jejuni colonization of broiler chickens. Appl Environ Microbiol 71: 6554–6563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Wagenaar JA, van Bergen MA, Mueller MA, Wassenaar TM, Carlton RM (2005) Phage therapy reduces Campylobacter jejuni colonization in broilers. Vet Microbiol 109: 275–283. [DOI] [PubMed] [Google Scholar]
- 26. Sklar IB Joerger RD (2001) Attempts to utilize bacteriophages to combat Salmonella enterica serovar Enteritidis in chickens. Journal of Food Safety 21: 15–30. [Google Scholar]
- 27. Smith HW, Huggins MB, Shaw KM (1987) The control of experimental Escherichia coli diarrhoea in calves by means of bacteriophages. J Gen Microbiol 133: 1111–1126. [DOI] [PubMed] [Google Scholar]
- 28. Svetoch EA, Stern NJ (2010) Bacteriocins to control Campylobacter spp. in poultry-A review. Poult Sci 89: 1763–1768. [DOI] [PubMed] [Google Scholar]
- 29. Santini C, Baffoni L, Gaggia F, Granata M, Gasbarri R, et al. (2010) Characterization of probiotic strains: an application as feed additives in poultry against Campylobacter jejuni. Int J Food Microbiol 141 Suppl 1 S98–108. [DOI] [PubMed] [Google Scholar]
- 30. Willis WL, Reid L (2008) Investigating the effects of dietary probiotic feeding regimens on broiler chicken production and Campylobacter jejuni presence. Poult Sci 87: 606–611. [DOI] [PubMed] [Google Scholar]
- 31. Castillo SL, Heredia N, Contreras JF, Garcia S (2011) Extracts of edible and medicinal plants in inhibition of growth, adherence, and cytotoxin production of Campylobacter jejuni and Campylobacter coli. J Food Sci 76: M421–M426. [DOI] [PubMed] [Google Scholar]
- 32. Kurekci C, Bishop-Hurley SL, Vercoe PE, Durmic Z, Al Jassim RA, et al. (2012) Screening of Australian plants for antimicrobial activity against Campylobacter jejuni. Phytother Res 26: 186–190. [DOI] [PubMed] [Google Scholar]
- 33. Messaoudi S, Kergourlay G, Rossero A, Ferchichi M, Prevost H, et al. (2011) Identification of lactobacilli residing in chicken ceca with antagonism against Campylobacter. Int Microbiol 14: 103–110. [DOI] [PubMed] [Google Scholar]
- 34. Solis de los SF, Donoghue AM, Venkitanarayanan K, Metcalf JH, Reyes-Herrera I, et al. (2009) The natural feed additive caprylic acid decreases Campylobacter jejuni colonization in market-aged broiler chickens. Poult Sci 88: 61–64. [DOI] [PubMed] [Google Scholar]
- 35. Hermans D, Martel A, Van Deun K, Verlinden M, Van Immerseel F, et al. (2010) Intestinal mucus protects Campylobacter jejuni in the ceca of colonized broiler chickens against the bactericidal effects of medium-chain fatty acids. Poult Sci 89: 1144–1155. [DOI] [PubMed] [Google Scholar]
- 36. Molatova Z, Skrivanova E, Macias B, McEwan NR, Brezina P, et al. (2010) Susceptibility of Campylobacter jejuni to organic acids and monoacylglycerols. Folia Microbiol (Praha) 55: 215–220. [DOI] [PubMed] [Google Scholar]
- 37. Clark JD, Oakes RD, Redhead K, Crouch CF, Francis MJ, et al. (2012) Eimeria species parasites as novel vaccine delivery vectors: anti-Campylobacter jejuni protective immunity induced by Eimeria tenella-delivered CjaA. Vaccine 30: 2683–2688. [DOI] [PubMed] [Google Scholar]
- 38. Layton SL, Morgan MJ, Cole K, Kwon YM, Donoghue DJ, et al. (2011) Evaluation of Salmonella-vectored Campylobacter peptide epitopes for reduction of Campylobacter jejuni in broiler chickens. Clin Vaccine Immunol 18: 449–454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Rice BE, Rollins DM, Mallinson ET, Carr L, Joseph SW (1997) Campylobacter jejuni in broiler chickens: colonization and humoral immunity following oral vaccination and experimental infection. Vaccine 15: 1922–1932. [DOI] [PubMed] [Google Scholar]
