

Abstract
Transient transformation is simpler, more efficient and economical in analyzing protein subcellular localization than stable transformation. Fluorescent fusion proteins were often used in transient transformation to follow the in vivo behavior of proteins. Onion epidermis, which has large, living and transparent cells in a monolayer, is suitable to visualize fluorescent fusion proteins. The often used transient transformation methods included particle bombardment, protoplast transfection and Agrobacterium-mediated transformation. Particle bombardment in onion epidermis was successfully established, however, it was expensive, biolistic equipment dependent and with low transformation efficiency. We developed a highly efficient in planta transient transformation method in onion epidermis by using a special agroinfiltration method, which could be fulfilled within 5 days from the pretreatment of onion bulb to the best time-point for analyzing gene expression. The transformation conditions were optimized to achieve 43.87% transformation efficiency in living onion epidermis. The developed method has advantages in cost, time-consuming, equipment dependency and transformation efficiency in contrast with those methods of particle bombardment in onion epidermal cells, protoplast transfection and Agrobacterium-mediated transient transformation in leaf epidermal cells of other plants. It will facilitate the analysis of protein subcellular localization on a large scale.
Introduction
Onion (Allium cepa L.), one kind of biennial herb Liliaceae plant, has been used as classical experimental materials in analyzing structure of plant cells, distribution location of DNA and RNA, reducing sugar of plant tissues [1], plasmolysis and recovery of plant cells [2], [3], karyotype [4], protein subcellular localization and interaction [5]–[7].
Imaging subcellular localization of proteins in living cells has become an important tool for defining protein function. Fluorescent fusion proteins are ideal marker non-enzymatic protein systems for imaging protein subcellular localization in living cells, which have many apparent advantages, such as stable fluorescence properties, easy observation, visualization in living cells, non-toxic to cells, non-specifity for species, without interference to false positives and no substrate etc. In addition, expression of fluorescent fusion proteins had also been used to investigate protein interaction, trafficking, turnover, movement and inheritance in living cells [8].
Transient transformation assays, which were conducted by using particle bombardment [9], [10], protoplast transfection [11], and Agrobacterium-infiltration [12], [13], [14], had been used to analyze gene function and protein-protein interactions in Arabidopsis [15], [16] and rice [11], [17], hybrid aspen [18], maize [19], [20], potato [21], [22], soybean [23]–[25], tomato [26]–[28], wheat [23], [29], [30], white spruce [31], celery and carrot [32].
However, most transient transformation methods have certain disadvantages, such as the lower transformation efficiency, equipment dependency and auxiliary material needed for particle bombardment, complex preparation procedures required for protoplasts transfection. In addition, for transient transformation in plants having complex outline of epidermal cells and the bushy epidermal hairs, laser scanning confocal microscope was usually needed to get ideal micro-images, which increased the reliability on expensive equipments. To avoid these disadvantages, we developed an in planta transient transformation method in living onion epidermal cells by using Agrobacterium-mediated infiltration. In this protocol, infiltration liquid of Agrobacterium carrying constructed vectors were injected into the interface between adaxial epidermis and mesophyll of onion bulb scales, which played an important role in yielding high transformation efficiency, and kept in the living onion bulb for about three days. With this simple method a higher frequency of transformation was achieved without expensive equipments in comparison with other transient transformation methods like protoplast transfection of Spinacia oleracea [33], A. thaliana [34] and Populus euphratica [35], agroinfiltration of Arabidopsis epidermal cells and particle bombardment of onion cells in vitro. In addition, following this method it took about 3 days for agroinfiltration to get ideal transient transformation efficiency in living onion epidermis. Therefore, the developed method was rapid, highly efficient, equipment independent and low-cost, which will benefit to analyze protein subcellular localization in a large-scale manner.
Results and Discussion
Special agroinfiltration method could benefit transient transformation
We first modified the agroinfiltration method previously used in Nicotiana tabacum [36], [37] in order to increase the efficiency of transient transformation. About 200 µl agroinfiltration liquid was slowly injected into the interfaces between adaxial epidermis and mesophyll of onion bulb scales by using a plastic syringe with needle, which resulted in an agroinfiltration bubble at the injection spot filled with infiltration liquid occupying about 1 cm2 area of epidermis (Figure 1). The formation of agroinfiltration bubble was demonstrated in the schematic diagram (Figure 2). We found that 1 cm2 agroinfiltration area with forming bubble gave higher transformation efficiency than a larger agroinfiltration area without forming bubble. The reason could be that forming agroinfiltration bubble filled with agroinfiltration liquid gave the infiltrated epidermal cells more chance to be infected by agrobacteria because the ratio of agrobacteria to infiltrated epidermal cells was higher. The special injection method could contribute to the high transformation efficiency of the developed transient transformation method. In contrast with other plant material, the adaxial epidermis and mesophyll of onion bulb scales are more flexible to be separated to form an interface bubble, which can take more agroinfiltration liquid.
Figure 1. Agrobacterium mediated in planta transient transoformationin living onion epidermal cells.

(A–F) Operational process of the modified agroinfiltration, (A) Onion bulb without outer scales, (B, C) The cut onion bulb prepared for subsequent injection, (D) The injection of Agrobacteria, (E) Bind injected cut scales together with elastic for further incubation, scale bar = 2.5 cm. (F) The magnification of injection location, scale bar = 4 mm. (G–V) Onion epidermal cells were transformed with constructs of pCM1205-RFP (G, H, I and J), pLPGM202 (K, L, M and N), pLPGM413 (O, P, Q and R) and pLPGM113 (S, T, U and V). Bright field images (G, K, O and S), UV excited fluorescence images (L, P, T and H), UV excited DAPI staining images (I, M, Q and U) and the merged images of fluorescence and DAPI (J, N, R and V), scale bar = 10 µm.
Figure 2. Schematic diagram of forming agroinfiltration bubble by injecting agroinfiltration liquid into the interface between adaxial epidermis and mesophyll of onion bulb scale.
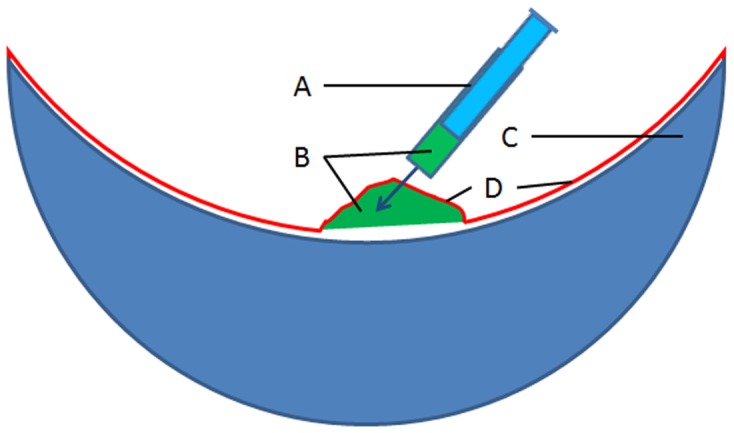
(A) Syringe. (B) Agroinfiltration liquid. (C) Mesophyll of onion bulb scale. (D) Adaxial epidermis of onion bulb scale.
Optimization of optical density (OD) of bacteria and agroinfiltration duration
The OD of bacteria and agroinfiltration duration were optimized. It showed that among the different agroinfiltration durations (24 h, 48 h, 72 h and 96 h), three-day agroinfiltration gave the significantly higher transformation efficiency (Table 1). While among three different concentrations of bacteria, agroinfiltration liquid with OD600 of 0.10 resulted in significantly higher transformation efficiency (Table 1). Considering both elements, the conditions of agroinfiltration liquid with OD600 of 0.10 and three-day infiltration were adopted to achieve the highest transformation efficiency of 43.87% (Table 1).
Table 1. Effects of bacterial concentrations and durations of agroinfiltration.
| Agroinfiltration durations | Transient transformation efficiencies (%) by using infiltration of Agrobacterium with different concentrations | ||
| 0.05 (OD600) | 0.10 (OD600) | 0.15 (OD600) | |
| 24 h | 3.30±0.259 cC | 12.03±0.282 dD | 5.10±0.216 dD |
| 48 h | 4.30±0.240 bB | 23.77±0.274 cC | 12.37±0.256 bB |
| 72 h | 13.10±0.326 aA | 43.87±0.431 aA | 22.53±0.335 aA |
| 96 h | 4.77±0.164 bB | 17.97±0.282 bB | 7.70±0.199 cC |
Note: three samples per combination of bacterial concentration and infiltration duration were investigated for ten 2 mm2 epidermal areas. Each efficiency value represents the mean of transformation efficiencies of thirty replicates of 2 mm2 epidermal areas from three samples, and the standard errors were calculated by using Excel. Different capital and lowercase letters within the same column exhibit significant difference at the 1% and 5% probability level according to the Duncan test of SPSS 10.0 statistic analysis.
Agroinfiltration liquid with special combination of components contributed to the high efficiency of transient transformation
The components of infiltration liquid and transfection conditions were referenced to relevant reports [13], [38]–[42] and with special modification. Based on agroinfiltration liquid for floral dip stable transformation and other transient transformation assays, the modified agroinfiltration liquid included 6-benzylaminopurine (BAP), Silwet L-77, D-glucose, acetosyringone (AS), magnesium chloride hexahydrate (MgCl2), calcium chloride dehydrate (CaCl2) and MES-KOH. To our knowledge, adding plant growth regulator BAP into the agroinfiltration liquid for transient transformation was seldom, although it had previously been applied in floral dip stable transformation. As an analogue of cytokinin, BAP plays roles in promoting cell division and prevent cell senescence, adding of low concentration of BAP in the agroinfiltration liquid could benefit to keep onion epidermal cells vigorous during agroinfiltration. Surfactant, glucose, osmotic buffer, calcium and magnesium ions, and acetosyringone were also supplemented to benefit the transient transformation.
In order to confirm the necessary roles of all the components, the transformation efficiencies of different infiltration liquids without one of the components were investigated (Table 2). The results indicated that all the components of agroinfiltration liquid were necessary and significantly contributed to the high efficiency of Agrobaterium-mediated transient transformation in onion epidermal cells.