- 40. Widders PR, Perry R, Muir WI, Husband AJ, Long KA (1996) Immunisation of chickens to reduce intestinal colonisation with Campylobacter jejuni. Br Poult Sci 37: 765–778. [DOI] [PubMed] [Google Scholar]
- 41. Zeng X, Xu F, Lin J (2010) Development and Evaluation of CmeC Subunit Vaccine against Campylobacter jejuni. J Vaccines Vaccin 1: 112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Nurmi E, Rantala M (1973) New aspects of Salmonella infection in broiler production. Nature 241: 210–211. [DOI] [PubMed] [Google Scholar]
- 43. Zhang G, Ma L, Doyle MP (2007) Potential competitive exclusion bacteria from poultry inhibitory to Campylobacter jejuni and Salmonella. J Food Prot 70: 867–873. [DOI] [PubMed] [Google Scholar]
- 44. Boyd Y, Herbert EG, Marston KL, Jones MA, Barrow PA (2005) Host genes affect intestinal colonisation of newly hatched chickens by Campylobacter jejuni. Immunogenetics 57: 248–253. [DOI] [PubMed] [Google Scholar]
- 45. Behring EA (1890) Ueber das zustandekommen der diptherie-immunität und der tetanus-immunität bei thieren. Deutch Med Woch 49: 1113–1114. [PubMed] [Google Scholar]
- 46. Casadevall A, Dadachova E, Pirofski LA (2004) Passive antibody therapy for infectious diseases. Nat Rev Microbiol 2: 695–703. [DOI] [PubMed] [Google Scholar]
- 47. Burr DH, Caldwell MB, Bourgeois AL, Morgan HR, Wistar R Jr, et al. (1988) Mucosal and systemic immunity to Campylobacter jejuni in rabbits after gastric inoculation. Infect Immun 56: 99–105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Pavlovskis OR, Rollins DM, Haberberger RL Jr, Green AE, Habash L, et al. (1991) Significance of flagella in colonization resistance of rabbits immunized with Campylobacter spp. Infect Immun 59: 2259–2264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Rollwagen FM, Pacheco ND, Clements JD, Pavlovskis O, Rollins DM, et al. (1993) Killed Campylobacter elicits immune response and protection when administered with an oral adjuvant. Vaccine 11: 1316–1320. [DOI] [PubMed] [Google Scholar]
- 50. Dolby JM, Newell DG (1986) The protection of infant mice from colonization with Campylobacter jejuni by vaccination of the dams. J Hyg (Lond) 96: 143–151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Ueki Y, Umeda A, Fujimoto S, Mitsuyama M, Amako K (1987) Protection against Campylobacter jejuni infection in suckling mice by anti-flagellar antibody. Microbiol Immunol 31: 1161–1171. [DOI] [PubMed] [Google Scholar]
- 52. Stern NJ, Meinersmann RJ, Dickerson HW (1990) Influence of antibody treatment of Campylobacter jejuni on the dose required to colonize chicks. Avian Dis 34: 595–601. [PubMed] [Google Scholar]
- 53. Cawthraw SA, Lind L, Kaijser B, Newell DG (2000) Antibodies, directed towards Campylobacter jejuni antigens, in sera from poultry abattoir workers. Clin Exp Immunol 122: 55–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Hamers-Casterman C, Atarhouch T, Muyldermans S, Robinson G, Hamers C, et al. (1993) Naturally occurring antibodies devoid of light chains. Nature 363: 446–448. [DOI] [PubMed] [Google Scholar]
- 55. Dooley H, Flajnik MF, Porter AJ (2003) Selection and characterization of naturally occurring single-domain (IgNAR) antibody fragments from immunized sharks by phage display. Mol Immunol 40: 25–33. [DOI] [PubMed] [Google Scholar]