Table 2. Effects of different components of agroinfiltration liquid (OD600 = 0.10).
| Infiltration components | Transient transformation efficiencies (%) of different agroinfiltration durations | |||
| 24 h | 48 h | 72 h | 96 h | |
| Complete components | 12.93±0.262 aA | 22.97±0.376 aA | 43.88±0.330 aA | 15.68±0.070 aA |
| D-glucose- | 0.15±0.014 fF | 0.91±0.013 gG | 2.61±0.040 gG | 2.40±0.031 hG |
| CaCl2- | 1.21±0.050 eE | 2.43±0.047 fF | 5.99±0.036 fF | 2.55±0.045 gG |
| MES-KOH- | 3.30±0.048 cC | 5.24±0.059 dD | 13.32±0.089 bB | 6.88±0.081 cC |
| BAP- | 2.13±0.062 dD | 4.21±0.049 eE | 8.28±0.043 dD | 5.67±0.043 dD |
| Silwet L-77- | 7.22±0.083 bB | 10.07±0.126 bB | 12.71±0.082 cC | 9.43±0.045 bB |
| MgCl2- | 2.42±0.065 dD | 6.34±0.040 cC | 8.39±0.040 dD | 4.32±0.029 eE |
| AS- | 1.15±0.041 eE | 4.39±0.044 eE | 7.29±0.253 eE | 3.42±0.054 fF |
Note: three samples per unit of infiltration components were investigated for ten 2 mm2 epidermal areas. Each efficiency value represents the mean of transformation efficiencies of thirty replicates of 2 mm2 epidermal areas from three samples (the percentage of positive cells in total cells per unit area), and the standard errors were calculated by using Excel. Different capital and lowercase letters within the same column exhibit significant difference at the 1% and 5% probability level according to the Duncan test of SPSS 10.0 statistic analysis. The symbol “-”, represents that the agroinfiltration liquid included all the components except the referred component.
The developed method is advantageous to particle bombardment and onion is more suitable for the developed method than tobacco and Arabidopsis
Through analyzing the side by side experiments of transforming pLPGM413 into onion epidermal cells with the developed method and particle bombardment, it indicated that the developed method resulted in significantly high transformation efficiency and showed advantages on cost and equipment needed (Table 3).
Table 3. Comparison of agroinfiltration and particle bombardment methods on transformation in onion epidermis.
| Comparison items | Transient transformation methods | |
| Agroinfiltration | Particle bombardment | |
| Transformation time | One day (about 12.93% transformation efficiency) | One day (about 4.67% transformation efficiency) |
| Cost | Low | High |
| Special equipment | Not needed | Biolistic equipment |
| Transformation efficiency (%) | 43.73±0.23 | 4.67±0.11 |
Note: three samples per unit of infiltration components were investigated for ten 2 mm2 epidermal areas. The efficiency value represents the mean of transformation efficiencies of thirty replicates of 2 mm2 epidermal areas from three samples (the percentage of positive cells in total cells per unit area) in the part of transformation efficiency, and the standard errors were calculated by using Excel.
The side by side experiments of Agrobacterium mediated transient transformation in onion, tobacco and Arabidopsis [14] with the same agroinfiltration liquid showed that the developed transient transformation method of onion epidermis gave significantly high transformation efficiency than tobacco and Arabidopsis (Table 4). The developed method of transformation in onion epidermis took only 5 days from the pretreatment of onion bulb to the best time-point for visualizing FFP signals, which is shorter than those in tobacco and Arabidopsis if considering the preparation time of plants [14]. In addition, the developed method is easier to get ideal images because onion epidermis has transparent cells arranged in a monolayer. Therefore, the developed method of onion has advantage to those of tobacco and Arabidopsis.
Table 4. Comparison of transient transformation efficiencies in different plant materials by using agroinfiltration.
| Plant materials | Transient transformation efficiencies (%) by using infiltration of Agrobacterium (OD600 = 0.10) with different agroinfiltration durations | |||
| 24 h | 48 h | 72 h | 96 h | |
| Arabidopsis(Col-0) | 1.31±0.041 cC | 2.56±0.066 cC | 1.79±0.043 cC | 1.63±0.053 cC |
| Tobacco(Xanthi) | 1.93±0.044 bB | 4.97±0.051 bB | 6.54±0.074 bB | 5.22±0.035 bB |
| Onion(Hongtaiyang) | 12.34±0.098 aA | 23.38±0.243 aA | 43.35±0.343 aA | 14.99±0.082 aA |
Note: three samples per material were investigated for ten 2 mm2 areas of agroinfiltrated epidermal cells. Each efficiency value represents the mean of transformation efficiencies of thirty replicates of 2 mm2 epidermal areas from three samples (the percentage of positive cells in total cells per unit area), and the standard errors were calculated by using Excel. Different capital and lowercase letters within the same column exhibit significant difference at the 1% and 5% probability level according to the Duncan test of SPSS 10.0 statistic analysis. For onion two-day pretreatment was conducted before agroinfiltration.
Pretreatment of onion bulb benefited the Agrobacterium mediated transient transformation in onion epidermis
The effects of different pretreatment times of onion bulb before injection of agroinfiltration liquid were evaluated (Table 5). It indicated that with the increase of pretreatment time the transformation efficiencies were increased (Table 5). However, 48-h and 72-h pretreatments resulted in similar transformation efficiencies, which are significantly higher than those resulted from 0-h and 24-h pretreatments (Table 5). It implied that 48-h pretreatment could be the suitable pretreatment time, which helped to achieve the highest transformation efficiency with the least time (Table 5).
Table 5. Effects of different pretreatment time of onion before Agrobacterium infection.
| Pretreatment time | Transient transformation efficiencies (%) by using infiltration of Agrobacterium concentration (OD600 = 0.10) with different agroinfiltration durations | |||
| 24 h | 48 h | 72 h | 96 h | |
| 0 h | 2.78±0.052 cC | 5.97±0.066 cC | 8.74±0.056 dC | 5.20±0.048 dD |
| 24 h | 8.10±0.068 bB | 15.01±0.158 bB | 24.10±0.279 cB | 13.17±0.272 cC |
| 48 h | 12.93±0.203 aA | 22.43±0.436 aA | 42.40±0.309 bA | 15.77±0.298 bB |
| 72 h | 13.03±0.286 aA | 22.10±0.480 aA | 43.57±0.513 aA | 17.71±0.276 aA |
Note: three repeat samples for each treatment were investigated by observing ten 2 mm2 epidermal areas of each sample. Each efficiency value represents the mean of transformation efficiencies of thirty replicates of 2 mm2 epidermal areas from three samples (the percentage of positive cells in total cells per unit area), and the standard errors were calculated by using Excel. Different capital and lowercase letters within the same column exhibit significant difference at the 1% and 5% probability level according to the Duncan test of SPSS 10.0 statistic analysis.
Monitoring of transformation and subcellular localization confirmation
The process of transient transformation was monitored by using four different plasmids carrying report genes mediated by A. tumefaciens strain GV3101, which constructed as the following expression binary vectors, pLPGM413 carrying RcSERK1 (somatic embryogenesis receptor-like kinase 1 of Rosa canina)[43] and GFP (green fluorescent proteins), pLPGM413 modified from pSAT6-GFP-N1 [44]–[47] by adding T-DNA border region of pCAMBIA1303, pLPGM113 modified from pEZS-NL-GFP [48] by adding T-DNA border region of pCAMBIA1303 and pCM1205-RFP (red fluorescent protein) carrying RFP [49], [50]. The Agrobacterium cells harboring above vectors were grown in liquid yeast extract and beef extract medium (YEB) until stationary phase and then re-suspended in infiltration liquid with above special components. Two to three days after injection of infiltration liquid, obvious GFP/RFP signals corresponding to different report genes could be observed in epidermal cells (Figure 1). It indicated that the developed agroinfiltration method can efficiently mediate in vivo transient expression of genes in living onion epidermal cells. For pCM1205-RFP, pLPGM202 and pLPGM113, the proteins encoded by their harboring genes were localized throughout the cells (Figure 1), and the fusion protein encoded by RcSERK1-GFP in pLPGM413 was localized in cell membrane (Figure 1). The results were the same as those of previous studies [43]–[50], suggesting the authenticity of the developed method.
The fluorescence signals of the nuclei in the cells transformed by pCM1205-RFP, pLPGM202 and pLPGM113 overlapped with those nuclei stained by 4′,6-diamidino-2-phenylindole (DAPI) (Figure 1), while no fluorescence signals were observed in nuclei of the cells transformed by pLPGM413. It further confirmed the credibility of the developed transient transformation method. It indicated that the developed transient transformation method in onion epidermal cells could be used to determine subcellular localization of the target proteins with the consistence to other transient transformation systems.
Conclusions
We have developed a rapid and low-cost transient transformation assay by using in-planta agroinfiltratoin in living onion epidermal cells, which transformation efficiency is higher than those of the conventional transient assays. It demonstrated that subcellular localization of proteins evaluated by using this method was same as that by other transient transformation assays and DAPI staining. Since it is rapid, efficient and low-cost, this method is suitable for large-scale analyses of protein subcelluar localization.
Materials and Methods
Plant material
Cultivated onion “Hongtaiyang” (A. cepa L.), which is a kind of Chinese onion cultivar with red skin, was used as experimental material. Before the injection of agroinfiltration liquid the onion bulbs pre-grew in darkness at 28°C for different days to evaluate the effect of different pretreatment time of onion bulb (Table 5). After injection the onion bulbs were incubated at 28°C in darkness for the agroinfiltration.
Tobacco (Nicotiana tabacum L. var. Xanthi nc.) was grown under 25°C with photoperiod of 16 h light (200 µmol m−2 s−1) and 8 h darkness. The leaves of fifty-day old plants were injected with agroinfiltration liquid from abaxial surface with syringe.
A. thaliana (Colombia ecotype) was grown under 20°C with photoperiod of 16 h light (120 µmol m−2s−1) and 8 h darkness. The leaves of 15 days old plants, the injection method was same as that of tobacco.