- 56. Greenberg AS, Avila D, Hughes M, Hughes A, McKinney EC, et al. (1995) A new antigen receptor gene family that undergoes rearrangement and extensive somatic diversification in sharks. Nature 374: 168–173. [DOI] [PubMed] [Google Scholar]
- 57. Arbabi GM, Desmyter A, Wyns L, Hamers R, Muyldermans S (1997) Selection and identification of single domain antibody fragments from camel heavy-chain antibodies. FEBS Lett 414: 521–526. [DOI] [PubMed] [Google Scholar]
- 58. Muyldermans S (2001) Single domain camel antibodies: current status. J Biotechnol 74: 277–302. [DOI] [PubMed] [Google Scholar]
- 59. Zhang J, Li Q, Nguyen TD, Tremblay TL, Stone E, et al. (2004) A pentavalent single-domain antibody approach to tumor antigen discovery and the development of novel proteomics reagents. J Mol Biol 341: 161–169. [DOI] [PubMed] [Google Scholar]
- 60. Penner JL, Hennessy JN (1980) Passive hemagglutination technique for serotyping Campylobacter fetus subsp. jejuni on the basis of soluble heat-stable antigens. J Clin Microbiol 12: 732–737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Holliger P, Hudson PJ (2005) Engineered antibody fragments and the rise of single domains. Nat Biotechnol 23: 1126–1136. [DOI] [PubMed] [Google Scholar]
- 62. Vanlandschoot P, Stortelers C, Beirnaert E, Ibanez LI, Schepens B, et al. (2011) Nanobodies(R): new ammunition to battle viruses. Antiviral Res 92: 389–407. [DOI] [PubMed] [Google Scholar]
- 63. Harmsen MM, Ruuls RC, Nijman IJ, Niewold TA, Frenken LG, et al. (2000) Llama heavy-chain V regions consist of at least four distinct subfamilies revealing novel sequence features. Mol Immunol 37: 579–590. [DOI] [PubMed] [Google Scholar]
- 64. Black RE, Levine MM, Clements ML, Hughes TP, Blaser MJ (1988) Experimental Campylobacter jejuni infection in humans. J Infect Dis 157: 472–479. [DOI] [PubMed] [Google Scholar]
- 65. Szymanski CM, King M, Haardt M, Armstrong GD (1995) Campylobacter jejuni motility and invasion of Caco-2 cells. Infect Immun 63: 4295–4300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Beery JT, Hugdahl MB, Doyle MP (1988) Colonization of gastrointestinal tracts of chicks by Campylobacter jejuni. Appl Environ Microbiol 54: 2365–2370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Harmsen MM, van Solt CB, Hoogendoorn A, van Zijderveld FG, Niewold TA, et al. (2005) Escherichia coli F4 fimbriae specific llama single-domain antibody fragments effectively inhibit bacterial adhesion in vitro but poorly protect against diarrhoea. Vet Microbiol 111: 89–98. [DOI] [PubMed] [Google Scholar]
- 68. Harmsen MM, van Solt CB, van Zijderveld-van Bemmel AM, Niewold TA, van Zijderveld FG (2006) Selection and optimization of proteolytically stable llama single-domain antibody fragments for oral immunotherapy. Appl Microbiol Biotechnol 72: 544–551. [DOI] [PubMed] [Google Scholar]
- 69. Power ME, Guerry P, McCubbin WD, Kay CM, Trust TJ (1994) Structural and antigenic characteristics of Campylobacter coli FlaA flagellin. J Bacteriol 176: 3303–3313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Arbabi-Ghahroudi M, Tanha J, Mackenzie R (2009) Isolation of monoclonal antibody fragments from phage display libraries. Methods Mol Biol 502: 341–364. [DOI] [PubMed] [Google Scholar]
- 71.Sambrook J, Russell D (2001) Molecular cloning: a laboratory manual, 3rd edn. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- 72. Guerry P, Ewing CP, Schirm M, Lorenzo M, Kelly J, et al. (2006) Changes in flagellin glycosylation affect Campylobacter autoagglutination and virulence. Mol Microbiol 60: 299–311. [DOI] [PMC free article] [PubMed] [Google Scholar]