GFP/RFP-based constructs
The binary expression vector pLPGM202 harboring GFP genes was modified from pSAT6-GFP-N1 provided by Prof. Tao Wang (State Key Laboratory of Agro-biotechnology, China Agricultural University) by adding T-DNA border region of pCAMBIA1303, which contains the right border, kanamycin (Kan) resistance gene for bacterial resistance selection and the left border. The steps of creating pLPGM202 was as follows: the T-DNA border region (2888-9172) of pCAMBIA1303 was amplified by using PrimeSTAR® GXL DNA Polymerase (TaKaRa, Japan) with PCR primers of Fsat-YR (NotI-GTAAACCTAAGAGAAAAGAG) and Rsat-YR (PI-PspI-TTTGCCTGTTTACACCACAAT) carrying the restriction enzyme cutting sites of NotI and PI-PspI, respectively. The PCR products were ligated into pSIMPLE-19 EcoRV/BAP vector by T4 DNA Ligase (TaKaRa, Japan) and sequenced to confirm the success of ligation. The constructed recombinant pSIMPLE-19 EcoRV/BAP with the T-DNA border region of pCAMBIA1303 was transformed into Escherichia coli DH5α for propagation. The pSAT6-GFP-N1 were digested with partial digestion method by using NotI and PI-PspI (NEB, USA), the target digested pSAT6-GFP-N1 was obtained with electrophoresis method. The T-DNA border region of pCAMBIA1303 was obtained from the recombinant pSIMPLE-19 EcoRV/BAP through complete digestion with NotI and PI-PspI, and cloned into the target partially-digested pSAT6-GFP-N1 to form pLPGM202 (Figure S1). The successful construction of pLPGM202 was confirmed by sequencing (Sangon Biotech, China).
The binary expression vector pLPGM113 was modified from pEZS-NL-GFP provided by Dr. Liang Zhang (Department of Life Science, Henan Normal University) by adding T-DNA border region (2888-9172) of pCAMBIA1303 same as that for pLPGM202 described above. The steps of creating pLPGM113 were similar as those of pLPGM202 to be simply described as follows: The T-DNA border region of pCAMBIA1303 was amplified with the primers Fnl-YR (PstI-GTAAACCTAAGAGAAAAGAG) and Rnl-YR (SpeI-TGTTTACACCACAATATATCC) carrying the restriction enzyme cutting sites of PstI and SpeI, respectively. The PCR products were ligated into pSIMPLE-19 EcoRV/BAP vector by T4 DNA Ligase (TaKaRa, Japan) and propagated in E. coli DH5α. The pEZS-NL-GFP were digested with partial digestion method by using PstI and SpeI (NEB, USA), the target digested pEZS-NL-GFP was obtained with electrophoresis method. The T-DNA border region of pCAMBIA1303 was obtained from the recombinant pSIMPLE-19 EcoRV/BAP through complete digestion with PstI and SpeI, and cloned into the target partially-digested pEZS-NL-GFP to form pLPGM113 (Figure S2). The successful construction of pLPGM113 was confirmed by sequencing (Sangon Biotech, China).
The binary expression vector pCM1205-RFP harboring RFP gene was provided by Dr. Wencai Qi (Bioengineering Department, Zhengzhou University).
The cDNA of RcSERK1 was isolated from cDNA library of R. canian PLBs and its sequence was deposited in GenBank as accession number of HM802242. Coding sequence of this gene (without stop codon) was amplified by PCR using Platinum pfx DNA polymerase (Invitrogen, USA) with the following primers: RcSERK1 (F-SKs, forward: 5′-CCCAAGCCTCATGGATAGCAGGCTT-3′; R-SKs, reverse: 5′-TCCCCCGGGCCTTGGACCAGATAAC-3′). The PCR products were digested with HindIII and SmaI, subsequently ligated into the HindIII/SmaI sites between the CaMV 35S promoter and GFP of pLPGM202 to construct the expression binary vector pLPGM413.
Agrobacterium infiltration
The vectors, pCM1205-RFP, pLPGM113, pLPGM202 and pLPGM413, were transformed into A. tumefaciens strain GV3101 for further transient transformation in onion epidermis. Positive Agrobacterium harboring pCM1205-RFP was selected and cultivated in YEB media supplemented with 100 mg/L rifampicin and 25 mg/L chloramphenicol. Positive Agrobacterium harboring pLPGM113, pLPGM202 and pLPGM413 were selected and cultivated in YEB media supplemented with 100 mg/L rifampicin and 100 mg/L Kan. Positive Agrobacterium cultivated overnight at 28°C were harvested at OD600 of 1.5 to 2.0, centrifuged at 5000 rpm for 10 min and re-suspended in 50 ml of infiltration liquid, and the centrifugation and resuspension procedure was repeated three to five times. Finally, Agrobacterium cells were diluted in agroinfiltration liquid to appropriate concentration for agroinfiltration. Different agroinfiltration durations (24 h, 48 h, 72 h and 96 h) and Agrobacterium concentrations (OD600 0.05, 0.10 and 0.15) were evaluated to determine conditions to obtain high transformation efficiency (Table 1).
The complete infiltration liquid was made as following: 41.65 mM D-glucose, 100 mM CaCl2, 100 mM MES-KOH (pH 5.6) stock solution, 0.011 µM BAP, 0.01% Silwet L-77, 0.05 mM MgCl2 and 12.5 mM AS (made with DMF, dimethylformamide) stock solution, and suitable amount of ddH2O to make final volume to 20 ml. About 200 µl infiltration liquid with Agrobacterium carrying constructed vectors were injected into the interface of adaxial epidermis and mesophyll of onion scales to make a bubble for agroinfiltration. In order to investigate whether all the components are necessary, different infiltration liquid without one of the components were used to evaluated the effect of different components of the infiltration liquid (Table 2).
Particle bombardment
The adaxial epidermis was obtained from onion bulb and placed on MS medium for 1-day incubation. The binary expression vector pLPGM413 was transformed into the onion epidermal cells with particle bombardment method as described by [51].
DAPI staining
To visualize nuclei, the epidermis was stained with DAPI (5 µg/mL, sigma, USA). Materials were soaked in the dye liquid phosphate buffer solution (PBS) (pH 7.0; DAPI: PBS (v/v) = 1∶1000) and kept in darkness for 20 min. Pieces of onion epidermis were arranged on slides to make wet mounts, the made slides were observed and photographed in dark-field of fluorescence microscope (Olympus BX 61, Japan).
Microscopic investigation
Transformation efficiency was determined by calculating the proportion of positive epidermal cells with fluorescent signals among the cells in 2 mm2 epidermis area that was measured with micro ruler under microscope. Images of epidermal cells were taken by using a motorized fluorescence microscope with a mirror unit (U-MNU2), dichroic mirror (DM400), excitation filter (BP360) and barrier filter (BA420) for DAPI (DNA staining). Images of epidermal cells positive for GFP were taken with a mirror unit (U-MSWB2), dichroic mirror (DM500), excitation filter (BP470-490) and barrier filter (BA520IP) for GFP. Images of epidermal cells positive for RFP were taken with a mirror unit (U-MSWG2), dichroic mirror (DM570), excitation filter (BP530-550) and barrier filter (BA590).
Statistical analysis
All statistical analyses were performed with SPSS (version 10.0).
Supporting Information
The sketch map of pLPGM202 originated from pCAMBIA1303 and pSAT6-GFP-N1. Details of creating the vector are given in the Materials and methods.
(TIF)
The sketch map of pLPGM113 originated from pCAMBIA1303 and pEZS-NL-GFP. Details of creating the vector are given in the Materials and methods.
(TIF)
Acknowledgments
The vectors, pSAT6-GFP-N1, pEZS-NL-GFP and pCM1205-RFP, were provided by Prof. Tao Wang (State Key Laboratory of Agro-biotechnology, China Agricultural University), Dr. Liang Zhang (Department of Life Science, Henan Normal University) and Dr. Wen-Cai Qi (Bioengineering Department, Zhengzhou University), respectively.
Funding Statement
The research was supported by National Natural Science Foundation of China (31071807, 31272168) (http://www.nsfc.gov.cn), Plan for Scientific Innovation Talent of Henan Province (124100510021) (http://www.hnkjt.gov.cn), Doctoral Scientific Research Starting Foundation of Zhoukou Normal University (zksybscx201108)(http://www.zknu.edu.cn), Scientific Research and Innovation Fund Projects of Zhoukou Normal University (zksykycx201306) (http://www.zknu.edu.cn) and the Science and Technology Research Major Projects of Department of Education of Henan Province (13B210270)(http://www.haedu.gov.cn). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Mitra J, Shrivastava SL, Rao PS (2012) Onion dehydration: a review. Int J Food Sci Tech 49: 267–277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Oparka KJ (1994) Plasmolysis: new insights into an old process. New Phytol 126(4): 571–591. [Google Scholar]
- 3. McLusky SR, Bennett MH, Beale MH, Lewis MJ, Gaskin P, et al. (1999) Cell wall alterations and localized accumulation of feruloyl-3′-methoxytyramine in onion epidermis at sites of attempted penetration by Botrytis allii are associated with actin polarisation, peroxidase activity and suppression of flavonoid biosynthesis. Plant J 17: 523–534. [Google Scholar]
- 4. Keller ERJ, Schubert I, Fuchs J, Meister A (1996) Interspecific crosses of onion with distant Allium species and characterization of the presumed hybrids by means of flow cytometry, karyotype analysis and genomic in situ hybridization. Theor Appl Genet 92: 417–424. [DOI] [PubMed] [Google Scholar]
- 5. Lee LY, Fang MJ, Kuang LY, Gelvin SB (2008) Vectors for multi-color bimolecular fluorescence complementation to investigate protein-protein interactions in living plant cells. Plant Methods 4: 24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Hollender CA, Liu Z (2010) Bimolecular fluorescence complementation (BiFC) assay for protein-protein interaction in onion cells using the helios gene gun. J Vis Exp 40: 1963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Eady CC, Lister CE, Suo Y, Schaper D (1996) Transient expression of uidA constructs in in vitro onion (Allium cepa L.) cultures following particle bombardment and Agrobacterium-mediated DNA delivery. Plant Cell Rep 15(12): 958–962. [DOI] [PubMed] [Google Scholar]
- 8. Hu CD, Kerppola TK (2003) Simultaneous visualization of multiple protein interactions in living cells using multicolor fluorescence complementation analysis. Nat Biotechnol 21: 539–545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Ueki S, Lacroix B, Krichevsky A, Lazarowitz SG, Citovsky V (2009) Functional transient genetic transformation of Arabidopsis leaves by biolistic bombardment. Nat Protoc 4: 71–77. [DOI] [PubMed] [Google Scholar]
- 10. Zhang G, Lu S, Chen TA, Funk CR, Meyer WA (2003) Transformation of triploid bermudagrass (Cynodon dactylon X C. transvaalensis cv. TifEagle) by means of biolistic bombardment. Plant Cell Rep 21: 860–864. [DOI] [PubMed] [Google Scholar]
- 11. Chen S, Tao L, Zeng L, Vega-Sanchez ME, Umemura K, et al. (2006) A highly efficient transient protoplast system for analyzing defence gene expression and protein-protein interactions in rice. Mol Plant Pathol 7: 417–427. [DOI] [PubMed] [Google Scholar]
- 12. Li JF, Park E, von Arnim AG, Nebenführ A (2009) The FAST technique: a simplified Agrobacterium-based transformation method for transient gene expression analysis in seedlings of Arabidopsis and other plant species. Plant Methods 5: 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Marion J, Bach L, Bellec Y, Meyer C, Gissot L, et al. (2008) Systematic analysis of protein subcellular localization and interaction using high-throughput transient transformation of Arabidopsis seedlings. Plant J 56: 169–179. [DOI] [PubMed] [Google Scholar]
- 14. Ye GN, Stone D, Pang SZ, Creely W, Gonzalez K, et al. (1999) Arabidopsis ovule is the target for Agrobacterium in planta vacuum infiltration transformation. Plant J 19: 249–257. [DOI] [PubMed] [Google Scholar]
- 15. Abel S, Theologis A (1994) Transient transformation of Arabidopsis leaf protoplasts: a versatile experimental system to study gene expression. Plant J 5: 421–427. [DOI] [PubMed] [Google Scholar]
- 16. Blachutzik JO, Demir F, Kreuzer I, Hedrich R, Harms GS (2012) Methods of staining and visualization of sphingolipid enriched and non-enriched plasma membrane regions of Arabidopsis thaliana with fluorescent dyes and lipid analogues. Plant Methods 8: 28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Zhang Y, Su J, Duan S, Ao Y, Dai J, et al. (2011) A highly efficient rice green tissue protoplast system for transient gene expression and studying light/chloroplast-related processes. Plant Methods 7: 30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Takata N, Eriksson ME (2012) A simple and efficient transient transformation for hybrid aspen (Populus tremula × P. tremuloides). Plant Methods 8: 30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Reggiardo MI, Arana JL, Orsaria LM, Permingeat HR, Spitteler MA, et al. (1991) Transient transformation of maize tissues by microparticle bombardment. Plant Sci 75: 237–243. [Google Scholar]
- 20. Hamilton DA, Roy M, Rueda J, Sindhu RK, Sanford J, et al. (1992) Dissection of a pollen-specific promoter from maize by transient transformation assays. Plant Mol Biol 18: 211–218. [DOI] [PubMed] [Google Scholar]
- 21. Sidorov VA, Kasten D, Pang SZ, Hajdukiewicz PT, Staub JM, et al. (1999) Stable chloroplast transformation in potato: use of green fluorescent protein as a plastid marker. Plant J 19: 209–216. [DOI] [PubMed] [Google Scholar]
- 22. Bhaskar PB, Venkateshwaran M, Wu L, Ané JM, Jiang J (2009) Agrobacterium-mediated transient gene expression and silencing: a rapid tool for functional gene assay in potato. PLoS One 4: e5812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Wang YC, Klein TM, Fromm M, Cao J, Sanford JC, et al. (1988) Transient expression of foreign genes in rice, wheat and soybean cells following particle bombardment. Plant Mol Biol 11: 433–439. [DOI] [PubMed] [Google Scholar]
- 24. Santarem ER, Trick HN, Essig JS, Finer JJ (1998) Sonication-assisted Agrobacterium-mediated transformation of soybean immature cotyledons: optimization of transient expression. Plant Cell Rep 17: 752–759. [DOI] [PubMed] [Google Scholar]
- 25. Trick HN, Finer JJ (1998) Sonication-assisted Agrobacterium-mediated transformation of soybean [Glycine max (L.) Merrill] embryogenic suspension culture tissue. Plant Cell Rep 17: 482–488. [DOI] [PubMed] [Google Scholar]
- 26. Baum K, Gröning B, Meier I (1997) Improved ballistic transient transformation conditions for tomato fruit allow identification of organ-specific contributions of I-box and G-box to the RBCS2 promoter activity. Plant J 12: 463–469. [DOI] [PubMed] [Google Scholar]
- 27. Wroblewski T, Tomczak A, Michelmore R (2005) Optimization of Agrobacterium-mediated transient assays of gene expression in lettuce, tomato and Arabidopsis. Plant Biotechnol J 3: 259–273. [DOI] [PubMed] [Google Scholar]
- 28. Orzaez D, Mirabel S, Wieland WH, Granell A (2006) Agroinjection of tomato fruits. A tool for rapid functional analysis of transgenes directly in fruit. Plant Physiol140: 3–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Rasco-Gaunt S, Riley A, Barcelo P, Lazzeri PA (1999) Analysis of particle bombardment parameters to optimise DNA delivery into wheat tissues. Plant Cell Rep 19: 118–7. [DOI] [PubMed] [Google Scholar]
- 30. Amoah BK, Wu H, Sparks C, Jones HD (2001) Factors influencing Agrobacterium-mediated transient expression of uidA in wheat inflorescence tissue. J Exp Bot 52: 1135–1142. [DOI] [PubMed] [Google Scholar]
- 31. Li YH, Tremblay FM, Séguin A (1994) Transient transformation of pollen and embryogenic tissues of white spruce (Picea glauca (Moench.) Voss) resulting from microprojectile bombardment. Plant Cell Rep 13: 661–665. [DOI] [PubMed] [Google Scholar]
- 32. Liu CN, Li XQ, Gelvin SB (1992) Multiple copies of virG enhance the transient transformation of celery, carrot and rice tissues by Agrobacterium tumefaciens . Plant Mol Biol 20: 1071–1087. [DOI] [PubMed] [Google Scholar]
- 33. Ohlrogge JB, Kuhn DN, Stumpf PK (1979) Subcellular localization of acyl carrier protein in leaf protoplasts of Spinacia oleracea . P Natl Acad Sci 76: 1194–1198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Pommerrenig B, Popko J, Heilmann M, Schulmeister S, Dietel K, et al. (2013) SUCROSE TRANSPORTER 5 supplies Arabidopsis embryos with biotin and affects triacylglycerol accumulation. Plant J 73: 392–404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Zhang H, Lv F, Han X, Xia X, Yin W (2013) The calcium sensor PeCBL1, interacting with PeCIPK24/25 and PeCIPK26, regulates Na+/K+ homeostasis in Populus euphratica . Plant Cell Rep 32: 611–621. [DOI] [PubMed] [Google Scholar]
- 36. Yang KY, Liu Y, Zhang S (2001) Activation of a mitogen-activated protein kinase pathway is involved in disease resistance in tobacco. P Natl Acad Sci 98: 741–746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Noël LD, Cagna G, Stuttmann J, Wirthmüller L, Betsuyaku S, et al. (2007) Interaction between SGT1 and cytosolic/nuclear HSC70 chaperones regulates Arabidopsis immune responses. Plant Cell 19: 4061–4076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Chen X, Equi R, Baxter H, Berk K, Han J, et al. (2010) A high-throughput transient gene expression system for switchgrass (Panicum virgatum L.) seedlings. Biotechnol Biofuels 3: 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Tsuda K, Qi Y, Nguyen LV, Bethke G, Tsuda Y, et al. (2012) An efficient Agrobacterium-mediated transient transformation of Arabidopsis. Plant J 69: 713–719. [DOI] [PubMed] [Google Scholar]
- 41. Hosein FN, Lennon AM, Umaharan P (2012) Optimization of an Agrobacterium-mediated transient assay for gene expression studies in Anthurium andraeanum . J Am Soc Hortic Sci 137: 263–272. [Google Scholar]
- 42. Shah K, Gadella JTW, van EH, Hecht V, de Vries SC (2001) Subcellular localization and oligomerization of the Arabidopsis thaliana somatic embryogenesis receptor kinase 1 protein. J Mol Biol 309: 641–655. [DOI] [PubMed] [Google Scholar]
- 43. Xu KD, Liu QL, Yang HF, Zeng L, Dong LL, et al. (2011) Isolation and molecular characterization of RcSERK1: A Rosa canina gene transcriptionally induced during initiation of protocorm-like bodies. African J of Biotech 10: 4011–4017. [Google Scholar]
- 44. Liu QL, Xu KD, Ma N, Zeng L, Zhao LJ (2010) Isolation and functional characterization of DgZFP: a gene encoding a Cys2/His2-type zinc finger protein in chrysanthemum. Mol Biol Rep 37: 1137–1142. [DOI] [PubMed] [Google Scholar]
- 45. Liu QL, Xu KD, Zhao LJ, Pan YZ, Jiang BB, et al. (2011) Overexpression of a novel chrysanthemum NAC transcription factor gene enhances salt tolerance in tobacco. Biotechnol Lett 33: 2073–2082. [DOI] [PubMed] [Google Scholar]
- 46. Han DG, Yang GH, Xu KD, Shao Q, Yu ZY, et al. (2013) Overexpression of a Malus xiaojinensis Nas1 gene influences flower development and tolerance to iron stress in transgenic tobacco. Plant Mol Biol Rep 31: 802–809. [Google Scholar]
- 47. Blanvillain R, Kim JH, Wu S, Lima A, Ow DW (2009) OXIDATIVE STRESS 3 is a chromatin-associated factor involved in tolerance to heavy metals and oxidative stress. Plant J 57: 654–665. [DOI] [PubMed] [Google Scholar]
- 48. Song S, Chen Y, Zhao M, Zhang WH (2012) A novel Medicago truncatula HD-Zip gene, MtHB2, is involved in abiotic stress responses. Environ Exp Bot 80: 1–9. [Google Scholar]
- 49. Zhou C, Wang H, Zhu J, Liu Z (2013) Molecular cloning, subcellular localization and functional analysis of ThCLC-a from Thellungiella halophila . Plant Mol Biol Rep 31: 783–790. [Google Scholar]
- 50. Tsai AYL, Gazzarrini S (2012) AKIN10 and FUSCA3 interact to control lateral organ development and phase transitions in Arabidopsis. Plant J 69: 809–821. [DOI] [PubMed] [Google Scholar]
- 51.Wang GL, Fang HY (2002). Gene engineering in plant (the 2nd edition). Beijing: China Science Press, Beijing, pp. 734–736. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The sketch map of pLPGM202 originated from pCAMBIA1303 and pSAT6-GFP-N1. Details of creating the vector are given in the Materials and methods.
(TIF)
The sketch map of pLPGM113 originated from pCAMBIA1303 and pEZS-NL-GFP. Details of creating the vector are given in the Materials and methods.
(TIF)